
Physicochemical Principles of Adhesion Mechanisms in the Brain

Abstract
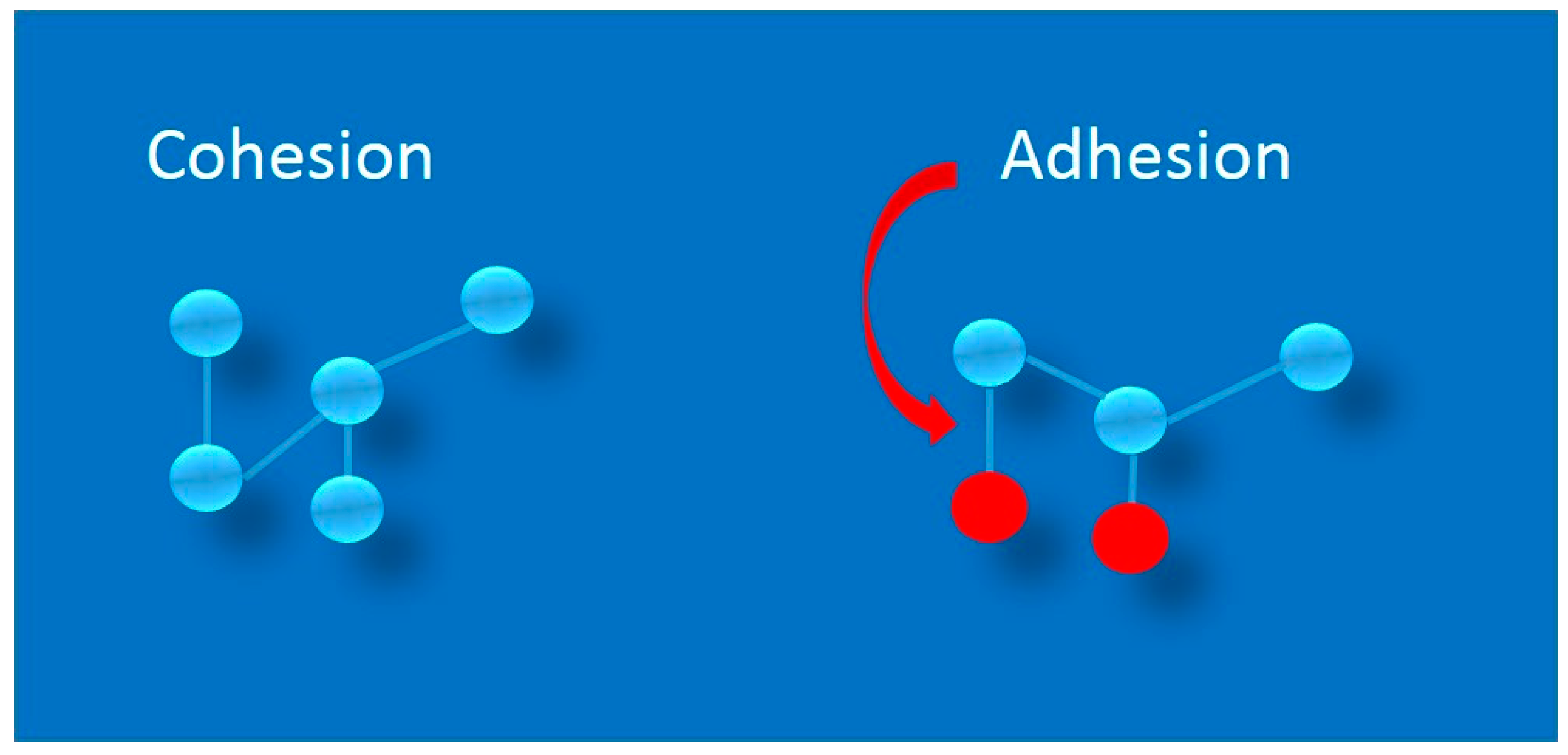
1. Adhesion as a Physicochemical Phenomenon
2. Families of Cell Adhesion Molecules Present in the Brain
3. CAMs in a Health and a Disease
- Cancer–One of the CAMs that plays a vital role in cancer is cadherins [18]. It was shown that the E-cadherin-catenin complex is a fundamental part of epithelial monolayers, upholding homeostasis and maintenance [18]. Losing E-cadherins results in cancer progression [18]. One of the mechanisms in that action is the formation of mature junctions that suppress tumor invasion [18]. Furthermore, a loss of E-cadherins releases cellular signals that promote tumor growth and infiltration, e.g., Rho GTPases, PI3K et cetera [18]. The significant role of selectins in tumorigenesis and inflammation has also been demonstrated [16,32]. Selectins, via adhesive mechanisms, protect the organism from bacterial infections and are involved in trafficking cells of the innate immune system, T lymphocytes, and platelets [16]. E-selectin expression increases up to 6 h after inflammatory stimuli, e.g., interleukin-1 [32]. The mechanism selectins utilize is a rolling adhesion through selectin-ligand interactions, described in detail by McEver [23]. Selectins’ role in tumorigenesis has also been established [16,32]. They facilitate the hematogenous dissemination of tumor cells and their arrest in the microvasculature by activating adhesive mechanisms between selectin-expressing host cells and ligands of tumor cells [32].
- Neurological disorders–The role of CAMs in neurological disorders is well documented. The connections with depression, schizophrenia, Parkinson’s disease, autism spectrum disorders, Alzheimer’s disease, Down syndrome, epilepsy, and others were found [12,22,25,33,34,35]. When NCAMs are attached to the polysialic acid (PolySia-NCAMs), they play a leading role in schizophrenia as vital regulators of synaptic plasticity, cell migration, axon guidance, and synapse formation [35]. The loss of PolySia-NCAM in NCAM knockout mice resulted in impaired learning, memory, LTP, or LTD [35]. On the other side, increased PolySia-NCAM immunoreactivity was documented in the schizophrenic hippocampus (HC) or prefrontal cortex (PFC) [35]. NCAM isoforms and soluble NCAM fragments were shown as concentrated in the cerebrospinal fluid of schizophrenic patients [35]. The changes in the level of NCAMs or their metabolism correspond with changes in the volume of the left PFC (Brodmann area 46) in schizophrenic patients [35]. Furthermore, PolySia-NCAMs were shown as involved in negative symptoms of schizophrenia [35]. Social affiliative behavior undergoes sCAMs regulation via mechanisms of synapse formation, development, and plastic changes [25].
- Learning and memory–involve a vast array of CAMs [12,22,29]; therefore, we will present only selected examples here. Following Wu and Reedy [22], integrins are involved in learning and memory through a few mechanisms. One of the mechanisms is the modulation of long-term potentiation (LTP) [22]. The effect of integrins on LTP was demonstrated by a knockout of the gene encoding α-integrin in Drosophila, resulting in impaired short-term olfactory learning [22,41]. Similar results were documented in mice [22]. Integrins may regulate LTP via modulating N-methyl-D-aspartic acid receptors (NMDARs) or α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors (AMPARs) activity [29]. Furthermore, any functional changes associated with integrins result in morphological changes in a synapse structure and function [22]. This is due to the available combination and influence on the scaffolding proteins and actin in spine heads and necks [29]. Cadherins, similarly to integrins, regulate synapse behavior upon NMDA activation [29]. However, their action is required for LTP and spine enlargement, but not for LTD and spine density and morphology, which indicates their participation in synapse plasticity [29]. IgSFs, on the other hand, e.g., NCAMs, regulate synapses via crosslinking to NMDARs and CaMKII via postsynaptic scaffolds [29]. NCAMs may interact with dyneins, enhancing synapse stability, and as states to SynCAMs enlarge during LTD controlling cleft diameter [29]. Our studies found connections between mGluR5, which are postsynaptically localized with DSCAM in a context of spatial learning and depression [42].
- Circadian clock functions–are controlled by the Ephrin receptor: EphA4, a CAMs are expressed in the suprachiasmatic nucleus (SCN) [43]. Its documented role in this phenomenon is to regulate the connections of neurons and astrocytes in the SCN [43]. The role of EphA4 in circadian clock functions was documented with EphA4-/- mice use [43]. EphA4-/- mice are characterized by reduced running activity, more extended endogenous periods under constant darkness, and shorter periods under continuous light conditions [43]. In addition, the inactivation of PSA-NCAM (polysialylated neural cell adhesion molecule) was able to extend the length of the circadian clock periods in mice [43,44].
- Adhesive tissue engineered scaffolds (ATEs)-have been developed to repair damaged tissues and guide tissue regeneration after injury and degeneration [45]. ATEs have found applications in nerve regeneration, cartilage repair, corneal regeneration, skin regeneration, cardiac tissue repair, bone repair and others [45]. Adhesive scaffolds can be created using various types of biomaterials (hydrogels, assembled microgel spheres, foams, electrospheres) [45]. ATEs provide a three-dimensional (3D) biomimetic and highly biocompatible environment for cell adhesion, growth, proliferation, differentiation, secretion of extracellular matrix proteins, and remodeling or scaffold replacement with regenerated tissue during matrix degradation [45]. This topic was comprehensively described by Chen et al. [45].Table 1 presents selected conditions and functions in which CAMs have been confirmed to be involved.
4. The Role of CAMs in a Synapse
5. The Idea of Assessing the Physicochemical Phenomenon of Adhesion in Mental and Cognitive Disorders
6. Selected Methods for Evaluating Cell Adhesion
7. Conclusive Remarks
Funding
Conflicts of Interest
References
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Tadmor, R.; Das, R.; Gulec, S.; Liu, J.; N’guessan, H.E.; Shah, M.; Wasnik, P.F.; Yadav, S.B. Solid–Liquid Work of Adhesion. Langmuir 2017, 33, 3594–3600. [Google Scholar] [CrossRef] [PubMed]
- Kendall, K.; Roberts, A.D. Van der Waals forces influencing adhesion of cells. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140078. [Google Scholar] [CrossRef] [PubMed]
- Varenberg, M.; Peressadko, A.; Gorb, S.; Arzt, E. Effect of real contact geometry on adhesion. Appl. Phys. Lett. 2006, 89, 121905. [Google Scholar] [CrossRef]
- Ladoux, B.; Nicolas, A. Physically based principles of cell adhesion mechanosensitivity in tissues. Rep. Prog. Phys. 2012, 75, 116601. [Google Scholar] [CrossRef]
- Havlica, J.; Kramolis, D.; Huchet, F. A revisit of the electro-diffusional theory for the wall shear stress measurement. Int. J. Heat Mass Transf. 2021, 165, 120610. [Google Scholar] [CrossRef]
- Stachowicz, K. The role of polyunsaturated fatty acids in neuronal signaling in depression and cognitive processes. Arch. Biochem. Biophys. 2023, 737, 109555. [Google Scholar] [CrossRef]
- Azuri, I.; Meirzadeh, E.; Ehre, D.; Cohen, S.R.; Rappe, A.M.; Lahav, M.; Lubomirsky, I.; Kronik, L. Usually large Young’s moduli of amino acid molecular crystals. Angew. Chem. Int. Ed. 2015, 54, 13566–13570. [Google Scholar] [CrossRef]
- Blom, K.; Godec, A. Criticality in Cell Adhesion. Phys. Rev. X 2021, 11, 031067. [Google Scholar] [CrossRef]
- McEver, R.P.; Luscinskas, F.W. Hematology, 7th ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 127–134. [Google Scholar]
- Dalva, M.B.; McClelland, A.C.; Kayser, M.S. Cell adhesion molecules: Signalling functions at the synapse. Nat. Rev. Neurosci. 2007, 8, 206–220. [Google Scholar] [CrossRef]
- Stachowicz, K. The role of DSCAM in the regulation of synaptic plasticity: Possible involvement in neuropsychiatric disorders. Acta Neurobiol. Exp. 2018, 78, 210–219. [Google Scholar] [CrossRef]
- Nielsen, H.; Wennström, M. Cell adhesion molecules in Alzheimer’s disease. Degener. Neurol. Neuromuscul. Dis. 2012, 2, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Dieterle, M.P.; Husari, A.; Rolauffs, B.; Steinberg, T.; Tomakidi, P. Integrins, cadherins and channels in cartilage mechanotransduction: Perspectives for future regeneration strategies. Expert Rev. Mol. Med. 2021, 23, e14. [Google Scholar] [CrossRef]
- Hynes, R.O. Integrins: Bidirectional, Allosteric Signaling Machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef]
- Ley, K. The role of selectins in inflammation and disease. Trends Mol. Med. 2003, 9, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Barclay, A.N. Membrane proteins with immunoglobulin-like domains—A master superfamily of interaction molecules. Semin. Immunol. 2003, 15, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Kourtidis, A.; Lu, R.; Pence, L.J.; Anastasiadis, P.Z. A central role for cadherin signaling in cancer. Exp. Cell Res. 2017, 358, 78–85. [Google Scholar] [CrossRef]
- Nose, A.; Nagafuchi, A.; Takeichi, M. Isolation of placental cadherin cDNA: Identification of a novel gene family of cell-cell adhesion molecules. EMBO J. 1987, 6, 3655–3661. [Google Scholar] [CrossRef]
- Hirano, S.; Takeichi, M. Cadherins in Brain Morphogenesis and Wiring. Physiol. Rev. 2012, 92, 597–634. [Google Scholar] [CrossRef]
- Campbell, I.D.; Humphries, M.J. Integrin structure, activation, and interactions. Cold Springs Harb. Perspect. Biol. 2011, 3, a004994. [Google Scholar] [CrossRef]
- Wu, X.; Reddy, D.S. Integrins as receptor targets for neurological disorders. Pharmacol. Ther. 2012, 134, 68–81. [Google Scholar] [CrossRef]
- McEver, R.P. Selectins: Initiators of leucocyte adhesion and signalling at the vascular wall. Cardiovasc. Res. 2015, 107, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Gout, S.; Tremblay, P.-L.; Huot, J. Selectins and selectin ligands in extravasation of cancer cells and organ selectivity of metastasis. Clin. Exp. Metastasis 2007, 25, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.C.; Ferri, S.L.; Grewal, M.; Smernoff, Z.; Bucan, M.; Weiner, J.A.; Abel, T.; Brodkin, E.S. The Role of Synaptic Cell Adhesion Molecules and Associated Scaffolding Proteins in Social Affiliative Behaviors. Biol. Psychiatry 2020, 88, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.M.; Wang, Y.; Lee, J.T.H.; Huang, Z.; Wu, D.; Xu, A.; Lam, K.S.L. Adropin is a brain membrane-bound protein regulating physical activity via the NB-3/Notch signaling pathway in mice. J. Biol. Chem. 2014, 289, 25976–25986. [Google Scholar] [CrossRef] [PubMed]
- Vactor, D.V. Adhesion and signaling in axonal fasciculation. Curr. Opin. Neurobiol. 1998, 8, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.A.; Fuertes, A.J.; Short, G.; Donohue, K.C.; Shao, H.; Quintanilla, J.; Malakzadeh, P.; Cohen-Cory, S. DSCAM differentially modulates pre- and postsynaptic structural and functional central connectivity during visual system wiring. Neur. Dev. 2018, 13, 22. [Google Scholar] [CrossRef]
- Kilinc, D. The Emerging Role of Mechanics in Synapse Formation and Plasticity. Front. Cell. Neurosci. 2018, 12, 483. [Google Scholar] [CrossRef]
- Malik, B.R.; Hodge, J.J.L. CASK and CaMKII function in Drosophila memory. Front. Neurosci. 2014, 8, 178. [Google Scholar] [CrossRef]
- Lilja, J.; Ivaska, J. Integrin activity in neuronal connectivity. J. Cell Sci. 2018, 131, jcs212803. [Google Scholar] [CrossRef]
- Cheung, L.S.-L.; Raman, P.S.; Balzer, E.M.; Wirtz, D.; Konstantopoulos, K. Biophysics of selectin–ligand interactions in inflammation and cancer. Phys. Biol. 2011, 8, 015013. [Google Scholar] [CrossRef]
- Ganesh, R.A.; Venkataraman, K.; Sirdeshmukh, R. GPR56: An adhesion GPCR involved in brain development, neurological disorders and cancer. Brain Res. 2020, 1747, 147055. [Google Scholar] [CrossRef] [PubMed]
- Izumi, Y.; Wakita, S.; Kanbara, C.; Nakai, T.; Akaike, A.; Kume, T. Integrin α5β1 expression on dopaminergic neurons is involved in dopaminergic neurite outgrowth on striatal neurons. Sci. Rep. 2017, 7, 42111. [Google Scholar] [CrossRef] [PubMed]
- Piras, F.; Schiff, M.; Chiapponi, C.; Bossù, P.; Mühlenhoff, M.; Caltagirone, C.; Gerardy-Schahn, R.; Hildebrandt, H.; Spalletta, G. Brain structure, cognition and negative symptoms in schizophrenia are associated with serum levels of polysialic acid-modified NCAM. Transl. Psychiatry 2015, 5, e658. [Google Scholar] [CrossRef] [PubMed]
- Alves-Sampaio, A.; Troca-Marin, J.A.; Montesinos, M.L. NMDA-mediated regulation of DSCAM dendritic local translation is lost in a mouse model of Down’s syndrome. J. Neurosci. 2010, 30, 13537–13548. [Google Scholar] [CrossRef]
- Antonarakis, S.E.; Lyle, R.; Dermitzakis, E.T.; Reymond, A.; Deutsch, S. Chromosome 21 and Down syndrome: From genomics to pathophysiology. Nat. Rev. Genet. 2004, 5, 725–738. [Google Scholar] [CrossRef]
- Belichenko, P.V.; Kieschevnikov, A.M.; Becker, A.; Wagner, G.E.; Lysenko, L.V.; Yu, Y.E.; Mobley, W.C. Down syndrome cognitive phenotypes modeled in mice trisomic for AII HAS 21 homologues. PLoS ONE 2015, 10, e0134861. [Google Scholar] [CrossRef]
- Siarey, R.J.; Kline-Burgess, A.; Cho, M.; Balbo, A.; Best, T.K.; Harashima, C.; Klann, E.; Galdzicki, Z. Altered signaling pathways underlying abnormal hippocampal synaptic plasticity in the Ts65Dn mouse model of Down syndrome. J. Neurochem. 2006, 98, 1266–1277. [Google Scholar] [CrossRef]
- Liu, W.; Zheng, Y.; Zhang, F.; Zhu, M.; Guo, Q.; Xu, H.; Liu, C.; Chen, H.; Wang, X.; Hu, Y.; et al. A Preliminary Investigation on Plasma Cell Adhesion Molecules Levels by Protein Microarray Technology in Major Depressive Disorder. Front. Psychiatry 2021, 12, 627469. [Google Scholar] [CrossRef]
- Grotewiel, M.S.; Beck, C.D.O.; Wu, K.H.; Zhu, X.-R.; Davis, R.L. Integrin-mediated short-term memory in Drosophila. Nature 1998, 391, 455–460. [Google Scholar] [CrossRef]
- Stachowicz, K.; Pańczyszyn-Trzewik, P.; Rzeźniczek, S.; Sowa-Kućma, M. Short Communication. Cyclooxygenase-2 inhibition affects the expression of Down syndrome cell adhesion molecule (DSCAM) through interaction with mGluR5. JPP 2022, 73, 319–326. [Google Scholar] [CrossRef]
- Kiessling, S.; O’Callaghan, E.K.; Freyburger, M.; Cermakian, N.; Mongrain, V. The cell adhesion molecule EphA4 is involved in circadian clock functions. Genes Brain Behav. 2018, 17, 82–92. [Google Scholar] [CrossRef]
- Shen, H.; Watanabe, M.; Tomasiewicz, H.; Rutishauser, U.; Magnuson, T.; Glass, J.D. Role of Neural Cell Adhesion Molecule and Polysialic Acid in Mouse Circadian Clock Function. J. Neurosci. 1997, 17, 5221–5229. [Google Scholar] [CrossRef]
- Chen, S.; Gil, C.J.; Ning, L.; Jin, L.; Perez, L.; Kabboul, G.; Tomov, M.L.; Serpooshan, V. Adhesive Tissue Engineered Scaffolds: Mechanisms and Applications. Front. Bioeng. Biotechnol. 2021, 9, 683079. [Google Scholar] [CrossRef]
- Danen, E.H. Integrins: An overview of structural and functional aspects. In Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Stachowicz, K. Is PSD-95 entangled in the side effects of antidepressants? Neurochem. Int. 2022, 159, 105391. [Google Scholar] [CrossRef] [PubMed]
- Gires, O.; Pan, M.; Schinke, H.; Canis, M.; Baeuerle, P.A. Expression and function of epithelial cell adhesion molecule EpCAM: Where are we after 40 years? Cancer Met. Rev. 2020, 39, 969–987. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, T.; Matsuda, H.; Tabira, T.; Asada, T.; Uno, M. Changes in brain morphology in Alzheimer disease and normal aging: Is Alzheimer disease an exaggerated aging process? AJNR Am. J. Neuroradiol. 2001, 22, 1680–1685. [Google Scholar] [PubMed]
- Miguel-Hidalgo, J.J.; Rajkowska, G. Morphological brain changes in depression, Can antidepressants reverse them? CNS Drugs 2002, 16, 361–372. [Google Scholar] [CrossRef]
- Duman, C.H.; Duman, R.S. Spine synapse remodeling in the pathophysiology and treatment of depression. Neurosci. Lett. 2015, 601, 20–29. [Google Scholar] [CrossRef]
- Popoli, M.; Yan, Z.; McEwen, B.S.; Sanacora, G. The stressed synapse: The impact of stress and glucocorticoids on glutamate transmission. Nat. Rev. Neurosci. 2013, 13, 22–37. [Google Scholar] [CrossRef]
- Dong, X.; Liu, O.W.; Howell, A.S.; Shen, K. An Extracellular Adhesion Molecule Complex Patterns Dendritic Branching and Morphogenesis. Cell 2013, 155, 296–307. [Google Scholar] [CrossRef]
- Stachowicz, K. Deciphering the mechanisms of regulation of an excitatory synapse via cyclooxygenase-2. A review. Biochem. Pharmacol. 2021, 192, 114729. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.A.; Calderwood, D.A. Forces and Bond Dynamics in Cell Adhesion. Science 2007, 316, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Luo, B.-H.; Xiao, T.; Zhang, C.; Nishida, N.; Springer, T.A. Structure of a Complete Integrin Ectodomain in a Physiologic Resting State and Activation and Deactivation by Applied Forces. Mol. Cell 2008, 32, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Nir, S.; Andersen, M. Van der Waals interactions between cell surfaces. J. Membr. Biol. 1977, 31, 1–18. [Google Scholar] [CrossRef]
- Alon, R.; Dustin, M. Force as a Facilitator of Integrin Conformational Changes during Leukocyte Arrest on Blood Vessels and Antigen-Presenting Cells. Immunity 2007, 26, 17–27. [Google Scholar] [CrossRef]
- Santamaria, F.; Gonzalez, J.; Augustine, G.J.; Raghavachari, S. Quantifying the Effects of Elastic Collisions and Non-Covalent Binding on Glutamate Receptor Trafficking in the Post-Synaptic Density. PLOS Comput. Biol. 2010, 6, e1000780. [Google Scholar] [CrossRef]
- Gupta, R. Self-crowding of AMPA receptors in the excitatory postsynaptic density can effectuate anomalous receptor sub-diffusion. PLOS Comput. Biol. 2018, 14, e1005984. [Google Scholar] [CrossRef]
- Li, T.P.; Song, Y.; MacGillavry, H.D.; Blanpied, T.A.; Raghavachari, S. Protein Crowding within the Postsynaptic Density Can Impede the Escape of Membrane Proteins. J. Neurosci. 2016, 36, 4276–4295. [Google Scholar] [CrossRef]
- Ungai-Salánki, R.; Peter, B.; Gerecsei, T.; Orgovan, N.; Horvath, R.; Szabó, B. A practical review on the measurement tools for cellular adhesion force. Adv. Colloid Interface Sci. 2019, 269, 309–333. [Google Scholar] [CrossRef]
- Khalili, A.A.; Ahmad, M.R. A Review of Cell Adhesion Studies for Biomedical and Biological Applications. Int. J. Mol. Sci. 2015, 16, 18149–18184. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family of CAMs | Role/Disease | References |
|---|---|---|
| Cadherins | -cell recognition, signaling, communication, morphogenesis, angiogenesis, neurotransmission, cancer | [14,18] |
| Integrins | -cell migration/motility–embryonic development, immune response, tissue repair, cancers -signal transduction cascades–differentiation, proliferation, survival -learning and memory, Alzheimer’s disease, Down’s syndrome, injury and stroke, epilepsy | [22,46] |
| Selectins | -hematogenous dissemination of tumor cells/cancer | [32] |
| IgSFs | -cell-cell reorganization, immune system functioning -neuropsychiatric disorders, LTP, synaptic plasticity -changes in the level of DSCAM were found in Down syndrome, Alzheimer’s disease, depression, PTSD -sCAMS as a representing several protein families of CAMs–participate in maintenance, function and elimination of synapses -autism, schizophrenia, social phobia, intellectual disability, attention deficits, bipolar depression, circadian clock functions -changes in a spine density and number, PSD-95 binding, scaffolds regulation | [12,17,25,43] |
| Methodology | Description | Application | Ref. |
|---|---|---|---|
| Cell attachment events | |||
| polyacrylamide-traction force microscopy (PA-TFM) | -technique examines single traction forces-through contact to substrate surface -cells are grown on a polyacrylamide gel with a functionalized cell adhesion ligand and fluorescent beads embedded near the gel surface -as a result of adhesion, the cells generate traction forces that move the beads, and the movement is quantified by tracking the displacement of the fluorescent beads | -real time observation -adaptable to a large variety of cells | [63] |
| micropatterning | -it relies on basic elastic beam theory -controls micro-patterning approaches involve controlling cellular shape, attachment, spreading | -single cell studies -multi-cellular arrangements in populations of cells -sensitivity and cell response to microenvironment | |
| three-dimensional traction force quantification (3D-TFM) | -gel-3D cell culture -cells grow inside a gel matrix embedded with fluorescent beads surrounding the cell | -real-time observation -adaptable to a large variety of cells -traction map of tumor cells -the effects of drugs on cells -cell motility -stem cell functions | |
| wash assay | -static and dynamic culture -after washing–cells that remain adhered to the substrate are analyzed | -for cell count, quantification of DNA content, protein count, antibody binding | |
| resonance frequency | -changes in resonance frequency are detected at the interaction between cell membrane and substrate | -real-time observation and measurement | |
| microfluidics | -applies fluid movement during the cell culturing and adhesion -the balance between the adhesive forces generated by the interactions of membrane-bound receptors and their ligands with the dispersive hydrodynamic forces determines cell adhesion | -real-time observation and measurement -cancer cells migration -tumor cell adhesion | |
| Cell detachment events | |||
| micropipette aspiration technique | -technique detaches an immobilized cell by applying suction force to a portion of the cell surface employed by micropipette suction under observation via a microscope -adhesion strength–minimum force needed to detach a single cell from its substrate | -real-time observation and measurement -mechanical properties of single cells | |
| single cell spectroscopy (SCFS) | -a microscope is used to observe the cell, while a force is applied to detach the cell using a nanomanipulator, micromanipulator, or micropipette | -real-time observation for short-term adhesion -to study the structures and mechanics of isolated biomolecules, cell nucleus, cytoskeleton | |
| AFM probe force measurement | -the single cell is immobilized on the AFM cantilever, the live cell is transformed into a probe to measure the adhesion force between cells or the cell matrix | -real-time observation for short-term adhesion -stiffness and adhesion strength against mechanical force | |
| bio-membrane force probe (BFP) | -using a force transducer (biotinylated erythrocyte) held by a glass micropipette | -real-time observation for short-term adhesion -quantification of a single molecular bond | |
| optical tweezers (OT) | -using a highly focused laser beam to trap and manipulate particles | -real-time observation for short-term adhesion | |
| centrifugation assay | -cultures of cells-to assess the strength of adhesion, the number of cells is quantified before and after the load is applied in the centrifuge | -many analysis in parallel | |
| spinning disc | -takes advantage of the shear stresses generated by the rotating disk device | -high stresses | |
| flow chamber | -radial or parallel flow chambers | -rea-time cell detachment observation | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stachowicz, K. Physicochemical Principles of Adhesion Mechanisms in the Brain. Int. J. Mol. Sci. 2023, 24, 5070. https://doi.org/10.3390/ijms24065070
Stachowicz K. Physicochemical Principles of Adhesion Mechanisms in the Brain. International Journal of Molecular Sciences. 2023; 24(6):5070. https://doi.org/10.3390/ijms24065070
Chicago/Turabian StyleStachowicz, Katarzyna. 2023. "Physicochemical Principles of Adhesion Mechanisms in the Brain" International Journal of Molecular Sciences 24, no. 6: 5070. https://doi.org/10.3390/ijms24065070
APA StyleStachowicz, K. (2023). Physicochemical Principles of Adhesion Mechanisms in the Brain. International Journal of Molecular Sciences, 24(6), 5070. https://doi.org/10.3390/ijms24065070