
Comparative Transcriptomic and Physiological Analyses Reveal Key Factors for Interstocks to Improve Grafted Seedling Growth in Tangor

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Morphological Changes
2.2. Changes in Photosynthetic Indexes and the Related Physiological Indexes
2.3. Changes in Endogenous Hormones
2.4. Transcriptome Analysis and Enrichment Analysis
2.5. Transcription Factors
2.6. Genes Involved in Photosynthesis–Antenna Proteins and Photosynthesis
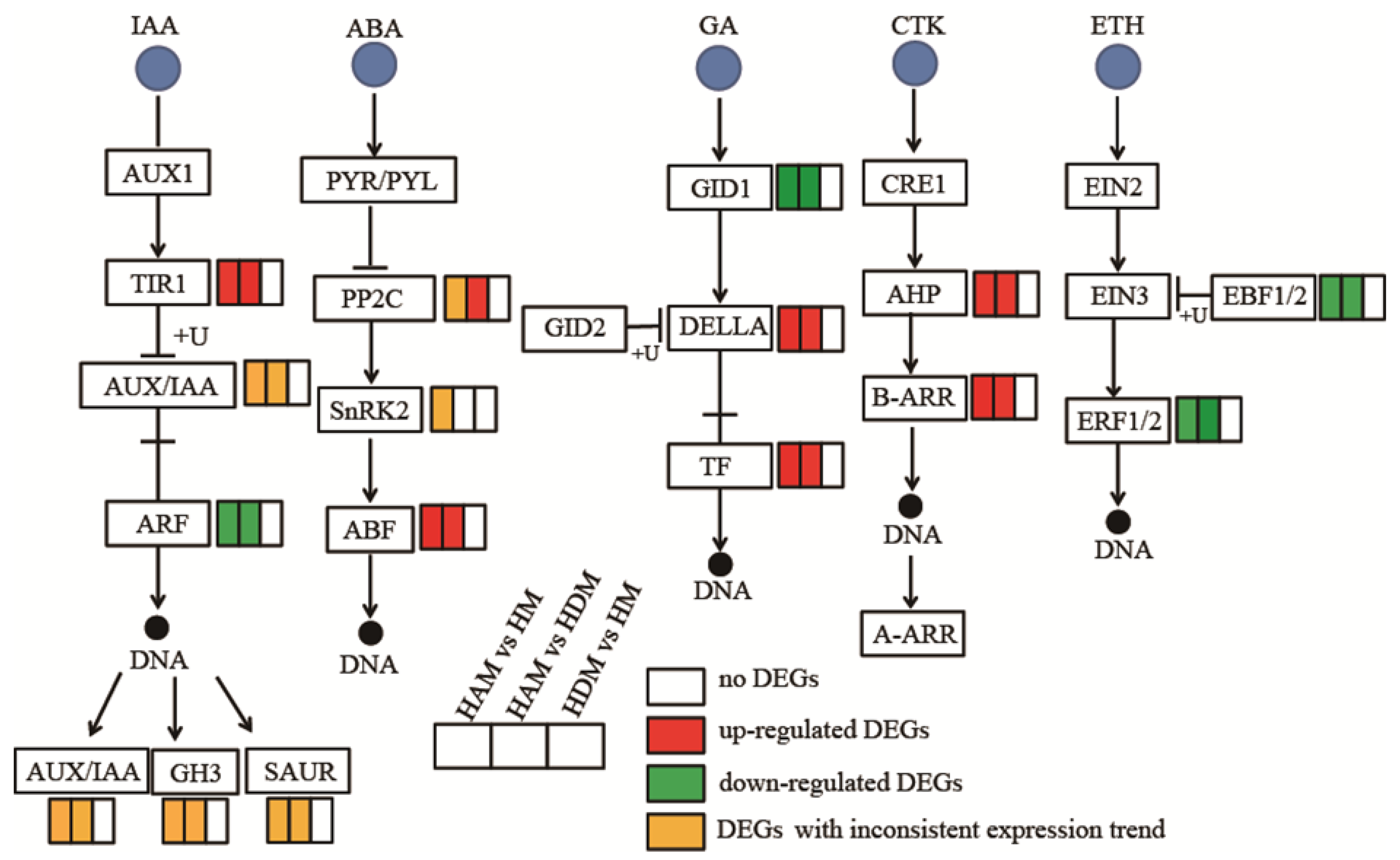
2.7. Genes Involved in Phytohormone Signaling
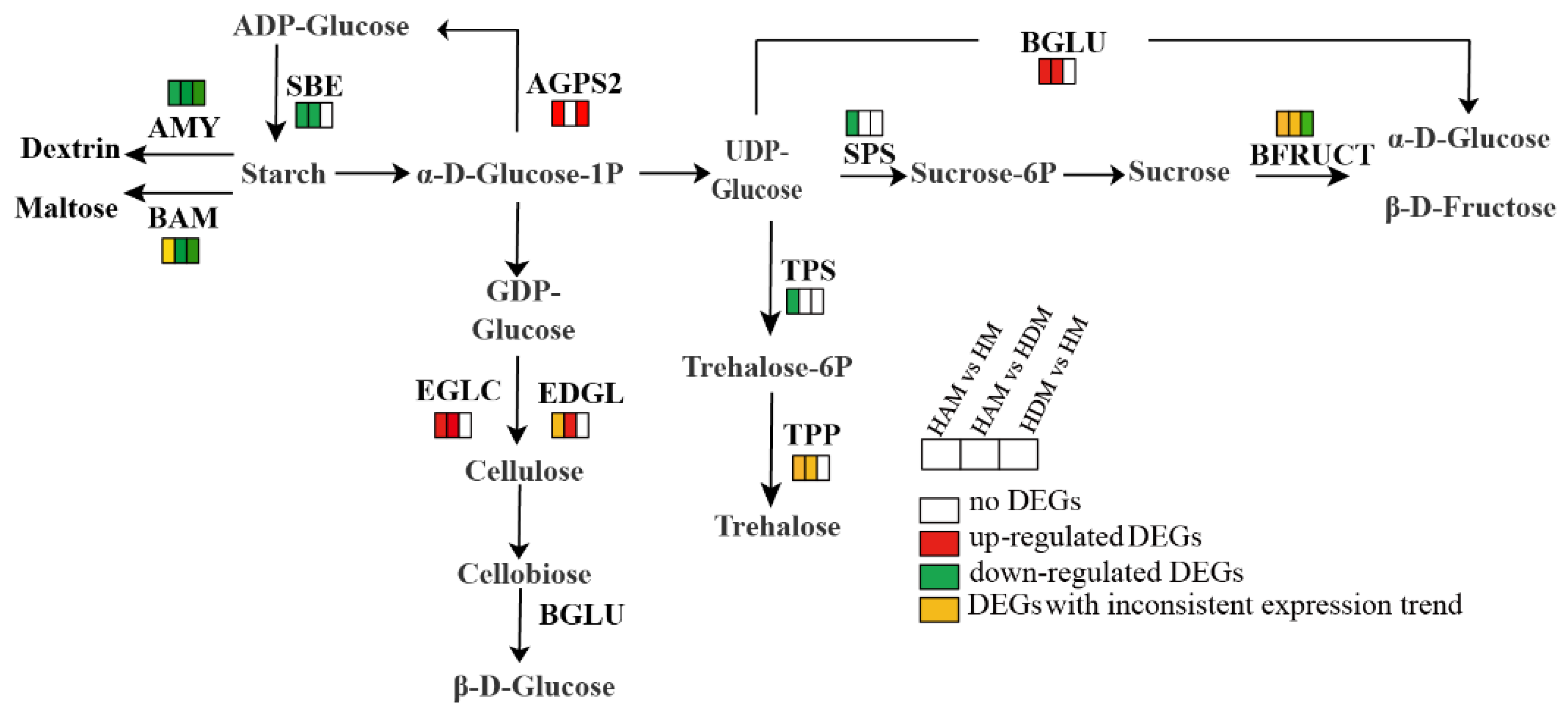
2.8. Genes Involved in Starch and Sucrose Metabolism, Amino Sugar and Nucleotide Sugar Metabolism, and Other Glycan Degradation Pathways
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Experimental Design
4.2. Analysis of Morphological Indicators and Measurement of Physiological Indexes
4.3. Anatomical Observation
4.4. Total RNA Extraction and Sequencing
4.5. Data Quality Control, Differentially Expressed Gene Analysis and Enrichment Analysis
4.6. Quantitative Real-Time PCR (qRT-PCR) Validation
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koshita, Y.; Morinaga, K.; Tsuchida, Y.; Asakura, T.; Yakushiji, H.; Azuma, A. Selection of Interstocks for Dwarfing Japanese Persimmon (Diospyros kaki Thunb.) Trees. Engei Gakkai Zasshi 2007, 76, 288–293. [Google Scholar] [CrossRef]
- Gil-Izquierdo, A.; Riquelme, M.T.; Porras, I.; Ferreres, F. Effect of the Rootstock and Interstock Grafted in Lemon Tree (Citrus limon (L.) Burm.) on the Flavonoid Content of Lemon Juice. J. Agric. Food Chem. 2004, 52, 324–331. [Google Scholar] [CrossRef]
- Shen, Y.; Zhuang, W.; Tu, X.; Gao, Z.; Qu, S. Transcriptomic analysis of interstock-induced dwarfism in Sweet Persimmon (Diospyros kaki Thunb.). Hortic. Res. 2019, 6, 1086–1102. [Google Scholar] [CrossRef] [PubMed]
- Calderón, F.; Weibel, A.M.; Trentacoste, E.R. Effects of different interstock length on vegetative growth and flowering in peach cv. Pavie Catherine. Sci. Hortic. 2021, 285, 110174. [Google Scholar] [CrossRef]
- Seleznyova, A.N.; Tustin, D.S.; Thorp, T.G. Apple Dwarfing Rootstocks and Interstocks Affect the Type of Growth Units Produced during the Annual Growth Cycle: Precocious Transition to Flowering Affects the Composition and Vigour of Annual Shoots. Ann. Bot. 2008, 101, 679–687. [Google Scholar] [CrossRef]
- Shokrollah, H.; Abdullah, T.L.; Sijam, K.; Abdullah, S. Potential use of selected citrus rootstocks and interstocks against HLB disease in Malaysia. Crop Prot. 2011, 30, 521–525. [Google Scholar] [CrossRef]
- Elias-Roman, R.D.; Calderon-Zavala, G.; Guzman-Mendoza, R.; Vallejo-Perez, M.R.; Mora-Aguilera, J.A. ‘Mondragon’: A clonal plum rootstock to enhance management of Armillaria root disease in peach orchards of Mexico. Crop Prot. 2019, 121, 89–95. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Liu, Z.Z.; Gao, H.; Zhao, Z.Y. Effect of Different Dwarfed Interstocks on the Growth Characteristics and Precocity of M. pumila ‘Changfu 2’. J. Longdong Univ. 2019, 30, 85–89. [Google Scholar]
- Almqvist, C. Interstock effects on topgraft vitality and strobili production after topgrafting in Pinus sylvestris. Can. J. For. Res. 2013, 43, 584–588. [Google Scholar] [CrossRef]
- Rebolledo-Martínez, A.; Peralta-Antonio, N.; Rebolledo-Martínez, L.; AlbertoBecerril-Román, E.; LauraRebolledo-García, R. Effect of rootstock in tree growth, dry matter, flowering, yield and quality of ‘Manila’ mango. Sci. Hortic. 2019, 251, 155–161. [Google Scholar] [CrossRef]
- Rogers, W.S.; Beakbane, A.B. Stock and Scion Relations. Annu. Rev. Plant Physiol. 2003, 8, 217–236. [Google Scholar] [CrossRef]
- He, W.; Wang, Y.; Chen, Q.; Sun, B.; Tang, H.R.; Pan, D.M.; Wang, X.R. Dissection of the Mechanism for Compatible and Incompatible Graft Combinations of Citrus grandis (L.) Osbeck (‘Hongmian Miyou’). Int. J. Mol. Sci. 2018, 19, 505. [Google Scholar] [CrossRef]
- Hong, L.L.; He, Z.; Yu, C.; Li, M.; Yi, W.; Xin, Z.Z.; Zhen, H.H. Possible roles of auxin and zeatin for initiating the dwarfing effect of M9 used as apple rootstock or interstock. Acta Physiol. Plant. 2012, 34, 235–244. [Google Scholar]
- Liu, G.D.; Wang, R.D.; Wu, L.S.; Peng, S.A.; Wang, Y.H.; Jiang, C.C. Boron distribution and mobility in navel orange grafted on citrange and trifoliate orange. Plant Soil 2012, 360, 123–133. [Google Scholar] [CrossRef]
- Shi, J.; Li, F.F.; Ma, H.; Li, Z.Y.; Xu, J.Z. Effects of Different Interstocks on Key Enzymes Activities and the Expression of Genes Related to Malic Acid Metabolism in Apple Fruit. Acta Hortic. Sin. 2016, 43, 132–140. [Google Scholar]
- Chi, C.; He, C.; Hongli, C. The content of hormone and auxin transport gene pin1 of SH40 as the interstock of apple. J. China Agric. Univ. 2012, 17, 80–84. [Google Scholar]
- Huang, L.; Grosser, J.; Gmitter, F.G., Jr.; Sims, C.A.; Wang, Y. Effects of Scion/Rootstock Combination on Flavor Quality of Orange Juice from Huanglongbing (HLB)-Affected Trees: A Two-Year Study of the Targeted Metabolomics. J. Agric. Food Chem. 2020, 68, 3286–3296. [Google Scholar] [CrossRef]
- Feng, S.; Niu, L.; Suh, J.H.; Hung, W.L.; Wang, Y. Comprehensive Metabolomics Analysis of Mandarins (Citrus reticulata) as a Tool for Variety, Rootstock, and Grove Discrimination. J. Agric. Food Chem. 2018, 66, 10317–10326. [Google Scholar] [CrossRef] [PubMed]
- Al-Khayri, J.M.; Johnson, S.M.; Dennis, V. Advances in Plant Breeding Strategies: Mandarin (Citrus reticulata Blanco) Breeding. Fruits 2018, 3, 465–533. [Google Scholar]
- da Cruz, M.A.; Neves, C.; de Carvalho, D.U.; Colombo, R.C.; Bai, J.; Yada, I.F.U.; Leite Junior, R.P.; Tazima, Z.H. Five Rootstocks for “Emperor” Mandarin Under Subtropical Climate in Southern Brazil. Front. Plant Sci. 2021, 12, 777871. [Google Scholar] [CrossRef]
- Rouphaela, Y.; Schwarzb, D.; Krumbeinb, A.; Collac, G. Impact of grafting on product quality of fruit vegetables. Sci. Hortic. 2010, 127, 172–179. [Google Scholar] [CrossRef]
- Souza, J.; Silva, E.; Filho, M.; Morillon, R.; Gesteira, A. Different adaptation strategies of two citrus scion/rootstock combinations in response to drought stress. PLoS ONE 2017, 12, e0177993. [Google Scholar]
- Dubey, A.K.; Sharma, R.M. Effect of rootstocks on tree growth, yield, quality and leaf mineral composition of lemon (Citrus limon (L.) Burm.). Sci. Hortic. 2016, 200, 131–136. [Google Scholar] [CrossRef]
- Emmanouilidou, M.G.; Kyriacou, M.C. Rootstock-modulated yield performance, fruit maturation and phytochemical quality of ‘Lane Late’ and ‘Delta’ sweet orange. Sci. Hortic. 2017, 225, 112–121. [Google Scholar] [CrossRef]
- Lastdrager, J.; Hanson, J.; Smeekens, S. Sugar signals and the control of plant growth and development. J. Exp. Bot. 2014, 65, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Amelie, R.; Clemence, H.; Khaoula, B.B.; Eric, M.; Wassim, A.; Jeremy, L.; Latifa, H.; Rachid, B.; Nathalie, L.; Sandrine, P.T. Insight into the Role of Sugars in Bud Burst Under Light in the Rose. Plant Cell Physiol. 2012, 53, 1068–1082. [Google Scholar]
- Kebrom, T.H.; Mullet, J.E. Photosynthetic Leaf Area Modulates Tiller Bud Outgrowth In Sorghum. Plant Cell Environ. 2014, 38, 1471–1478. [Google Scholar] [CrossRef]
- Foster, T.M.; Mcatee, P.A.; Waite, C.N.; Boldingh, H.L.; Mcghie, T.K. Apple dwarfing rootstocks exhibit an imbalance in carbohydrate allocation and reduced cell growth and metabolism. Hortic. Res. 2017, 4, 17009. [Google Scholar] [CrossRef]
- Zombardo, A.; Crosatti, C.; Bagnaresi, P.; Bassolino, L.; Mica, E. Transcriptomic and biochemical investigations support the role of rootstock-scion interaction in grapevine berry quality. BMC Genom. 2020, 21, 468. [Google Scholar] [CrossRef]
- He, W.; Xie, R.; Wang, Y.; Chen, Q.; Wang, H.; Yang, S.; Luo, Y.; Zhang, Y.; Tang, H.; Gmitter, F.G. Comparative transcriptomic analysis on compatible/incompatible grafts in Citrus. Hortic. Res. 2022, 9, uhab072. [Google Scholar] [CrossRef]
- Ensminger, I.; Busch, F.; Huner, N.P.A. Photostasis and cold acclimation: Sensing low temperature through photosynthesis. Physiol. Plant. 2006, 126, 28–44. [Google Scholar] [CrossRef]
- Xiao, X.; Lv, J.; Xie, J.; Feng, Z.; Calderón-Urrea, A. Transcriptome Analysis Reveals the Different Response to Toxic Stress in Rootstock Grafted and Non-Grafted Cucumber Seedlings. Int. J. Mol. Sci. 2020, 21, 774. [Google Scholar] [CrossRef]
- Kornarzyński, K.; Dziwulska-Hunek, A.; Kornarzyńska-Gregorowicz, A.; Sujak, A. Effect of Electromagnetic Stimulation of Amaranth Seeds of Different Initial Moisture on the Germination Parameters and Photosynthetic Pigments Content. Entific Rep. 2018, 8, 14023. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xiong, B.; Tan, L.; Yang, Y.; Wang, J. Effects of interstocks on growth and photosynthetic characteristics in ‘Yuanxiaochun’ Citrus seedlings. Funct. Plant Biol. 2020, 47, 977–987. [Google Scholar] [CrossRef]
- Xi, Y.; Kong, F.; Chi, Z. ROS Induce beta-Carotene Biosynthesis Caused by Changes of Photosynthesis Efficiency and Energy Metabolism in Dunaliella salina Under Stress Conditions. Front. Bioeng. Biotechnol. 2020, 8, 613768. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chen, X.; Ge, Q.; Li, B.; Tong, Y.; Zhang, A.; Li, Z.; Kuang, T.; Lu, C. Tolerance of photosynthesis to photoinhibition, high temperature and drought stress in flag leaves of wheat: A comparison between a hybridization line and its parents grown under field conditions. Plant Sci. Int. J. Exp. Plant Biol. 2006, 171, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Yang, C.; Pan, Z.; Liu, Y.; Peng, S. Boron deficiency in woody plants: Various responses and tolerance mechanisms. Front. Plant Sci. 2015, 6, 916. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, J.W.; Guo, L.X.; Liu, Y.Z.; Jin, L.F.; Hussain, S.B.; Wei, D.; Deng, Z.; Peng, S.A. Transcriptome Changes Associated with Boron Deficiency in Leaves of Two Citrus Scion-Rootstock Combinations. Front. Plant Sci. 2017, 8, 317–327. [Google Scholar] [CrossRef]
- Ma, J.; Qiu, D.; Gao, H.; Wen, H.; Wu, Y.; Pang, Y.; Wang, X.; Qin, Y. Over-expression of a γ-tocopherol methyltransferase gene in vitamin E pathway confers PEG-simulated drought tolerance in alfalfa. BMC Plant Biol. 2020, 20, 226. [Google Scholar] [CrossRef]
- Zhao, Y.; Kong, H.; Guo, Y.; Zou, Z. Light-harvesting chlorophyll a/b-binding protein-coding genes in jatropha and the comparison with castor, cassava and arabidopsis. PeerJ 2020, 8, e8465. [Google Scholar] [CrossRef]
- Guan, Z.; Mou, S.; Zhang, X.; Xu, D.; Fan, X.; Wang, Y.; Wang, D.; Ye, N. Identification and expression analysis of four light harvesting-like (Lhc) genes associated with light and desiccation stress in Ulva linza. J. Exp. Mar. Biol. Ecol. 2016, 478, 10–15. [Google Scholar] [CrossRef]
- Wientjes, E.; Roest, G.; Croce, R. From red to blue to far-red in Lhca4: How does the protein modulate the spectral properties of the pigments? BBA—Bioenerg. 2012, 1817, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Hallik, L.; Niinemets, Ü.; Kull, O. Photosynthetic acclimation to light in woody and herbaceous species: A comparison of leaf structure, pigment content and chlorophyll fluorescence characteristics measured in the field. Plant Biol. 2012, 14, 88–99. [Google Scholar] [CrossRef]
- Tang, C.; Yang, C.; Yu, H.; Tian, S.; Huang, X.; Wang, W.; Cai, P. Electromagnetic Radiation Disturbed the Photosynthesis of Microcystis aeruginosa at the Proteomics Level. Sci. Rep. 2018, 8, 479. [Google Scholar] [CrossRef] [PubMed]
- Leelavathi, S.; Bhardwaj, A.; Kumar, S.; Dass, A.; Pathak, R.; Pandey, S.S.; Tripathy, B.C.; Padmalatha, K.V.; Dhandapani, G.; Kanakachari, M. Genome-wide transcriptome and proteome analyses of tobacco psaA and psbA deletion mutants. Plant Mol. Biol. 2011, 76, 407–423. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.Y.; Gong, Q.Y.; Pei, H.; Liao, C.J.; Huang, J. Comparative Transcriptome Analysis Reveals Regulatory Networks during the Maize Ear Shank Elongation Process. Int. J. Mol. Sci. 2021, 22, 7029. [Google Scholar] [CrossRef] [PubMed]
- Rocha, F.R.; Papini-Terzi, F.S.; Nishiyama, M.Y.; Vêncio, R.; Vicentini, R.; Duarte, R.D.; Rosa, V.; Vinagre, F.; Barsalobres, C.; Medeiros, A.H. Signal transduction-related responses to phytohormones and environmental challenges in sugarcane. BMC Genom. 2007, 8, 71. [Google Scholar] [CrossRef]
- Bohn-Courseau, I. Auxin: A major regulator of organogenesis. C. R. Biol. 2010, 333, 290–296. [Google Scholar] [CrossRef]
- Burks, D.J.; Azad, R.K. Identification and Network-Enabled Characterization of Auxin Response Factor Genes in Medicago truncatula. Front. Plant Sci. 2016, 7, 1857. [Google Scholar] [CrossRef]
- Wu, B.; Sun, M.; Zhang, H.; Yang, D.; Lin, C.; Khan, I.; Wang, X.; Zhang, X.; Nie, G.; Feng, G.; et al. Transcriptome analysis revealed the regulation of gibberellin and the establishment of photosynthetic system promote rapid seed germination and early growth of seedling in pearl millet. Biotechnol. Biofuels 2021, 14, 94. [Google Scholar] [CrossRef]
- Li, K.; Yu, R.; Fan, L.M.; Wei, N.; Chen, H.; Deng, X.W. DELLA-mediated PIF degradation contributes to coordination of light and gibberellin signalling in Arabidopsis. Nat. Commun. 2016, 7, 11868. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Owens, R.A.; Sun, Q.; Song, H.; Liu, Y.; Eamens, A.L.; Feng, H.; Tian, H.; Wang, M.B.; Zhang, R. Silencing of transcription factor encoding gene StTCP23 by small RNAs derived from the virulence modulating region of potato spindle tuber viroid is associated with symptom development in potato. PLoS Pathog. 2019, 15, e1008110. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.; Urao, T.; Feng, Q.; Maruyama, K.; Kakimoto, T.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis. Proc. Natl. Acad. Sci. USA 2008, 104, 20623–20628. [Google Scholar] [CrossRef]
- Guo, S.; Sun, H.; Tian, J.; Zhang, G.; Xu, Y. Grafting Delays Watermel on Fruit Ripening by Altering Gene Expression of ABA Centric Phytohormone Signaling. Front. Plant Sci. 2021, 12, 624319. [Google Scholar] [CrossRef] [PubMed]
- Salehin, M.; Li, B.; Tang, M.; Katz, E.; Song, L.; Ecker, J.R.; Kliebenstein, D.J.; Estelle, M. Auxin-sensitive Aux/IAA proteins mediate drought tolerance in Arabidopsis by regulating glucosinolate levels. Nat. Commun. 2019, 10, 4021. [Google Scholar] [CrossRef]
- Nag, R.; Maity, M.K.; Dasgupta, M. Dual DNA binding property of ABA insensitive 3 like factors targeted to promoters responsive to ABA and auxin. Plant Mol. Biol. 2005, 59, 821–838. [Google Scholar] [CrossRef]
- Tang, X.; Hao, Y.J.; Lu, J.X.; Lu, G.; Zhang, T. Transcriptomic analysis reveals the mechanism of thermosensitive genic male sterility (TGMS) of Brassica napus under the high temperature inducement. BMC Genom. 2019, 20, 644. [Google Scholar] [CrossRef]
- Gao, Q.Y.; Yan, C.; Zhang, B.X.; Wang, Z.W.; Wang, X.J.; Zhang, Y.; University, N.A. Changes of Endogenous Hormone Content during the Growth of Raspberry Leaves. Hubei Agric. Sci. 2015, 54, 6280–6283. [Google Scholar]
- Nan, J.; Hang, T.; Peng, Z.; Yi, W.; Wu, T.; Zhang, X. Changes in IAA and ABA Levels and PpPINs Genes Expression During the Bud Dormancy Induction in Peach. Chin. Agric. Sci. Bull. 2015, 31, 100–106. [Google Scholar]
- Soto, A.; Ruiz, K.B.; Ravaglia, D.; Costa, G.; Torrigiani, P. ABA may promote or delay peach fruit ripening through modulation of ripening- and hormone-related gene expression depending on the developmental stage. Plant Physiol. Biochem. 2013, 64, 11–24. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Harshavardhan, V.T.; Govind, G.; Seiler, C.; Kohli, A. Contrapuntal role of ABA: Does it mediate stress tolerance or plant growth retardation under long-term drought stress? Gene 2012, 506, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, Q.; Chen, W.; Guo, Q.; Xia, Y.; Wang, S.; Jing, D.; Liang, G. Physiological and transcription analyses reveal the regulatory mechanism of melatonin in inducing drought resistance in loquat (Eriobotrya japonica Lindl.) seedlings. Environ. Exp. Bot. 2021, 181, 104291. [Google Scholar] [CrossRef]
- Song, L.; Yu, H.; Dong, J.; Che, X.; Dong, L. The Molecular Mechanism of Ethylene-Mediated Root Hair Development Induced by Phosphate Starvation. PLoS Genet. 2016, 12, e1006194. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Q.; Yu, C.Y.; Dong, J.G.; Hu, S.W.; Xu, A.X. Acetolactate Synthase-Inhibiting Gametocide Amidosulfuron Causes Chloroplast Destruction, Tissue Autophagy, and Elevation of Ethylene Release in Rapeseed. Front. Plant Sci. 2017, 8, 1625. [Google Scholar] [CrossRef]
- Dai, Y.; Shen, Z.; Liu, Y.; Wang, L.; Hannaway, D.; Lu, H. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environ. Exp. Bot. 2009, 65, 177–182. [Google Scholar] [CrossRef]
- Wang, X.; Peng, F.; Li, M.; Yang, L.; Li, G. Expression of a heterologous SnRK1 in tomato increases carbon assimilation, nitrogen uptake and modifies fruit development. J. Plant Physiol. 2012, 169, 1173–1182. [Google Scholar] [CrossRef]
- Si, C.C.; Liang, Q.G.; Liu, H.J.; Wang, N.; Kumar, S.; Chen, Y.L.; Zhu, G.P. Response Mechanism of Endogenous Hormones of Potential Storage Root to Phosphorus and Its Relationship With Yield and Appearance Quality of Sweetpotato. Front. Plant Sci. 2022, 13, 872422. [Google Scholar] [CrossRef]
- Yu, B.; Liu, J.; Wu, D.; Liu, Y.; Cen, W.; Wang, S.; Li, R.; Luo, J. Weighted gene coexpression network analysis-based identification of key modules and hub genes associated with drought sensitivity in rice. BMC Plant Biol. 2020, 20, 478. [Google Scholar] [CrossRef]
- Liu, L.; Shi, H.; Li, S.; Sun, M.; Zhang, R.; Wang, Y.; Ren, F. Integrated Analysis of Molybdenum Nutrition and Nitrate Metabolism in Strawberry. Front. Plant Sci. 2020, 11, 1117. [Google Scholar] [CrossRef]
- Yang, L.; Smyth, G.K.; Wei, S. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rong, Y.; Liao, L.; Li, S.; Wei, W.; Bi, X.; Sun, G.; He, S.; Wang, Z. Comparative Transcriptomic and Physiological Analyses Reveal Key Factors for Interstocks to Improve Grafted Seedling Growth in Tangor. Int. J. Mol. Sci. 2023, 24, 6533. https://doi.org/10.3390/ijms24076533
Rong Y, Liao L, Li S, Wei W, Bi X, Sun G, He S, Wang Z. Comparative Transcriptomic and Physiological Analyses Reveal Key Factors for Interstocks to Improve Grafted Seedling Growth in Tangor. International Journal of Molecular Sciences. 2023; 24(7):6533. https://doi.org/10.3390/ijms24076533
Chicago/Turabian StyleRong, Yi, Ling Liao, Sichen Li, Wen Wei, Xiaoyi Bi, Guochao Sun, Siya He, and Zhihui Wang. 2023. "Comparative Transcriptomic and Physiological Analyses Reveal Key Factors for Interstocks to Improve Grafted Seedling Growth in Tangor" International Journal of Molecular Sciences 24, no. 7: 6533. https://doi.org/10.3390/ijms24076533
APA StyleRong, Y., Liao, L., Li, S., Wei, W., Bi, X., Sun, G., He, S., & Wang, Z. (2023). Comparative Transcriptomic and Physiological Analyses Reveal Key Factors for Interstocks to Improve Grafted Seedling Growth in Tangor. International Journal of Molecular Sciences, 24(7), 6533. https://doi.org/10.3390/ijms24076533