
Novel Risk Variants in the Oxytocin Receptor Gene (OXTR) Possibly Linked to and Associated with Familial Type 2 Diabetes



Abstract
1. Background
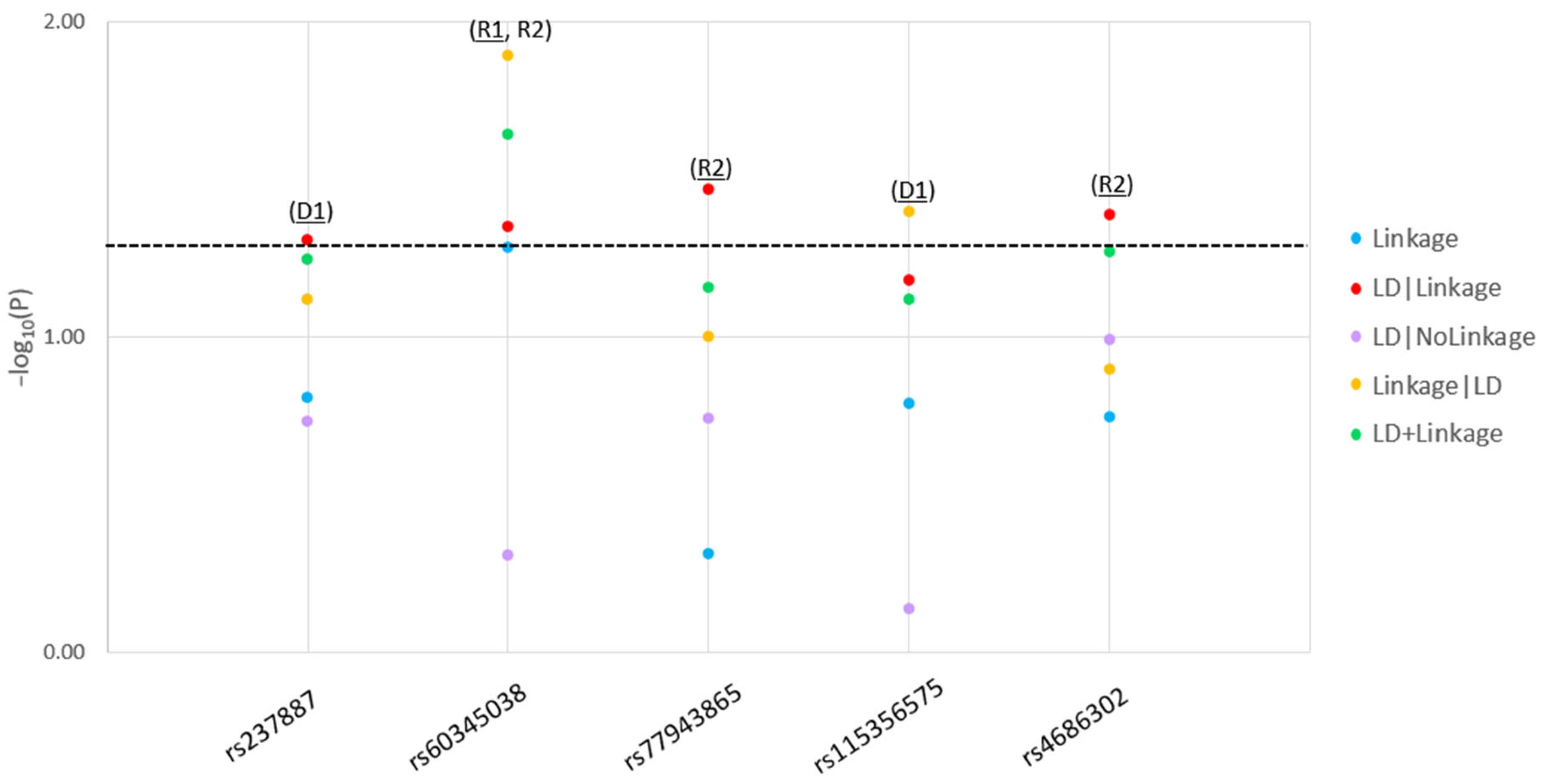
2. Results and Discussion
3. Methods
3.1. Patients
3.2. Genetic Testing and Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baribeau, D.A.; Anagnostou, E. Oxytocin and vasopressin: Linking pituitary neuropeptides and their receptors to social neurocircuits. Front. Neurosci. 2015, 9, 335. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.; Barrera, I.; Brothers, S.; Ring, R.; Wahlestedt, C. Oxytocin and social functioning. Dialogues Clin. Neurosci. 2017, 19, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Veening, J.G.; de Jong, T.R.; Waldinger, M.D.; Korte, S.M.; Olivier, B. The role of oxytocin in male and female reproductive behavior. Eur. J. Pharmacol. 2015, 753, 209–228. [Google Scholar] [CrossRef] [PubMed]
- Onaka, T.; Takayanagi, Y. Role of oxytocin in the control of stress and food intake. J. Neuroendocrinol. 2019, 31, e12700. [Google Scholar] [CrossRef]
- Lee, E.S.; Uhm, K.O.; Lee, Y.M.; Kwon, J.; Park, S.H.; Soo, K.H. Oxytocin stimulates glucose uptake in skeletal muscle cells through the calcium-CaMKK-AMPK pathway. Regul. Pept. 2008, 151, 71–74. [Google Scholar] [CrossRef]
- Ding, C.; Leow, M.K.; Magkos, F. Oxytocin in metabolic homeostasis: Implications for obesity and diabetes management. Obes. Rev. 2019, 20, 22–40. [Google Scholar] [CrossRef]
- Mohan, S.; Khan, D.; Moffett, R.C.; Irwin, N.; Flatt, P.R. Oxytocin is present in islets and plays a role in beta-cell function and survival. Peptides 2018, 100, 260–268. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, C.; Chen, Q.; Chen, X.; Xu, Z.; Wu, J.; Cai, D. Treatment of obesity and diabetes using oxytocin or analogs in patients and mouse models. PLoS ONE 2013, 8, e61477. [Google Scholar] [CrossRef]
- Chatterjee, O.; Patil, K.; Sahu, A.; Gopalakrishnan, L.; Mol, P.; Advani, J.; Mukherjee, S.; Christopher, R.; Prasad, T.S. An overview of the oxytocin-oxytocin receptor signaling network. J. Cell Commun. Signal. 2016, 10, 355–360. [Google Scholar] [CrossRef]
- Kimura, T.; Saji, F.; Nishimori, K.; Ogita, K.; Nakamura, H.; Koyama, M.; Murata, Y. Molecular regulation of the oxytocin receptor in peripheral organs. J. Mol. Endocrinol. 2003, 30, 109–115. [Google Scholar] [CrossRef]
- Watanabe, S.; Wei, F.Y.; Matsunaga, T.; Matsunaga, N.; Kaitsuka, T.; Tomizawa, K. Oxytocin Protects against Stress-Induced Cell Death in Murine Pancreatic β-Cells. Sci. Rep. 2016, 6, 25185. [Google Scholar]
- Davis, C.; Patte, K.; Zai, C.; Kennedy, J.L. Polymorphisms of the oxytocin receptor gene and overeating: The intermediary role of endophenotypic risk factors. Nutr. Diabetes 2017, 7, e279. [Google Scholar] [CrossRef]
- Jacondino, C.B.; Borges, C.A.; Rosemberg, L.S.; da Silva, I.G.; da Luz Correa, B.; Valle Gottlieb, M.G. Association of oxytocin levels and oxytocin receptor gene polymorphism (rs2254298) with cardiovascular risk factors in Brazilian elderly from Primary Health Care. Arch. Gerontol. Geriatr. 2019, 84, 103903. [Google Scholar] [CrossRef] [PubMed]
- Salonen, J.T.; Uimari, P.; Aalto, J.M.; Pirskanen, M.; Kaikkonen, J.; Todorova, B.; Hyppönen, J.; Korhonen, V.P.; Asikainen, J.; Devine, C.; et al. Type 2 diabetes whole-genome association study in four populations: The DiaGen consortium. Am. J. Hum. Genet. 2007, 81, 338–345. [Google Scholar]
- Chang, H.H.; Chang, W.H.; Chi, M.H.; Peng, Y.C.; Huang, C.C.; Yang, Y.K.; Chen, P.S. The OXTR Polymorphism Stratified the Correlation of Oxytocin and Glucose Homeostasis in Non-Diabetic Subjects. Diabetes Metab. Syndr. Obes. 2019, 12, 2707–2713. [Google Scholar] [CrossRef]
- Saravani, R.; Esmaeeli, E.; Kordi Tamendani, M.; Nejad, M.N. Oxytocin Receptor Gene Polymorphisms in Patients with Diabetes. Gene Cell Tissue 2015, 2, e60171. [Google Scholar] [CrossRef]
- Ng, P.C.; Henikoff, S. SIFT: Predicting amino acid changes that affect protein function. Nucleic Acids Res. 2003, 31, 3812–3814. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, K.; Kyriazopoulou Panagiotopoulou, S.; McRae, J.F.; Darbandi, S.F.; Knowles, D.; Li, Y.I.; Kosmicki, J.A.; Arbelaez, J.; Cui, W.; Schwartz, G.B.; et al. Predicting Splicing from Primary Sequence with Deep Learning. Cell 2019, 176, 535–548.e24. [Google Scholar] [CrossRef] [PubMed]
- Dayem Ullah, A.Z.; Oscanoa, J.; Wang, J.; Nagano, A.; Lemoine, N.R.; Chelala, C. SNPnexus: Assessing the functional relevance of genetic variation to facilitate the promise of precision medicine. Nucleic Acids Res. 2018, 46, W109–W113. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Ambrosini, G.; Bucher, P. SNP2TFBS—A database of regulatory SNPs affecting predicted transcription factor binding site affinity. Nucleic Acids Res. 2017, 45, D139–D144. [Google Scholar] [PubMed]
- Boyle, A.P.; Hong, E.L.; Hariharan, M.; Cheng, Y.; Schaub, M.A.; Kasowski, M.; Karczewski, K.J.; Park, J.; Hitz, B.C.; Weng, S.; et al. Annotation of functional variation in personal genomes using RegulomeDB. Genome Res. 2012, 22, 1790–1797. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, F.; Li, T.; Lu, M.; Wang, L.; Yue, W.; Zhang, D. MirSNP, a database of polymorphisms altering miRNA target sites, identifies miRNA-related SNPs in GWAS SNPs and eQTLs. BMC Genom. 2012, 13, 661. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Hiekkalinna, T.; Schaffer, A.A.; Lambert, B.; Norrgrann, P.; Goring, H.H.; Terwilliger, J.D. PSEUDOMARKER: A powerful program for joint linkage and/or linkage disequilibrium analysis on mixtures of singletons and related individuals. Hum. Hered. 2011, 71, 256–266. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Model 1 | SNP | Position | Ref | Alt | Risk Allele | Consequence | RAF 2 | P 3 | Reported in T2D? |
|---|---|---|---|---|---|---|---|---|---|
| D1 | rs237887 | 8755356 | G | A | A | Intronic | 0.54 | 0.04 | Yes [14] |
| R1, R2 | rs60345038 | 8760830 | C | T | C | Intronic | 0.62 | 0.01 | Novel |
| R2 | rs77943865 | 8762293 | G | A | G | Intronic | 0.95 | 0.03 | Novel |
| D1 | rs115356575 | 8764840 | G | A | G | Intronic | 0.96 | 0.04 | Novel |
| R2 | rs4686302 | 8767536 | C | T | C | Missense | 0.89 | 0.04 | Novel |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amin, M.; Wu, R.; Gragnoli, C. Novel Risk Variants in the Oxytocin Receptor Gene (OXTR) Possibly Linked to and Associated with Familial Type 2 Diabetes. Int. J. Mol. Sci. 2023, 24, 6282. https://doi.org/10.3390/ijms24076282
Amin M, Wu R, Gragnoli C. Novel Risk Variants in the Oxytocin Receptor Gene (OXTR) Possibly Linked to and Associated with Familial Type 2 Diabetes. International Journal of Molecular Sciences. 2023; 24(7):6282. https://doi.org/10.3390/ijms24076282
Chicago/Turabian StyleAmin, Mutaz, Rongling Wu, and Claudia Gragnoli. 2023. "Novel Risk Variants in the Oxytocin Receptor Gene (OXTR) Possibly Linked to and Associated with Familial Type 2 Diabetes" International Journal of Molecular Sciences 24, no. 7: 6282. https://doi.org/10.3390/ijms24076282
APA StyleAmin, M., Wu, R., & Gragnoli, C. (2023). Novel Risk Variants in the Oxytocin Receptor Gene (OXTR) Possibly Linked to and Associated with Familial Type 2 Diabetes. International Journal of Molecular Sciences, 24(7), 6282. https://doi.org/10.3390/ijms24076282