

The Oligomerization Domains of the APC Protein Mediate Liquid-Liquid Phase Separation That Is Phosphorylation Controlled


Abstract
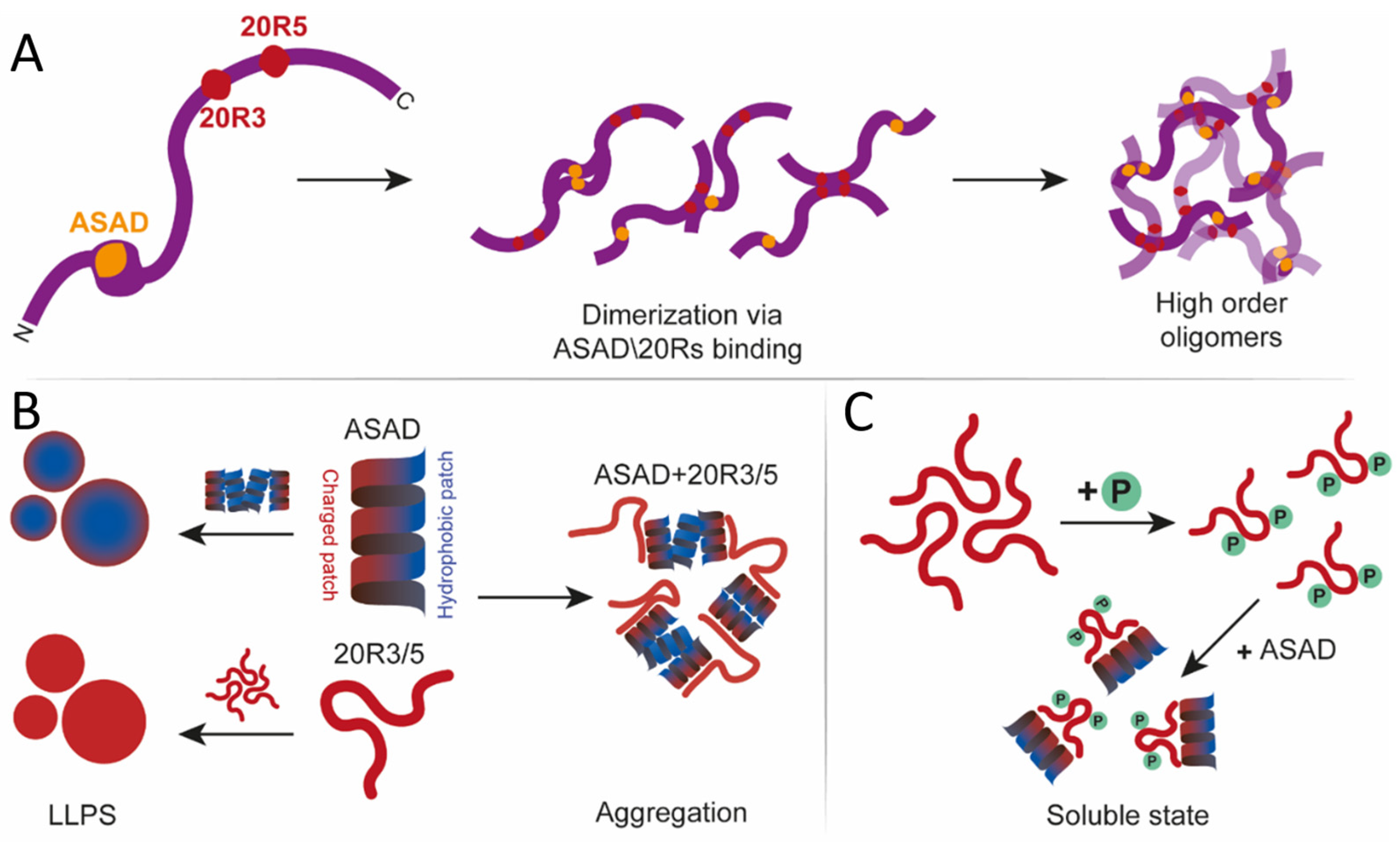
1. Introduction
2. Results
2.1. The ASAD Binds Itself and Forms Droplets via Hydrophobic Interactions
2.2. 20R3 and 20R5 form Droplets in an Ionic-Strength-Dependent Manner
2.3. LLPS of the 20R Peptides Is Regulated by Phosphorylation and Is Kinase Dependent
2.4. The ASAD and 20R Peptides Do Not form Droplets When Co-Incubated
2.5. The SLSXL Repeat within the 20R Peptides Binds the ASAD and Is Responsible for Droplet Formation of 20R3 and 20R5
3. Discussion
3.1. A peptide Approach Reveals New Oligomerization Domains in APC
3.2. Droplet Formation of APC Is Mediated by Different Domains and Mechanisms
3.3. The Role of the SLSSL Residues in Mediating Droplet Formation of the 20R Domain
3.4. Phosphorylation Regulates Droplet Formation by the 20R Domains of APC
4. Materials and Methods
4.1. Peptide Synthesis, Labeling, Stapling, and Purification
4.2. Peptide Array
4.3. DIC Microscopy
4.4. Circular Dichroism
4.5. Contact Angle Measurements
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uversky, V.N. Unusual biophysics of intrinsically disordered proteins. Biochim. Biophys. Acta 2013, 1834, 932–951. [Google Scholar] [CrossRef] [PubMed]
- Dyson, H.J.; Wright, P.E. Intrinsically unstructured proteins and their functions. Nat. Rev. Mol. Cell Biol. 2005, 6, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Forman-Kay, J.D.; Mittag, T. From sequence and forces to structure, function, and evolution of intrinsically disordered proteins. Structure 2013, 21, 1492–1499. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, A.; Dogra, P.; Maity, S.; Mukhopadhyay, S. Liquid-Liquid Phase Separation Is Driven by Large-Scale Conformational Unwinding and Fluctuations of Intrinsically Disordered Protein Molecules. J. Phys. Chem. Lett. 2019, 10, 3929–3936. [Google Scholar] [CrossRef]
- Uversky, V.N. Intrinsically disordered proteins in overcrowded milieu: Membrane-less organelles, phase separation, and intrinsic disorder. Curr. Opin. Struct. Biol. 2017, 44, 18–30. [Google Scholar] [CrossRef]
- Wei, M.T.; Elbaum-Garfinkle, S.; Holehouse, A.S.; Chen, C.C.; Feric, M.; Arnold, C.B.; Priestley, R.D.; Pappu, R.V.; Brangwynne, C.P. Phase behaviour of disordered proteins underlying low density and high permeability of liquid organelles. Nat. Chem. 2017, 9, 1118–1125. [Google Scholar] [CrossRef]
- Elbaum-Garfinkle, S.; Kim, Y.; Szczepaniak, K.; Chen, C.C.H.; Eckmann, C.R.; Myong, S.; Brangwynne, C.P. The disordered P granule protein LAF-1 drives phase separation into droplets with tunable viscosity and dynamics. Proc. Natl. Acad. Sci. USA 2015, 112, 7189–7194. [Google Scholar] [CrossRef]
- Drobot, B.; Iglesias-Artola, J.M.; Le Vay, K.; Mayr, V.; Kar, M.; Kreysing, M.; Mutschler, H.; Tang, T.D. Compartmentalised RNA catalysis in membrane-free coacervate protocells. Nat. Commun. 2018, 9, 3643. [Google Scholar] [CrossRef]
- Poudyal, R.R.; Guth-Metzler, R.M.; Veenis, A.J.; Frankel, E.A.; Keating, C.D.; Bevilacqua, P.C. Template-directed RNA polymerization and enhanced ribozyme catalysis inside membraneless compartments formed by coacervates. Nat. Commun. 2019, 10, 490. [Google Scholar] [CrossRef]
- Nakashima, K.K.; Baaij, J.F.; Spruijt, E. Reversible generation of coacervate droplets in an enzymatic network. Soft Matter 2018, 14, 361–367. [Google Scholar] [CrossRef]
- Protter, D.S.W.; Parker, R. Principles and Properties of Stress Granules. Trends Cell Biol. 2016, 26, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular condensates: Organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Oliva, R.; Mukherjee, S.K.; Ostermeier, L.; Pazurek, L.A.; Kriegler, S.; Bader, V.; Prumbaum, D.; Raunser, S.; Winklhofer, K.F.; Tatzelt, J.; et al. Remodeling of the Fibrillation Pathway of alpha-Synuclein by Interaction with Antimicrobial Peptide LL-III. Chemistry 2021, 27, 11845–11851. [Google Scholar] [CrossRef]
- Savastano, A.; Flores, D.; Kadavath, H.; Biernat, J.; Mandelkow, E.; Zweckstetter, M. Disease-Associated Tau Phosphorylation Hinders Tubulin Assembly within Tau Condensates. Angew. Chem. Int. Ed. 2021, 60, 726–730. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, S.; Gu, J.; Tong, Y.; Li, Y.; Gui, X.; Long, H.; Wang, C.; Zhao, C.; Lu, J.; et al. Hsp27 chaperones FUS phase separation under the modulation of stress-induced phosphorylation. Nat. Struct. Mol. Biol. 2020, 27, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Nelson, S.; Näthke, I.S. Interactions and functions of the adenomatous polyposis coli (APC) protein at a glance. J. Cell Sci. 2013, 15, 873–878. [Google Scholar] [CrossRef]
- Rubinfeld, B.; Albert, I.; Porfiri, E.; Fiol, C.; Munemitsu, S.; Polakis, P. Binding of GSK3β to the APC-β-catenin complex and regulation of complex assembly. Science 1996, 17, 1023–1026. [Google Scholar] [CrossRef]
- Fodde, R. The APC gene in colorectal cancer. Eur. J. Cancer 2002, 38, 867–871. [Google Scholar] [CrossRef]
- Minde, D.P.; Anvarian, Z.; Rüdiger, S.G.D.; Maurice, M.M. Messing up disorder: How do missense mutations in the tumor suppressor protein APC lead to cancer? Mol. Cancer 2011, 10, 101. [Google Scholar] [CrossRef]
- Jiang, H.; Deng, R.; Yang, X.; Shang, J.; Lu, S.; Zhao, Y.; Song, K.; Liu, X.; Zhang, Q.; Chen, Y.; et al. Peptidomimetic inhibitors of APC-Asef interaction block colorectal cancer migration. Nat. Chem. Biol. 2017, 13, 994–1001. [Google Scholar] [CrossRef]
- Day, C.L.; Alber, T. Crystal structure of the amino-terminal coiled-coil domain of the APC tumor suppressor. J. Mol. Biol. 2000, 301, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kroboth, K.; Newton, I.P.; Näthke, I.S. Novel self-association of the APC molecule affects APC clusters and cell migration. J. Cell Sci. 2008, 11, 1916–1925. [Google Scholar] [CrossRef]
- McCartney, B.M.; Nathke, I.S. Cell regulation by the Apc protein Apc as master regulator of epithelia. Curr. Opin. Cell Biol. 2008, 20, 186–193. [Google Scholar] [CrossRef]
- Fearnhead, N.S. The ABC of APC. Hum. Mol. Genet. 2001, 10, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.O. Regulation of the APC and the exit from mitosis. Nat. Cell Biol. 1999, 1, E47–E53. [Google Scholar] [CrossRef] [PubMed]
- Peifer, M.; Polakis, P. Wnt signaling in oncogenesis and embryogenesis—A look outside the nucleus. Science 2000, 287, 1606–1609. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.D.; Nusse, R. Wnt signaling: Multiple pathways, multiple receptors, and multiple transcription factors. J. Biol. Chem. 2006, 281, 22429–22433. [Google Scholar] [CrossRef] [PubMed]
- Kunttas-Tatli, E.; Roberts, D.M.; McCartney, B.M. Self-association of the APC tumor suppressor is required for the assembly, stability, and activity of the Wnt signaling destruction complex. Mol. Biol. Cell 2014, 25, 3424–3436. [Google Scholar] [CrossRef] [PubMed]
- Zumbrunn, J.; Kinoshita, K.; Hyman, A.A.; Näthke, I.S. Binding of the adenomatous polyposis coli protein to microtubules increases microtubule stability and is regulated by GSK3β phosphorylation. Curr. Biol. 2001, 25, 44–49. [Google Scholar] [CrossRef]
- Ha, N.C.; Tonozuka, T.; Stamos, J.L.; Choi, H.J.; Weis, W.I. Mechanism of phosphorylation-dependent binding of APC to β-catenin and its role in β-catenin degradation. Mol. Cell 2004, 15, 511–521. [Google Scholar] [CrossRef]
- Xing, Y.; Clements, W.K.; Kimelman, D.; Xu, W. Crystal structure of a beta-catenin/axin complex suggests a mechanism for the beta-catenin destruction complex. Genes Dev. 2003, 17, 2753–2764. [Google Scholar] [CrossRef]
- Xing, Y.; Clements, W.K.; Le Trong, I.; Hinds, T.R.; Stenkamp, R.; Kimelman, D.; Xu, W. Crystal structure of a β-catenin/APC complex reveals a critical role for APC phosphorylation in APC function. Mol. Cell 2004, 15, 523–533. [Google Scholar] [CrossRef]
- Liu, J.; Xing, Y.; Hinds, T.R.; Zheng, J.; Xu, W. The Third 20 Amino Acid Repeat Is the Tightest Binding Site of APC for β-Catenin. J. Mol. Biol. 2006, 360, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Mimori-Kiyosue, Y.; Shiina, N.; Tsukita, S. Adenomatous polyposis coli (APC) protein moves along microtubules and concentrates at their growing ends in epithelial cells. J. Cell Biol. 2000, 148, 505–518. [Google Scholar] [CrossRef]
- Serre, L.; Stoppin-Mellet, V.; Arnal, I. Adenomatous Polyposis Coli as a Scaffold for Microtubule End-Binding Proteins. J. Mol. Biol. 2019, 431, 1993–2005. [Google Scholar] [CrossRef]
- Mimori-Kiyosue, Y.; Shiina, N.; Tsukita, S. The dynamic behavior of the APC-binding protein EB1 on the distal ends of microtubules. Curr. Biol. 2000, 10, 865–868. [Google Scholar] [CrossRef]
- Efimova, N.; Yang, C.; Chia, J.X.; Li, N.; Lengner, C.J.; Neufeld, K.L.; Svitkina, T.M. Branched actin networks are assembled on microtubules by adenomatous polyposis coli for targeted membrane protrusion. J. Cell Biol. 2020, 219, e202003091. [Google Scholar] [CrossRef]
- Fang, X.; Svitkina, T.M. Adenomatous Polyposis Coli (APC) in cell migration. Eur. J. Cell Biol. 2022, 101, 151228. [Google Scholar] [CrossRef] [PubMed]
- Pronobis, M.I.; Rusan, N.M.; Peifer, M. A novel GSK3-regulated APC:Axin interaction regulates Wnt signaling by driving a catalytic cycle of efficient betacatenin destruction. Elife 2015, 4, e08022. [Google Scholar] [CrossRef] [PubMed]
- Li, T.M.; Ren, J.; Husmann, D.; Coan, J.P.; Gozani, O.; Chua, K.F. Multivalent tumor suppressor adenomatous polyposis coli promotes Axin biomolecular condensate formation and efficient beta-catenin degradation. Sci. Rep. 2020, 10, 17425. [Google Scholar] [CrossRef] [PubMed]
- Huber, A.H. The Structure of the b-Catenin/E-Cadherin Complex and the Molecular Basis of Diverse Ligand Recognition by b-Catenin. Cell Press 2001, 105, 391–402. [Google Scholar] [CrossRef]
- Mitrea, D.M.; Chandra, B.; Ferrolino, M.C.; Gibbs, E.B.; Tolbert, M.; White, M.R.; Kriwacki, R.W. Methods for Physical Characterization of Phase-Separated Bodies and Membrane-less Organelles. J. Mol. Biol. 2018, 430, 4773–4805. [Google Scholar] [CrossRef] [PubMed]
- Lamiable, A.; Thevenet, P.; Rey, J.; Vavrusa, M.; Derreumaux, P.; Tuffery, P. PEP-FOLD3: Faster de novo structure prediction for linear peptides in solution and in complex. Nucleic Acids Res. 2016, 44, W449–W454. [Google Scholar] [CrossRef] [PubMed]
- Näthke, I.; Adams, C.L.; Polakis, P.; Sellin, J.H.; Nelson, W.J. The Adenomatous Polyposis Coli Tumor Suppressor Protein Localizes to Plasma Membrane Sites Involved in Active Cell Migration. J. Cell Biol. 1996, 134, 165–179. [Google Scholar] [CrossRef]
- Penman, G.A.; Leung, L.; Näthke, I.S. The adenomatous polyposis coli protein (APC) exists in two distinct soluble complexes with different functions. J. Cell Sci. 2005, 118, 4741–4750. [Google Scholar] [CrossRef]
- Dikovskaya, D.; Li, Z.; Newton, I.P.; Davidson, I.; Hutchins, J.R.A.; Kalab, P.; Clarke, P.R.; Näthke, I.S. Microtubule assembly by the Apc protein is regulated by importin-β-RanGTP. J. Cell Sci. 2010, 123, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Madinya, J.J.; Chang, L.-W.; Perry, S.L.; Sing, C.E. Sequence-dependent self-coacervation in high charge-density polyampholytes. Mol. Syst. Des. Eng. 2020, 5, 632–644. [Google Scholar] [CrossRef]
- Simon, J.R.; Carroll, N.J.; Rubinstein, M.; Chilkoti, A.; Lopez, G.P. Programming molecular self-assembly of intrinsically disordered proteins containing sequences of low complexity. Nat. Chem. 2017, 9, 509–515. [Google Scholar] [CrossRef]
- Pearlman, S.M.; Serber, Z.; Ferrell, J.E., Jr. A mechanism for the evolution of phosphorylation sites. Cell 2011, 147, 934–946. [Google Scholar] [CrossRef]
- Tang, Y.; Bera, S.; Yao, Y.; Zeng, J.; Lao, Z.; Dong, X.; Gazit, E.; Wei, G. Prediction and characterization of liquid-liquid phase separation of minimalistic peptides. Cell Rep. Phys. Sci. 2021, 2, 100579. [Google Scholar] [CrossRef]
- Li, Z.; Näthke, I.S. Tumor-associated NH2-terminal fragments are the most stable part of the adenomatous polyposis coli protein and can be regulated by interactions with COOH-terminal domains. Cancer Res. 2005, 65, 5195–5204. [Google Scholar] [CrossRef]
- Abbott, J.; Näthke, I.S. The adenomatous polyposis coli protein 3o years on. bioRxiv 2022. [Google Scholar] [CrossRef]
- Nathke, I.S. The adenomatous polyposis coli protein: The Achilles heel of the gut epithelium. Annu. Rev. Cell Dev. Biol. 2004, 20, 337–366. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.; Lipinski, W.P.; Nakashima, K.K.; Huck, W.T.S.; Spruijt, E. A short peptide synthon for liquid-liquid phase separation. Nat. Chem. 2021, 13, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Mayer, G.; Shpilt, Z.; Kowalski, H.; Tshuva, E.Y.; Friedler, A. Targeting Protein Interaction Hotspots Using Structured and Disordered Chimeric Peptide Inhibitors. ACS Chem. Biol. 2022, 17, 1811–1823. [Google Scholar] [CrossRef] [PubMed]
- Weber, P.J.A.; Bader, J.E.; Folkers, G.; Beck-Sickinger, A.G. A fast and inexpensive method for N-terminal fluorescein-labeling of peptides. Bioorg. Med. Chem. Lett. 1998, 8, 597–600. [Google Scholar] [CrossRef] [PubMed]
- Gabizon, R.; Mor, M.; Rosenberg, M.M.; Britan, L.; Hayouka, Z.; Kotler, M.; Shalev, D.E.; Friedler, A. Using peptides to study the interaction between the p53 tetramerization domain and HIV-1 Tat. Biopolymers 2008, 90, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Grunhaus, D.; Molina, E.R.; Cohen, R.; Stein, T.; Friedler, A.; Hurevich, M. Accelerated Multiphosphorylated Peptide Synthesis. Org. Process. Res. Dev. 2022, 26, 2492–2497. [Google Scholar] [CrossRef]
- Boeynaems, S.; De Decker, M.; Tompa, P.; Van Den Bosch, L. Arginine-rich Peptides Can Actively Mediate Liquid-liquid Phase Separation. Bio-Protocol 2017, 7, e2525. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence |
|---|---|
| 20R3 | GFSCSSSLSALSLDE |
| 20R3CK1(pS1504,1507) | GFSCSpSSLpSALSLDE |
| 20R3GSK3β(pS1501,1505) | GFpSCSSpSLSALSLDE |
| 20R5 | CFSRNDSLSSLDFDD |
| 20R5CK1(pS1863) | CFSRNDSLpSSLDFDD |
| 20R5GSK3β(pS1857,1861) | CFpSRNDpSLSSLDFDD |
| 20R4 | NFSTATSLSDLTIES |
| Peptide | Sequence | Wheel Projection |
|---|---|---|
| Human APC | LHLLEQIRAYCETCWEWQEA |  |
| Drosophila APC2 | LRLLDQILDYCNFLHTQLQS | |
| Consensus | HPHHPPH – – P – – – – – P – – P – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bressler, S.G.; Mitrany, A.; Wenger, A.; Näthke, I.; Friedler, A. The Oligomerization Domains of the APC Protein Mediate Liquid-Liquid Phase Separation That Is Phosphorylation Controlled. Int. J. Mol. Sci. 2023, 24, 6478. https://doi.org/10.3390/ijms24076478
Bressler SG, Mitrany A, Wenger A, Näthke I, Friedler A. The Oligomerization Domains of the APC Protein Mediate Liquid-Liquid Phase Separation That Is Phosphorylation Controlled. International Journal of Molecular Sciences. 2023; 24(7):6478. https://doi.org/10.3390/ijms24076478
Chicago/Turabian StyleBressler, Shachar G., Amit Mitrany, Alon Wenger, Inke Näthke, and Assaf Friedler. 2023. "The Oligomerization Domains of the APC Protein Mediate Liquid-Liquid Phase Separation That Is Phosphorylation Controlled" International Journal of Molecular Sciences 24, no. 7: 6478. https://doi.org/10.3390/ijms24076478
APA StyleBressler, S. G., Mitrany, A., Wenger, A., Näthke, I., & Friedler, A. (2023). The Oligomerization Domains of the APC Protein Mediate Liquid-Liquid Phase Separation That Is Phosphorylation Controlled. International Journal of Molecular Sciences, 24(7), 6478. https://doi.org/10.3390/ijms24076478