Abstract
Obesity and obesity-associated disorders pose a major public health issue worldwide. Apart from conventional weight loss drugs, next-generation probiotics (NGPs) seem to be very promising as potential preventive and therapeutic agents against obesity. Candidate NGPs such as Akkermansia muciniphila, Faecalibacterium prausnitzii, Anaerobutyricum hallii, Bacteroides uniformis, Bacteroides coprocola, Parabacteroides distasonis, Parabacteroides goldsteinii, Hafnia alvei, Odoribacter laneus and Christensenella minuta have shown promise in preclinical models of obesity and obesity-associated disorders. Proposed mechanisms include the modulation of gut flora and amelioration of intestinal dysbiosis, improvement of intestinal barrier function, reduction in chronic low-grade inflammation and modulation of gut peptide secretion. Akkermansia muciniphila and Hafnia alvei have already been administered in overweight/obese patients with encouraging results. However, safety issues and strict regulations should be constantly implemented and updated. In this review, we aim to explore (1) current knowledge regarding NGPs; (2) their utility in obesity and obesity-associated disorders; (3) their safety profile; and (4) their therapeutic potential in individuals with overweight/obesity. More large-scale, multicentric and longitudinal studies are mandatory to explore their preventive and therapeutic potential against obesity and its related disorders.
1. Introduction
Obesity has become a major global public threat that is associated with a variety of disorders, including metabolic syndrome, type 2 diabetes mellitus, cardiovascular disease, nonalcoholic fatty liver disease, polycystic ovary syndrome, cancer, autoimmune disorders, and the severity of COVID-19 [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15]. Based on recent WHO data, the worldwide prevalence of overweight and obesity in the adult population is greater than 39% and 13%, respectively [16].
As the obesity epidemic is on the rise worldwide, novel treatment strategies are being developed [12,17]. Apart from classical treatment options, such as glucagon-like peptide-1 (GLP-1) receptor agonists [18,19,20], researchers have also focused on the microbiome as a potential target to combat obesity [12,20,21,22,23].
The term microbiome refers to the sum of genes from the total amount of bacteria, viruses, fungi, archaea and protozoa inhabiting the human body. Amongst them, most of the microorganisms in the human body reside in the gut, comprising the human gut microbiota [21,22,23,24]. There, trillions of microorganisms interact with each other and produce a variety of metabolites, which play a crucial physiological role [25,26,27]. More specifically, microorganisms in the gut produce short-chain fatty acids (SCFAs), such as butyric acid, propionic acid and acetate, which account for several beneficial functions. SCFAs have been implicated in strengthening the integrity of the gut barrier as well as in reducing serum endotoxin levels and serum inflammatory markers [27,28,29]. Nowadays, the notion that the exogenous administration of these beneficial microbes can improve human health is widely accepted. In 1974, the term “probiotics” was introduced to describe “live microorganisms that confer a health benefit when consumed in adequate amounts”. This was the final definition proposed by the World Health Organization (WHO) in 2002 [30]. The most famous probiotics are Lactobacillus strains and Bifidobacteria, which have been used in dietary supplements and functional foods [31]. Saccharomyces boulardii is another well-known probiotic that has been administered for the prophylaxis of antibiotic-associated diarrhea [32]. Lactobacillus and Bifidobacterium strains have been reported to ameliorate obesity and obesity-associated disorders in rodents [33,34,35,36,37,38,39,40,41,42,43,44]. In addition, Saccharomyces boulardii has also been demonstrated to reduce fat mass and systemic inflammation in rodents with obesity and diabetes [45]. However, with the recent advent of multi-omics and other technological developments, new species with beneficial health effects have been identified, called next-generation probiotics (NGPs). Currently, there has been a tendency to identify and test these NGPs in the context of specific diseases. NGPs are currently in the spotlight of research, and there is a growing body of evidence with regard to their therapeutic potential in obesity and its related disorders. In this review, we aim to explore (1) current knowledge regarding NGPs; (2) their utility in obesity and obesity-associated disorders; (3) their safety profile; and (4) their therapeutic potential in individuals with overweight/obesity.
2. A Brief Synopsis of Classical Probiotics and Their Effects on Obesity and Associated Disorders
The available preclinical and clinical evidence points toward the presence of the beneficial effects of certain classical probiotics on obesity and related disorders. Reported results have been variable and partially strain-specific, with heterogeneity amongst studies being an additional factor contributing to equivocal results. The proposed mechanisms of action for classical probiotics in obesity include the modulation of gut flora and amelioration of intestinal dysbiosis, improved intestinal barrier function with a reduction in endotoxemia, low-grade inflammation and oxidative stress, which, in turn, beneficially affect aspects of energy metabolism in adipose and other target tissues [46,47,48,49].
The administration of different strains of Lactobacillus rhamnosus, plantarum, curvatus or gasseri in diet-induced obese (DIO) mice has been associated with a multitude of benefits regarding adiposity and its related complications, such as a reduction in weight and visceral fat as well as a decrease in glucose, insulin and triglyceride levels. These probiotics have also been associated with a reduction in insulin resistance (IR) and proinflammatory cytokines, as well as a concomitant increase in interleukin-10 (IL-10) and an improvement in fatty liver indices [43,50,51,52,53,54,55]. Regarding human studies, a 12-week supplementation of L. gasseri BNR17 in adults with overweight or grade I obesity resulted in significant reductions in visceral fat and weight circumference in comparison to a placebo without affecting circulating biochemical profiles [56]. In another randomized, placebo-controlled trial, a 24-week supplementation of the L. rhamnosus CGMCC1.3724 strain to individuals with obesity led to significant weight reductions among female participants [33]. On the other hand, the integration of an L. rhamnosus probiotic supplement on a 20-week, low-glycemic index, hypocaloric dietary intervention in women with overweight or obesity and polycyctic ovary syndrome had no additive effects on anthropometry and circulating lipid profiles after 20 weeks [57]. Additionally, a recent meta-analysis of nine clinical trials on Lactobacillus supplementation in individuals who are overweight and obese has shown beneficial effects on total and low-density lipoprotein (LDL) cholesterol, fasting glucose and triglyceride concentrations [58].
The supplementation of different Bifidobacterium species has been studied in rodent obesity models with strain-dependent effects. In DIO mice, Bifidobacteria FS31-12, L66-5, M13-4, and L75-4 reduced serum triglycerides and liver fat content, although only L66-5 showed weight-loss properties [59]. Furthermore, B. adolescentis supplementation has shown weight-lowering effects and an improvement in steatosis and steatohepatitis in mouse and rat models [60,61]. The treatment of rodents with other Bifidobacterium strains has resulted in the reduction in visceral fat and IR, as well as glucose-lowering and diabetes-preventing effects [62,63,64]. In human trials, the supplementation of Bifidobacteria-containing probiotic formulations (B. bifidum; B. breve, B. longum, B. animalis subsp. lactis) has been associated with improvements in adiposity, glycemia, insulin resistance and fatty liver indices, with potential species-specific effects [65].
The administration of Saccharomyces boulardii to db/db mice with diabetes for 4 weeks resulted in reductions in the body weight and fat mass, improvements in fatty liver, and lowering of inflammatory indices. These effects were likely mediated through the modulation of gut microbiota [45]. Furthermore, S. boulardii reduced oxidative stress and liver injury in mice with streptozotocin-induced diabetes mellitus [66]. In a placebo-controlled randomized trial, the co-administration of S. boulardii and superoxide dismutase for 60 days among patients with obesity resulted in a reduced fat mass with the preservation of lean mass and improvements in IR [67].
3. Next-Generation Probiotics: The Contribution of Technology
The majority of microorganisms inhabiting the gut have remained unidentified until recently, as they are mostly anaerobic bacteria, which are difficult to cultivate [68]. However, modern microbiology has revolutionized our ability to identify previously unidentified microbes of the gut microbiota. In particular, with the advent of the polymerase chain reaction (PCR) of the 16 S rRNA gene, as well as next-generation sequencing (NGS) and the use of bioinformatics, we are now able to accurately identify various bacterial strains. Thus, PCR and NGS, with the use of various platforms, such as Illumina, have made possible the detection of the bacteria residing in the gut [69]. In this way, we can characterize the microorganisms associated with several disease states, including obesity and obesity-related disorders. In addition, we can identify species that are less abundant in obesity and its associated disorders, which may be associated with beneficial effects regarding obesity. Among these, Akkermansia muciniphila, Faecalibacterium prausnitzii, Eubacterium hallii (recently reclassified into Anaerobutyricum hallii and Anaerobutyricum soenhgenii [70], Bacteroides uniformis, Bacteroides coprocola, Parabacteroides distasonis, Parabacteroides goldsteinii, Hafnia alvei, Odoribacter laneus and Christensenella minuta have been proposed as NGPs, which could prove especially useful in the treatment of obesity and obesity-associated disorders [71]. Table 1 depicts major studies investigating the administration of various NGPs and their effects on overweight/obese patients. Notably, apart from culturomics, the multi-omics approach, i.e., the identification of genes (genomics), their phenotypes (epigenomics), mRNA (transcriptomics), proteins (proteomics) and various metabolites (metabolomics), can improve our understanding of the composition of the gut microbiota [71]. Collectively, high-throughput technology has shed light on the interplay between bacteria, viruses, fungi, archaea and protozoa comprising the gut microbiota.

Table 1.
List of main studies associating the administration of NGPs and amelioration of metabolic dysfunction parameters.
4. Preventive and Therapeutic Potential of NGPs in Obesity and Obesity-Related Disorders
4.1. Akkermansia muciniphila
Among NGPs, Akkermansia muciniphila (A. muciniphila) has been the best-studied and perhaps the most promising NGP until now. A. muciniphila is a Gram-negative, anaerobic and non-motile, oval-shaped bacterium which belongs to the phylum Verrucomicrobia [72]. It colonizes the gut very early, during the first year of life, via human milk [73]. Thereby, it is highly abundant in the gut of infants and healthy adults, accounting for 0.5–5% of the human gut microbiota [72,73,74]. Its beneficial effects are mainly attributed to its ability to degrade mucin: a major component of mucus in the gut [74]. The degradation of mucin leads to a counterbalanced increased production of mucin in the gut, which results in the restoration of intestinal barrier integrity [75,76]. A. muciniphila uses mucin as a carbon and nitrogen source, as well as an energy source. It improves intestinal barrier integrity by the increased expression of tight junction proteins such as claudin-3, -4, -15 and occludin [75,76,77]. Furthermore, it reduces the production of bacterial lipopolysaccharide (LPS) in the gut, exerting anti-inflammatory effects [78,79]. Notably, its administration to overweight/obese individuals has been shown to decrease serum LPS levels as well as white blood cell count (WBC) [78,79]. These reductions have been associated with the anti-inflammatory properties of A. muciniphila [78,79].
Being an anaerobe bacterium, A. muciniphila requires specific conditions in order to be cultivated. Apart from the anaerobic or microaerophilic environment, another requirement is the addition of mucin from animal sources. However, since the first isolation of A. muciniphila in 2004, novel synthetic media have been applied to facilitate its cultivation. These synthetic media do not preclude their administration to human beings, as safety is always an absolute priority for NGPs [72,80,81]. In a comparison between this synthetic medium and an animal mucin-based counterpart, it was documented that A. muciniphila was cultivated in both media and was equally effective in mice. In particular, Plovier et al. demonstrated that its administration in rodents was associated with less weight gain and improved glucose tolerance [80]. In addition, they showed that, contrary to the complete inactivation of A. muciniphila by autoclaving, pasteurization resulted in the sustained and surprisingly augmented beneficial effects of this NGP. More specifically, rodents fed with a high lipid diet, which received a pasteurized form, exhibited the same weight gain as mice who were fed with a controlled diet. This augmentation was attributed to a decrease in the energy absorption caused by this NGP. In addition, the pasteurized form improved glucose tolerance and hepatic insulin resistance; it also diminished diet-induced endotoxemia [80].
Apart from its use in animal models, A. muciniphila has been administered to overweight/obese participants. In a landmark study, Depommier et al. administered 1010 bacteria daily for 3 months in the alive or pasteurized form in 40 participants. Thirty-two overweight/obese participants completed the study. The administration of A. muciniphila for 3 months was safe and well tolerated. The pasteurized form of A. muciniphila improved insulin sensitivity, insulinemia by approximately 30%, and total cholesterol levels while decreasing body weight and fat mass as well [79]. Moreover, decreases in serum gamma-glutamyl-transferase (γ-GT) and serum aspartate-aminotransferase (AST) levels were noted in the group that received A. muciniphila in its pasteurized form compared to a placebo. These improvements in hepatic indices, especially the 24% decrease in serum γ-GT levels, may indicate an amelioration in the deposition of fatty acids in the liver and, thus, the beneficial effect of non-alcoholic fatty liver disease (NAFLD) [79,80,81]. In addition, WBC, as well as serum LPS levels, were also statistically significantly reduced [79]. Regarding adverse effects, nausea and flatulence were rarely reported [79].
Overall, the administration of A. muciniphila as an NGP may have a promising future in the treatment of obesity and its related disorders [72,73,74,75,76,78,79,80,81,82] (Figure 1).
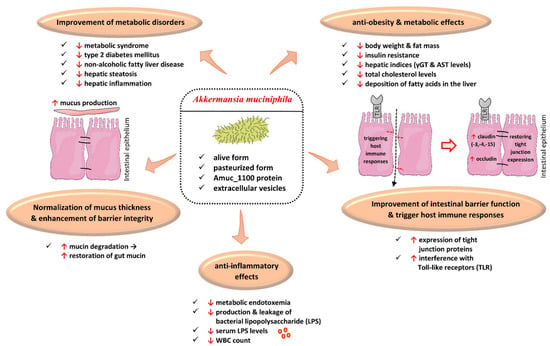
Figure 1.
Effects of Akkermansia muciniphila and its derived products in obesity and its related disorders. ↑, augmentation; ↓, reduction; AST, aspartate-aminotransferase; γGT, Gamma-Glutamyl-Transferase; WBC, white blood cell. (All images originate from the free medical site http://smart.servier.com/ by Servier licensed under a Creative Commons Attribution 3.0 Unported License; accessed on 2 February 2023).
However, there are many issues still to be resolved. A pili-like protein on the outer membrane of A. muciniphila named Amuc_1100 was demonstrated to interfere with Toll-like receptors 2 and 4 (TLR-2 and TLR-4) [83,84,85,86], which are associated with host immune responses. With the use of proteomics, the Amuc_1100 protein has been shown to be among the most abundant proteins in the outer membrane of A. muciniphila. In addition, A. muciniphila extracellular vesicles can play a crucial role in host–bacteria interactions. For example, these extracellular vesicles have been implicated in a reduction in body weight as well as in the alleviation of inflammation and the amelioration of intestinal barrier integrity in obese animal models [87]. However, to date, it remains unknown whether these extracellular vesicles contain the Amuc_1100 protein or other proteins that exhibit the abovementioned potential [83,84,85,86].
These unanswered questions must be clarified before the wide therapeutic application of A. muciniphila, either alive, in its pasteurized form, or as extracellular vesicles for the treatment of obesity and its related disorders. As there is a growing interest in NGPs regarding obesity and obesity-associated disorders, future research will shed light on unresolved issues.
4.2. Faecalibacterium prausnitzii
Faecalibacterium prausnitzii (F. prausnitzii) is a Gram-positive, anaerobe, non-motile rod which accounts for 5–15% of the human gut microbiota composition [88,89,90]. Despite its relatively high abundance in healthy adults, it has been documented to decrease among patients with obesity, metabolic syndrome and NAFLD [90,91,92,93]. Its presence has been associated with anti-inflammatory properties as well as improved gut barrier integrity [21,25,93,94]. Indeed, F. prausnitzii’s administration in animal models has resulted in improvements in indices of NAFLD [95]. More specifically, among the 12 strains administered to HFD-mice (high-fat diet-fed mice) for 3 months, five specific strains led to an amelioration of various metabolic characteristics, such as glucose intolerance, hepatic steatosis, inflammation and oxidative stress in an animal model of NAFLD. Moreover, two strains, LC49 and LB8, increased the production of SCFAs and modified the composition of gut microbiota [95]. Nowadays, it has been widely accepted that the increased production of SCFAs, such as butyric acid, propionate and acetate, is implicated in the restoration of the imbalance of gut homeostasis. SCFAs are suggested to be amongst the major contributing factors of the homeostatic state (gut symbiosis) as opposed to the state of gut dysbiosis [21,25,94,95,96]. Collectively, F. prausnitzii has been associated with an enhancement in SCFAs, especially butyrate, and an amelioration of NAFLD markers. In addition, it has been documented to inhibit the activation of Nuclear Factor kappa B (NF-κB), thereby, exerting anti-inflammatory properties and diminishing chronic inflammation in the gut [97,98]. Apart from its anti-inflammatory potential, F. prausnitzii can promote the integrity of the intestinal barrier by increasing the tight junction proteins occludin and E-cadherin. In this way, it can mitigate the permeability of the gut barrier, enhancing the production of mucous [98,99].
4.3. Eubacterium hallii
Eubacterium hallii (E. hallii) is a Gram-positive, anaerobe bacterium, which is capable of producing butyrate from lactate and acetate in low pH conditions, as well as from monosaccharides, in the intestine [100,101]. As a butyrate producer, it has gained much attention lately as a potential NGP. Udayappan et al. administered E. hallii in rodents in a live and heat-killed bacterial form. The alive form of E. hallii was documented to increase the production of butyrate, improve the metabolism of bile acid and modulate the composition of the gut microbiota in diabetic mice [102]. Furthermore, the live form of E. hallii ameliorated insulin sensitivity while increasing energy expenditure in diabetic mice [102]. It has been demonstrated that glucose intolerance and hyperglycemia may increase the intestinal barrier’s permeability, leading to chronic gut inflammation [103,104]. Therefore, this improvement in insulin sensitivity and energy expenditure achieved by the administration of E. hallii may account for its potential beneficial effects [102,103,104]. Nevertheless, despite its promising features, there is a long way ahead before E. hallii can be used as an NGP.
4.4. Bacteroides uniformis and Bacteroides coprocola
Bacteroides are Gram negative, obligatory anaerobe rods, with a high abundance in the human gut microbiota, which are estimated to comprise the vast majority of its composition [105]. Yan et al. enrolled 41 individuals with a normal body mass index (BMI) divided into two groups: one with a low visceral fat area (L-VFA) and one with a high visceral fat area (H-VFA) according to computed tomography estimations. They further studied gut composition in the two groups and found significant differences. In particular, with the use of whole genome shotgun sequencing, they showed that Bacteroides plebeius (B. plebeius) and Bacteroides uniformis (B. uniformis) were highly abundant in those with L-VFA. Therefore, they concluded that B. plebeius and B. uniformis might serve as NGPs. Furthermore, B. uniformis was negatively correlated with serum LDL-cholesterol levels [106]. In general, B. uniformis has already been documented to possess beneficial properties against obesity and metabolic disorders [107]. B. uniformis has been related to weight loss, reduced serum cholesterol and triglycerides levels and an improvement in hepatic steatosis in animal models. In addition, it has been associated with decreased serum leptin levels, a fasting glucose concentration, and improved glucose tolerance [107]. B. plebeius has been suggested as a potential therapeutic target for patients with type 2 diabetes mellitus (T2DM) based on its considerably higher abundance in patients with obesity and T2DM compared with the controls [108]. Bacteroides coprocola (B. coprocola) has been negatively associated with liver stiffness among male patients with metabolic dysfunction-associated fatty liver disease (MAFLD) [108]. In particular, among 85 individual males with MAFLD, patients with increased liver stiffness measurements (LSM), as defined by LSM > 7.4 kPa, had a decreased abundance of B. coprocola in the gut using whole genome sequencing [109]. Furthermore, Cuffaro et al. showed that B. coprocola AS101, B. uniformis PF-BaE8 and PF-BaE13 strains exhibited combined anti-inflammatory properties as well as an improvement in the intestinal barrier function [71]. Therefore, these specific strains might be NGP candidates for future research in obesity and its associated disorders.
4.5. Parabacteroides distasonis and Parabacteroides goldsteinii
Parabacteroides are Gram-negative, obligatory anaerobe, non-motile rods that colonize the human intestine [110]. Parabacteroides distasonis (P. distansonis) may be another promising NGP candidate due to its ability to reduce weight gain, hyperglycemia and hepatic steatosis in obese and HFD-mice [111]. More specifically, the oral administration of P. distasonis CGMCC 1.30169 reduced liver cholesterol, free fatty acids and triglyceride concentrations while also enhancing the production of succinate and secondary bile acids in the gut. In this animal model, all the beneficial properties resulted from the administration of living rather than heat-killed P. distasonis [111]. In addition, P. distasonis AS93 was demonstrated to exhibit anti-inflammatory properties as well as protective effects on the gut barrier function. Moreover, the same study reported that it also promoted the secretion of GLP-1 [71]. In addition, P. distasonis was suggested to exert its anti-inflammatory effects through the inhibition of NF-κB and the PI3K/Akt/mTOR pathways [112,113]. P. goldsteinii has been shown to correlate with HDL-cholesterol and IL-10, whereas it was inversely related to serum glucose and triglyceride levels in HFD-mice [114,115]. In addition, it has been related to less weight gain and reductions in inflammatory indices, such as the Tumor Necrosis Factor (TNF)-a and interleukin (IL)-1β [114,115]. Moreover, the oral administration of a live form of P. goldsteinii ATCC BAA-1180 in HFD-mice has been associated with reduced weight gain and the amelioration of various metabolic parameters, such as inflammation and insulin resistance [114,115]. Overall, despite the fact that both P. distasonis and P. goldsteinii have just started to be in the spotlight of microbiome research, they have shown promising effects regarding obesity and its related disorders.
4.6. Hafnia alvei
Hafnia alvei (H. alvei) is a Gram-negative, facultative anaerobic rod. The genus Hafnia is one of 40 members of the Enterobacterales order [116,117]. Lucas et al. reported that the oral administration of the H. alvei HA4597TM strain in HFD-mice resulted in reduced weight and fat mass gain and decreased food intake [116]. In addition, the group that was administered with H. alvei HA4597TM showed decreases in serum glucose, triglycerides and anino-transferase levels [116]. Legrand et al. found that oral gavage with H. alvei in ob/ob and HFD-mice for 18 and 46 days, respectively, decreased the fat mass in both mice and reduced food intake in hyperphagic ob/ob mice. Overall, the administration of H. alvei HA4597TM was well tolerated and had weight-lowering effects [117]. Based on the abovementioned findings, Dechelotte et al. embarked on the study of 236 overweight patients who were administered H. alvei HA4597®® in conjunction with a moderately hypocaloric diet for 12 weeks. They observed a significant weight loss of at least 3% and even 4% of the baseline body weight [118]. This constitutes an important finding, considering that a weight loss of 3% to 5% of body weight is recommended by international guidelines for overweight patients [119]. This has also been related to reductions in the prevalence of T2DM and cardiovascular risk [119]. In addition, Dechelotte et al. documented a decrease in hip circumference and a subtle reduction in waist circumference and serum glucose levels [118]. The observed decrease in body weight may be attributed to metabolic properties in the eating behavior of the ClpB protein produced by H. alvei [118]. Notably, the development of this NGP has been based on the overproduction of the ClpB protein to enhance satiety pathways via the activation of the melanocortin receptor [117]. Indeed, the melanocortin network plays a crucial role in energy metabolism. This is mediated through the transmission of anorexigenic signals as well as an increase in the energy expenditure associated with peripheral lipolytic effects [118,119,120,121,122,123,124]. It is noteworthy that melanocortin receptors are also present in the intestinal mucosa, where they are readily accessible to gut bacteria-derived products, such as ClpB [118,119,120,121,122,123,124]. Other studies have also confirmed that the ClpB-like gene function in fecal microbiota is inversely associated with BMI and fat mass. In addition, obese patients have been documented to exhibit a low abundance of bacterial taxa expressing ClpB with the alpha-Melanocyte-stimulating hormone (MSH) homology. Overall, these findings support the fact that a high abundance of bacteria expressing the ClpB gene in the gut microbiota may be associated with increased satiety and decreased body weight [118,119,120,121,122,123,124]. Therefore, H. alvei could be a promising NGP candidate for obesity and its related disorders.
4.7. Odoribacter laneus
Odoribacter laneus (O. laneus) is a Gram-negative anaerobe rod that was isolated from the human feces of a healthy Japanese adult donor in 2010 [125]. Very recently, in 2022, Hueber-Ruano et al. characterized O. laneus as an NGP due to its ability to ameliorate insulin sensitivity and decrease inflammatory markers in a mouse model [126]. By administering O. laneus through oral gavage in obese mice, they documented a significant reduction in serum succinate levels. They concluded that excess succinate might be the result of a disrupted intestinal barrier integrity, especially in cases with gut dysbiosis. The augmentation of serum succinate levels could be attributed to the increased permeability of the gut barrier in the context of a leaking gut. Therefore, the administration of succinate-consuming bacteria, such as O. laneus, could be beneficial in T2DM and obesity. The increase in serum succinate levels has been associated with T2DM and obesity, characterized by chronic low-grade inflammation [127,128]. Additionally, succinate receptor 1 (SUCNR1) may play a pivotal role in the pathogenesis of T2DM and obesity. Indeed, it has been documented that the activation of SUCNR1 in macrophages could lead to the inflammation of adipose tissue and its infiltration with inflammatory cells [127,128]. Overall, O. laneus may present a promising potential based on its succinate-consuming properties.
4.8. Christensenella minuta
Christensenella minuta (C. minuta) is a Gram-negative, non-motile and non-spore-forming anaerobe rod belonging to the order Clostridiales of the family Christensenellaceae [129,130]. The family Christensenellaceae has been related to decreases in serum LDL-cholesterol, triglycerides, and alanine-aminotransferase (ALT) levels [131]. In addition, it is highly abundant in the gut of lean individuals [131,132]. Therefore, C. minuta has been proposed as an NGP. C. minuta DSM33407 has been very recently studied as a potential therapeutic agent in a diet-induced obesity animal model. Indeed, Mazier et al. confirmed that C. minuta DSM33407 decreased the obesogenic effects of diet in a mouse model while it improved glycemia and serum leptin levels, inhibited de novo lipogenesis and ameliorated the intestinal barrier integrity. Moreover, in a humanized simulator of the human intestinal microbial ecosystem model, they demonstrated that anti-obesity effects were also related to a decrease in the Furmicutes/Bacteroidetes ratio [130]. In a plethora of studies, the Firmicutes/Bacteroidetes ratio has been found to be increased in obesity. On the contrary, there is a decrease in the Firmicutes/Bacteroidetes ratio after weight loss in individuals with obesity [130,131,132,133].
4.9. Parabacteroides distasonis
P. distansonis is a Gram-negative, non-sporeforming, anaerbobic, rod-shaped bacterium that was originally isolated in the 1930s form clinical specimens and reclassified to the genus Parabacteroides in 2006 [134]. Wang et al. reported that treatment of ob/ob and DIO mice with P. distansonis resulted in weight loss and improvements in insulin sensitivity and hepatic steatosis. This was mediated through the production of succinate in the gut, which is bound to fructose-1,6-bisphosphatase, stimulating intestinal gluconeogenesis, as well as the activation of the Farnesoid X receptor pathway by secondary bile acids [111]. In humans, P. distasonis has been found to be more abundant in children with obesity compared to those with obesity and metabolic syndrome [135]. Haro et al. reported a significant increase in P. distansonis abundance in men with obesity after a 12-month adherence to a Mediterranean diet. It should be noted that no significant changes in anthropometric parameters were noted over the course of the study; however, there was an increase in the Insulin Sensitivity Index at the end of the intervention compared to the baseline, indicating improvements in whole-body insulin sensitivity [136]. Nevertheless, S. distansonis has not been universally associated with a beneficial metabolic profile; metformin treatment has been shown to decrease its abundance in high-fat-fed mice [137], while free access to a sugar-sweetened beverage in adolescent mice produced the opposite effect [138]. Furthermore, in contrast to the previously presented results, an increased abundance of S. distansonis was noted in patients with obesity and T2DM compared to lean control individuals in a case–control study by Yang et al. [139], while similar findings were reported in women with gestational diabetes mellitus by Kuang et al. [140].
5. Safety Profiles and Implementation of NGPs in the Prevention and Treatment of Obesity and Obesity-Related Disorders
NGPs are mostly anaerobic rods that are difficult to cultivate in practice. Additionally, there are other issues that need to be clarified. First, the exact dose of each NGP to exert a beneficial effect on humans needs to be determined in colony-forming units (CFU). In the study by Depommier et al., the dose was defined as 1010 CFU for A. muciniphila [78]. The exact dose of other NGPs in CFU also needs to be determined. Second, the optimal formulation of each NGP needs to be ascertained in order to increase the survival of live bacteria during storage and transit through the unfavorable conditions of the gastrointestinal microenvironment (low pH, peptic enzymes, bile acids, etc.) as well as their engraftment into the intestinal flora, when needed [141]. Third, more studies on the co-administration of two or more NGPs are required in order to determine synergistic actions which augment their beneficial potential. It is tempting to speculate that a combination of H. alvei HA4597®®, A. muciniphila, and other probiotics may provide beneficial additive or synergistic anti-obesity and metabolic effects in patients with obesity. The combination of two or more NGPs may maximize their beneficial effects; nevertheless, more studies are required to delineate any subsequent adverse effects. Probiotic-associated adverse effects, although scarce and typically of mild severity, are likely under-reported in human trials [142]. The available reports showing mild gastrointestinal disorders have been derived from the use of probiotics for indications other than obesity and dysmetabolism. Rarely, more severe complications due to invasive infections by formulated probiotic strains, such as bacteremia, overt sepsis or endocarditis, have been noted and typically observed in immunocompromised patients. Other speculative concerns include the lateral transmission of antibiotic-resistant genes from probiotic strains to gut bacteria and excessive immune stimulation in susceptible individuals, which may lead to autoimmune or autoinflammatory phenomena, although, to date, no such events have been reported [143]. Subsequently, safety regulations need to be strictly implemented and updated. For example, in the European Union, the regulations from the European Parliament and the Council of the European Union, regulation number 1924/2006, should be followed. In the United States, regulations from the Food and Drug Administration (FDA), with the characterization of “generally recognized as safe” (GRAS), should be implemented [69]. However, as new NGPs are currently emerging, the regulation of these should be constantly updated, while the presumed safety of NGPs should be harmonized with the needs as well as the safety of individuals who are overweight/obese [69,144].
6. Conclusions
As overweight/obesity poses a major public health problem worldwide, there is currently an urgent need for more preventive and therapeutic agents in our armamentarium against obesity and its related disorders. In this context, diet is the cornerstone of preventive and therapeutic interventions. A diet rich in fiber, such as vegetables and fruits, and low in animal proteins and fat should be adopted and followed. The Mediterranean diet and, lately, ketogenic diets are increasingly being recognized as important parameters in the fight against obesity [145,146,147]. Besides the use of agents with weight loss properties, such as GLP-1 analogs or tirzepatide [148,149], which possess agonist activities in both GLP-1 and glucose-dependent insulinotropic polypeptide (GIP) receptors, there is an emerging interest in the use of NGPs. The advent of sophisticated molecular methods has revolutionized the study of NGPs as potential preventive and therapeutic anti-obesity agents. Notably, there are numerous, mostly anaerobic bacteria that have been associated with weight loss. The study of NGPs has revealed a promising potential in the context of obesity, metabolic syndrome, NAFLD, and T2DM. However, there is much to be conducted in this research field. Currently available data have emerged from animal studies that may not present translational potential in humans [150]. Additionally, corresponding human trials are limited by their small sample sizes, monocentric design and short follow-up duration. The latter is particularly relevant regarding the assessment of adverse events, which are, in turn, likely to be under-reported [142]. Safety issues, as well as more eligible international regulations regarding their use, have to be clarified and implemented in the near future. Further large-scale, longitudinal studies with a multi-omics approach are mandatory in order to shed light on their therapeutic potential in obesity and obesity-associated disorders.
Author Contributions
N.G.V. conceived the idea of the manuscript, was involved in literature search, and wrote major parts of the manuscript. D.K., D.T., F.P., A.E., I.K. and M.D. were involved in writing parts of the manuscript. G.S.C. prepared the Table and figures of the manuscript. M.D. planned the content of the review and reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
ALT: Alanine-aminotransferase; AST: Aspartate-aminotransferase; BMI: Body Mass Index; CFU: Colony Forming Units; γGT: Gamma-Glutamyl-Transferase; FDA: Food and Drug Administration; GIP: Glucose-dependent Insulinotropic Polypeptide; GLP-1: Glucagon-like peptide-1; GRAS: Generally Recognized As Safe; HFD: High Fat Diet; H-VFA: High Visceral Fat Area; IL-10: Interleukin-10; IR: Insulin Resistance; LDL: Low-density lipoprotein; LSM: Liver Stiffness Measurement; LPS: Lipopolysaccharide; L-VFA: Low Visceral Fat Area; MAFLD: Metabolic dysfunction Associated Fatty Liver Disease; MSH: Melanocyte-stimulating hormone; NAFLD: Non Alcoholic Fatty Liver Disease; NF-κB: Nuclear Factor kappa B; NGPs: Next-Generation Probiotics; NGS: Next-Generation Sequencing; PCR: Polymerase Chain Reaction; SCFAs: Short Chain Fatty Acids; SUCNR1: Succinate receptor 1; T2DM: Type 2 Diabetes Mellitus; TNF-α: Tumor Necrosis Factor-alpha; WBC: white blood cell count; WHO: World Health Organization.
References
- Powell-Wiley, T.M.; Poirier, P.; Burke, L.E.; Després, J.P.; Gordon-Larsen, P.; Lavie, C.J.; Lear, S.A.; Ndumele, C.E.; Neeland, I.J.; Sanders, P.; et al. Obesity and Cardiovascular Disease: A Scientific Statement From the American Heart Association. Circulation 2021, 143, e984–e1010. [Google Scholar] [CrossRef] [PubMed]
- Marouga, A.; Dalamaga, M.; Kastania, A.N.; Antonakos, G.; Thrasyvoulides, A.; Kontelia, G.; Dimas, C.; Vlahakos, D.V. Correlates of serum resistin in elderly, non-diabetic patients with chronic kidney disease. Clin. Lab. 2013, 59, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Paroutoglou, K.; Papadavid, E.; Christodoulatos, G.S.; Dalamaga, M. Deciphering the Association Between Psoriasis and Obesity: Current Evidence and Treatment Considerations. Curr. Obes. Rep. 2020, 9, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Hroussalas, G.; Kassi, E.; Dalamaga, M.; Delimaris, I.; Zachari, A.; Dionyssiou-Asteriou, A. Leptin, soluble leptin receptor, adiponectin and resistin in relation to OGTT in overweight/obese postmenopausal women. Maturitas 2008, 59, 339–349. [Google Scholar] [CrossRef]
- Tsigalou, C.; Vallianou, N.; Dalamaga, M. Autoantibody Production in Obesity: Is There Evidence for a Link Between Obesity and Autoimmunity? Curr. Obes. Rep. 2020, 9, 245–254. [Google Scholar] [CrossRef]
- Papadavid, E.; Gazi, S.; Dalamaga, M.; Stavrianeas, N.; Ntelis, V. Palmoplantar and scalp psoriasis occurring during anti-tumour necrosis factor-alpha therapy: A case series of four patients and guidelines for management. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 380–382. [Google Scholar] [CrossRef]
- Dalamaga, M.; Karmaniolas, K.; Chamberland, J.; Nikolaidou, A.; Lekka, A.; Dionyssiou-Asteriou, A.; Mantzoros, C.S. Higher fetuin-A, lower adiponectin and free leptin levels mediate effects of excess body weight on insulin resistance and risk for myelodysplastic syndrome. Metabolism 2013, 62, 1830–1839. [Google Scholar] [CrossRef]
- Kassi, E.; Dalamaga, M.; Faviou, E.; Hroussalas, G.; Kazanis, K.; Nounopoulos, C.; Dionyssiou-Asteriou, A. Circulating oxidized LDL levels, current smoking and obesity in postmenopausal women. Atherosclerosis 2009, 205, 279–283. [Google Scholar] [CrossRef]
- Pavlidou, A.; Dalamaga, M.; Kroupis, C.; Konstantoudakis, G.; Belimezi, M.; Athanasas, G.; Dimas, K. Survivin isoforms and clinicopathological characteristics in colorectal adenocarcinomas using real-time qPCR. World J. Gastroenterol. 2011, 17, 1614–1621. [Google Scholar] [CrossRef]
- Papadavid, E.; Katsimbri, P.; Kapniari, I.; Koumaki, D.; Karamparpa, A.; Dalamaga, M.; Tzannis, K.; Boumpas, D.; Rigopoulos, D. Prevalence of psoriatic arthritis and its correlates among patients with psoriasis in Greece: Results from a large retrospective study. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 1749–1752. [Google Scholar] [CrossRef]
- Dalamaga, M.; Papadavid, E.; Basios, G.; Vaggopoulos, V.; Rigopoulos, D.; Kassanos, D.; Trakakis, E. Ovarian SAHA syndrome is associated with a more insulin-resistant profile and represents an independent risk factor for glucose abnormalities in women with polycystic ovary syndrome: A prospective controlled study. J. Am. Acad. Dermatol. 2013, 69, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, N.; Dalamaga, M.; Stratigou, T.; Karampela, I.; Tsigalou, C. Do Antibiotics Cause Obesity Through Long-term Alterations in the Gut Microbiome? A Review of Current Evidence. Curr. Obes. Rep. 2021, 10, 244–262. [Google Scholar] [CrossRef] [PubMed]
- Dalamaga, M.; Christodoulatos, G.S.; Karampela, I.; Vallianou, N.; Apovian, C.M. Understanding the Co-Epidemic of Obesity and COVID-19: Current Evidence, Comparison with Previous Epidemics, Mechanisms, and Preventive and Therapeutic Perspectives. Curr. Obes. Rep. 2021, 10, 214–243. [Google Scholar] [CrossRef] [PubMed]
- Karampela, I.; Vallianou, N.; Magkos, F.; Apovian, C.M.; Dalamaga, M. Obesity, Hypovitaminosis D, and COVID-19: The Bermuda Triangle in Public Health. Curr. Obes. Rep. 2022, 11, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Tsilingiris, D.; Vallianou, N.G.; Karampela, I.; Dalamaga, M. Vaccine induced thrombotic thrombocytopenia: The shady chapter of a success story. Metabol. Open 2021, 11, 100101. [Google Scholar] [CrossRef]
- Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight/ (accessed on 8 February 2023).
- Vallianou, N.G.; Tsilingiris, D.; Kounatidis, D.; Lempesis, I.G.; Karampela, I.; Dalamaga, M. Sodium-glucose cotransporter-2 inhibitors in obesity and associated cardiometabolic disorders: Where do we stand? Pol. Arch. Intern. Med 2022, 132, 16342. [Google Scholar] [CrossRef]
- Tsilingiris, D.; Liatis, S.; Dalamaga, M.; Kokkinos, A. The Fight Against Obesity Escalates: New Drugs on the Horizon and Metabolic Implications. Curr. Obes. Rep. 2020, 9, 136–149. [Google Scholar] [CrossRef]
- Karamanakos, G.; Kokkinos, A.; Dalamaga, M.; Liatis, S. Highlighting the Role of Obesity and Insulin Resistance in Type 1 Diabetes and Its Associated Cardiometabolic Complications. Curr. Obes. Rep. 2022, 11, 180–202. [Google Scholar] [CrossRef]
- Fotis, D.; Liu, J.; Dalamaga, M. Could gut mycobiome play a role in NAFLD pathogenesis? Insights and therapeutic perspectives. Metabol. Open 2022, 14, 100178. [Google Scholar] [CrossRef]
- Vallianou, N.; Stratigou, T.; Christodoulatos, G.S.; Dalamaga, M. Understanding the Role of the Gut Microbiome and Microbial Metabolites in Obesity and Obesity-Associated Metabolic Disorders: Current Evidence and Perspectives. Curr. Obes. Rep. 2019, 8, 317–332. [Google Scholar] [CrossRef]
- Koliaki, C.; Liatis, S.; Dalamaga, M.; Kokkinos, A. The Implication of Gut Hormones in the Regulation of Energy Homeostasis and Their Role in the Pathophysiology of Obesity. Curr. Obes. Rep. 2020, 9, 255–271. [Google Scholar] [CrossRef]
- Spyrou, N.; Vallianou, N.; Kadillari, J.; Dalamaga, M. The interplay of obesity, gut microbiome and diet in the immune check point inhibitors therapy era. Semin. Cancer Biol. 2021, 73, 356–376. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef]
- Vallianou, N.; Christodoulatos, G.S.; Karampela, I.; Tsilingiris, D.; Magkos, F.; Stratigou, T.; Kounatidis, D.; Dalamaga, M. Understanding the Role of the Gut Microbiome and Microbial Metabolites in Non-Alcoholic Fatty Liver Disease: Current Evidence and Perspectives. Biomolecules 2021, 12, 56. [Google Scholar] [CrossRef]
- Verhaar, B.J.H.; Collard, D.; Prodan, A.; Levels, J.H.M.; Zwinderman, A.H.; Bäckhed, F.; Vogt, L.; Peters, M.J.L.; Muller, M.; Nieuwdorp, M.; et al. Associations between gut microbiota, faecal short-chain fatty acids, and blood pressure across ethnic groups: The HELIUS study. Eur. Heart J. 2020, 41, 4259–4267. [Google Scholar] [CrossRef]
- Santisteban, M.M.; Qi, Y.; Zubcevic, J.; Kim, S.; Yang, T.; Shenoy, V.; Cole-Jeffrey, C.T.; Lobaton, G.O.; Stewart, D.C.; Rubiano, A.; et al. Hypertension-Linked Pathophysiological Alterations in the Gut. Circ. Res. 2017, 120, 312–323. [Google Scholar] [CrossRef]
- Silveira-Nunes, G.; Durso, D.F.; Jr, L.; Cunha, E.H.M.; Maioli, T.U.; Vieira, A.T.; Speziali, E.; Corrêa-Oliveira, R.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; et al. Hypertension Is Associated With Intestinal Microbiota Dysbiosis and Inflammation in a Brazilian Population. Front. Pharmacol. 2020, 11, 258. [Google Scholar] [CrossRef]
- Zhai, S.; Qin, S.; Li, L.; Zhu, L.; Zou, Z.; Wang, L. Dietary butyrate suppresses inflammation through modulating gut microbiota in high-fat diet-fed mice. FEMS Microbiol. Lett. 2019, 366, fnz153. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef]
- Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: What Makes It Tick as Successful Probiotic? J. Fungi 2020, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.; Darimont, C.; Drapeau, V.; Emady-Azar, S.; Lepage, M.; Rezzonico, E.; Ngom-Bru, C.; Berger, B.; Philippe, L.; Ammon-Zuffrey, C.; et al. Effect of Lactobacillus rhamnosus CGMCC1.3724 supplementation on weight loss and maintenance in obese men and women. Br. J. Nutr. 2014, 111, 1507–1519. [Google Scholar] [CrossRef] [PubMed]
- Toral, M.; Gómez-Guzmán, M.; Jiménez, R.; Romero, M.; Sánchez, M.; Utrilla, M.P.; Garrido-Mesa, N.; Rodríguez-Cabezas, M.E.; Olivares, M.; Gálvez, J.; et al. The probiotic Lactobacillus coryniformis CECT5711 reduces the vascular pro-oxidant and pro-inflammatory status in obese mice. Clin. Sci. 2014, 127, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tang, H.; Zhang, C.; Zhao, Y.; Derrien, M.; Rocher, E.; van-Hylckama Vlieg, J.E.; Strissel, K.; Zhao, L.; Obin, M.; et al. Modulation of gut microbiota during probiotic-mediated attenuation of metabolic syndrome in high fat diet-fed mice. ISME J. 2015, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Yun, S.I.; Park, M.H.; Park, J.H.; Jeong, S.Y.; Park, H.O. Anti-obesity effect of Lactobacillus gasseri BNR17 in high-sucrose diet-induced obese mice. PLoS ONE 2013, 8, e54617. [Google Scholar] [CrossRef]
- Kondo, S.; Kamei, A.; Xiao, J.Z.; Iwatsuki, K.; Abe, K. Bifidobacterium breve B-3 exerts metabolic syndrome-suppressing effects in the liver of diet-induced obese mice: A DNA microarray analysis. Benef. Microbes 2013, 4, 247–251. [Google Scholar] [CrossRef]
- Okubo, T.; Takemura, N.; Yoshida, A.; Sonoyama, K. KK/Ta Mice Administered Lactobacillus plantarum Strain No. 14 Have Lower Adiposity and Higher Insulin Sensitivity. Biosci. Microbiota Food Health 2013, 32, 93–100. [Google Scholar] [CrossRef]
- Park, D.Y.; Ahn, Y.T.; Park, S.H.; Huh, C.S.; Yoo, S.R.; Yu, R.; Sung, M.K.; McGregor, R.A.; Choi, M.S. Supplementation of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 in diet-induced obese mice is associated with gut microbial changes and reduction in obesity. PLoS ONE 2013, 8, e59470. [Google Scholar] [CrossRef]
- Poutahidis, T.; Kleinewietfeld, M.; Smillie, C.; Levkovich, T.; Perrotta, A.; Bhela, S.; Varian, B.J.; Ibrahim, Y.M.; Lakritz, J.R.; Kearney, S.M.; et al. Microbial reprogramming inhibits Western diet-associated obesity. PLoS ONE 2013, 8, e68596. [Google Scholar] [CrossRef]
- Sakai, T.; Taki, T.; Nakamoto, A.; Shuto, E.; Tsutsumi, R.; Toshimitsu, T.; Makino, S.; Ikegami, S. Lactobacillus plantarum OLL2712 regulates glucose metabolism in C57BL/6 mice fed a high-fat diet. J. Nutr. Sci. Vitaminol. 2013, 59, 144–147. [Google Scholar] [CrossRef]
- Yadav, H.; Lee, J.H.; Lloyd, J.; Walter, P.; Rane, S.G. Beneficial metabolic effects of a probiotic via butyrate-induced GLP-1 hormone secretion. J. Biol. Chem. 2013, 288, 25088–25097. [Google Scholar] [CrossRef]
- Yoo, S.R.; Kim, Y.J.; Park, D.Y.; Jung, U.J.; Jeon, S.M.; Ahn, Y.T.; Huh, C.S.; McGregor, R.; Choi, M.S. Probiotics, L. plantarum and L. curvatus in combination alter hepatic lipid metabolism and suppress diet-induced obesity. Obesity 2013, 21, 2571–2578. [Google Scholar] [CrossRef]
- Lindsay, K.L.; Kennelly, M.; Culliton, M.; Smith, T.; Maguire, O.C.; Shanahan, F.; Brennan, L.; McAuliffe, F.M. Probiotics in obese pregnancy do not reduce maternal fasting glucose: A double-blind, placebo-controlled, randomized trial (Probiotics in Pregnancy Study). Am. J. Clin. Nutr. 2014, 99, 1432–1439. [Google Scholar] [CrossRef]
- Everard, A.; Matamoros, S.; Geurts, L.; Delzenne, N.M.; Cani, P.D. Saccharomyces boulardii administration changes gut microbiota and reduces hepatic steatosis, low-grade inflammation, and fat mass in obese and type 2 diabetic db/db mice. mBio 2014, 5, e01011–e01014. [Google Scholar] [CrossRef]
- Salles, B.I.M.; Cioffi, D.; Ferreira, S.R.G. Probiotics supplementation and insulin resistance: A systematic review. Diabetol. Metab. Syndr. 2020, 12, 98. [Google Scholar] [CrossRef]
- Wiciński, M.; Gębalski, J.; Gołębiewski, J.; Malinowski, B. Probiotics for the Treatment of Overweight and Obesity in Humans-A Review of Clinical Trials. Microorganisms 2020, 8, 1148. [Google Scholar] [CrossRef]
- Vallianou, N.; Stratigou, T.; Christodoulatos, G.S.; Tsigalou, C.; Dalamaga, M. Probiotics, Prebiotics, Synbiotics, Postbiotics, and Obesity: Current Evidence, Controversies, and Perspectives. Curr. Obes. Rep. 2020, 9, 179–192. [Google Scholar] [CrossRef]
- Kang, Y.; Cai, Y. The development of probiotics therapy to obesity: A therapy that has gained considerable momentum. Hormones 2018, 17, 141–151. [Google Scholar] [CrossRef]
- Lee, H.Y.; Park, J.H.; Seok, S.H.; Baek, M.W.; Kim, D.J.; Lee, K.E.; Paek, K.S.; Lee, Y.; Park, J.H. Human originated bacteria, Lactobacillus rhamnosus PL60, produce conjugated linoleic acid and show anti-obesity effects in diet-induced obese mice. Biochim. Biophys. Acta 2006, 1761, 736–744. [Google Scholar] [CrossRef]
- Lee, K.; Paek, K.; Lee, H.Y.; Park, J.H.; Lee, Y. Antiobesity effect of trans-10,cis-12-conjugated linoleic acid-producing Lactobacillus plantarum PL62 on diet-induced obese mice. J. Appl. Microbiol. 2007, 103, 1140–1146. [Google Scholar] [CrossRef]
- Park, S.Y.; Seong, K.S.; Lim, S.D. Anti-obesity Effect of Yogurt Fermented by Lactobacillus plantarum Q180 in Diet-induced Obese Rats. Korean J. Food Sci. Anim. Resour. 2016, 36, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Oh, S.H.; Cha, Y.S. Lactobacillus plantarum LG42 isolated from gajami sik-hae decreases body and fat pad weights in diet-induced obese mice. J. Appl. Microbiol. 2014, 116, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Ben Salah, R.; Trabelsi, I.; Hamden, K.; Chouayekh, H.; Bejar, S. Lactobacillus plantarum TN8 exhibits protective effects on lipid, hepatic and renal profiles in obese rat. Anaerobe 2013, 23, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, M.; Ogawa, A.; Higurashi, S.; Kadooka, Y. Anti-obesity effect of Lactobacillus gasseri SBT2055 accompanied by inhibition of pro-inflammatory gene expression in the visceral adipose tissue in diet-induced obese mice. Eur. J. Nutr. 2014, 53, 599–606. [Google Scholar] [CrossRef]
- Kim, J.; Yun, J.M.; Kim, M.K.; Kwon, O.; Cho, B. Lactobacillus gasseri BNR17 Supplementation Reduces the Visceral Fat Accumulation and Waist Circumference in Obese Adults: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Med. Food 2018, 21, 454–461. [Google Scholar] [CrossRef]
- Łagowska, K.; Drzymała-Czyż, S. A low glycemic index, energy-restricted diet but not Lactobacillus rhamnosus supplementation changes fecal short-chain fatty acid and serum lipid concentrations in women with overweight or obesity and polycystic ovary syndrome. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 917–926. [Google Scholar] [CrossRef]
- Qiu, X.; Wu, Q.; Li, W.; Tang, K.; Zhang, J. Effects of Lactobacillus supplementation on glycemic and lipid indices in overweight or obese adults: A systematic review and meta-analysis. Clin. Nutr. 2022, 41, 1787–1797. [Google Scholar] [CrossRef]
- Yin, Y.N.; Yu, Q.F.; Fu, N.; Liu, X.W.; Lu, F.G. Effects of four Bifidobacteria on obesity in high-fat diet induced rats. World J. Gastroenterol. 2010, 16, 3394–3401. [Google Scholar] [CrossRef]
- Reichold, A.; Brenner, S.A.; Spruss, A.; Förster-Fromme, K.; Bergheim, I.; Bischoff, S.C. Bifidobacterium adolescentis protects from the development of nonalcoholic steatohepatitis in a mouse model. J. Nutr. Biochem. 2014, 25, 118–125. [Google Scholar] [CrossRef]
- Chen, J.; Wang, R.; Li, X.F.; Wang, R.L. Bifidobacterium adolescentis supplementation ameliorates visceral fat accumulation and insulin sensitivity in an experimental model of the metabolic syndrome. Br. J. Nutr. 2012, 107, 1429–1434. [Google Scholar] [CrossRef]
- Kondo, S.; Xiao, J.Z.; Satoh, T.; Odamaki, T.; Takahashi, S.; Sugahara, H.; Yaeshima, T.; Iwatsuki, K.; Kamei, A.; Abe, K. Antiobesity effects of Bifidobacterium breve strain B-3 supplementation in a mouse model with high-fat diet-induced obesity. Biosci. Biotechnol. Biochem. 2010, 74, 1656–1661. [Google Scholar] [CrossRef]
- Amar, J.; Chabo, C.; Waget, A.; Klopp, P.; Vachoux, C.; Bermúdez-Humarán, L.G.; Smirnova, N.; Bergé, M.; Sulpice, T.; Lahtinen, S.; et al. Intestinal mucosal adherence and translocation of commensal bacteria at the early onset of type 2 diabetes: Molecular mechanisms and probiotic treatment. EMBO Mol. Med. 2011, 3, 559–572. [Google Scholar] [CrossRef]
- Kikuchi, K.; Ben Othman, M.; Sakamoto, K. Sterilized bifidobacteria suppressed fat accumulation and blood glucose level. Biochem. Biophys. Res. Commun. 2018, 501, 1041–1047. [Google Scholar] [CrossRef]
- Koutnikova, H.; Genser, B.; Monteiro-Sepulveda, M.; Faurie, J.M.; Rizkalla, S.; Schrezenmeir, J.; Clément, K. Impact of bacterial probiotics on obesity, diabetes and non-alcoholic fatty liver disease related variables: A systematic review and meta-analysis of randomised controlled trials. BMJ Open 2019, 9, e017995. [Google Scholar] [CrossRef]
- Barssotti, L.; Abreu, I.; Brandão, A.B.P.; Albuquerque, R.; Ferreira, F.G.; Salgado, M.A.C.; Dias, D.D.S.; De Angelis, K.; Yokota, R.; Casarini, D.E.; et al. Saccharomyces boulardii modulates oxidative stress and renin angiotensin system attenuating diabetes-induced liver injury in mice. Sci. Rep. 2021, 11, 9189. [Google Scholar] [CrossRef]
- Rondanelli, M.; Miraglia, N.; Putignano, P.; Castagliuolo, I.; Brun, P.; Dall’Acqua, S.; Peroni, G.; Faliva, M.A.; Naso, M.; Nichetti, M.; et al. Effects of 60-Day Saccharomyces boulardii and Superoxide Dismutase Supplementation on Body Composition, Hunger Sensation, Pro/Antioxidant Ratio, Inflammation and Hormonal Lipo-Metabolic Biomarkers in Obese Adults: A Double-Blind, Placebo-Controlled Trial. Nutrients 2021, 13, 2512. [Google Scholar] [CrossRef]
- Amrane, S.; Hocquart, M.; Afouda, P.; Kuete, E.; Pham, T.P.; Dione, N.; Ngom, I.I.; Valles, C.; Bachar, D.; Raoult, D.; et al. Metagenomic and culturomic analysis of gut microbiota dysbiosis during Clostridium difficile infection. Sci. Rep. 2019, 9, 12807. [Google Scholar] [CrossRef]
- López-Moreno, A.; Acuña, I.; Torres-Sánchez, A.; Ruiz-Moreno, Á.; Cerk, K.; Rivas, A.; Suárez, A.; Monteoliva-Sánchez, M.; Aguilera, M. Next Generation Probiotics for Neutralizing Obesogenic Effects: Taxa Culturing Searching Strategies. Nutrients 2021, 13, 1617. [Google Scholar] [CrossRef]
- Shetty, S.A.; Zuffa, S.; Bui, T.P.N.; Aalvink, S.; Smidt, H.; De Vos, W.M. Reclassification of Eubacterium hallii as Anaerobutyricum hallii gen. nov., comb. nov., and description of Anaerobutyricum soehngenii sp. nov., a butyrate and propionate-producing bacterium from infant faeces. Int. J. Syst. Evol. Microbiol. 2018, 68, 3741–3746. [Google Scholar] [CrossRef]
- Cuffaro, B.; Assohoun, A.L.W.; Boutillier, D.; Peucelle, V.; Desramaut, J.; Boudebbouze, S.; Croyal, M.; Waligora-Dupriet, A.J.; Rhimi, M.; Grangette, C.; et al. Identification of New Potential Biotherapeutics from Human Gut Microbiota-Derived Bacteria. Microorganisms 2021, 9, 565. [Google Scholar] [CrossRef]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Isolauri, E.; Laitinen, K.; Salminen, S. Distinct composition of gut microbiota during pregnancy in overweight and normal-weight women. Am. J. Clin. Nutr. 2008, 88, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Sunder, S.; Verma, S.R. Disease-associated dysbiosis and potential therapeutic role of Akkermansia muciniphila, a mucus degrading bacteria of gut microbiome. Folia Microbiol. 2022, 67, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Reunanen, J.; Kainulainen, V.; Huuskonen, L.; Ottman, N.; Belzer, C.; Huhtinen, H.; de Vos, W.M.; Satokari, R. Akkermansia muciniphila Adheres to Enterocytes and Strengthens the Integrity of the Epithelial Cell Layer. Appl. Environ. Microbiol. 2015, 81, 3655–3662. [Google Scholar] [CrossRef]
- Shi, M.; Yue, Y.; Ma, C.; Dong, L.; Chen, F. Pasteurized Akkermansia muciniphila Ameliorate the LPS-Induced Intestinal Barrier Dysfunction via Modulating AMPK and NF-κB through TLR2 in Caco-2 Cells. Nutrients 2022, 14, 764. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Cani, P.D.; Depommier, C.; Derrien, M.; Everard, A.; de Vos, W.M. Akkermansia muciniphila: Paradigm for next-generation beneficial microorganisms. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 625–637. [Google Scholar] [CrossRef]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef]
- Everard, A.; Plovier, H.; Rastelli, M.; Van Hul, M.; de Wouters d’Oplinter, A.; Geurts, L.; Druart, C.; Robine, S.; Delzenne, N.M.; Muccioli, G.G.; et al. Intestinal epithelial N-acylphosphatidylethanolamine phospholipase D links dietary fat to metabolic adaptations in obesity and steatosis. Nat. Commun. 2019, 10, 457. [Google Scholar] [CrossRef]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef]
- Ottman, N.; Reunanen, J.; Meijerink, M.; Pietilä, T.E.; Kainulainen, V.; Klievink, J.; Huuskonen, L.; Aalvink, S.; Skurnik, M.; Boeren, S.; et al. Pili-like proteins of Akkermansia muciniphila modulate host immune responses and gut barrier function. PLoS ONE 2017, 12, e0173004. [Google Scholar] [CrossRef]
- Kuipers, M.E.; Hokke, C.H.; Smits, H.H.; Nolte-‘t Hoen, E.N.M. Pathogen-Derived Extracellular Vesicle-Associated Molecules That Affect the Host Immune System: An Overview. Front. Microbiol. 2018, 9, 2182. [Google Scholar] [CrossRef]
- Macia, L.; Nanan, R.; Hosseini-Beheshti, E.; Grau, G.E. Host- and Microbiota-Derived Extracellular Vesicles, Immune Function, and Disease Development. Int. J. Mol. Sci. 2019, 21, 107. [Google Scholar] [CrossRef]
- Yan, J.; Sheng, L.; Li, H. Akkermansia muciniphila: Is it the Holy Grail for ameliorating metabolic diseases? Gut Microbes 2021, 13, 1984104. [Google Scholar] [CrossRef]
- Chelakkot, C.; Choi, Y.; Kim, D.K.; Park, H.T.; Ghim, J.; Kwon, Y.; Jeon, J.; Kim, M.S.; Jee, Y.K.; Gho, Y.S.; et al. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp. Mol. Med. 2018, 50, e450. [Google Scholar] [CrossRef]
- Martín, R.; Miquel, S.; Benevides, L.; Bridonneau, C.; Robert, V.; Hudault, S.; Chain, F.; Berteau, O.; Azevedo, V.; Chatel, J.M.; et al. Functional Characterization of Novel Faecalibacterium prausnitzii Strains Isolated from Healthy Volunteers: A Step Forward in the Use of F. prausnitzii as a Next-Generation Probiotic. Front. Microbiol. 2017, 8, 1226. [Google Scholar] [CrossRef]
- Miquel, S.; Martín, R.; Rossi, O.; Bermúdez-Humarán, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef]
- Leylabadlo, H.E.; Ghotaslou, R.; Feizabadi, M.M.; Farajnia, S.; Moaddab, S.Y.; Ganbarov, K.; Khodadadi, E.; Tanomand, A.; Sheykhsaran, E.; Yousefi, B.; et al. The critical role of Faecalibacterium prausnitzii in human health: An overview. Microb. Pathog. 2020, 149, 104344. [Google Scholar] [CrossRef]
- Grabherr, F.; Grander, C.; Effenberger, M.; Adolph, T.E.; Tilg, H. Gut Dysfunction and Non-alcoholic Fatty Liver Disease. Front. Endocrinol. 2019, 10, 611. [Google Scholar] [CrossRef]
- Pircalabioru, G.G.; Ilie, I.; Oprea, L.; Picu, A.; Petcu, L.M.; Burlibasa, L.; Chifiriuc, M.C.; Musat, M. Microbiome, Mycobiome and Related Metabolites Alterations in Patients with Metabolic Syndrome-A Pilot Study. Metabolites 2022, 12, 218. [Google Scholar] [CrossRef] [PubMed]
- Caudet, J.; Trelis, M.; Cifre, S.; Soriano, J.M.; Rico, H.; Merino-Torres, J.F. Interplay between Intestinal Bacterial Communities and Unicellular Parasites in a Morbidly Obese Population: A Neglected Trinomial. Nutrients 2022, 14, 3211. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, N.; Liu, J.; Dalamaga, M. What are the key points in the association between the gut microbiome and nonalcoholic fatty liver disease? Metabol. Open 2019, 1, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Gao, W.; Liu, Z.; Fang, Z.; Wang, H.; Zhao, J.; Zhang, H.; Lu, W.; Chen, W. Specific Strains of Faecalibacterium prausnitzii Ameliorate Nonalcoholic Fatty Liver Disease in Mice in Association with Gut Microbiota Regulation. Nutrients 2022, 14, 2945. [Google Scholar] [CrossRef] [PubMed]
- Machiels, K.; Joossens, M.; Sabino, J.; De Preter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Van Immerseel, F.; Verbeke, K.; et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Quévrain, E.; Maubert, M.A.; Michon, C.; Chain, F.; Marquant, R.; Tailhades, J.; Miquel, S.; Carlier, L.; Bermúdez-Humarán, L.G.; Pigneur, B.; et al. Identification of an anti-inflammatory protein from Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn’s disease. Gut 2016, 65, 415–425. [Google Scholar] [CrossRef]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841–852. [Google Scholar] [CrossRef]
- Wrzosek, L.; Miquel, S.; Noordine, M.L.; Bouet, S.; Joncquel Chevalier-Curt, M.; Robert, V.; Philippe, C.; Bridonneau, C.; Cherbuy, C.; Robbe-Masselot, C.; et al. Bacteroides thetaiotaomicron and Faecalibacterium prausnitzii influence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biol. 2013, 11, 61. [Google Scholar] [CrossRef]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 2004, 70, 5810–5817. [Google Scholar] [CrossRef]
- Louis, P.; Young, P.; Holtrop, G.; Flint, H.J. Diversity of human colonic butyrate-producing bacteria revealed by analysis of the butyryl-CoA:acetate CoA-transferase gene. Environ. Microbiol. 2010, 12, 304–314. [Google Scholar] [CrossRef]
- Udayappan, S.; Manneras-Holm, L.; Chaplin-Scott, A.; Belzer, C.; Herrema, H.; Dallinga-Thie, G.M.; Duncan, S.H.; Stroes, E.S.G.; Groen, A.K.; Flint, H.J.; et al. Oral treatment with Eubacterium hallii improves insulin sensitivity in db/db mice. NPJ Biofilms Microbiomes 2016, 2, 16009. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Levy, M.; Grosheva, I.; Zheng, D.; Soffer, E.; Blacher, E.; Braverman, S.; Tengeler, A.C.; Barak, O.; Elazar, M.; et al. Hyperglycemia drives intestinal barrier dysfunction and risk for enteric infection. Science 2018, 359, 1376–1383. [Google Scholar] [CrossRef]
- Martin, A.; Devkota, S. Hold the Door: Role of the Gut Barrier in Diabetes. Cell Metab. 2018, 27, 949–951. [Google Scholar] [CrossRef]
- Wexler, H.M. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef]
- Yan, H.; Qin, Q.; Chen, J.; Yan, S.; Li, T.; Gao, X.; Yang, Y.; Li, A.; Ding, S. Gut Microbiome Alterations in Patients With Visceral Obesity Based on Quantitative Computed Tomography. Front. Cell Infect. Microbiol. 2021, 11, 823262. [Google Scholar] [CrossRef]
- Gómez Del Pulgar, E.M.; Benítez-Páez, A.; Sanz, Y. Safety Assessment of Bacteroides Uniformis CECT 7771, a Symbiont of the Gut Microbiota in Infants. Nutrients 2020, 12, 551. [Google Scholar] [CrossRef]
- Wang, T.Y.; Zhang, X.Q.; Chen, A.L.; Zhang, J.; Lv, B.H.; Ma, M.H.; Lian, J.; Wu, Y.X.; Zhou, Y.T.; Ma, C.C.; et al. A comparative study of microbial community and functions of type 2 diabetes mellitus patients with obesity and healthy people. Appl. Microbiol. Biotechnol. 2020, 104, 7143–7153. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, S.; Sheng, S.; Qin, Q.; Chen, J.; Li, W.; Li, T.; Gao, X.; Wang, L.; Ang, L.; et al. Comparison of gut microbiota in male MAFLD patients with varying liver stiffness. Front. Cell Infect. Microbiol. 2022, 12, 873048. [Google Scholar] [CrossRef]
- Sakamoto, M.; Benno, Y. Reclassification of Bacteroides distasonis, Bacteroides goldsteinii and Bacteroides merdae as Parabacteroides distasonis gen. nov., comb. nov., Parabacteroides goldsteinii comb. nov. and Parabacteroides merdae comb. nov. Int. J. Syst. Evol. Microbiol. 2006, 56, 1599–1605. [Google Scholar] [CrossRef]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids. Cell Rep. 2019, 26, 222–235.e225. [Google Scholar] [CrossRef]
- Li, M.; Hu, F.C.; Qiao, F.; Du, Z.Y.; Zhang, M.L. Sodium acetate alleviated high-carbohydrate induced intestinal inflammation by suppressing MAPK and NF-κB signaling pathways in Nile tilapia (Oreochromis niloticus). Fish Shellfish. Immunol. 2020, 98, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Qiu, Y.B.; Gao, Z.Q.; Wu, Y.X.; Wan, B.B.; Liu, G.; Chen, J.L.; Zhou, Q.; Yu, R.Q.; Pang, Q.F. Sodium Propionate Attenuates the Lipopolysaccharide-Induced Epithelial-Mesenchymal Transition via the PI3K/Akt/mTOR Signaling Pathway. J. Agric. Food Chem. 2020, 68, 6554–6563. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.R.; Lin, C.S.; Chang, C.J.; Lin, T.L.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Lu, C.C.; Young, J.D.; Lai, H.C. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut 2019, 68, 248–262. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhang, L.; Wang, X.; Yi, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Roles of intestinal Parabacteroides in human health and diseases. FEMS Microbiol. Lett. 2022, 369, fnac072. [Google Scholar] [CrossRef] [PubMed]
- Lucas, N.; Legrand, R.; Deroissart, C.; Dominique, M.; Azhar, S.; Le Solliec, M.A.; Léon, F.; do Rego, J.C.; Déchelotte, P.; Fetissov, S.O.; et al. Hafnia alvei HA4597 Strain Reduces Food Intake and Body Weight Gain and Improves Body Composition, Glucose, and Lipid Metabolism in a Mouse Model of Hyperphagic Obesity. Microorganisms 2019, 8, 15. [Google Scholar] [CrossRef]
- Legrand, R.; Lucas, N.; Dominique, M.; Azhar, S.; Deroissart, C.; Le Solliec, M.A.; Rondeaux, J.; Nobis, S.; Guérin, C.; Léon, F.; et al. Commensal Hafnia alvei strain reduces food intake and fat mass in obese mice-a new potential probiotic for appetite and body weight management. Int. J. Obes. 2020, 44, 1041–1051. [Google Scholar] [CrossRef]
- Déchelotte, P.; Breton, J.; Trotin-Picolo, C.; Grube, B.; Erlenbeck, C.; Bothe, G.; Fetissov, S.O.; Lambert, G. The Probiotic Strain H. alvei HA4597(®®) Improves Weight Loss in Overweight Subjects under Moderate Hypocaloric Diet: A Proof-of-Concept, Multicenter Randomized, Double-Blind Placebo-Controlled Study. Nutrients 2021, 13, 1902. [Google Scholar] [CrossRef]
- Jensen, M.D.; Ryan, D.H.; Apovian, C.M.; Ard, J.D.; Comuzzie, A.G.; Donato, K.A.; Hu, F.B.; Hubbard, V.S.; Jakicic, J.M.; Kushner, R.F.; et al. 2013 AHA/ACC/TOS guideline for the management of overweight and obesity in adults: A report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines and The Obesity Society. Circulation 2014, 129, S102–S138. [Google Scholar] [CrossRef]
- Anderson, E.J.; Çakir, I.; Carrington, S.J.; Cone, R.D.; Ghamari-Langroudi, M.; Gillyard, T.; Gimenez, L.E.; Litt, M.J. 60 YEARS OF POMC: Regulation of feeding and energy homeostasis by α-MSH. J. Mol. Endocrinol. 2016, 56, T157–T174. [Google Scholar] [CrossRef]
- Panaro, B.L.; Tough, I.R.; Engelstoft, M.S.; Matthews, R.T.; Digby, G.J.; Møller, C.L.; Svendsen, B.; Gribble, F.; Reimann, F.; Holst, J.J.; et al. The melanocortin-4 receptor is expressed in enteroendocrine L cells and regulates the release of peptide YY and glucagon-like peptide 1 in vivo. Cell Metab. 2014, 20, 1018–1029. [Google Scholar] [CrossRef]
- Kühnen, P.; Clément, K.; Wiegand, S.; Blankenstein, O.; Gottesdiener, K.; Martini, L.L.; Mai, K.; Blume-Peytavi, U.; Grüters, A.; Krude, H. Proopiomelanocortin Deficiency Treated with a Melanocortin-4 Receptor Agonist. N. Engl. J. Med. 2016, 375, 240–246. [Google Scholar] [CrossRef]
- Arnoriaga-Rodríguez, M.; Mayneris-Perxachs, J.; Burokas, A.; Pérez-Brocal, V.; Moya, A.; Portero-Otin, M.; Ricart, W.; Maldonado, R.; Fernández-Real, J.M. Gut bacterial ClpB-like gene function is associated with decreased body weight and a characteristic microbiota profile. Microbiome 2020, 8, 59. [Google Scholar] [CrossRef]
- Dominique, M.; Breton, J.; Guérin, C.; Bole-Feysot, C.; Lambert, G.; Déchelotte, P.; Fetissov, S. Effects of Macronutrients on the In Vitro Production of ClpB, a Bacterial Mimetic Protein of α-MSH and Its Possible Role in Satiety Signaling. Nutrients 2019, 11, 2115. [Google Scholar] [CrossRef]
- Nagai, F.; Morotomi, M.; Watanabe, Y.; Sakon, H.; Tanaka, R. Alistipes indistinctus sp. nov. and Odoribacter laneus sp. nov., common members of the human intestinal microbiota isolated from faeces. Int. J. Syst. Evol. Microbiol. 2010, 60, 1296–1302. [Google Scholar] [CrossRef]
- Huber-Ruano, I.; Calvo, E.; Mayneris-Perxachs, J.; Rodríguez-Peña, M.M.; Ceperuelo-Mallafré, V.; Cedó, L.; Núñez-Roa, C.; Miro-Blanch, J.; Arnoriaga-Rodríguez, M.; Balvay, A.; et al. Orally administered Odoribacter laneus improves glucose control and inflammatory profile in obese mice by depleting circulating succinate. Microbiome 2022, 10, 135. [Google Scholar] [CrossRef]
- Serena, C.; Ceperuelo-Mallafré, V.; Keiran, N.; Queipo-Ortuño, M.I.; Bernal, R.; Gomez-Huelgas, R.; Urpi-Sarda, M.; Sabater, M.; Pérez-Brocal, V.; Andrés-Lacueva, C.; et al. Elevated circulating levels of succinate in human obesity are linked to specific gut microbiota. ISME J. 2018, 12, 1642–1657. [Google Scholar] [CrossRef]
- Van Diepen, J.A.; Robben, J.H.; Hooiveld, G.J.; Carmone, C.; Alsady, M.; Boutens, L.; Bekkenkamp-Grovenstein, M.; Hijmans, A.; Engelke, U.F.H.; Wevers, R.A.; et al. SUCNR1-mediated chemotaxis of macrophages aggravates obesity-induced inflammation and diabetes. Diabetologia 2017, 60, 1304–1313. [Google Scholar] [CrossRef]
- Ndongo, S.; Khelaifia, S.; Fournier, P.E.; Raoult, D. Christensenella massiliensis, a new bacterial species isolated from the human gut. New Microbes New Infect. 2016, 12, 69–70. [Google Scholar] [CrossRef]
- Mazier, W.; Corf, K.L.; Martinez, C.; Tudela, H.; Kissi, D.; Kropp, C.; Coubard, C.; Soto, M.; Elustondo, F.; Rawadi, G.; et al. A New Strain of Christensenella minuta as a Potential Biotherapy for Obesity and Associated Metabolic Diseases. Cells 2021, 10, 823. [Google Scholar] [CrossRef]
- Li, X.; Li, Z.; He, Y.; Li, P.; Zhou, H.; Zeng, N. Regional distribution of Christensenellaceae and its associations with metabolic syndrome based on a population-level analysis. PeerJ 2020, 8, e9591. [Google Scholar] [CrossRef]
- Peters, B.A.; Shapiro, J.A.; Church, T.R.; Miller, G.; Trinh-Shevrin, C.; Yuen, E.; Friedlander, C.; Hayes, R.B.; Ahn, J. A taxonomic signature of obesity in a large study of American adults. Sci. Rep. 2018, 8, 9749. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Ezeji, J.C.; Sarikonda, D.K.; Hopperton, A.; Erkkila, H.L.; Cohen, D.E.; Martinez, S.P.; Cominelli, F.; Kuwahara, T.; Dichosa, A.E.K.; Good, C.E.; et al. Parabacteroides distasonis: Intriguing aerotolerant gut anaerobe with emerging antimicrobial resistance and pathogenic and probiotic roles in human health. Gut Microbes 2021, 13, 1922241. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Becerra, L.; Cornejo-Granados, F.; García-López, R.; Valdez-Lara, A.; Bikel, S.; Canizales-Quinteros, S.; López-Contreras, B.E.; Mendoza-Vargas, A.; Nielsen, H.; Ochoa-Leyva, A. Metatranscriptomic analysis to define the Secrebiome, and 16S rRNA profiling of the gut microbiome in obesity and metabolic syndrome of Mexican children. Microb. Cell Factories 2020, 19, 61. [Google Scholar] [CrossRef] [PubMed]
- Haro, C.; Montes-Borrego, M.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.F.; Gómez-Delgado, F.; Pérez-Martínez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Tinahones, F.J.; Landa, B.B.; et al. Two Healthy Diets Modulate Gut Microbial Community Improving Insulin Sensitivity in a Human Obese Population. J. Clin. Endocrinol. Metab. 2016, 101, 233–242. [Google Scholar] [CrossRef]
- Wu, H.; Wang, X.; Fang, X.; Lian, F.; Li, M.; Liao, J.; Dai, D.; Tian, J. Metformin modulates the gut microbiome in a mice model of high-fat diet-induced glycolipid metabolism disorder. BMJ Open Diabetes Res. Care 2022, 10, e003149. [Google Scholar] [CrossRef]
- Noble, E.E.; Olson, C.A.; Davis, E.; Tsan, L.; Chen, Y.W.; Schade, R.; Liu, C.; Suarez, A.; Jones, R.B.; de La Serre, C.; et al. Gut microbial taxa elevated by dietary sugar disrupt memory function. Transl. Psychiatry 2021, 11, 194. [Google Scholar] [CrossRef]
- Yang, K.; Niu, J.; Zuo, T.; Sun, Y.; Xu, Z.; Tang, W.; Liu, Q.; Zhang, J.; Ng, E.K.W.; Wong, S.K.H.; et al. Alterations in the Gut Virome in Obesity and Type 2 Diabetes Mellitus. Gastroenterology 2021, 161, 1257–1269. [Google Scholar] [CrossRef]
- Kuang, Y.S.; Lu, J.H.; Li, S.H.; Li, J.H.; Yuan, M.Y.; He, J.R.; Chen, N.N.; Xiao, W.Q.; Shen, S.Y.; Qiu, L.; et al. Connections between the human gut microbiome and gestational diabetes mellitus. Gigascience 2017, 6, gix058. [Google Scholar] [CrossRef]
- Olofsson, L.E.; Bäckhed, F. The Metabolic Role and Therapeutic Potential of the Microbiome. Endocr. Rev. 2022, 43, 907–926. [Google Scholar] [CrossRef]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The pros, cons, and many unknowns of probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef]
- Doron, S.; Snydman, D.R. Risk and safety of probiotics. Clin. Infect. Dis. 2015, 60 (Suppl. S2), S129–S134. [Google Scholar] [CrossRef]
- Veiga, P.; Suez, J.; Derrien, M.; Elinav, E. Moving from probiotics to precision probiotics. Nat. Microbiol. 2020, 5, 878–880. [Google Scholar] [CrossRef]
- Verde, L.; Dalamaga, M.; Capó, X.; Annunziata, G.; Hassapidou, M.; Docimo, A.; Savastano, S.; Colao, A.; Muscogiuri, G.; Barrea, L. The Antioxidant Potential of the Mediterranean Diet as a Predictor of Weight Loss after a Very Low-Calorie Ketogenic Diet (VLCKD) in Women with Overweight and Obesity. Antioxidants 2022, 12, 18. [Google Scholar] [CrossRef]
- Dalamaga, M.; Nikolaidou, A.; Karmaniolas, K.; His, A.; Chamberland, J.; Dionyssiou-Asteriou, A.; Mantzoros, C.S. Circulating adiponectin and leptin in relation to myelodysplastic syndrome: A case-control study. Oncology 2007, 73, 26–32. [Google Scholar] [CrossRef]
- Desli, E.; Spilioti, M.; Evangeliou, A.; Styllas, F.; Magkos, F.; Dalamaga, M. The Efficacy and Safety of Ketogenic Diets in Drug-Resistant Epilepsy in Children and Adolescents: A Systematic Review of Randomized Controlled Trials. Curr. Nutr. Rep. 2022, 11, 102–116. [Google Scholar] [CrossRef]
- Lempesis, I.G.; Liu, J.; Dalamaga, M. The catcher in the gut: Tirzepatide, a dual incretin analog for the treatment of type 2 diabetes mellitus and obesity. Metabol. Open 2022, 16, 100220. [Google Scholar] [CrossRef]
- Kounatidis, D.; Vallianou, N.G.; Tsilingiris, D.; Christodoulatos, G.S.; Geladari, E.; Stratigou, T.; Karampela, I.; Dalamaga, M. Therapeutic Potential of GLP-2 Analogs in Gastrointestinal Disorders: Current Knowledge, Nutritional Aspects, and Future Perspectives. Curr. Nutr. Rep. 2022, 11, 618–642. [Google Scholar] [CrossRef]
- Lempesis, I.G.; Tsilingiris, D.; Liu, J.; Dalamaga, M. Of mice and men: Considerations on adipose tissue physiology in animal models of obesity and human studies. Metabol. Open 2022, 15, 100208. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).