
Molecular Mechanisms of Neurogenic Inflammation of the Skin

 ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Neuroimmune Communication (NIC)
2.1. Neuropeptides (NPs)
2.2. Neurotrophins (NTs)
2.2.1. Calcitonin Gene-Related Peptide (CGRP)
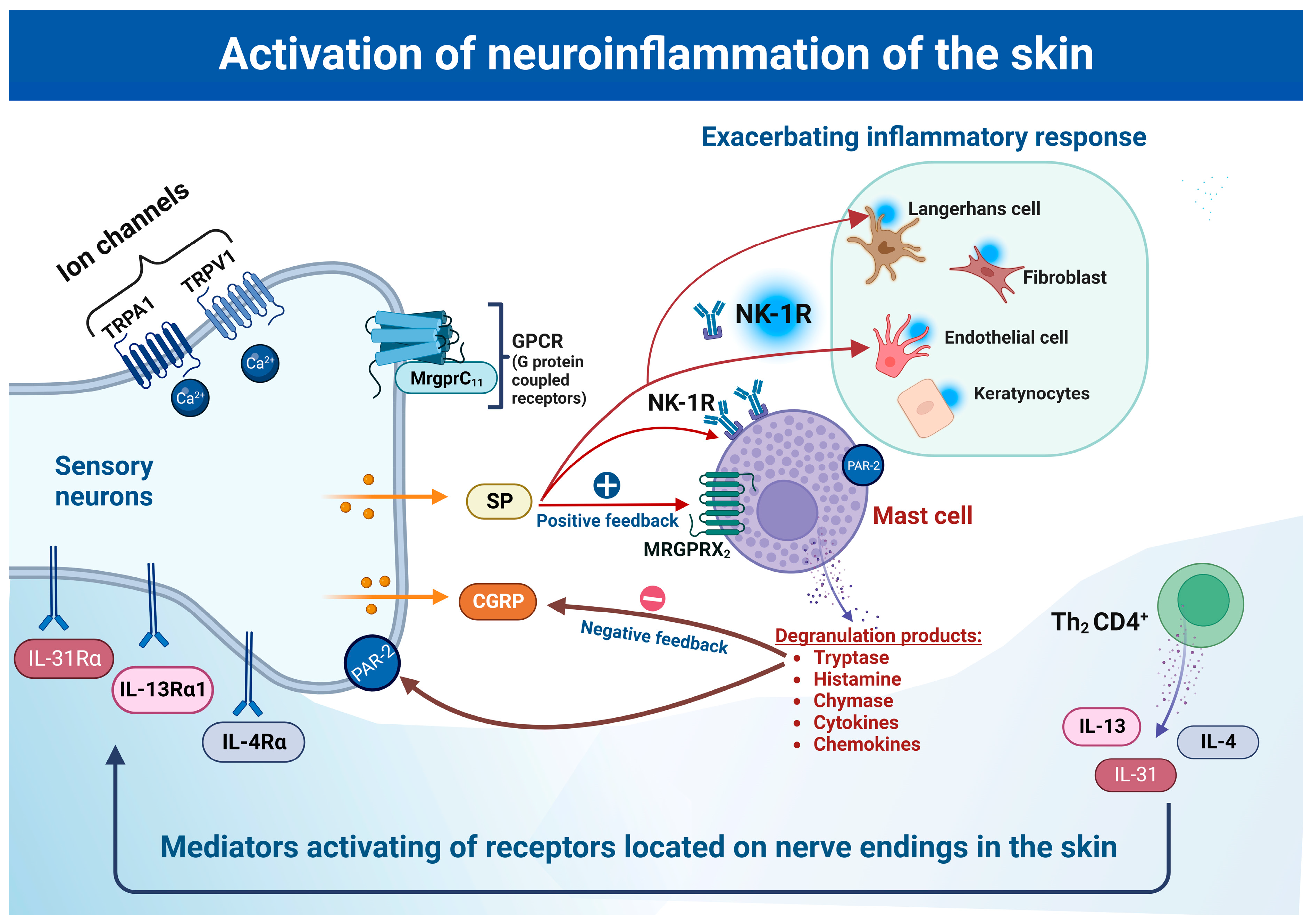
2.2.2. Substance P (SP) and Its Role in Neuroinflammation
3. Receptors in Neuro-Immune Interaction
3.1. Neurokinin Receptor (NK-R)
3.2. Tropomyosin Receptor Kinase A
3.3. Mas-Related G-Coupled Protein Receptors (Mrgprs)
3.4. Transient Receptor Potential (TRP)
3.5. Role of Protease-Activated Receptors (PARs)
4. Mast Cells as Major Mediators of Neuroimmune Crosstalk
4.1. The Role of Mast Cells in Neuroinflammation
4.2. Mast Cell-Induced Disease
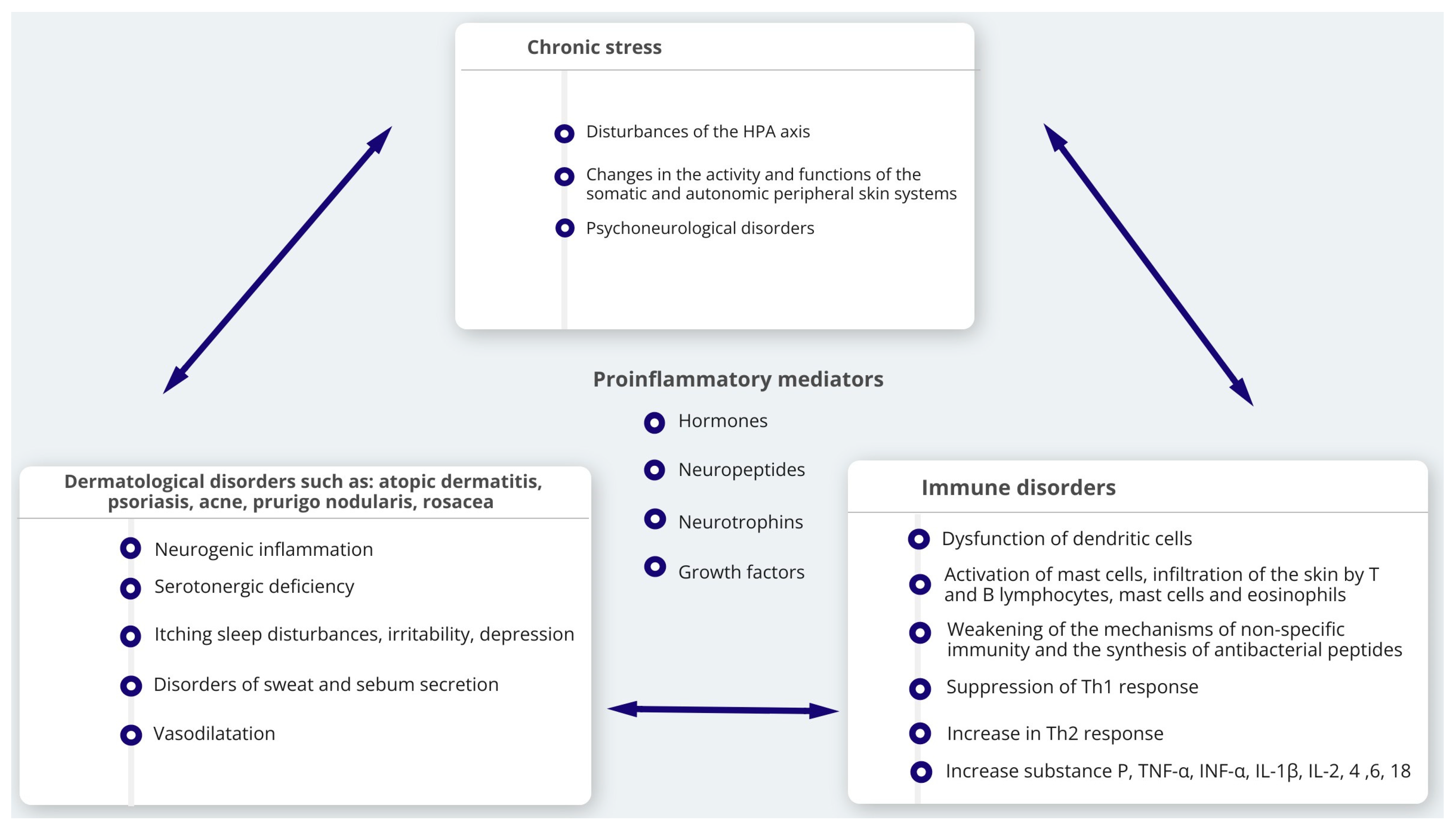
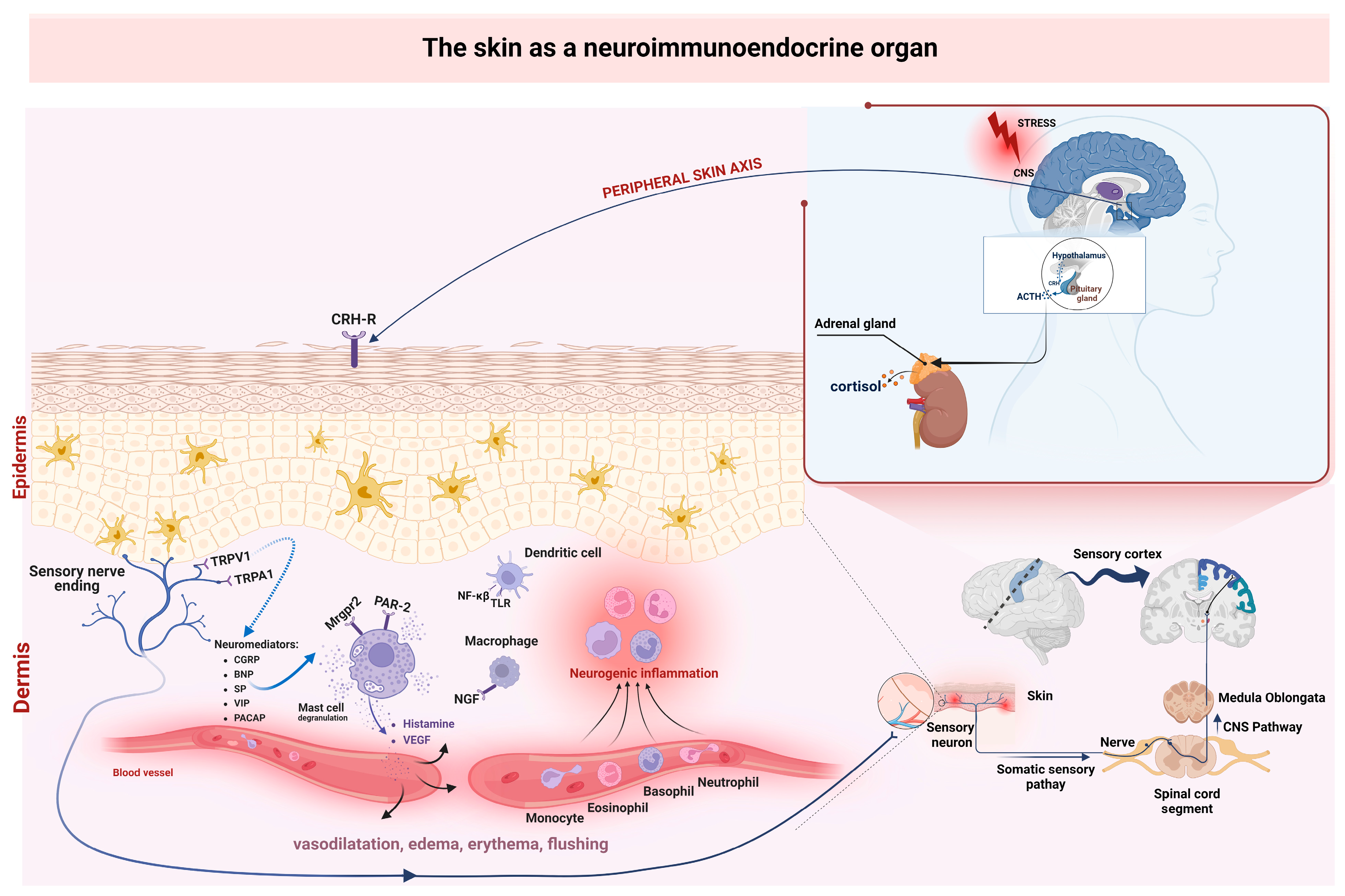
5. Influence of Stress in Formation of Neurogenic Skin Inflammation
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| α-MSH | α-melanocyte stimulating hormone |
| AD | atopic dermatitis |
| AP-1 | activating protein-1 |
| BDNF | brain-derived neurotrophic factor |
| CGRP | Calcitonin gene-related peptide |
| CGRP-KCM | CGRP-stimulated keratinocytes |
| CNI | cutaneous neurogenic inflammation |
| CRF/-H | corticotropin-releasing factor/- hormone |
| CSU | chronic spontaneous urticaria |
| DRG | dorsal root ganglion |
| DAMPs | damage-associated molecular patterns |
| ICAM | intracellular adhesion molecule |
| iCa2+ | intracellular Ca2+ |
| HMGB1 | high mobility group box 1 |
| HPA | hypothalamic-pituitary-adrenal |
| HSP | heat shock protein |
| MCAS | mast cell activation syndrome |
| MCs | mast cells |
| Mrgprs | Mas-related G-coupled protein receptors |
| MRGPRX2 | Mas-related G protein-coupled receptor X2 |
| MMPs | matrix metalloproteinases |
| NF-κB | nuclear factor κB |
| NGF | nerve growth factor |
| NGF-TrkA | nerve growth factor (NGF)-Tropomyosin-related kinase A receptor (TrkA) |
| NIC | neuro-immuno-cutaneous |
| NICE | neuro-immuno-cutaneous-endocrine |
| NK1- 3R | neurokinin-1−3 receptors |
| NPY | neuropeptide Y |
| NT-3/4 | Neurotrophin-3/-4 |
| PAF | platelet-activating factor |
| PAMP | pathogen-associated molecular pattern |
| PAR | protease-activated receptor |
| PG | prostaglandin |
| POMC | proopiomelanocortin |
| PRR | pattern-recognition receptor |
| SM | systemic mastocytosis |
| SP | substance P |
| SST | Somatostatin |
| TLR | Toll-like receptors |
| TrkA | tropomyosin receptor kinase A |
| TRP | transient receptor potential |
| TRPA | transient receptor potential Ankyrin |
| TRPV | transient receptor potential vanilloid |
| TSLP | thymic stromal lymphopoietin |
| VCAM | vascular cell adhesion molecule |
| VEGF | vascular endothelial growth factor |
| VIP | vasoactive intestinal peptide |
References
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the Environment: Regulation of Local and Global Homeostasis by the Skin’s Neuroendocrine System. Adv. Anat. Embryol. Cell Biol. 2012, 212, 1–115. [Google Scholar] [CrossRef]
- Slominski, A.T.; Slominski, R.M.; Raman, C.; Chen, J.Y.; Athar, M.; Elmets, C. Neuroendocrine Signaling in the Skin with a Special Focus on the Epidermal Neuropeptides. Am. J. Physiol.-Cell Physiol. 2022, 323, C1757–C1776. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J. Neuroendocrinology of the Skin. Endocr. Rev. 2000, 21, 457–487. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.E.; di Nardo, A. Skin Neurogenic Inflammation. Semin. Immunopathol. 2018, 40, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Botchkarev, V.A.; Yaar, M.; Peters, E.M.J.; Raychaudhuri, S.P.; Botchkareva, N.V.; Marconi, A.; Raychaudhuri, S.K.; Paus, R.; Pincelli, C. Neurotrophins in Skin Biology and Pathology. J. Investig. Dermatol. 2006, 126, 1719–1727. [Google Scholar] [CrossRef]
- Roosterman, D.; Goerge, T.; Schneider, S.W.; Bunnett, N.W.; Steinhoff, M. Neuronal Control of Skin Function: The Skin as a Neuroimmunoendocrine Organ. Physiol. Rev. 2006, 86, 1309–1379. [Google Scholar] [CrossRef]
- Cevikbas, F.; Steinhoff, A.; Homey, B.; Steinhoff, M. Neuroimmune Interactions in Allergic Skin Diseases. Curr. Opin. Allergy Clin. Immunol. 2007, 7, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Liezmann, C.; Klapp, B.; Peters, E. Stress, Atopy and Allergy: A Re-Evaluation from a Psychoneuroimmunologic Persepective. Derm.-Endocrinol. 2011, 3, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.M.; Kim, C.W. The Effects of Substance P and Vasoactive Intestinal Peptide on Interleukin-6 Synthesis in Cultured Human Keratinocytes. J. Dermatol. Sci. 1999, 22, 17–23. [Google Scholar] [CrossRef]
- Burbach, G.J.; Kim, K.H.; Zivony, A.S.; Kim, A.; Aranda, J.; Wright, S.; Naik, S.M.; Caughman, S.W.; Ansel, J.C.; Armstrong, C.A. The Neurosensory Tachykinins Substance P and Neurokinin a Directly Induce Keratinocyte Nerve Growth Factor. J. Investig. Dermatol. 2001, 117, 1075–1082. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, C.; Wang, Z.J. Proteinase-Activated Receptor 2 Sensitizes Transient Receptor Potential Vanilloid 1, Transient Receptor Potential Vanilloid 4, and Transient Receptor Potential Ankyrin 1 in Paclitaxel-Induced Neuropathic Pain. Neuroscience 2011, 193, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, M.; Neisius, U.; Ikoma, A.; Fartasch, M.; Heyer, G.; Skov, P.S.; Luger, T.A.; Schmelz, M. Proteinase-Activated Receptor-2 Mediates Itch: A Novel Pathway for Pruritus in Human Skin. J. Neurosci. 2003, 23, 6176–6180. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Eberlin, S.; Polettini, A.J.; Pereira, A.F.d.C.; Pereira, C.S.; Ferreira, N.M.C.; Dolis, E.; Torloni, L.B.O. Neuromodulatory and Anti-Inflammatory Ingredient for Sensitive Skin: In Vitro Assessment. Inflamm. Allergy Drug Targets 2014, 13, 191–198. [Google Scholar] [CrossRef]
- Holmes, A.D.; Steinhoff, M. Integrative Concepts of Rosacea Pathophysiology, Clinical Presentation and New Therapeutics. Exp. Dermatol. 2017, 26, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, K.; Yu, Z.; Chai, J.; Zhang, Z.; Zhang, Y.; Min, P. Dramatic Effect of Botulinum Toxin Type A on Hypertrophic Scar: A Promising Therapeutic Drug and Its Mechanism Through the SP-NK1R Pathway in Cutaneous Neurogenic Inflammation. Front. Med. 2022, 9, 820817. [Google Scholar] [CrossRef]
- Vidal Yucha, S.E.; Tamamoto, K.A.; Kaplan, D.L. The Importance of the Neuro-immuno-cutaneous System on Human Skin Equivalent Design. Cell Prolif. 2019, 52, e12677. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, H.; Jiang, B.; Yan, S.; Lu, J. A Promising Therapeutic Target for Psoriasis: Neuropeptides in Human Skin. Int. Immunopharmacol. 2020, 87, 106755. [Google Scholar] [CrossRef]
- Mehta, D.; Granstein, R.D. Immunoregulatory Effects of Neuropeptides on Endothelial Cells: Relevance to Dermatological Disorders. Dermatology 2019, 235, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Talamantes, A.K.; Gómez-González, B.A.; Uriarte-Mayorga, D.F.; Martínez-Guzman, M.A.; Wheber-Hidalgo, K.A.; Alvarado-Navarro, A. Neurotransmitters, Neuropeptides and Their Receptors Interact with Immune Response in Healthy and Psoriatic Skin. Neuropeptides 2020, 79, 102004. [Google Scholar] [CrossRef]
- Graefe, S.B.; Mohiuddin, S.S. Biochemistry, Substance P; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- O’Connor, T.M.; O’Connell, J.; O’Brien, D.I.; Goode, T.; Bredin, C.P.; Shanahan, F. The Role of Substance P in Inflammatory Disease. J. Cell. Physiol. 2004, 201, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, M.S.; von Mentzer, B.; Geppetti, P.; Pothoulakis, C.; Bunnett, N.W. Tachykinins and Their Receptors: Contributions to Physiological Control and the Mechanisms of Disease. Physiol. Rev. 2014, 94, 265–301. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.J.; Dawbarn, D. Clinical Relevance of the Neurotrophins and Their Receptors. Clin. Sci. 2006, 110, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Theoharides, T.C.; Arck, P.C. Neuroimmunoendocrine Circuitry of the ‘Brain-Skin Connection’. Trends Immunol. 2006, 27, 32–39. [Google Scholar] [CrossRef]
- Kee, Z.; Kodji, X.; Brain, S.D. The Role of Calcitonin Gene Related Peptide (CGRP) in Neurogenic Vasodilation and Its Cardioprotective Effects. Front. Physiol. 2018, 9, 1249. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.R.; Brain, S.D. A Calcitonin Gene-Related Peptide (CGRP) Antagonist (CGRP8-37) Inhibits Microvascular Responses Induced by CGRP and Capsaicin in Skin. Br. J. Pharmacol. 1991, 104, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, M.; Ahmad, F.; Pandey, A.; Datsi, A.; AlHammadi, A.; Al-Khawaga, S.; Al-Malki, A.; Meng, J.; Alam, M.; Buddenkotte, J. Neuroimmune Communication Regulating Pruritus in Atopic Dermatitis. J. Allergy Clin. Immunol. 2022, 149, 1875–1898. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E. DAMPs, PAMPs and Alarmins: All We Need to Know about Danger. J. Leukoc. Biol. 2007, 81, 1–5. [Google Scholar] [CrossRef]
- Zindel, J.; Kubes, P. DAMPs, PAMPs, and LAMPs in Immunity and Sterile Inflammation. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 493–518. [Google Scholar] [CrossRef]
- Gong, T.; Liu, L.; Jiang, W.; Zhou, R. DAMP-Sensing Receptors in Sterile Inflammation and Inflammatory Diseases. Nat. Rev. Immunol. 2020, 20, 95–112. [Google Scholar] [CrossRef]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-ΚB Pathway for the Therapy of Diseases: Mechanism and Clinical Study. Signal Transduct. Target Ther. 2020, 5, 209. [Google Scholar] [CrossRef]
- Minnone, G.; de Benedetti, F.; Bracci-Laudiero, L. NGF and Its Receptors in the Regulation of Inflammatory Response. Int. J. Mol. Sci. 2017, 18, 1028. [Google Scholar] [CrossRef]
- Furue, M.; Yamamura, K.; Kido-Nakahara, M.; Nakahara, T.; Fukui, Y. Emerging Role of Interleukin-31 and Interleukin-31 Receptor in Pruritus in Atopic Dermatitis. Allergy 2018, 73, 29–36. [Google Scholar] [CrossRef]
- Jin, R.; Luo, L.; Zheng, J. The Trinity of Skin: Skin Homeostasis as a Neuro–Endocrine–Immune Organ. Life 2022, 12, 725. [Google Scholar] [CrossRef] [PubMed]
- Pennefather, J.N.; Lecci, A.; Candenas, M.L.; Patak, E.; Pinto, F.M.; Maggi, C.A. Tachykinins and Tachykinin Receptors: A Growing Family. Life Sci. 2004, 74, 1445–1463. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Recio, S.; Gascón, P. Biological and Pharmacological Aspects of the NK1-Receptor. BioMed Res. Int. 2015, 2015, 1–14. [Google Scholar] [CrossRef]
- Harding, S.D.; Sharman, J.L.; Faccenda, E.; Southan, C.; Pawson, A.J.; Ireland, S.; Gray, A.J.G.; Bruce, L.; Alexander, S.P.H.; Anderton, S.; et al. The IUPHAR/BPS Guide to Pharmacology in 2018: Updates and Expansion to Encompass the New Guide to Immunopharmacology. Nucleic Acids Res. 2018, 46, D1091–D1106. [Google Scholar] [CrossRef]
- Li, W.-W.; Guo, T.-Z.; Liang, D.; Sun, Y.; Kingery, W.S.; Clark, J.D. Substance P Signaling Controls Mast Cell Activation, Degranulation, and Nociceptive Sensitization in a Rat Fracture Model of Complex Regional Pain Syndrome. Anesthesiology 2012, 116, 882–895. [Google Scholar] [CrossRef]
- Hsin, L.; Fernandopulle, N.A.; Ding, J.; Lumb, C.; Veldhuis, N.; Karas, J.A.; Northfield, S.E.; Mackay, G.A. The Effect of Substance P and Its Common in Vivo-formed Metabolites on MRGPRX2 and Human Mast Cell Activation. Pharmacol. Res. Perspec. 2022, 10, e00990. [Google Scholar] [CrossRef] [PubMed]
- Perner, C.; Flayer, C.H.; Zhu, X.; Aderhold, P.A.; Dewan, Z.N.A.; Voisin, T.; Camire, R.B.; Chow, O.A.; Chiu, I.M.; Sokol, C.L. Substance P Release by Sensory Neurons Triggers Dendritic Cell Migration and Initiates the Type-2 Immune Response to Allergens. Immunity 2020, 53, 1063–1077.e7. [Google Scholar] [CrossRef]
- Theoharides, T.C. The Impact of Psychological Stress on Mast Cells. Ann. Allergy Asthma Immunol. 2020, 125, 388–392. [Google Scholar] [CrossRef]
- Kim, K.H.; Park, K.C.; Chung, J.H.; Choi, H.R. The Effect of Substance P on Peripheral Blood Mononuclear Cells in Patients with Atopic Dermatitis. J. Dermatol. Sci. 2003, 32, 115–124. [Google Scholar] [CrossRef]
- Siiskonen, H.; Harvima, I. Mast Cells and Sensory Nerves Contribute to Neurogenic Inflammation and Pruritus in Chronic Skin Inflammation. Front. Cell Neurosci. 2019, 13, 422. [Google Scholar] [CrossRef]
- Kim, Y.J.; Granstein, R.D. Roles of Calcitonin Gene-Related Peptide in the Skin, and Other Physiological and Pathophysiological Functions. Brain Behav. Immun.-Health 2021, 18, 100361. [Google Scholar] [CrossRef]
- Luger, T.A. Neuromediators—A Crucial Component of the Skin Immune System. J. Dermatol. Sci. 2002, 30, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Meixiong, J.; Anderson, M.; Limjunyawong, N.; Sabbagh, M.F.; Hu, E.; Mack, M.R.; Oetjen, L.K.; Wang, F.; Kim, B.S.; Dong, X. Activation of Mast-Cell-Expressed Mas-Related G-Protein-Coupled Receptors Drives Non-Histaminergic Itch. Immunity 2019, 50, 1163–1171.e5. [Google Scholar] [CrossRef]
- Liu, Q.; Weng, H.-J.; Patel, K.N.; Tang, Z.; Bai, H.; Steinhoff, M.; Dong, X. The Distinct Roles of Two GPCRs, MrgprC11 and PAR2, in Itch and Hyperalgesia. Sci. Signal. 2011, 4, ra45. [Google Scholar] [CrossRef] [PubMed]
- Tausk, F.; Elenkov, I.; Moynihan, J. Psychoneuroimmunology. Dermatol. Ther. 2008, 21, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y.; Nam, Y.J.; Kang, S.; Choi, E.J.; Han, I.; Kim, J.; Kim, D.H.; An, J.H.; Lee, S.; Lee, M.H.; et al. The Local Hypothalamic–Pituitary–Adrenal Axis in Cultured Human Dermal Papilla Cells. BMC Mol. Cell Biol. 2020, 21, 42. [Google Scholar] [CrossRef]
- Huang, J.; Polgár, E.; Solinski, H.J.; Mishra, S.K.; Tseng, P.-Y.; Iwagaki, N.; Boyle, K.A.; Dickie, A.C.; Kriegbaum, M.C.; Wildner, H.; et al. Circuit Dissection of the Role of Somatostatin in Itch and Pain. Nat. Neurosci. 2018, 21, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Lotti, T.; D’Erme, A.M.; Hercogová, J. The Role of Neuropeptides in the Control of Regional Immunity. Clin. Dermatol. 2014, 32, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Ruppenstein, A.; Limberg, M.M.; Loser, K.; Kremer, A.E.; Homey, B.; Raap, U. Involvement of Neuro-Immune Interactions in Pruritus with Special Focus on Receptor Expressions. Front. Med. 2021, 8, 627985. [Google Scholar] [CrossRef] [PubMed]
- Ständer, S.; Luger, T.A. NK-1 Antagonists and itch. In Pharmacology of Itch; Handbook of Experimental Pharmacology; Cowan, A., Yosipovitch, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; Volume 226, pp. 237–255. ISBN 978-3-662-44604-1. [Google Scholar]
- Pojawa-Gołąb, M.; Jaworecka, K.; Reich, A. NK-1 Receptor Antagonists and Pruritus: Review of Current Literature. Dermatol. Ther. 2019, 9, 391–405. [Google Scholar] [CrossRef]
- Yosipovitch, G.; Rosen, J.D.; Hashimoto, T. Itch: From Mechanism to (Novel) Therapeutic Approaches. J. Allergy Clin. Immunol. 2018, 142, 1375–1390. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, J.; Zhu, W.; Xu, C.; He, S. Upregulated Expression of Substance P in Basophils of the Patients with Chronic Spontaneous Urticaria: Induction of Histamine Release and Basophil Accumulation by Substance P. Cell Biol. Toxicol. 2016, 32, 217–228. [Google Scholar] [CrossRef]
- Zhang, Z.; Zheng, W.; Xie, H.; Chai, R.; Wang, J.; Zhang, H.; He, S. Up-Regulated Expression of Substance P in CD8+ T Cells and NK1R on Monocytes of Atopic Dermatitis. J. Transl. Med. 2017, 15, 93. [Google Scholar] [CrossRef] [PubMed]
- Oishi, N.; Iwata, H.; Kambe, N.; Kobayashi, N.; Fujimoto, K.; Sato, H.; Hisaka, A.; Ueno, K.; Yamaura, K. Expression of Precipitating Factors of Pruritus Found in Humans in an Imiquimod-Induced Psoriasis Mouse Model. Heliyon 2019, 5, e01981. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, H.; Huang, S. Role of NGF and Its Receptors in Wound Healing (Review). Exp. Ther. Med. 2021, 21, 599. [Google Scholar] [CrossRef]
- Datta-Mitra, A.; Kundu-Raychaudhuri, S.; Mitra, A.; Raychaudhuri, S.P. Cross Talk between Neuroregulatory Molecule and Monocyte: Nerve Growth Factor Activates the Inflammasome. PLoS ONE 2015, 10, e0121626. [Google Scholar] [CrossRef]
- Aarão, T.L.d.S.; de Sousa, J.R.; Falcão, A.S.C.; Falcão, L.F.M.; Quaresma, J.A.S. Nerve Growth Factor and Pathogenesis of Leprosy: Review and Update. Front. Immunol. 2018, 9, 939. [Google Scholar] [CrossRef] [PubMed]
- Conroy, J.N.; Coulson, E.J. High-Affinity TrkA and P75 Neurotrophin Receptor Complexes: A Twisted Affair. J. Biol. Chem. 2022, 298, 101568. [Google Scholar] [CrossRef]
- Gostynska, N.; Pannella, M.; Rocco, M.L.; Giardino, L.; Aloe, L.; Calzà, L. The Pleiotropic Molecule NGF Regulates the in Vitro Properties of Fibroblasts, Keratinocytes, and Endothelial Cells: Implications for Wound Healing. Am. J. Physiol.-Cell Physiol. 2020, 318, C360–C371. [Google Scholar] [CrossRef]
- Lorenzini, L.; Baldassarro, V.A.; Stanzani, A.; Giardino, L. Nerve growth factor: The first molecule of the neurotrophin family. In Recent Advances in NGF and Related Molecules: The Continuum of the NGF “Saga”; Advances in Experimental Medicine and, Biology; Calzà, L., Aloe, L., Giardino, L., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 3–10. ISBN 978-3-030-74046-7. [Google Scholar]
- Zhao, P.; Metcalf, M.; Bunnett, N.W. Biased Signaling of Protease-Activated Receptors. Front. Endocrinol. 2014, 5, 67. [Google Scholar] [CrossRef]
- Fu, Q.; Cheng, J.; Gao, Y.; Zhang, Y.; Chen, X.; Xie, J. Protease-Activated Receptor 4: A Critical Participator in Inflammatory Response. Inflammation 2015, 38, 886–895. [Google Scholar] [CrossRef]
- Solinski, H.J.; Gudermann, T.; Breit, A. Pharmacology and Signaling of MAS-Related G Protein–Coupled Receptors. Pharmacol. Rev. 2014, 66, 570–597. [Google Scholar] [CrossRef] [PubMed]
- Bader, M.; Alenina, N.; Andrade-Navarro, M.A.; Santos, R.A. Mas and Its Related G Protein–Coupled Receptors, Mrgprs. Pharmacol. Rev. 2014, 66, 1080–1105. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Kang, H.J.; Singh, I.; Chen, H.; Zhang, C.; Ye, W.; Hayes, B.W.; Liu, J.; Gumpper, R.H.; Bender, B.J.; et al. Structure, Function and Pharmacology of Human Itch GPCRs. Nature 2021, 600, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Tang, Z.; Surdenikova, L.; Kim, S.; Patel, K.N.; Kim, A.; Ru, F.; Guan, Y.; Weng, H.-J.; Geng, Y.; et al. Sensory Neuron-Specific GPCR Mrgprs Are Itch Receptors Mediating Chloroquine-Induced Pruritus. Cell 2009, 139, 1353–1365. [Google Scholar] [CrossRef]
- Wilson, S.R.; Gerhold, K.A.; Bifolck-Fisher, A.; Liu, Q.; Patel, K.N.; Dong, X.; Bautista, D.M. TRPA1 Is Required for Histamine-Independent, Mas-Related G Protein–Coupled Receptor–Mediated Itch. Nat. Neurosci. 2011, 14, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Chen, C.; Zhou, W.; Ma, X.; Pu, X.; Zeng, Y.; Zhou, W.; Lv, F. TRPA1 Deficiency Alleviates Inflammation of Atopic Dermatitis by Reducing Macrophage Infiltration. Life Sci. 2021, 266, 118906. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Escalera, J.; Balakrishna, S.; Fan, L.; Caceres, A.I.; Robinson, E.; Sui, A.; McKay, M.C.; McAlexander, M.A.; Herrick, C.A.; et al. TRPA1 Controls Inflammation and Pruritogen Responses in Allergic Contact Dermatitis. FASEB J. 2013, 27, 3549–3563. [Google Scholar] [CrossRef]
- Weidinger, S.; Beck, L.A.; Bieber, T.; Kabashima, K.; Irvine, A.D. Atopic Dermatitis. Nat. Rev. Dis. Prim. 2018, 4, 1. [Google Scholar] [CrossRef]
- Greb, J.E.; Goldminz, A.M.; Elder, J.T.; Lebwohl, M.G.; Gladman, D.D.; Wu, J.J.; Mehta, N.N.; Finlay, A.Y.; Gottlieb, A.B. Psoriasis. Nat. Rev. Dis. Prim. 2016, 2, 16082. [Google Scholar] [CrossRef] [PubMed]
- Komiya, E.; Tominaga, M.; Kamata, Y.; Suga, Y.; Takamori, K. Molecular and Cellular Mechanisms of Itch in Psoriasis. Int. J. Mol. Sci. 2020, 21, 8406. [Google Scholar] [CrossRef] [PubMed]
- Silverman, H.A.; Chen, A.; Kravatz, N.L.; Chavan, S.S.; Chang, E.H. Involvement of Neural Transient Receptor Potential Channels in Peripheral Inflammation. Front. Immunol. 2020, 11, 590261. [Google Scholar] [CrossRef] [PubMed]
- Duitama, M.; Vargas-López, V.; Casas, Z.; Albarracin, S.L.; Sutachan, J.-J.; Torres, Y.P. TRP Channels Role in Pain Associated with Neurodegenerative Diseases. Front. Neurosci. 2020, 14, 782. [Google Scholar] [CrossRef] [PubMed]
- Boillat, A.; Alijevic, O.; Kellenberger, S. Calcium Entry via TRPV1 but Not ASICs Induces Neuropeptide Release from Sensory Neurons. Mol. Cell. Neurosci. 2014, 61, 13–22. [Google Scholar] [CrossRef]
- Gouin, O.; L’Herondelle, K.; Lebonvallet, N.; Le Gall-Ianotto, C.; Sakka, M.; Buhé, V.; Plée-Gautier, E.; Carré, J.-L.; Lefeuvre, L.; Misery, L.; et al. TRPV1 and TRPA1 in Cutaneous Neurogenic and Chronic Inflammation: Pro-Inflammatory Response Induced by Their Activation and Their Sensitization. Protein Cell 2017, 8, 644–661. [Google Scholar] [CrossRef]
- Meng, J.; Li, Y.; Fischer, M.J.M.; Steinhoff, M.; Chen, W.; Wang, J. Th2 Modulation of Transient Receptor Potential Channels: An Unmet Therapeutic Intervention for Atopic Dermatitis. Front. Immunol. 2021, 12, 696784. [Google Scholar] [CrossRef]
- Jain, A.; Brönneke, S.; Kolbe, L.; Stäb, F.; Wenck, H.; Neufang, G. TRP-Channel-Specific Cutaneous Eicosanoid Release Patterns. Pain 2011, 152, 2765–2772. [Google Scholar] [CrossRef]
- Landini, L.; Souza Monteiro de Araujo, D.; Titiz, M.; Geppetti, P.; Nassini, R.; de Logu, F. TRPA1 Role in Inflammatory Disorders: What Is Known So Far? Int. J. Mol. Sci. 2022, 23, 4529. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.R.; Thé, L.; Batia, L.M.; Beattie, K.; Katibah, G.E.; McClain, S.P.; Pellegrino, M.; Estandian, D.M.; Bautista, D.M. The Epithelial Cell-Derived Atopic Dermatitis Cytokine TSLP Activates Neurons to Induce Itch. Cell 2013, 155, 285–295. [Google Scholar] [CrossRef]
- Lamas, J.A.; Rueda-Ruzafa, L.; Herrera-Pérez, S. Ion Channels and Thermosensitivity: TRP, TREK, or Both? Int. J. Mol. Sci. 2019, 20, 2371. [Google Scholar] [CrossRef] [PubMed]
- Giniatullin, R. Ion Channels of Nociception. Int. J. Mol. Sci. 2020, 21, 3553. [Google Scholar] [CrossRef] [PubMed]
- Shirolkar, P.; Mishra, S.K. Role of TRP Ion Channels in Pruritus. Neurosci. Lett. 2022, 768, 136379. [Google Scholar] [CrossRef]
- Lebonvallet, N.; Fluhr, J.W.; le Gall-Ianotto, C.; Leschiera, R.; Talagas, M.; Reux, A.; Bataille, A.; Brun, C.; Oddos, T.; Pennec, J.-P.; et al. A Re-innervated in Vitro Skin Model of Non-histaminergic Itch and Skin Neurogenic Inflammation: PAR2-, TRPV1- and TRPA1-agonist Induced Functionality. Ski. Health Dis. 2021, 1, e66. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.-Y.; Tan, C.-H. TRP channels in nociception and pathological pain. In Advances in Pain Research: Mechanisms and Modulation of Chronic Pain; Advances in Experimental Medicine and, Biology; Shyu, B.-C., Tominaga, M., Eds.; Springer: Singapore, 2018; Volume 1099, pp. 13–27. ISBN 9789811317552. [Google Scholar]
- Mollanazar, N.K.; Smith, P.K.; Yosipovitch, G. Mediators of Chronic Pruritus in Atopic Dermatitis: Getting the Itch Out? Clinic Rev. Allerg. Immunol. 2016, 51, 263–292. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Munanairi, A.; Liu, X.-Y.; Zhang, J.; Hu, L.; Hu, M.; Bu, D.; Liu, L.; Xie, Z.; Kim, B.S.; et al. PAR2 Mediates Itch via TRPV3 Signaling in Keratinocytes. J. Investig. Dermatol. 2020, 140, 1524–1532. [Google Scholar] [CrossRef]
- Buhl, T.; Ikoma, A.; Kempkes, C.; Cevikbas, F.; Sulk, M.; Buddenkotte, J.; Akiyama, T.; Crumrine, D.; Camerer, E.; Carstens, E.; et al. Protease-Activated Receptor-2 Regulates Neuro-Epidermal Communication in Atopic Dermatitis. Front. Immunol. 2020, 11, 1740. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.P.; Amadesi, S.; Veldhuis, N.A.; Abogadie, F.C.; Lieu, T.; Darby, W.; Liedtke, W.; Lew, M.J.; McIntyre, P.; Bunnett, N.W. Protease-Activated Receptor 2 (PAR2) Protein and Transient Receptor Potential Vanilloid 4 (TRPV4) Protein Coupling Is Required for Sustained Inflammatory Signaling*. J. Biol. Chem. 2013, 288, 5790–5802. [Google Scholar] [CrossRef] [PubMed]
- Buddenkotte, J.; Stroh, C.; Engels, I.H.; Moormann, C.; Shpacovitch, V.M.; Seeliger, S.; Vergnolle, N.; Vestweber, D.; Luger, T.A.; Schulze-Osthoff, K.; et al. Agonists of Proteinase-Activated Receptor-2 Stimulate Upregulation of Intercellular Cell Adhesion Molecule-1 in Primary Human Keratinocytes via Activation of NF-Kappa B. J. Investig. Dermatol. 2005, 124, 38–45. [Google Scholar] [CrossRef]
- Kwatra, S.G.; Misery, L.; Clibborn, C.; Steinhoff, M. Molecular and Cellular Mechanisms of Itch and Pain in Atopic Dermatitis and Implications for Novel Therapeutics. Clin. Trans. Immunol. 2022, 11, e1390. [Google Scholar] [CrossRef]
- Maglie, R.; Souza Monteiro de Araujo, D.; Antiga, E.; Geppetti, P.; Nassini, R.; de Logu, F. The Role of TRPA1 in Skin Physiology and Pathology. Int. J. Mol. Sci. 2021, 22, 3065. [Google Scholar] [CrossRef] [PubMed]
- Moon, T.C.; St Laurent, C.D.; Morris, K.E.; Marcet, C.; Yoshimura, T.; Sekar, Y.; Befus, A.D. Advances in Mast Cell Biology: New Understanding of Heterogeneity and Function. Mucosal. Immunol. 2010, 3, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, E.; van Bergeijk, D.; Oosting, R.S.; Redegeld, F.A. Mast Cells in Neuroinflammation and Brain Disorders. Neurosci. Biobehav. Rev. 2017, 79, 119–133. [Google Scholar] [CrossRef]
- Silver, R.; Curley, J.P. Mast Cells on the Mind: New Insights and Opportunities. Trends Neurosci. 2013, 36, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Harvima, I.T. Mast Cell-Neural Interactions Contribute to Pain and Itch. Immunol. Rev. 2018, 282, 168–187. [Google Scholar] [CrossRef]
- Mittal, A.; Sagi, V.; Gupta, M.; Gupta, K. Mast Cell Neural Interactions in Health and Disease. Front. Cell Neurosci. 2019, 13, 110. [Google Scholar] [CrossRef]
- Kissel, C.L.; Kovács, K.J.; Larson, A.A. Evidence for the Modulation of Nociception in Mice by Central Mast Cells. Eur. J. Pain 2017, 21, 1743–1755. [Google Scholar] [CrossRef]
- Esposito, P.; Gheorghe, D.; Kandere, K.; Pang, X.; Connolly, R.; Jacobson, S.; Theoharides, T.C. Acute Stress Increases Permeability of the Blood–Brain-Barrier through Activation of Brain Mast Cells. Brain Res. 2001, 888, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Kempuraj, D.; Selvakumar, G.P.; Thangavel, R.; Ahmed, M.E.; Zaheer, S.; Raikwar, S.P.; Iyer, S.S.; Bhagavan, S.M.; Beladakere-Ramaswamy, S.; Zaheer, A. Mast Cell Activation in Brain Injury, Stress, and Post-Traumatic Stress Disorder and Alzheimer’s Disease Pathogenesis. Front. Neurosci. 2017, 11, 703. [Google Scholar] [CrossRef] [PubMed]
- Aich, A.; Afrin, L.; Gupta, K. Mast Cell-Mediated Mechanisms of Nociception. Int. J. Mol. Sci. 2015, 16, 29069–29092. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Rambasek, T.; Wöhrl, S. Mastocytosis: From a Molecular Point of View. Clin. Rev. Allerg. Immunol. 2018, 54, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Hoermann, G.; Sotlar, K.; Jawhar, M.; Kristensen, T.; Bachelot, G.; Nedoszytko, B.; Carter, M.C.; Horny, H.-P.; Bonadonna, P.; Sperr, W.R.; et al. Standards of Genetic Testing in the Diagnosis and Prognostication of Systemic Mastocytosis in 2022: Recommendations of the EU-US Cooperative Group. J. Allergy Clin. Immunol. Pract. 2022, 10, 1953–1963. [Google Scholar] [CrossRef] [PubMed]
- Pyatilova, P.; Akin, C.; Alvarez-Twose, I.; Arock, M.; Bonadonna, P.; Brockow, K.; Butterfield, J.H.; Broesby-Olsen, S.; Carter, M.C.; Castells, M.; et al. Refined Treatment Response Criteria for Indolent Systemic Mastocytosis Proposed by the ECNM-AIM Consortium. J. Allergy Clin. Immunol. Pract. 2022, 10, 2015–2024. [Google Scholar] [CrossRef] [PubMed]
- Valent, P.; Hartmann, K.; Bonadonna, P.; Gülen, T.; Brockow, K.; Alvarez-Twose, I.; Hermine, O.; Niedoszytko, M.; Carter, M.C.; Hoermann, G.; et al. Global Classification of Mast Cell Activation Disorders: An ICD-10-CM–Adjusted Proposal of the ECNM-AIM Consortium. J. Allergy Clin. Immunol. Pract. 2022, 10, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Romantowski, J.; Górska, A.; Niedoszytko, M.; Gulen, T.; Gruchała-Niedoszytko, M.; Nedoszytko, B.; Lange, M.; Brockow, K.; Arock, M.; Akin, C.; et al. A Challenge for Allergologist: Application of Allergy Diagnostic Methods in Mast Cell Disorders. Int. J. Mol. Sci. 2021, 22, 1454. [Google Scholar] [CrossRef]
- Nedoszytko, B.; Arock, M.; Lyons, J.; Bachelot, G.; Schwartz, L.; Reiter, A.; Jawhar, M.; Schwaab, J.; Lange, M.; Greiner, G.; et al. Clinical Impact of Inherited and Acquired Genetic Variants in Mastocytosis. Int. J. Mol. Sci. 2021, 22, 411. [Google Scholar] [CrossRef]
- Lyons, J.J.; Yi, T. Mast Cell Tryptases in Allergic Inflammation and Immediate Hypersensitivity. Curr. Opin. Immunol. 2021, 72, 94–106. [Google Scholar] [CrossRef]
- Shamji, M.H.; Durham, S.R. Mechanisms of Allergen Immunotherapy for Inhaled Allergens and Predictive Biomarkers. J. Allergy Clin. Immunol. 2017, 140, 1485–1498. [Google Scholar] [CrossRef]
- Modena, B.D.; Dazy, K.; White, A.A. Emerging Concepts: Mast Cell Involvement in Allergic Diseases. Transl. Res. 2016, 174, 98–121. [Google Scholar] [CrossRef]
- Redhu, D.; Franke, K.; Aparicio-Soto, M.; Kumari, V.; Pazur, K.; Illerhaus, A.; Hartmann, K.; Worm, M.; Babina, M. Mast Cells Instruct Keratinocytes to Produce Thymic Stromal Lymphopoietin: Relevance of the Tryptase/Protease-Activated Receptor 2 Axis. J. Allergy Clin. Immunol. 2022, 149, 2053–2061.e6. [Google Scholar] [CrossRef]
- Elieh Ali Komi, D.; Wöhrl, S.; Bielory, L. Mast Cell Biology at Molecular Level: A Comprehensive Review. Clin. Rev. Allerg. Immunol. 2020, 58, 342–365. [Google Scholar] [CrossRef]
- Gilfillan, A.M.; Beaven, M.A. Regulation of Mast Cell Responses in Health and Disease. Crit. Rev. Immunol. 2011, 31, 475–530. [Google Scholar] [CrossRef]
- McNeil, B.D.; Pundir, P.; Meeker, S.; Han, L.; Undem, B.J.; Kulka, M.; Dong, X. Identification of a Mast-Cell-Specific Receptor Crucial for Pseudo-Allergic Drug Reactions. Nature 2015, 519, 237–241. [Google Scholar] [CrossRef]
- Gupta, M.A.; Gupta, A.K. Psychiatric and Psychological Co-Morbidity in Patients with Dermatologic Disorders: Epidemiology and Management. Am. J. Clin. Dermatol. 2003, 4, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Zbytek, B.; Tobin, D.J.; Theoharides, T.C.; Rivier, J. Key Role of CRF in the Skin Stress Response System. Endocr. Rev. 2013, 34, 827–884. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Wortsman, J.; Luger, T.; Paus, R.; Solomon, S. Corticotropin Releasing Hormone and Proopiomelanocortin Involvement in the Cutaneous Response to Stress. Physiol. Rev. 2000, 80, 979–1020. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV Light Touches the Brain and Endocrine System Through Skin, and Why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef]
- Troubat, R.; Barone, P.; Leman, S.; Desmidt, T.; Cressant, A.; Atanasova, B.; Brizard, B.; El Hage, W.; Surget, A.; Belzung, C.; et al. Neuroinflammation and Depression: A Review. Eur. J. Neurosci. 2021, 53, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Grigore, O.; Mihailescu, A.; Solomon, I.; Boda, D.; Caruntu, C. Role of Stress in Modulation of Skin Neurogenic Inflammation. Exp. Ther. Med. 2018, 997–1003. [Google Scholar] [CrossRef]
- Bauer, M.E.; Teixeira, A.L. Neuroinflammation in Mood Disorders: Role of Regulatory Immune Cells. Neuroimmunomodulation 2021, 28, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P. Stress and Disorders of the Stress System. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Wortsman, J.; Tuckey, R.C.; Paus, R. Differential Expression of HPA Axis Homolog in the Skin. Mol. Cell. Endocrinol. 2007, 265–266, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A. A Nervous Breakdown in the Skin: Stress and the Epidermal Barrier. J. Clin. Investig. 2007, 117, 3166–3169. [Google Scholar] [CrossRef] [PubMed]
- Spencer, R.L.; Deak, T. A Users Guide to HPA Axis Research. Physiol. Behav. 2017, 178, 43–65. [Google Scholar] [CrossRef] [PubMed]
- Pariante, C.M. Why Are Depressed Patients Inflamed? A Reflection on 20 Years of Research on Depression, Glucocorticoid Resistance and Inflammation. Eur. Neuropsychopharmacol. 2017, 27, 554–559. [Google Scholar] [CrossRef]
- Cruz-Topete, D.; Cidlowski, J.A. One Hormone, Two Actions: Anti- and Pro-Inflammatory Effects of Glucocorticoids. Neuroimmunomodulation 2015, 22, 20–32. [Google Scholar] [CrossRef]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin Pigmentation in Mammalian Skin and Its Hormonal Regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef]
- Slominski, A.T.; Manna, P.R.; Tuckey, R.C. Cutaneous Glucocorticosteroidogenesis: Securing Local Homeostasis and the Skin Integrity. Exp. Dermatol. 2014, 23, 369–374. [Google Scholar] [CrossRef]
- Hannen, R.F.; Michael, A.E.; Jaulim, A.; Bhogal, R.; Burrin, J.M.; Philpott, M.P. Steroid Synthesis by Primary Human Keratinocytes; Implications for Skin Disease. Biochem. Biophys. Res. Commun. 2011, 404, 62–67. [Google Scholar] [CrossRef]
- Slominski, R.M.; Raman, C.; Elmets, C.; Jetten, A.M.; Slominski, A.T.; Tuckey, R.C. The Significance of CYP11A1 Expression in Skin Physiology and Pathology. Mol. Cell. Endocrinol. 2021, 530, 111238. [Google Scholar] [CrossRef]
- Slominski, A.; Zjawiony, J.; Wortsman, J.; Semak, I.; Stewart, J.; Pisarchik, A.; Sweatman, T.; Marcos, J.; Dunbar, C.; Tuckey, R.C. A Novel Pathway for Sequential Transformation of 7-Dehydrocholesterol and Expression of the P450scc System in Mammalian Skin: P450scc in the Skin. Eur. J. Biochem. 2004, 271, 4178–4188. [Google Scholar] [CrossRef]
- Slominski, A.; Zbytek, B.; Szczesniewski, A.; Semak, I.; Kaminski, J.; Sweatman, T.; Wortsman, J. CRH Stimulation of Corticosteroids Production in Melanocytes Is Mediated by ACTH. Am. J. Physiol.-Endocrinol. Metab. 2005, 288, E701–E706. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Zbytek, B.; Semak, I.; Sweatman, T.; Wortsman, J. CRH Stimulates POMC Activity and Corticosterone Production in Dermal Fibroblasts. J. Neuroimmunol. 2005, 162, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Slominski, R.M.; Tuckey, R.C.; Manna, P.R.; Jetten, A.M.; Postlethwaite, A.; Raman, C.; Slominski, A.T. Extra-Adrenal Glucocorticoid Biosynthesis: Implications for Autoimmune and Inflammatory Disorders. Genes Immun. 2020, 21, 150–168. [Google Scholar] [CrossRef]
- Slominski, A.; Zbytek, B.; Szczesniewski, A.; Wortsman, J. Cultured Human Dermal Fibroblasts Do Produce Cortisol. J. Investig. Dermatol. 2006, 126, 1177–1178. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Stewart, J.M.; Panagiotidou, S.; Melamed, I. Mast Cells, Brain Inflammation and Autism. Eur. J. Pharmacol. 2016, 778, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Kempuraj, D.; Mentor, S.; Thangavel, R.; Ahmed, M.E.; Selvakumar, G.P.; Raikwar, S.P.; Dubova, I.; Zaheer, S.; Iyer, S.S.; Zaheer, A. Mast Cells in Stress, Pain, Blood-Brain Barrier, Neuroinflammation and Alzheimer’s Disease. Front. Cell. Neurosci. 2019, 13, 54. [Google Scholar] [CrossRef]
- Conti, P.; Caraffa, A.; Ronconi, G.; Conti, C.M.; Kritas, S.K.; Mastrangelo, F.; Tettamanti, L.; Theoharides, T.C. Impact of Mast Cells in Depression Disorder: Inhibitory Effect of IL-37 (New Frontiers). Immunol. Res. 2018, 66, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D.; Facci, L.; Kee, W.J.; Strijbos, P.J.L.M. Potentiation by Histamine of Synaptically Mediated Excitotoxicity in Cultured Hippocampal Neurones: A Possible Role for Mast Cells: Histamine, Synaptic Activity and Neuronal Death. J. Neurochem. 2008, 76, 47–55. [Google Scholar] [CrossRef]
- Woźniak, E.; Owczarczyk-Saczonek, A.; Placek, W. Psychological Stress, Mast Cells, and Psoriasis—Is There Any Relationship? Int. J. Mol. Sci. 2021, 22, 13252. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Neuropeptide/ Mediator | Sources/Expressed by | Cell types Expressing the Receptor | Overall Function in Skin | Association with Skin Diseases | Ref. |
|---|---|---|---|---|---|
| SP (substance P) | Secreted by sensory C fibers and DRG - Endothelial cells, keratinocytes and MCs | - NK-1R (neurokinin-1 receptors) - Endothelial cells, connective tissue mast cells producing tryptase and chymase, fibroblasts and Langerhans cells | - Initiates the inflammatory response, leading to proliferation of specific T-lymphocytes, as well as the activation and degranulation of mast cells in early stages of psoriasis - Vasodilatation - Local inflammation - Increased cellular proliferation - Skin HPA axis activation | - Psoriasis - Rosacea - Atopic dermatitis - High concentration is observed in the blood of patients, which correlates with the activity of the disease process and the intensity of itching | [4,17,18,19,20,21,22,35,36] |
| NGF (nerve growth factor) neurotrophin | Secreted by cutaneous cells (keratinocytes, fibroblasts and adipocytes nerves) | NGF-TrkA keratinocytes, MCs, fibroblasts and eosinophils | - Correlated with the intensity of pruritus - Modulates nerve innervation and neuropeptide release, degranulates mast cells and induces keratinocyte hyperproliferation - Responsible for proliferation and growth of nerve cells - Favors the survival of certain immune cells in the cutaneous system | - Psoriasis - Atopic dermatitis - Allergic diseases - Prurigo nodularis | [6,16,23,24] |
| CGRP (calcitonin gene-related peptide) | - One of the main peptides involved in neurogenic inflammation - Is released after the activation of TRPV1 or TRPA1 - Secreted by sensory neurons | CGRP receptors and mast cell surface | - Induces vasodilatation - Can activate eosinophils to release proinflammatory mediators - Stimulates the proliferation of keratinocytes - Is responsible for the appearance of erythema | - Psoriasis - Prurigo nodularis - Atopic dermatitis - Rosacea | [18,25,26,32,33,34,44] |
| VIP (vasoactive intestinal peptide) | Secreted by: - Sensory and autonomic neurons - Keratinocytes, endothelial cells and T lymphocytes | Membrane receptors VPAC1 and 2R coupled with G protein are found on keratinocytes, T lymphocytes and mast cells | - MC degranulation and the production of proinflammatory cytokines - Induces vasodilatation | - Eczema - Psoriasis - Atopic dermatitis | [18,43,45] |
| CRF (corticotropin- releasing factor) the central neuropeptide of the HPA axis | - Keratinocytes, melanocytes, fibroblasts and mast cells | - CRF1 receptors are found on the cells of the epidermis (keratinocytes and melanocytes) and the dermis (fibroblasts and mast cells) - CRF2 receptors are found on the cells of skin appendages (hair follicles, sebaceous and sweat glands) | - Stimulates the production and release of MCs, keratinocytes and fibroblasts of cytokines with a Th2 profile | - Psoriasis - Acne - Alopecia areata - Atopic dermatitis - Vitiligo - Lichen planus - Seborrheic dermatitis - Rosacea - Urticaria | [46,47,48,49] |
| Substances that inhibit the neurogenic inflammatory process | |||||
| SST (somatostatin) | - Receptors (SSTR2) are found on T lymphocytes, Langerhans cells, epithelium cells and fibroblasts | - Sensory neurons - Merkel cells - Langerhans cells - Sweat gland cells | - Inhibits the secretion of pro-inflammatory cytokines - Inhibits the secretion endocrine glands - Inhibits intracellular cAMP | [18,19,50] | |
| (α-MSH)— α-melanocyte stimulating hormone | MC1R−MC5R melanocytes, keratinocytes, fibroblasts and immune cells | - Is formed on the skin by the POMC prohormone - Melanocytes, keratinocytes, MCs, macrophages and monocytes | - Inhibits the synthesis of cytokines with anti-inflammatory properties | [18,51] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marek-Jozefowicz, L.; Nedoszytko, B.; Grochocka, M.; Żmijewski, M.A.; Czajkowski, R.; Cubała, W.J.; Slominski, A.T. Molecular Mechanisms of Neurogenic Inflammation of the Skin. Int. J. Mol. Sci. 2023, 24, 5001. https://doi.org/10.3390/ijms24055001
Marek-Jozefowicz L, Nedoszytko B, Grochocka M, Żmijewski MA, Czajkowski R, Cubała WJ, Slominski AT. Molecular Mechanisms of Neurogenic Inflammation of the Skin. International Journal of Molecular Sciences. 2023; 24(5):5001. https://doi.org/10.3390/ijms24055001
Chicago/Turabian StyleMarek-Jozefowicz, Luiza, Bogusław Nedoszytko, Małgorzata Grochocka, Michał A. Żmijewski, Rafał Czajkowski, Wiesław J. Cubała, and Andrzej T. Slominski. 2023. "Molecular Mechanisms of Neurogenic Inflammation of the Skin" International Journal of Molecular Sciences 24, no. 5: 5001. https://doi.org/10.3390/ijms24055001
APA StyleMarek-Jozefowicz, L., Nedoszytko, B., Grochocka, M., Żmijewski, M. A., Czajkowski, R., Cubała, W. J., & Slominski, A. T. (2023). Molecular Mechanisms of Neurogenic Inflammation of the Skin. International Journal of Molecular Sciences, 24(5), 5001. https://doi.org/10.3390/ijms24055001