
Tumor Microenvironment in Sporadic Vestibular Schwannoma: A Systematic, Narrative Review
 ,
,  ,
,  , ,
, , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Protocol Registration
2.2. Search Strategy
2.3. Selection Criteria
2.4. Data Extraction and Quality Assessment
3. Results and Discussion
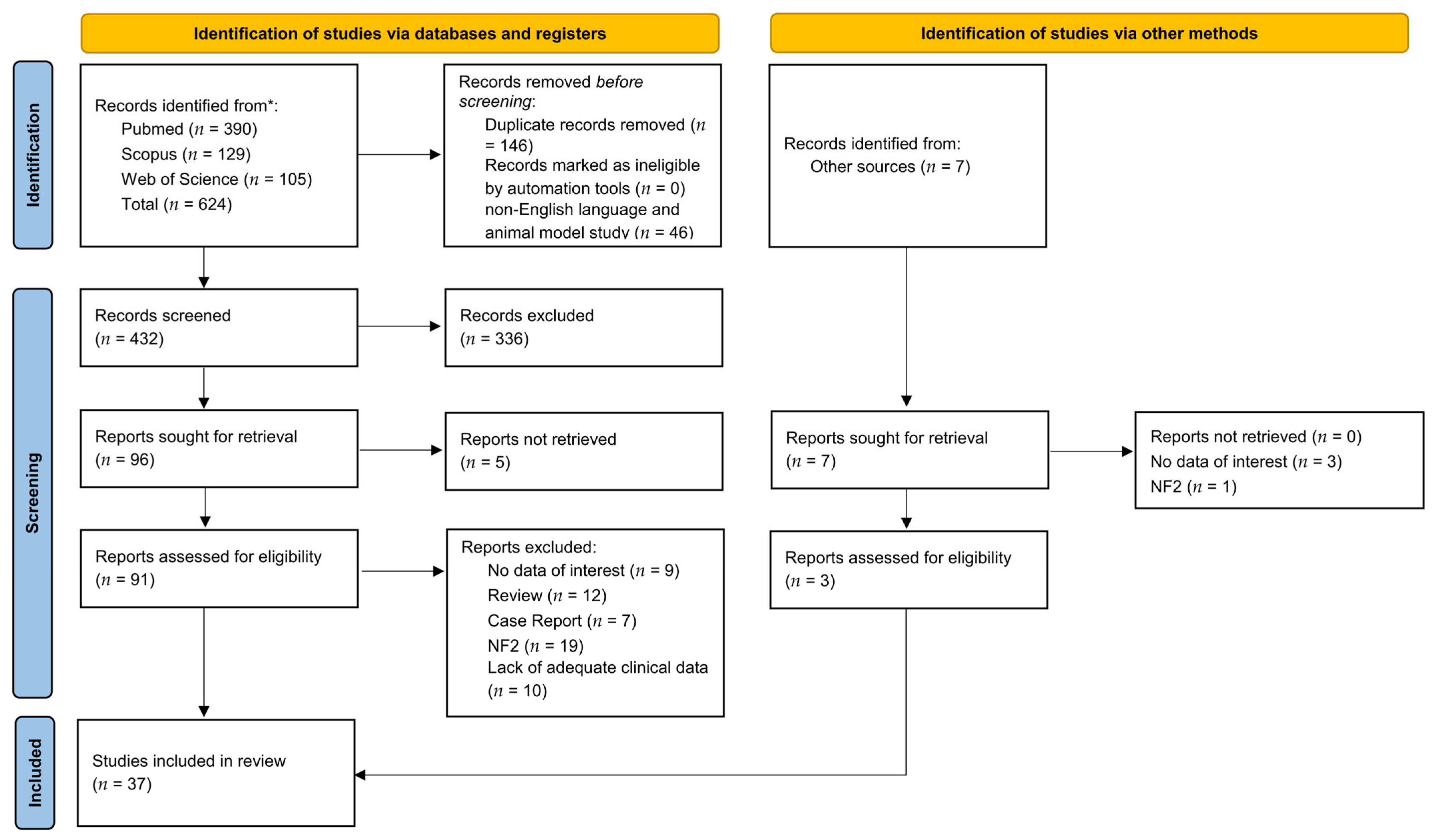
3.1. Search Results and Quality Assessment
3.2. Included Studies’ Characteristics
3.3. Angiogenesis

{kind=link}
| Author | Year | Country | Study Design | Cases (n) | Mean Age ± SD (Range) | Sex (M/F) | Tumor Size, Mean ± SD (Range) | Method of Marker Detection | Marker Studied | Key Findings |
|---|---|---|---|---|---|---|---|---|---|---|
| Brieger et al. [26] | 2003 | Germany | Ex vivo tissue study | 34 | 49 (19–72) | 20/14 | 16.5 mm (5–36) | IHC | VEGF VEGF-R1 VEGF-R2 TGF-β1 CD31 CD68 |
|
| Cayè-Thomasen et al. [28] | 2005 | Denmark | Ex vivo tissue study | 27 | 53 (35–61) | 11/16 | 1.431 cm2 (0.198–4.589) | ELISA | VEGF VEGF-R1 |
|
| Koutsimpelas et al. [38] | 2007 | Germany | Ex vivo tissue study | 17 | 51.5 ± 12.2 (28–71) | 11/6 | 424 ± 658 mm3 (36–2556) | IHC qPCR | VEGF bFGF |
|
| Koutsimpelas et al. [39] | 2012 | Germany | Ex vivo tissue study | 182 | 52 ± 10.6 (18–78) | 79/ 103 | 2.404 ± 2.329 mm3 (24–37.679) | Tissue microarray IHC | VEGF VEGFR-1 VEGFR-2 NP1 |
|
| Marioni et al. [46] | 2019 | Italy | Ex vivo tissue study | 71 | 52.8 ± 13.0 | 37/34 | 10 intra-meatal; 21 small (<1 cm); 27 medium-sized (1–2.5 cm); 13 large (>2.5 cm) | IHC | Endoglin (CD105) |
|
| Matsunaga et al. [48] | 1996 | Japan | Ex vivo tissue study | 29 | 51.38 ± 12.18 (8–66) | 13/16 | 19.03 ± 10.32 mm (5–40) | IHC | Blood vessels |
|
| Xia et al. [57] | 2020 | China | Ex vivo tissue study | 38 | NR | NR | 31.95 ± 1.74 (NR) | IHC | MMP-14 VEGF |
|
3.4. Immune Cells Infiltrates
| Author | Year | Country | Study Design | Cases (n) | Mean Age ± SD (Range) | Sex (M/F) | Tumor Size, Mean ± SD (Range) | Method of Marker Detection | Marker Studied | Key Findings |
|---|---|---|---|---|---|---|---|---|---|---|
| Amit et al. [23] | 2022 | USA | Ex vivo tissue study | 17 | 51.3 ± 11.22 | 6/11 | NR | CD4/CD8 CD20 CD68 CD1A | IF |
|
| De Vries et al. [29] | 2012 | Netherlands | Ex vivo tissue study | 67 | 49.04 ± 14.06 (15–72) | 26/41 | 24.03 ± 11.52 mm (5–50) | Histone H3 Ki-67 CD31 CD45 CD68 Hemosiderin | IHC |
|
| De Vries et al. [30] | 2019 | Netherlands | Ex vivo tissue study | 20 | 56.75 (39–81) | 4/16 | 13 ± 7.44 mL | M-CSF IL-34 | IHC |
|
| Gonçalves et al. [34] | 2021 | Germany | Ex vivo tissue study | 923 | NR | NR | 4.73 cm3 (0.04–52.14) | CD3 CD8 CD68 CD163 | IHC |
|
| Labit-Bouvier et al. [41] | 2000 | France | Ex vivo tissue study | 69 | 53 (median) (20–77) | 37/32 | 18 mm (median) (6–50) | CD34 CD45 ER, PR | IHC |
|
| Leisz et al. [43] | 2022 | Germany | Ex vivo tissue study | 74 | 53 (28–77) | 32/42 | 2.44 cm3 (0.1–18.8) | Ki-67 COX2 VEGF M-CSF GM-CSF CD163 CD68 | RT-PCR |
|
| Lewis et al. [44] | 2021 | UK | Ex vivo tissue study | 17 | 49.4 (median) (41.3–55-8) | 7/10 | 2.51 cm3 (median) (1.56–5-91) | Iba+ CD31 Fibrinogen TAM VEGF VEGFR-1 Ki-67 | IHC IF DCE-MRI |
|
| Perry et al. [53] | 2020 | USA | Ex vivo tissue study | 46 | 57 (median) (25–83) | 21/25 | Stable disease 3.3 cm (1.8–5.2) Tumor progression 2.9 cm (1.8–5.0) (median) | CD68 CD163 PD-L1 | IHC |
|
3.5. Molecular Regulators
| Author | Year | Country | Study Design | Cases (n) | Mean Age ± SD (Range) | Sex (M/F) | Tumor Size, Mean ± SD (Range) | Method of Marker Detection | Marker Studied | Key Findings |
|---|---|---|---|---|---|---|---|---|---|---|
| Ahmad et al. [21] | 2009 | Spain | Ex vivo tissue study | 34 | 49.5 (25–72) | 19/15 | 17 mm (3–40) | IHC, cell culture WB, IF, Flow citometry | p73 |
|
| Breun et al. [24] | 2018 | Germany | Ex vivo tissue study | 30 | 51 (NR) | 12/18 | 10 small tumors (T3A or smaller) 20 large tumors (T3B of larger) | IHC WB RT-qPCR | CXCR4 CXCL12 |
|
| Jabbour et al. [35] | 2016 | Australia | Ex vivo tissue study | 180 | 54 ± 13.9 (14–81) | 96/84 | NR | IHC | cyclin D1 cyclin D3 Ki-67 |
|
| Lassaletta et al. [42] | 2011 | Spain | Ex vivo tissue study | 64 | 49 (16–78) | 27/37 | 23 mm (5–55) | IHC | Cyclin D1 |
|
| Martini et al. [47] | 2017 | Italy | Ex vivo tissue study | 36 | 51.8 ± 12.4 (NR) | 17/19 | 4 intra-meatal; 8 small-sized (<1 cm); 19 medium-sized (1–2.5 cm); 5 large-sized (>2.5 cm) | IHC ceMRI | YAP TAZ AREG |
|
| Mawrin et al. [49] | 2002 | Germany | Ex vivo tissue study | 14 | 58.36 ± 5.35 (52–73) | 7/7 | NR | IHC | Fas-Fas-L Bcl-2 Bax MIB-1 |
|
| Neff et al. [52] | 2006 | USA | Ex vivo tissue study | 15 | NR | NR | NR | IHC | Cyclin D1 Cyclin D3 |
|
| Seol et al. [55] | 2005 | Korea | Ex vivo tissue study | 12 | NR | NR | NR | IHC | p53 Bax Bcl-2 Fas Fas-L Caspase-3 p27 p21 |
|
3.6. Growth Factors
| Author | Year | Country | Study Design | Cases (n) | Mean Age ± SD (Range) | Sex (M/F) | Tumor Size, Mean ± SD (Range) | Method of Marker Detection | Marker Studied | Key Findings |
|---|---|---|---|---|---|---|---|---|---|---|
| Altuna et al. [22] | 2011 | Spain | Ex vivo tissue study | 34 | 49.5 (25–72) | 19/15 | 17 mm (3–40) | IHC, cell culture and WB, IF, Colony Formation Assay | PDGF-R c-Kit |
|
| Diensthuber et al. [31] | 2004 | Germany | Ex vivo tissue study | 22 | 55.3 ± 11.8 (27–77) | 13/9 | 17 ± 6.8 mm (7–30) | IHC | TGF-β1 Glial Cell Line-Derived Neurotrophic Factor Ki-67 TβR II, GFRα-1 and Ret |
|
| Dilwali et al. [33] | 2013 | USA | Ex vivo tissue study | 16 gh 19 ph | NR | NR | NR | Cytokine Array ELISA | FGF2 |
|
| Kramer et al. [40] | 2010 | Germany | Ex vivo tissue study | 18 | 54 (20–77) | NR | NR | RT-qPCR IHC | BDNF, GDNF TGF-β1/β2 Ki-67 |
|
| Löttrich et al. [45] | 2007 | Germany | Ex vivo tissue study | 40 | NR | NR | NR | IHC qRT-PCR | TGF-β R1 TGF-β R2 |
|
| Taurone et al. [56] | 2015 | Italy | Ex vivo tissue study | 10 | NR (45–69) | 6/4 | NR | IHC | TGF-β1 IL-1β IL-6 TNF-α ICAM-1 VEGF |
|
3.7. Matrix Metalloproteinases
| Author | Year | Country | Study Design | Cases (n) | Mean Age ± SD (Range) | Sex (M/F) | Tumor Size, Mean ± SD (Range) | Method of Marker Detection | Marker Studied | Key Findings |
|---|---|---|---|---|---|---|---|---|---|---|
| Breun et al. [25] | 2020 | Germany | Ex vivo tissue study | 30 | 51 (NR) | 12/18 | 10 small tumors (T3A or smaller) 20 large tumors (T3B of larger) | IHC WB RT-qPCR | ADAM9 |
|
| Møller et al. [50] | 2010 | Denmark | Ex vivo tissue study | 34 | 53.61 ± 12.06 (25–75) | 14/20 | 1.686 ± 1.247 cm3 (0.11–4.716) | IHC ELISA | MMP-2 MMP-9 TIMP-1 |
|
| Moon et al. [51] | 2007 | South Korea | Ex vivo tissue study | 24 | 40.5 (16–71) | 12/12 | 43.8 mm (22–60) | Gelatin zymography IHC | MMP-2 |
|
| Sagers et al. [54] | 2019 | USA | Ex vivo tissue study | 80 | NR | NR | NR | IHC qRT-PCR | NLRP3 (CASP1, PYCARD, IL-18, NLRP3, NAIP, NLRC4, AIM2, IL-1β) |
|
3.8. Hormone Receptors
| Author | Year | Country | Study Design | Cases (n) | Mean Age ± SD (Range) | Sex (M/F) | Tumor Size, Mean ± SD (Range) | Method of Marker Detection | Marker Studied | Key Findings |
|---|---|---|---|---|---|---|---|---|---|---|
| Cafer et al. [27] | 2008 | Turkey | Ex vivo tissue study | 59 | 46.8 (14–75) | 27/32 | 21 small size tumors (<19 mm) 35 medium size tumors (20–39 mm) 3 large size tumors (>40 mm) | IHC | Ki-67 ER PR |
|
| Dillard et al. [32] | 2001 | USA | Ex vivo tissue study | 14 | NR | NR | 2.41 ± 1.53 cm (1–6) | IHC | EPO EPO-R |
|
| Jaiswal et al. [36] | 2016 | India | Ex vivo tissue study | 100 | 37.5 (12–77) | 63/37 | NR | IHC | ER PR |
|
| Klinken et al. [37] | 1990 | Denmark | Ex vivo tissue study | 18 | 52 (26–73) | 7/11 | NR | IHC | ER PR |
|
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roosli, C.; Linthicum, F.H.; Cureoglu, S.; Merchant, S.N. What is the site of origin of cochleovestibular schwannomas? Audiol. Neurootol. 2012, 17, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Carlson, M.L.; Link, M.J. Vestibular schwannomas. N. Engl. J. Med. 2021, 384, 1335–1348. [Google Scholar] [CrossRef] [PubMed]
- Reznitsky, M.; Petersen, M.M.B.S.; West, N.; Stangerup, S.E.; Cayé-Thomasen, P. Epidemiology of vestibular schwannomas—Prospective 40-year data from an unselected national cohort. Clin. Epidemiol. 2019, 11, 981–986. [Google Scholar] [CrossRef]
- Zanoletti, E.; Cazzador, D.; Faccioli, C.; Gallo, S.; Denaro, L.; D’Avella, D.; Martini, A.; Mazzoni, A. Multi-option therapy vs observation for small acoustic neuroma: Hearing-focused management. Acta Otorhinol. Ital. 2018, 38, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Zanoletti, E.; Concheri, S.; Tealdo, G.; Cazzador, D.; Denaro, L.; d’Avella, D.; Mazzoni, A. Early surgery and definitive cure in small sporadic vestibular schwannoma. Acta Otorhinol. Ital. 2022, 42, 481–486. [Google Scholar] [CrossRef]
- Zanoletti, E.; Mazzoni, A.; Chiumenti, F.A.; d’Avella, D.; Cazzador, D. Early Translabyrinthine surgery for small- and medium-sized vestibular schwannomas: Consecutive cohort analysis of outcomes. Otol. Neurotol. 2022, 43, 962–967. [Google Scholar] [CrossRef]
- Schwartz, M.S.; Lekovic, G.P.; Miller, M.E.; Slattery, W.H.; Wilkinson, E.P. Translabyrinthine microsurgical resection of small vestibular schwannomas. J. Neurosurg. 2018, 129, 128–136. [Google Scholar] [CrossRef]
- Carlson, M.L.; Jacob, J.T.; Pollock, B.E.; Neff, B.A.; Tombers, N.M.; Driscoll, C.L.; Link, M.J. Long-term hearing outcomes following stereotactic radiosurgery for vestibular schwannoma: Patterns of hearing loss and variables influencing audiometric decline. J. Neurosurg. 2013, 118, 579–587. [Google Scholar] [CrossRef]
- Lee, D.Y.; Lerner, D.K.; Naples, J.G.; Brant, J.A.; Bigelow, D.C.; Lee, J.Y.K.; Alonso-Basanta, M.; Ruckenstein, M.J. Vestibular schwannoma tumor size is associated with acute vestibular symptoms after gamma knife therapy. Otol. Neurotol. 2019, 40, 1088–1093. [Google Scholar] [CrossRef]
- Lerner, D.K.; Lee, D.; Naples, J.G.; Brant, J.A.; Bigelow, D.; Alonso-Basanta, M.; Ruckenstein, M.J. Factors associated with facial nerve paresis following gamma knife for vestibular schwannoma. Otol. Neurotol. 2020, 41, e83–e88. [Google Scholar] [CrossRef]
- Lu, V.M.; Ravindran, K.; Graffeo, C.S.; Perry, A.; Van Gompel, J.J.; Daniels, D.J.; Link, M.J. Efficacy and safety of bevacizumab for vestibular schwannoma in neurofibromatosis type 2: A systematic review and meta-analysis of treatment outcomes. J. Neurooncol. 2019, 144, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Goutagny, S.; Kalamarides, M. Medical treatment in neurofibromatosis type 2. Review of the literature and presentation of clinical reports. Neurochirurgie 2018, 64, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Gugel, I.; Kluwe, L.; Zipfel, J.; Teuber, C.; Tatagiba, M.; Mautner, V.F.; Schuhmann, M.U.; Grimm, F. Minimal effect of bevacizumab treatment on residual vestibular schwannomas after partial resection in young neurofibromatosis type 2 patients. Cancers 2019, 11, 1862. [Google Scholar] [CrossRef]
- Fridman, W.H.; Dieu-Nosjean, M.C.; Pagès, F.; Cremer, I.; Damotte, D.; Sautès-Fridman, C.; Galon, J. The immune microenvironment of human tumors: General significance and clinical impact. Cancer Microenviron. 2013, 6, 117–122. [Google Scholar] [CrossRef]
- Hanahan, D.; Coussens, L.M. Accessories to the crime: Functions of cells recruited to the tumor microenvironment. Cancer Cell. 2012, 21, 309–322. [Google Scholar] [CrossRef]
- Kandathil, C.K.; Dilwali, S.; Wu, C.C.; Ibrahimov, M.; McKenna, M.J.; Lee, H.; Stankovic, K.M. Aspirin intake correlates with halted growth of sporadic vestibular schwannoma in vivo. Otol. Neurotol. 2014, 35, 353–357. [Google Scholar] [CrossRef] [PubMed]
- MacKeith, S.; Wasson, J.; Baker, C.; Guilfoyle, M.; John, D.; Donnelly, N.; Mannion, R.; Jefferies, S.; Axon, P.; Tysome, J.R. Aspirin does not prevent growth of vestibular schwannomas: A case-control study. Laryngoscope 2018, 128, 2139–2144. [Google Scholar] [CrossRef] [PubMed]
- Dilwali, S.; Kao, S.Y.; Fujita, T.; Landegger, L.D.; Stankovic, K.M. Nonsteroidal anti-inflammatory medications are cytostatic against human vestibular schwannomas. Transl. Res. 2015, 166, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. PLoS Med. 2021, 18, e1003583. [Google Scholar] [CrossRef]
- NHLBI NIH. Study Quality Assessment Tools. Available online: https://www.nhlbi.nih.gov/health-topics/study-quality-assessment-tools (accessed on 15 November 2022).
- Ahmad, Z.K.; Altuna, X.; Lopez, J.P.; An, Y.; Wang-Rodriguez, J.; Juneja, V.R.; Chen, J.S.; Arandazi, M.J.; Aguilera, J.; Harris, J.P.; et al. p73 expression and function in vestibular schwannoma. Arch. Otolaryngol. Head Neck Surg. 2009, 135, 662–669. [Google Scholar] [CrossRef][Green Version]
- Altuna, X.; Lopez, J.P.; Yu, M.A.; Arandazi, M.J.; Harris, J.P.; Wang-Rodriguez, J.; An, Y.; Dobrow, R.; Doherty, J.K.; Ongkeko, W.M. Potential role of imatinib mesylate (Gleevec, STI-571) in the treatment of vestibular schwannoma. Otol. Neurotol. 2011, 32, 163–170. [Google Scholar] [CrossRef]
- Amit, M.; Xie, T.; Gleber-Netto, F.O.; Hunt, P.J.; Mehta, G.U.; Bell, D.; Silverman, D.A.; Yaman, I.; Ye, Y.; Burks, J.K.; et al. Distinct immune signature predicts progression of vestibular schwannoma and unveils a possible viral etiology. J. Exp. Clin. Cancer Res. 2022, 41, 292. [Google Scholar] [CrossRef] [PubMed]
- Breun, M.; Schwerdtfeger, A.; Martellotta, D.D.; Kessler, A.F.; Perez, J.M.; Monoranu, C.M.; Ernestus, R.I.; Matthies, C.; Löhr, M.; Hagemann, C. CXCR4: A new player in vestibular schwannoma pathogenesis. Oncotarget 2018, 9, 9940–9950. [Google Scholar] [CrossRef] [PubMed]
- Breun, M.; Schwerdtfeger, A.; Martellotta, D.D.; Kessler, A.F.; Monoranu, C.M.; Matthies, C.; Löhr, M.; Hagemann, C. ADAM9: A novel player in vestibular schwannoma pathogenesis. Oncol. Lett. 2020, 19, 1856–1864. [Google Scholar] [CrossRef] [PubMed]
- Brieger, J.; Bedavanija, A.; Lehr, H.; Maurer, J.; Mann, W. Expression of angiogenic growth factors in acoustic neurinoma. Acta Oto-Laryngol. 2003, 123, 1040–1045. [Google Scholar] [CrossRef]
- Cafer, S.; Bayramoglu, I.; Uzum, N.; Memis, L.; Uygur, K. Expression and clinical significance of Ki-67, oestrogen and progesterone receptors in acoustic neuroma. J. Laryngol. Otol. 2008, 122, 125–127. [Google Scholar] [CrossRef]
- Cayé-Thomasen, P.; Werther, K.; Nalla, A.; Bøg-Hansen, T.C.; Nielsen, H.J.; Stangerup, S.E.; Thomsen, J. VEGF and VEGF receptor-1 concentration in vestibular schwannoma homogenates correlates to tumor growth rate. Otol. Neurotol. 2005, 26, 98–101. [Google Scholar] [CrossRef]
- De Vries, W.M.; Hogendoorn, P.C.W.; Briaire-de Bruyn, I.H.; Malessy, M.J.A.; van der Mey, A.G.L. Intratumoral hemorrhage, vessel density, and the inflammatory reaction contribute to volume increase of sporadic vestibular schwannomas. Virchows Arch. 2012, 460, 629–636. [Google Scholar] [CrossRef]
- De Vries, W.M.; Briaire-de Bruijn, I.H.; van Benthem, P.P.G.; van der Mey, A.G.L.; Hogendoorn, P.C.W. M-CSF and IL-34 expression as indicators for growth in sporadic vestibular schwannoma. Virchows Arch. 2019, 474, 375–381. [Google Scholar] [CrossRef]
- Diensthuber, M.; Brandis, A.; Lenarz, T.; Stöver, T. Co-expression of transforming growth factor-beta1 and glial cell line-derived neurotrophic factor in vestibular schwannoma. Otol. Neurotol. 2004, 25, 359–365. [Google Scholar] [CrossRef]
- Dillard, D.G.; Venkatraman, G.; Cohen, C.; Delgaudio, J.; Gal, A.A.; Mattox, D.E. Immunolocalization of erythropoietin and erythropoietin receptor in vestibular schwannoma. Acta Otolaryngol. 2001, 121, 149–152. [Google Scholar] [CrossRef]
- Dilwali, S.; Lysaght, A.; Roberts, D.; Barker, F.G.; McKenna, M.J.; Stankovic, K.M. Sporadic vestibular schwannomas associated with good hearing secrete higher levels of fibroblast growth factor 2 than those associated with poor hearing irrespective of tumor size. Otol. Neurotol. 2013, 34, 748–754. [Google Scholar] [CrossRef]
- Gonçalves, V.M.; Suhm, E.M.; Ries, V.; Skardelly, M.; Tabatabai, G.; Tatagiba, M.; Schittenhelm, J.; Behling, F. Macrophage and lymphocyte infiltration is associated with volumetric tumor size but not with volumetric growth in the Tübingen schwannoma cohort. Cancers 2021, 13, 466. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, J.; Earls, P.; Biggs, N.; Gracie, G.; Fagan, P.; Bova, R. Role of cyclins D1 and D3 in vestibular schwannoma. J. Laryngol. Otol. 2016, 130, S2–S10. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Agrawal, V.; Jaiswal, A.K.; Pandey, R.; Mahapatra, A.K. Expression of estrogen and progesterone receptors in vestibular schwannomas and their clinical significance. J. Negat. Res. Biomed. 2009, 8, 9. [Google Scholar] [CrossRef]
- Klinken, L.; Thomsen, J.; Rasmussen, B.B.; Wiet, R.J.; Tos, M. Estrogen and progesterone receptors in acoustic neuromas. Arch. Otolaryngol. Head Neck Surg. 1990, 116, 202–204. [Google Scholar] [CrossRef]
- Koutsimpelas, D.; Stripf, T.; Heinrich, U.R.; Mann, W.J.; Brieger, J. Expression of vascular endothelial growth factor and basic fibroblast growth factor in sporadic vestibular schwannomas correlates to growth characteristics. Otol. Neurotol. 2007, 28, 1094–1099. [Google Scholar] [CrossRef] [PubMed]
- Koutsimpelas, D.; Bjelopavlovic, M.; Yetis, R.; Frauenknecht, K.; Adryan, B.; Schmidtmann, I.; Gouveris, H.; Fruth, K.; Heinrich, U.R.; Stauber, R.H.; et al. The VEGF/VEGF-R axis in sporadic vestibular schwannomas correlates with irradiation and disease recurrence. ORL J. Otorhinolaryngol. Relat. Spec. 2012, 74, 330–338. [Google Scholar] [CrossRef]
- Kramer, F.; Stöver, T.; Warnecke, A.; Diensthuber, M.; Lenarz, T.; Wissel, K. BDNF mRNA expression is significantly upregulated in vestibular schwannomas and correlates with proliferative activity. J. Neurooncol. 2010, 98, 31–39. [Google Scholar] [CrossRef]
- Labit-Bouvier, C.; Crebassa, B.; Bouvier, C.; Andrac-Meyer, L.; Magnan, J.; Charpin, C. Clinicopathologic growth factors in vestibular schwannomas: A morphologic and immunohistochemical study of 69 tumors. Acta Otolaryngol. 2000, 120, 950–954. [Google Scholar] [CrossRef]
- Lassaletta, L.; Del Rio, L.; Torres-Martin, M.; Rey, J.A.; Patrón, M.; Madero, R.; Roda, J.M.; Gavilan, J. Cyclin D1 expression and facial function outcome after vestibular schwannoma surgery. Otol Neurotol. 2011, 32, 136–140. [Google Scholar] [CrossRef]
- Leisz, S.; Klause, C.H.; Vital Dos Santos, T.; Haenel, P.; Scheer, M.; Simmermacher, S.; Mawrin, C.; Strauss, C.; Scheller, C.; Rampp, S. Vestibular schwannoma volume and tumor growth correlates with macrophage marker expression. Cancers 2022, 14, 4429. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.; Donofrio, C.A.; O’Leary, C.; Li, K.L.; Zhu, X.; Williams, R.; Djoukhadar, I.; Agushi, E.; Hannan, C.J.; Stapleton, E.; et al. The microenvironment in sporadic and neurofibromatosis type II-related vestibular schwannoma: The same tumor or different? A comparative imaging and neuropathology study. J. Neurosurg. 2020, 134, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Löttrich, M.; Mawrin, C.; Chamaon, K.; Kirches, E.; Dietzmann, K.; Freigang, B. Expression of transforming growth factor-beta receptor type 1 and type 2 in human sporadic vestibular Schwannoma. Pathol. Res. Pract. 2007, 203, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Marioni, G.; Blandamura, S.; Nicolè, L.; Denaro, L.; Cazzador, D.; Pavone, C.; Giacomelli, L.; Guzzardo, V.; Fassina, A.; Mazzoni, A.; et al. Endoglin-based assessment of neoangiogenesis in sporadic VIII cranial nerve schwannoma. Pathol. Res. Pract. 2019, 215, 152648. [Google Scholar] [CrossRef]
- Martini, A.; Marioni, G.; Zanoletti, E.; Cappellesso, R.; Stramare, R.; Fasanaro, E.; Faccioli, C.; Giacomelli, L.; Denaro, L.; D’Avella, D.; et al. YAP, TAZ and AREG expression in eighth cranial nerve schwannoma. Int. J. Biol. Markers 2017, 32, e319–e324. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, T.; Kanzaki, J.; Hosoda, Y. Angiogenesis from the eighth cranial nerve to vestibular schwannomas. Acta Otolaryngol. 1996, 116, 52–58. [Google Scholar] [CrossRef]
- Mawrin, C.; Kirches, E.; Dietzmann, K.; Roessner, A.; Boltze, C. Expression pattern of apoptotic markers in vestibular schwannomas. Pathol. Res. Pract. 2002, 198, 813–819. [Google Scholar] [CrossRef]
- Møller, M.N.; Werther, K.; Nalla, A.; Stangerup, S.E.; Thomsen, J.; Bøg-Hansen, T.C.; Nielsen, H.J.; Cayé-Thomasen, P. Angiogenesis in vestibular schwannomas: Expression of extracellular matrix factors MMP-2, MMP-9, and TIMP-1. Laryngoscope 2010, 120, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.S.; Jung, S.; Seo, S.K.; Jung, T.Y.; Kim, I.Y.; Ryu, H.H.; Jin, Y.H.; Jin, S.G.; Jeong, Y.I.; Kim, K.K.; et al. Cystic vestibular schwannomas: A possible role of matrix metalloproteinase-2 in cyst development and unfavorable surgical outcome. J. Neurosurg. 2007, 106, 866–871. [Google Scholar] [CrossRef]
- Neff, B.A.; Oberstien, E.; Lorenz, M.; Chaudhury, A.R.; Welling, D.B.; Chang, L.S. Cyclin D(1) and D(3) expression in vestibular schwannomas. Laryngoscope 2006, 116, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Perry, A.; Graffeo, C.S.; Carlstrom, L.P.; Raghunathan, A.; Driscoll, C.L.W.; Neff, B.A.; Carlson, M.L.; Parney, I.F.; Link, M.J.; Van Gompel, J.J. Predominance of M1 subtype among tumor-associated macrophages in phenotypically aggressive sporadic vestibular schwannoma. J. Neurosurg. 2019, 133, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Sagers, J.E.; Sahin, M.I.; Moon, I.; Ahmed, S.G.; Stemmer-Rachamimov, A.; Brenner, G.J.; Stankovic, K.M. NLRP3 inflammasome activation in human vestibular schwannoma: Implications for tumor-induced hearing loss. Hear. Res. 2019, 381, 107770. [Google Scholar] [CrossRef] [PubMed]
- Seol, H.J.; Jung, H.W.; Park, S.H.; Hwang, S.K.; Kim, D.G.; Paek, S.H.; Chung, Y.S.; Sub Lee, C. Aggressive vestibular schwannomas showing postoperative rapid growth—Their association with decreased p27 expression. J. Neurooncol. 2005, 75, 203–207. [Google Scholar] [CrossRef]
- Taurone, S.; Bianchi, E.; Attanasio, G.; di Gioia, C.; Ierinó, R.; Carubbi, C.; Galli, D.; Pastore, F.S.; Giangaspero, F.; Filipo, R.; et al. Immunohistochemical profile of cytokines and growth factors expressed in vestibular schwannoma and in normal vestibular nerve tissue. Mol. Med. Rep. 2015, 12, 737–745. [Google Scholar] [CrossRef]
- Xia, L.; Yang, S.; Wang, C.; Yu, E.; Zhang, H.; Zhang, Y.; Ruan, L.; Shi, L.; Ni, J.; Luo, J.; et al. Immunohistochemical profiles of matrix metalloproteinases and vascular endothelial growth factor overexpression in the Antoni B area of vestibular schwannomas. World Neurosurg. 2020, 144, e72–e79. [Google Scholar] [CrossRef]
- Dzobo, K.; Senthebane, D.A.; Dandara, C. The tumor microenvironment in tumorigenesis and therapy resistance revisited. Cancers 2023, 15, 376. [Google Scholar] [CrossRef]
- Alessandrini, L.; Ferrari, M.; Taboni, S.; Sbaraglia, M.; Franz, L.; Saccardo, T.; del Forno, B.M.; Agugiaro, F.; Frigo, A.C.; Dei Tos, A.P.; et al. Tumor-stroma ratio, neoangiogenesis and prognosis in laryngeal carcinoma. A pilot study on preoperative biopsies and matched surgical specimens. Oral Oncol. 2022, 132, 105982. [Google Scholar] [CrossRef]
- Folkman, J. Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [CrossRef]
- Folkman, J.; Merler, E.; Abernathy, C.; Williams, G. Isolation of a tumor factor responsible for angiogenesis. J. Exp. Med. 1971, 133, 275–288. [Google Scholar] [CrossRef]
- Marioni, G.; Marino, F.; Blandamura, S.; D’Alessandro, E.; Giacomelli, L.; Guzzardo, V.; Lionello, M.; De Filippis, C.; Staffieri, A. Neoangiogenesis in laryngeal carcinoma: Angiogenin and CD105 expression is related to carcinoma recurrence rate and disease-free survival. Histopathology 2010, 57, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Folkman, J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell 1996, 9, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, T.; Valter, M.M.; Wolf, H.K.; von Deimling, A.; Huang, H.J.; Cavenee, W.K.; Wiestler, O.D. Expression and distribution of vascular endothelial growth factor protein in human brain tumors. Acta Neuropathol. 1997, 93, 109–117. [Google Scholar] [CrossRef]
- Nishikawa, R.; Cheng, S.Y.; Nagashima, R.; Huang, H.J.; Cavenee, W.K.; Matsutani, M. Expression of vascular endothelial growth factor in human brain tumors. Acta Neuropathol. 1998, 96, 453–462. [Google Scholar] [CrossRef]
- Schratzberger, P.; Schratzberger, G.; Silver, M.; Curry, C.; Kearney, M.; Magner, M.; Alroy, J.; Adelman, L.S.; Weinberg, D.H.; Ropper, A.H.; et al. Favorable effect of VEGF gene transfer on ischemic peripheral neuropathy. Nat. Med. 2000, 6, 405–413. [Google Scholar] [CrossRef]
- Charabi, S. Acoustic neuroma/vestibular schwannoma in vivo and in vitro growth models. A clinical and experimental study. Acta Otolaryngol. 1997, 530, 1–27. [Google Scholar]
- Saito, K.; Kato, M.; Susaki, N.; Nagatani, T.; Nagasaka, T.; Yoshida, J. Expression of Ki-67 antigen and vascular endothelial growth factor in sporadic and neurofibromatosis type 2-associated schwannomas. Clin. Neuropathol. 2003, 22, 30–34. [Google Scholar] [PubMed]
- Caye-Thomasen, P.; Baandrup, L.; Jacobsen, G.K.; Thomsen, J.; Stangerup, S.E. Immunohistochemical demonstration of VEGF in vestibular schwannomas correlates to tumor growth rate. Laryngoscope 2003, 113, 2129–2134. [Google Scholar] [CrossRef] [PubMed]
- Marioni, G.; Gaio, E.; Giacomelli, L.; Marchese-Ragona, R.; Staffieri, C.; Staffieri, A.; Marino, F. Endoglin (CD105) expression in head and neck basaloid squamous cell carcinoma. Acta Otolaryngol. 2005, 125, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Archibald, D.J.; Neff, B.A.; Voss, S.G.; Splinter, P.L.; Driscoll, C.L.; Link, M.J.; Dong, H.; Kwon, E.D. B7-H1 expression in vestibular schwannomas. Otol. Neurotol. 2010, 31, 991–997. [Google Scholar] [CrossRef]
- Solinas, G.; Germano, G.; Mantovani, A.; Allavena, P. Tumor-associated macrophages (TAM) as major players of the cancer-related inflammation. J. Leuk. Biol. 2009, 86, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Lahmar, Q.; Keirsse, J.; Laoui, D.; Movahedi, K.; van Overmeire, E.; van Ginderachter, J.A. Tissue-resident versus monocyte-derived macrophages in the tumor microenvironment. Biochim. Biophys. Acta. 2016, 1865, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Kissil, J.L.; Johnson, K.C.; Eckman, M.S.; Jacks, T. Merlin phosphorylation by p21-activated kinase 2 and effects of phosphorylation on merlin localization. J. Biol. Chem. 2002, 277, 10394–10399. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Lopez-Lago, M.; Giancotti, F.G. Merlin/NF-2 mediates contact inhibition of growth by suppressing recruitment of Rac to the plasma membrane. J. Cell Biol. 2005, 171, 361–371. [Google Scholar] [CrossRef]
- Meng, Z.; Moroishi, T.; Guan, K.L. Mechanisms of Hippo pathway regulation. Genes Dev. 2016, 30, 1–17. [Google Scholar] [CrossRef]
- Zhao, F.; Yang, Z.; Chen, Y.; Zhou, Q.; Zhang, J.; Liu, J.; Wang, B.; He, Q.; Zhang, L.; Yu, Y.; et al. Deregulation of the Hippo pathway promotes tumor cell proliferation through YAP activity in human sporadic vestibular schwannoma. World Neurosurg. 2018, 117, e269–e279. [Google Scholar] [CrossRef]
- Hilton, D.A.; Hanemann, C.O. Schwannomas and their pathogenesis. Brain Pathol. 2014, 24, 205–220. [Google Scholar] [CrossRef]
- Boerboom, A.; Dion, V.; Chariot, A.; Franzen, R. Molecular mechanisms involved in schwann cell plasticity. Front. Mol. Neurosci. 2017, 10, 38. [Google Scholar] [CrossRef]
- Rundhaug, J.E. Matrix metalloproteinases and angiogenesis. J. Cell Mol. Med. 2005, 9, 267–285. [Google Scholar] [CrossRef]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; La Rosa, C.C.-D.; Ramirez-Acuña, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The roles of matrix metalloproteinases and their inhibitors in human diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef]
- Weskamp, G.; Krätzschmar, J.; Reid, M.S.; Blobel, C.P. MDC9, a widely expressed cellular disintegrin containing cytoplasmic SH3 ligand domains. J. Cell Biol. 1996, 132, 717–726. [Google Scholar] [CrossRef]
- Mazzocca, A.; Coppari, R.; de Franco, R.; Cho, J.Y.; Libermann, T.A.; Pinzani, M.; Toker, A. A secreted form of ADAM9 promotes carcinoma invasion through tumor-stromal interactions. Cancer Res. 2005, 65, 4728–4738. [Google Scholar] [CrossRef]
- Fan, X.; Wang, Y.; Zhang, C.; Liu, L.; Yang, S.; Wang, Y.; Lu, X.; Qian, Z.; Fang, S.; Qiao, H.; et al. ADAM9 expression is associated with glioma tumor grade and histological type, and acts as a prognostic factor in lower-grade gliomas. Int. J. Mol. Sci. 2016, 17, 1276. [Google Scholar] [CrossRef]
- Oria, V.O.; Lopatta, P.; Schmitz, T.; Preca, B.T.; Nyström, A.; Conrad, C.; Bartsch, J.W.; Kulemann, B.; Hoeppner, J.; Maurer, J.; et al. ADAM9 contributes to vascular invasion in pancreatic ductal adenocarcinoma. Mol. Oncol. 2019, 13, 456–479. [Google Scholar] [CrossRef] [PubMed]
- Dilwali, S.; Landegger, L.D.; Soares, V.Y.; Deschler, D.G.; Stankovic, K.M. Secreted factors from human vestibular schwannomas can cause cochlear damage. Sci. Rep. 2015, 5, 18599. [Google Scholar] [CrossRef] [PubMed]
- Baroja-Mazo, A.; Martín-Sánchez, F.; Gomez, A.I.; Martínez, C.M.; Amores-Iniesta, J.; Compan, V.; Barberà-Cremades, M.; Yagüe, J.; Ruiz-Ortiz, E.; Antón, J.; et al. The NLRP3 inflammasome is released as a particulate danger signal that amplifies the inflammatory response. Nat. Immunol. 2014, 15, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Boyle, S.T. RISING STARS: Hormonal regulation of the breast cancer microenvironment. J. Mol. Endocrinol. 2023, 70, e220174. [Google Scholar] [CrossRef]
- Kasantikul, V.; Brown, W.J. Estrogen receptors in acoustic neurilemmomas. Surg. Neurol. 1981, 15, 105–109. [Google Scholar] [CrossRef]
- Whittle, I.; Hawkins, R.A.; Miller, J.D. Sex hormone receptors in intracranial tumors and normal brain. Eur. J. Surg. Oncol. 1987, 13, 303–307. [Google Scholar]
- Allen, J.; Eldridge, R.; Koerber, T. Acoustic neuroma in the last months of pregnancy. Am. J. Obstet. Gynecol. 1974, 119, 516–520. [Google Scholar] [CrossRef]
- Monsell, E.W.; Wiet, R.J. Estrogen and progesterone binding in acoustic neuroma tissue. Otolaryngol. Head Neck Surg. 1990, 103, 377–379. [Google Scholar] [CrossRef]
- Beatty, C.W.; Scheithauer, B.W.; Katzmann, J.A.; Roche, P.C.; Kjeldahl, K.S.; Ebsersold, M.J. Acoustic schwannomas in pregnancy: A DNA flow cytometric, steroid hormone receptor and proliferation marker study. Laryngoscope 1995, 105, 693–700. [Google Scholar] [CrossRef]
- Carroll, R.S.; Zhang, J.P.; Black, P.M.L. Hormone receptors in vestibular schwannomas. Acta Neurochir. 1997, 139, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Curley, J.W.; Ramsden, R.T.; Howell, A.; Healy, K.; Lye, R.H. Oestrogen and progesterone receptors in acoustic neuroma. J. Laryngol. Otol. 1990, 104, 865–867. [Google Scholar] [CrossRef] [PubMed]
- Dalgorf, D.M.; Rowsell, C.; Bilbao, J.M.; Chen, J.M. Immunohistochemical investigation of hormonal receptors and vascular endothelial growth factor concentration in vestibular schwannoma. Skull Base 2008, 18, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Juul, S.E.; Yachnis, A.T.; Rojiani, A.M.; Christensen, R.D. Immunohistochemical localization of erythropoietin and its receptor in the developing human brain. Ped. Dev. Path. 1999, 2, 148–158. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cazzador, D.; Astolfi, L.; Daloiso, A.; Tealdo, G.; Simoni, E.; Mazzoni, A.; Zanoletti, E.; Marioni, G. Tumor Microenvironment in Sporadic Vestibular Schwannoma: A Systematic, Narrative Review. Int. J. Mol. Sci. 2023, 24, 6522. https://doi.org/10.3390/ijms24076522
Cazzador D, Astolfi L, Daloiso A, Tealdo G, Simoni E, Mazzoni A, Zanoletti E, Marioni G. Tumor Microenvironment in Sporadic Vestibular Schwannoma: A Systematic, Narrative Review. International Journal of Molecular Sciences. 2023; 24(7):6522. https://doi.org/10.3390/ijms24076522
Chicago/Turabian StyleCazzador, Diego, Laura Astolfi, Antonio Daloiso, Giulia Tealdo, Edi Simoni, Antonio Mazzoni, Elisabetta Zanoletti, and Gino Marioni. 2023. "Tumor Microenvironment in Sporadic Vestibular Schwannoma: A Systematic, Narrative Review" International Journal of Molecular Sciences 24, no. 7: 6522. https://doi.org/10.3390/ijms24076522
APA StyleCazzador, D., Astolfi, L., Daloiso, A., Tealdo, G., Simoni, E., Mazzoni, A., Zanoletti, E., & Marioni, G. (2023). Tumor Microenvironment in Sporadic Vestibular Schwannoma: A Systematic, Narrative Review. International Journal of Molecular Sciences, 24(7), 6522. https://doi.org/10.3390/ijms24076522