
Composite Coatings Based on Recombinant Spidroins and Peptides with Motifs of the Extracellular Matrix Proteins Enhance Neuronal Differentiation of Neural Precursor Cells Derived from Human Induced Pluripotent Stem Cells
 , and
, and
Abstract
1. Introduction
2. Results
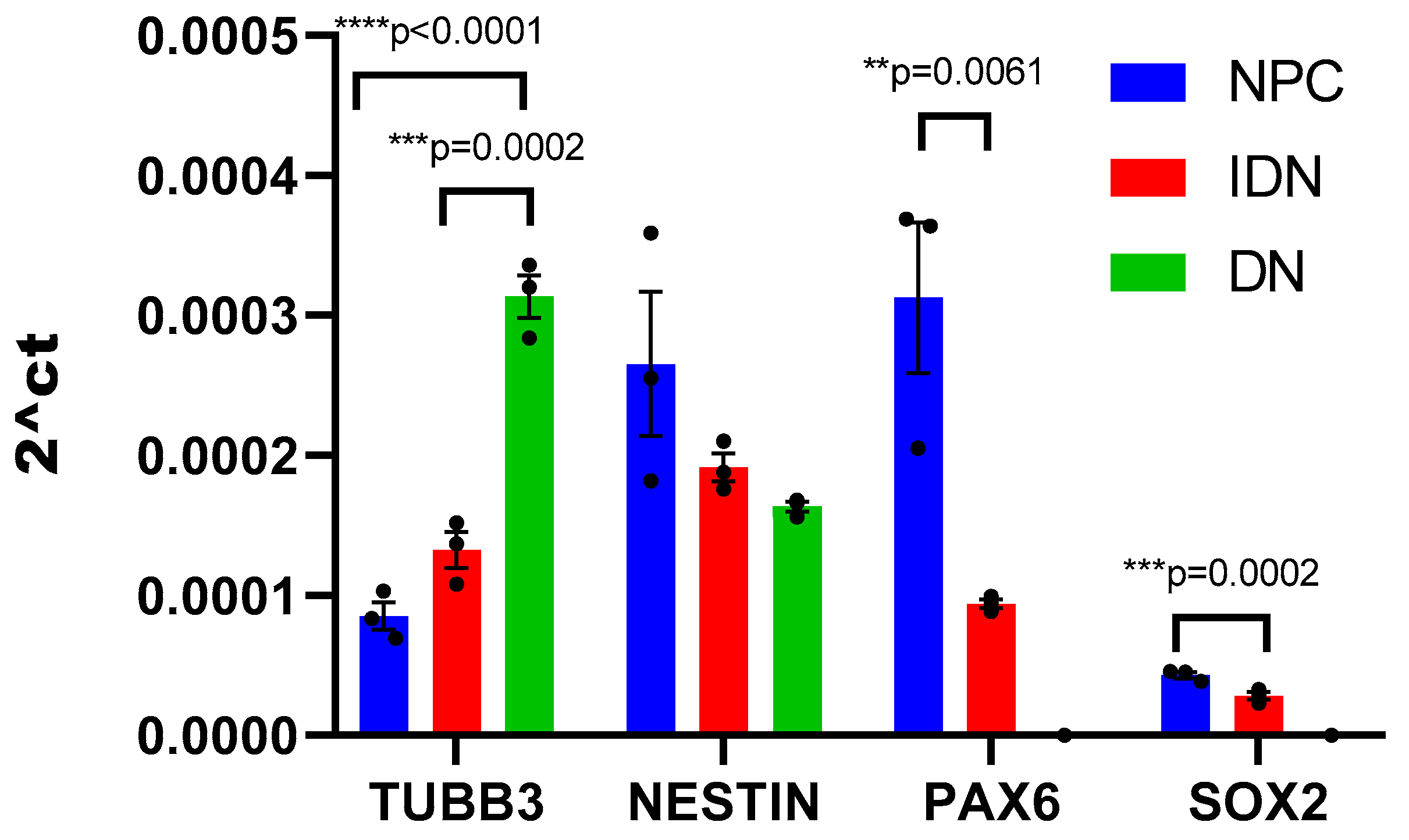
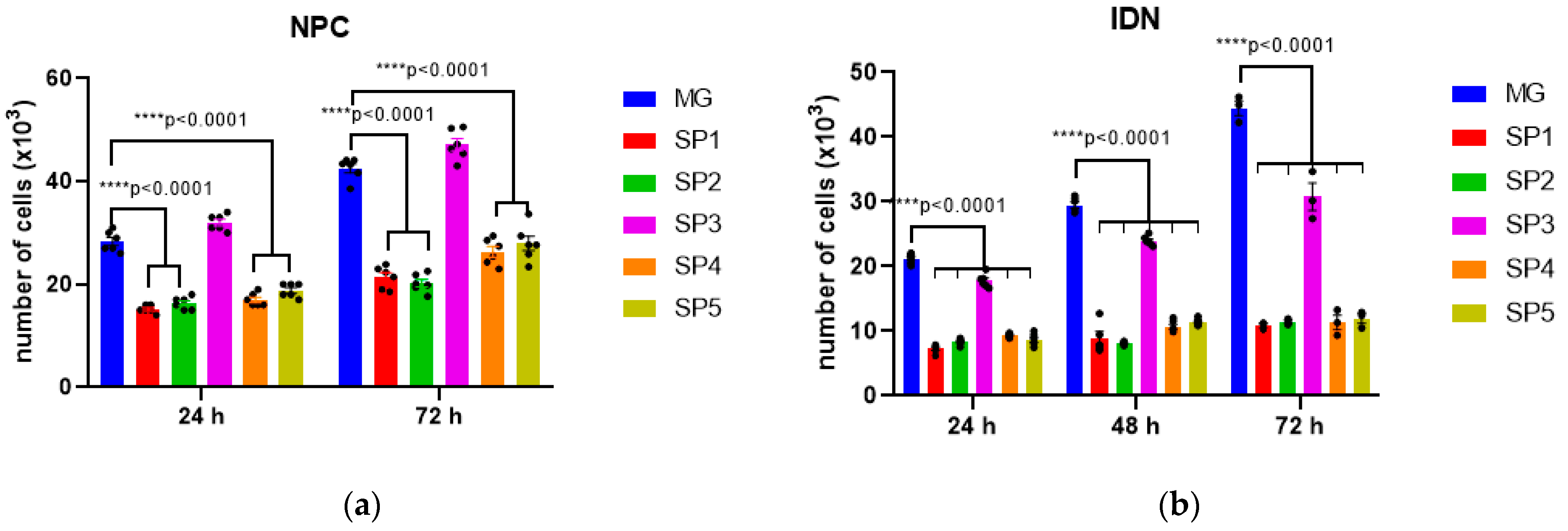
2.1. The Influence of Different CC Variants on the Proliferative Activity of NPCs Differentiating from iPSCs
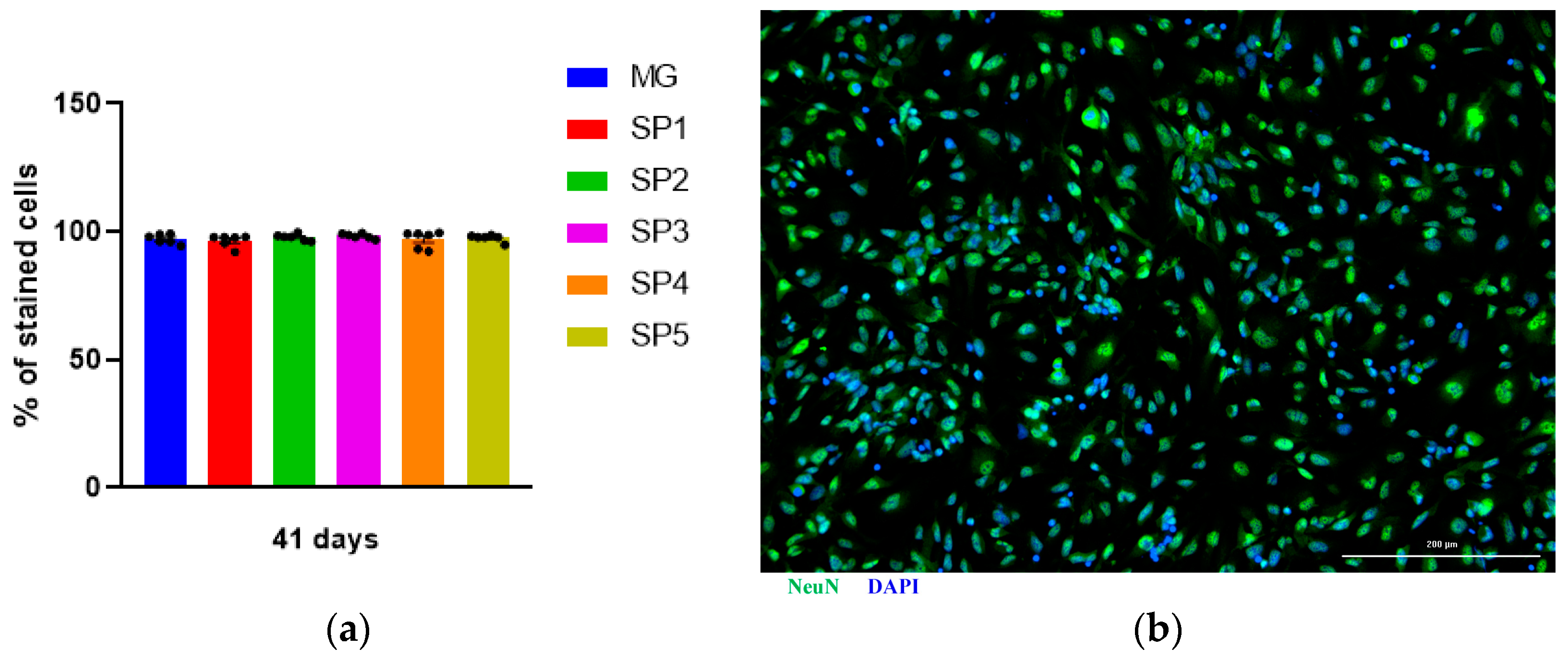
2.2. Study of the Effect of Cultivation on CCs on the Differentiation of NPS into IDNs
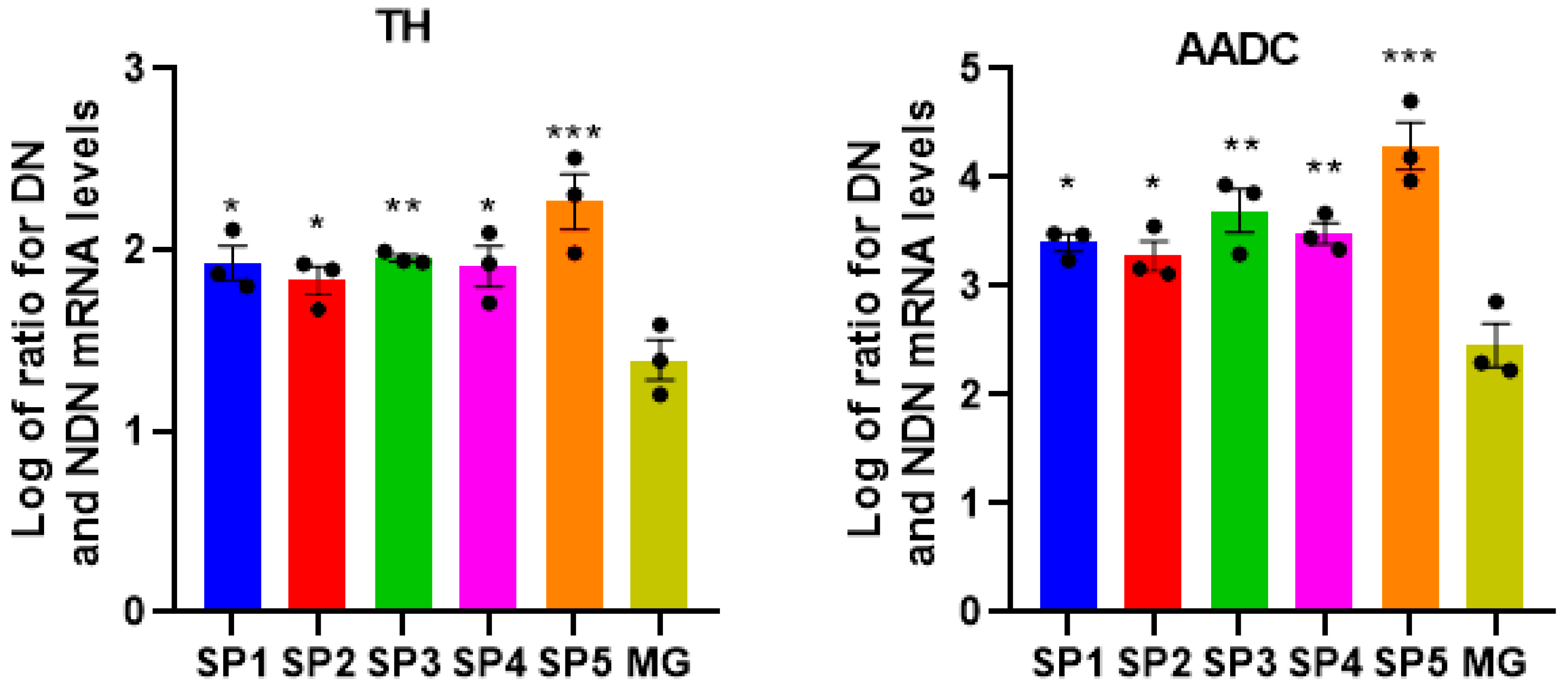
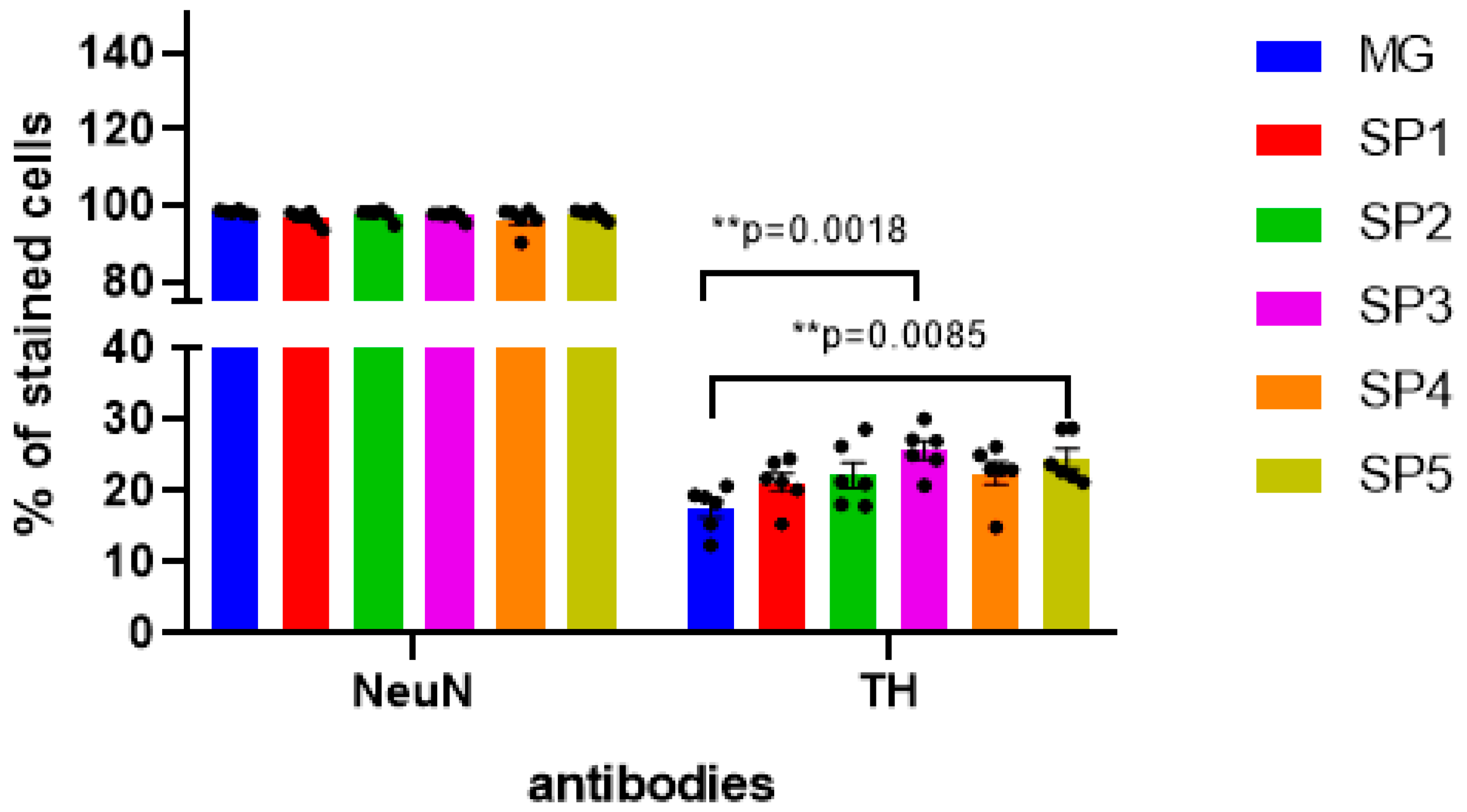
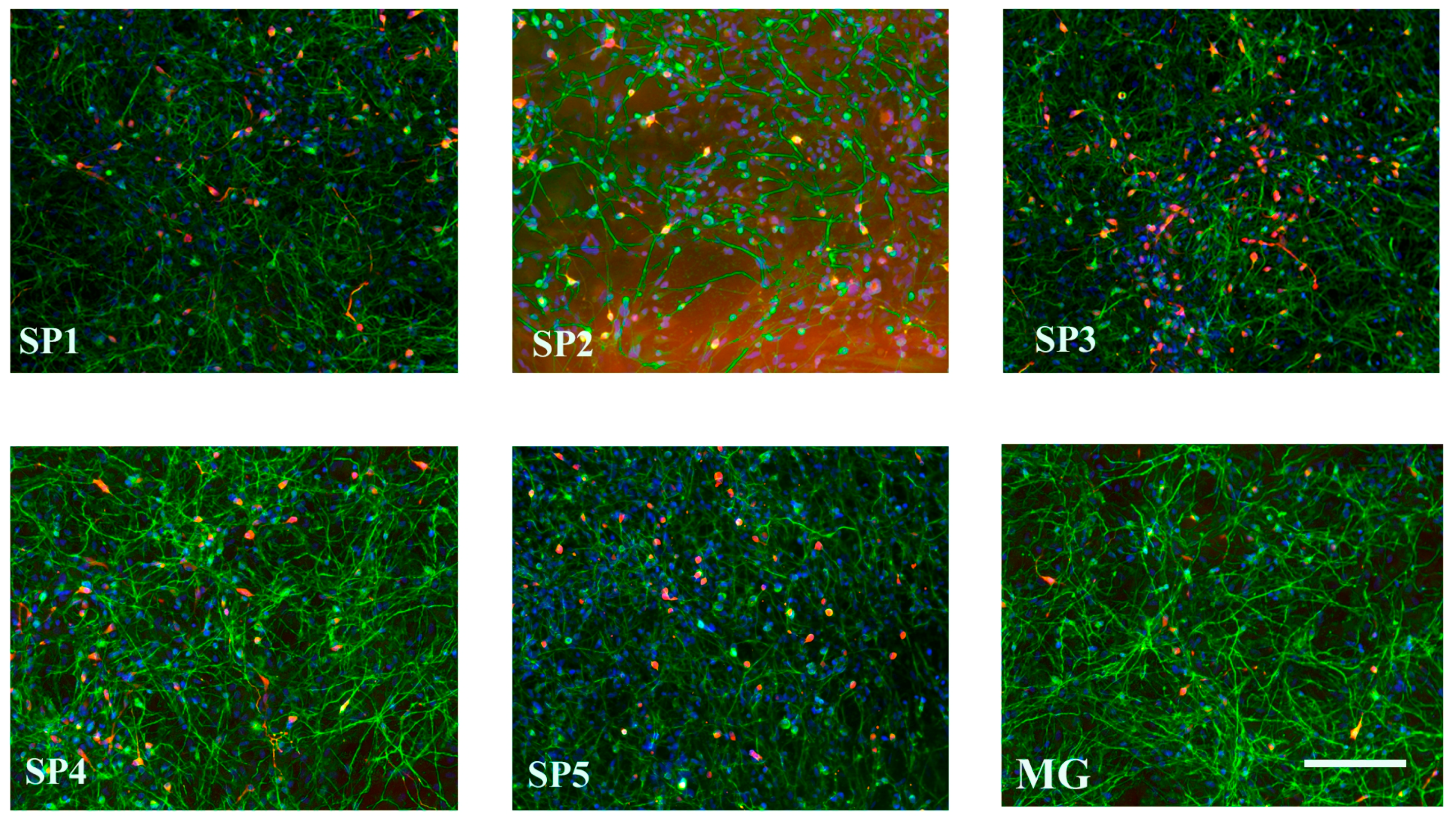
2.3. Influence of Cell Cultivation on CCs on Differentiation of NPCs into DA Neurons
2.4. Influence of Cell Cultivation on CCs on Synaptogenesis in Emerging NPCs and IDNs
2.5. Effect of FPs on the Efficiency of Synaptogenesis during Neuronal Differentiation on CCs
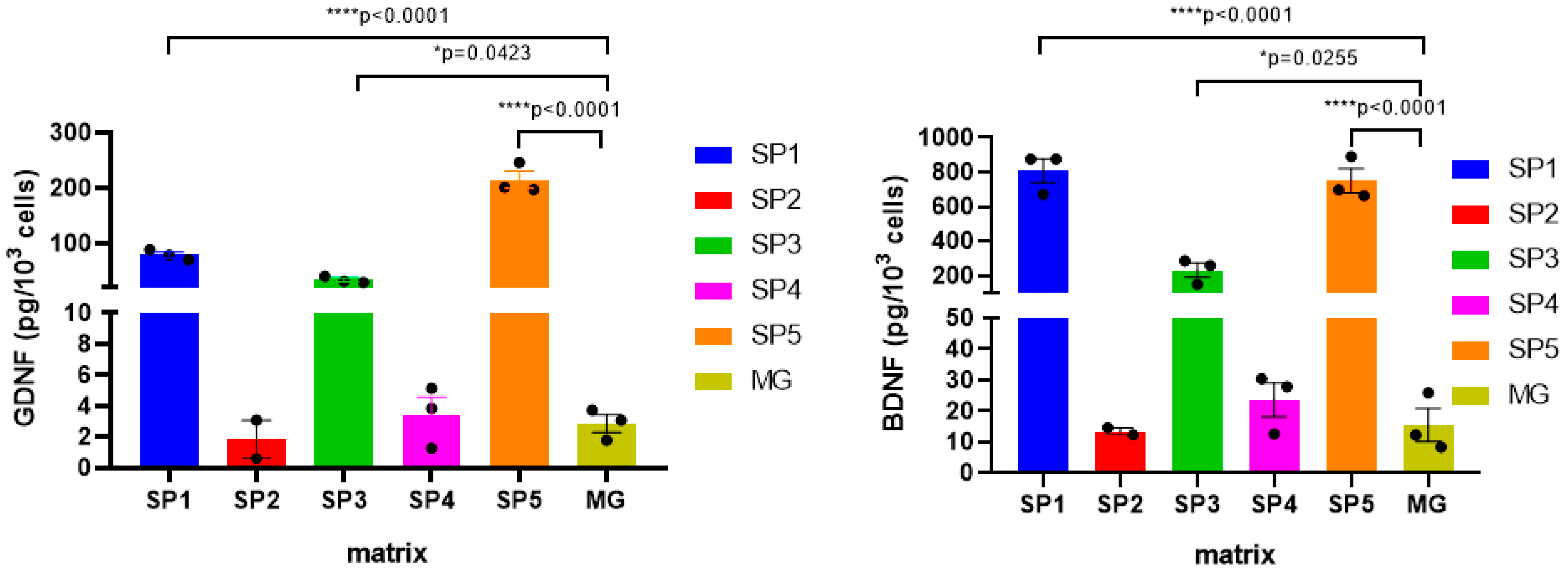
2.6. Effect of Cultivation on CCs on the Expression of Neurotrophic Factors (NTF) Genes in IDNs and DNs Formed on Them
3. Discussion
4. Methods and Materials
4.1. Isolation and Purification of Full Size RSs
4.2. Obtaining and Purification Fused Peptides (FPs)
4.3. Preparation of Mixed Protein Solutions for Dishes Coating
4.4. The Coating Preparation
4.5. Ethics Statement
4.6. Human Pluripotent Stem Cell Culture
4.7. Generation of Human iPSC-Derived Neural Stem Cells (NSCs)
4.8. Media Used for Cultivation and Differentiation of Neurons
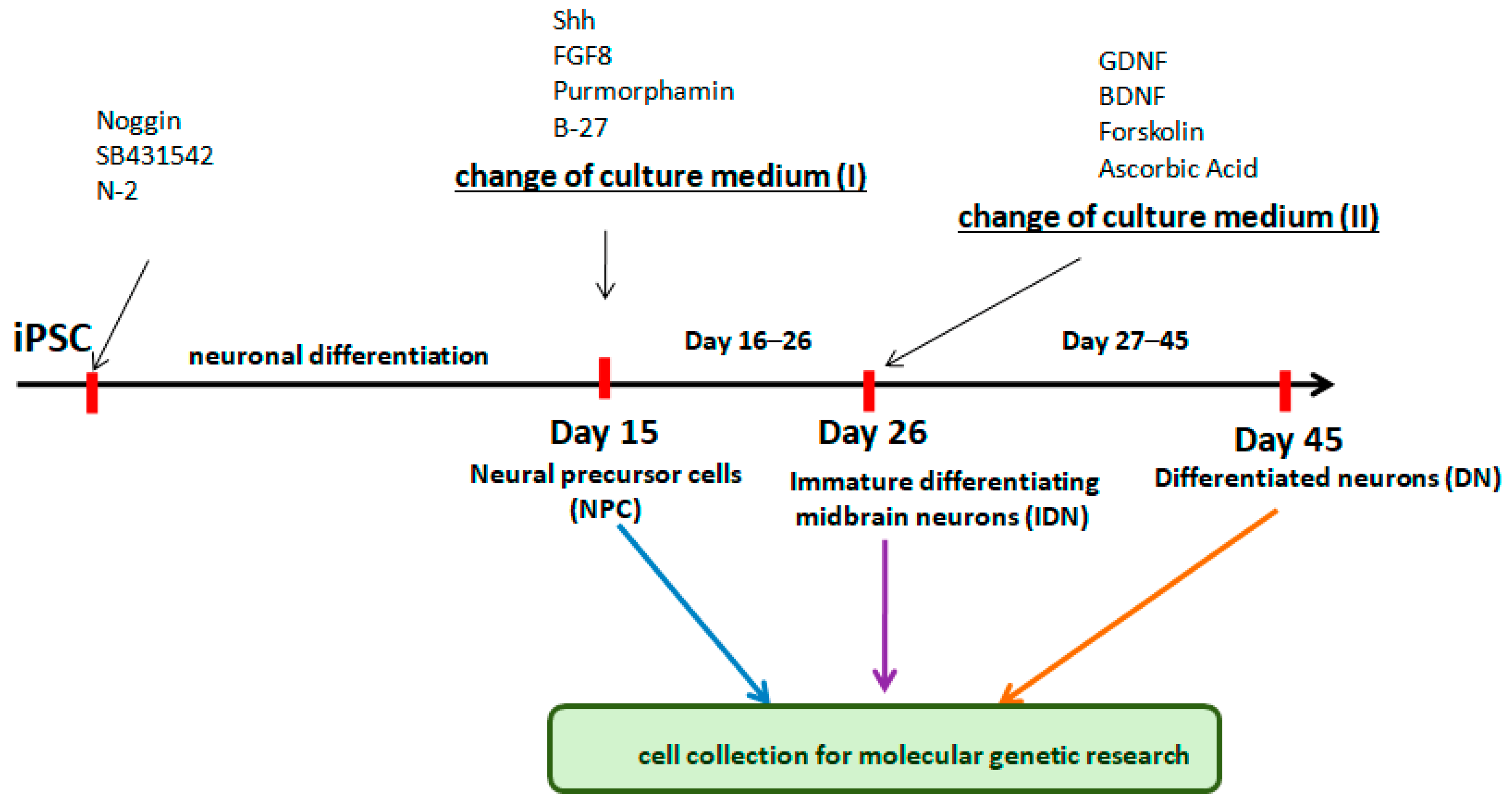
4.9. Targeted Differentiation of NPCs in IDN and DN
4.10. Immunocytochemistry
4.11. Quantification of BDNF and GDNF Protein Levels in Cultured Cells
4.12. Quantitative Real-Time PCR (qPCR)
4.13. MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) Assay
4.14. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gholipourmalekabadi, M.; Zhao, S.; Harrison, B.S.; Mozafari, M.; Seifalian, A.M. Oxygen-Generating Biomaterials: A New, Viable Paradigm for Tissue Engineering? Trends Biotechnol. 2016, 34, 1010–1021. [Google Scholar] [CrossRef]
- Blackledge, T.A. Spider silk: A brief review and prospectus on research linking biomechanics and ecology in draglines and orb webs. J. Arachnol. 2012, 40, 1–12. [Google Scholar] [CrossRef]
- Rising, A.; Widhe, M.; Johansson, J.; Hedhammar, M. Spider silk proteins: Recent advances in recombinant production, structure-function relationships and biomedical applications. Cell Mol. Life Sci. 2011, 68, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Debabov, V.G.; Bogush, V.G. Recombinant Spidroins as the Basis for New Materials. ACS Biomater. Sci. Eng. 2020, 6, 3745–3761. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Tang-Schomer, M.D.; Huang, W.; Xia, X.X.; Weiss, A.S.; Kaplan, D.L. Charge-Tunable Silk-Tropoelastin Protein Alloys That Control Neuron Cell Responses. Adv. Funct. Mater. 2013, 23, 3875–3884. [Google Scholar] [CrossRef] [PubMed]
- Jansson, R.; Thatikonda, N.; Lindberg, D.; Rising, A.; Johansson, J.; Nygren, P.; Hedhammar, M. Recombinant spider silk genetically functionalized with affinity domains. Biomacromolecules 2014, 15, 1696–1706. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Kleinman, H.K.; Martin, G.R. Matrigel: Basement membrane matrix with biological activity. Semin. Cancer Biol. 2005, 15, 378–386. [Google Scholar] [CrossRef]
- An, B.; Tang-Schomer, M.; Huang, W.; He, J.; Jones, J.; Lewis, R.V.; Kaplan, D.L. Physical and biological regulation of neuron regenerative growth and network formation on recombinant dragline silks. Biomaterials 2015, 48, 137–146. [Google Scholar] [CrossRef]
- Lewicka, M.; Hermanson, O.; Rising, A.U. Recombinant spider silk matrices for neural stem cell cultures. Biomaterials 2012, 33, 7712–7717. [Google Scholar] [CrossRef]
- Baklaushev, V.P.; Bogush, V.G.; Kalsin, V.A.; Sovetnikov, N.N.; Samoilova, E.M.; Revkova, V.A.; Sidoruk, K.V.; Konoplyannikov, M.A.; Timashev, P.S.; Kotova, S.L.; et al. Tissue Engineered Neural Constructs Composed of Neural Precursor Cells, Recombinant Spidroin and PRP for Neural Tissue Regeneration. Sci. Rep. 2019, 9, 3161. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, S.V.; Sullivan, A.M.; O’Keeffe, G.W. Midbrain dopaminergic neurons: A review of the molecular circuitry that regulates their development. Dev. Biol. 2013, 379, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Novosadova, E.; Antonov, S.; Arsenyeva, E.; Kobylanskiy, A.; Vanyushina, Y.; Malova, T.; Khaspekov, L.; Bobrov, M.; Bezuglov, V.; Tarantul, V.; et al. Neuroprotective and neurotoxic effects of endocannabinoid-like compounds, N-arachidonoyl dopamine and N-docosahexaenoyl dopamine in differentiated cultures of induced pluripotent stem cells derived from patients with Parkinson’s disease. Neurotoxicology 2021, 82, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Teplenin, A.; Krasheninnikova, A.; Agladze, N.; Sidoruk, K.; Agapova, O.; Agapov, I.; Bogush, V.; Agladze, K. Functional analysis of the engineered cardiac tissue grown on recombinant spidroin fiber meshes. PLoS ONE 2015, 10, e0121155. [Google Scholar] [CrossRef]
- Moisenovich, M.M.; Pustovalova, O.; Shackelford, J.; Vasiljeva, T.V.; Druzhinina, T.V.; Kamenchuk, Y.A.; Guzeev, V.V.; Sokolova, O.S.; Bogush, V.G.; Debabov, V.G.; et al. Tissue regeneration in vivo within recombinant spidroin 1 scaffolds. Biomaterials 2012, 33, 3887–3898. [Google Scholar] [CrossRef]
- Nosenko, M.A.; Moysenovich, A.M.; Zvartsev, R.V.; Arkhipova, A.Y.; Zhdanova, A.S.; Agapov, I., II.; Vasilieva, T.V.; Bogush, V.G.; Debabov, V.G.; Nedospasov, S.A.; et al. Novel Biodegradable Polymeric Microparticles Facilitate Scarless Wound Healing by Promoting Re-epithelialization and Inhibiting Fibrosis. Front. Immunol. 2018, 9, 2851. [Google Scholar] [CrossRef]
- Hosoyama, K.; Lazurko, C.; Muñoz, M.; McTiernan, C.D.; Alarcon, E.I. Peptide-Based Functional Biomaterials for Soft-Tissue Repair. Front. Bioeng. Biotechnol. 2019, 7, 205. [Google Scholar] [CrossRef]
- Revkova, V.A.; Sidoruk, K.V.; Kalsin, V.A.; Melnikov, P.A.; Konoplyannikov, M.A.; Kotova, S.; Frolova, A.A.; Rodionov, S.A.; Smorchkov, M.M.; Kovalev, A.V.; et al. Spidroin Silk Fibers with Bioactive Motifs of Extracellular Proteins for Neural Tissue Engineering. ACS Omega 2021, 6, 15264–15273. [Google Scholar] [CrossRef]
- Mikhailova, M.M.; Sydoruk, K.V.; Davydova, L.I.; Yastremsky, E.V.; Chvalun, S.N.; Debabov, V.G.; Bogush, V.G.; Panteleyev, A.A. Nonwoven spidroin materials as scaffolds for ex vivo cultivation of aortic fragments and dorsal root ganglia. J. Biomater. Sci. Polym. Ed. 2022, 33, 1685–1703. [Google Scholar] [CrossRef]
- Timpl, R.; Rohde, H.; Robey, P.G.; Rennard, S.I.; Foidart, J.M.; Martin, G.R. Laminin—A glycoprotein from basement membranes. J. Biol. Chem. 1979, 254, 9933–9937. [Google Scholar] [CrossRef]
- Silva, A.; Pereira, J.; Oliveira, C.R.; Relvas, J.B.; Rego, A.C. BDNF and extracellular matrix regulate differentiation of mice neurosphere-derived cells into a GABAergic neuronal phenotype. J. Neurosci. Res. 2009, 87, 1986–1996. [Google Scholar] [CrossRef]
- Pierret, C.; Morrison, J.A.; Rath, P.; Zigler, R.E.; Engel, L.A.; Fairchild, C.L.; Shi, H.; Maruniak, J.A.; Kirk, M.D. Developmental cues and persistent neurogenic potential within an in vitro neural niche. BMC Dev. Biol. 2010, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, J.; Ishihara, A.; Fukunaga, K.; Sasaki, K.; White, M.J.V.; Briquez, P.S.; Hubbell, J.A. Laminin heparin-binding peptides bind to several growth factors and enhance diabetic wound healing. Nat. Commun. 2018, 9, 2163. [Google Scholar] [CrossRef] [PubMed]
- Bothwell, M. Recent advances in understanding neurotrophin signaling. F1000Research 2016, 5, 1885. [Google Scholar] [CrossRef]
- Pramanik, S.; Sulistio, Y.A.; Heese, K. Neurotrophin Signaling and Stem Cells-Implications for Neurodegenerative Diseases and Stem Cell Therapy. Mol. Neurobiol. 2017, 54, 7401–7459. [Google Scholar] [CrossRef] [PubMed]
- Vilar, M.; Mira, H. Regulation of Neurogenesis by Neurotrophins during Adulthood: Expected and Unexpected Roles. Front. Neurosci. 2016, 10, 26. [Google Scholar] [CrossRef]
- Bogush, V.G.; Sokolova, O.S.; Davydova, L.I.; Klinov, D.V.; Sidoruk, K.V.; Esipova, N.G.; Neretina, T.V.; Orchanskyi, I.A.; Makeev, V.Y.; Tumanyan, V.G.; et al. A novel model system for design of biomaterials based on recombinant analogs of spider silk proteins. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2009, 4, 17–27. [Google Scholar] [CrossRef]
- Matsuda, A.; Kobayashi, H.; Itoh, S.; Kataoka, K.; Tanaka, J. Immobilization of laminin peptide in molecularly aligned chitosan by covalent bonding. Biomaterials 2005, 26, 2273–2279. [Google Scholar] [CrossRef]
- Mauri, E.; Sacchetti, A.; Vicario, N.; Peruzzotti-Jametti, L.; Rossi, F.; Pluchino, S. Evaluation of RGD functionalization in hybrid hydrogels as 3D neural stem cell culture systems. Biomater. Sci. 2018, 6, 501–510. [Google Scholar] [CrossRef]
- Moisenovich, M.M.; Silachev, D.N.; Moysenovich, A.M.; Arkhipova, A.Y.; Shaitan, K.V.; Bogush, V.G.; Debabov, V.G.; Latanov, A.V.; Pevzner, I.B.; Zorova, L.D.; et al. Effects of Recombinant Spidroin rS1/9 on Brain Neural Progenitors After Photothrombosis-Induced Ischemia. Front. Cell Dev. Biol. 2020, 8, 823. [Google Scholar] [CrossRef]
- Johnson, E.S.; Schwienhorst, I.; Dohmen, R.J.; Blobel, G. The ubiquitin-like protein Smt3p is activated for conjugation to other proteins by an Aos1p/Uba2p heterodimer. EMBO J. 1997, 16, 5509–5519. [Google Scholar] [CrossRef] [PubMed]
- Marblestone, J.G.; Edavettal, S.C.; Lim, Y.; Lim, P.; Zuo, X.; Butt, T.R. Comparison of SUMO fusion technology with traditional gene fusion systems: Enhanced expression and solubility with SUMO. Protein Sci. Publ. Protein Soc. 2006, 15, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Hirano, Y.; Okuno, M.; Hayashi, T.; Goto, K.; Nakajima, A. Cell-attachment activities of surface immobilized oligopeptides RGD, RGDS, RGDV, RGDT, and YIGSR toward five cell lines. J. Biomater. Sci. Polym. Ed. 1993, 4, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Wohlrab, S.; Müller, S.; Schmidt, A.; Neubauer, S.; Kessler, H.; Leal-Egaña, A.; Scheibel, T. Cell adhesion and proliferation on RGD-modified recombinant spider silk proteins. Biomaterials 2012, 33, 6650–6659. [Google Scholar] [CrossRef] [PubMed]
- Sapir, Y.; Kryukov, O.; Cohen, S. Integration of multiple cell-matrix interactions into alginate scaffolds for promoting cardiac tissue regeneration. Biomaterials 2011, 32, 1838–1847. [Google Scholar] [CrossRef] [PubMed]
- Huemmerich, D.; Slotta, U.; Scheibel, T. Processing and modification of films made from recombinant spider silk proteins. Appl. Phys. A 2006, 82, 219–222. [Google Scholar] [CrossRef]
- Novosadova, E.V.; Nekrasov, E.D.; Chestkov, I.; Surdina, A.V.; Vasina, E.M.; Bogomazova, A.; Manuilova, E.S.; Arsenyeva, E.L.; Simonova, V.V.; Konovalova, E.V.; et al. A Platform for Studying Molecular and Cellular Mechanisms of Parkinson’s Disease Based on Human Induced Pluripotent Stem Cells. Sovrem. Tehnol. V Med. 2016, 8, 157–166. [Google Scholar] [CrossRef]
- Pollock, G.S.; Vernon, E.; Forbes, M.E.; Yan, Q.; Ma, Y.T.; Hsieh, T.; Robichon, R.; Frost, D.O.; Johnson, J.E. Effects of early visual experience and diurnal rhythms on BDNF mRNA and protein levels in the visual system, hippocampus, and cerebellum. J. Neurosci. Off. J. Soc. Neurosci. 2001, 21, 3923–3931. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coating | PAX6 | SOX2 | NESTIN | TUBB3 |
|---|---|---|---|---|
| SP1/MG | 0.83 ± 0.22 | 0.83 ± 0.69 | 1.06 ± 0.38 | 0.71 ± 0.4 |
| SP2/MG | 0.96 ± 0.18 | 0.96 ± 0.32 | 1.26 ± 0.18 | 0.89 ± 0.16 |
| SP3/MG | 1.25 ± 0.14 | 1.25 ± 0.19 | 1.22 ± 0.14 | 0.78 ± 0.16 |
| SP4/MG | 1.06 ± 0.30 | 1.06 ± 0.42 | 1.27 ± 0.21 | 0.90 ± 0.02 |
| SP5/MG | 1.59 ± 0.25 | 1.59 ± 0.21 | 1.06 ± 0.16 | 0.79 ± 0.17 |
| Coating | SNAP 25 | STX1A | SNPT | SYN2 | SYN3 |
|---|---|---|---|---|---|
| Docking of Synaptic Vesicles with the Membrane | Synaptogenesis | ||||
| SP1/MG | 0.88 ± 0.14 | 0.62 ± 0.18 | 1.12 ± 0.34 | 0.97 ± 0.26 | 0.85 ± 0.34 |
| SP2/MG | 1.03 ± 0.19 | 0.82 ± 0.05 | 1.36 ± 0.34 | 1.30 ± 0.19 | 1.57 ± 0.39 |
| SP3/MG | 1.07 ± 0.18 | 0.94 ± 0.06 | 1.18 ± 0.30 | 1.18 ± 0.13 | 1.67 ± 0.35 |
| SP4/MG | 0.97 ± 0.17 | 0.88 ± 0.06 | 0.96 ± 0.04 | 1.08 ± 0.05 | 1.63 ± 0.88 |
| SP5/MG | 1.07 ± 0.17 | 0.81 ± 0.14 | 1.17 ± 0.59 | 1.25 ± 0.19 | 1.79 ± 0.69 |
| Coating | SNAP25 | STX1A | SNPT | PSD95 | GSG1L | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Docking of Synaptic Vesicles with the Membrane | Post Synaptic Sites | Post Synaptic Density | ||||||||
| IDN | DN | IDN | DN | IDN | DN | IDN | DN | IDN | DN | |
| SP1/MG | 2.07 *** | 0.87 | 1.67 ** | 1.18 | 1.39 | 2.49 ** | 1.04 | 0.91 | 3.91 *** | 1.23 |
| SP2/MG | 1.86 *** | 0.93 | 1.40 | 1.12 | 1.26 | 1.71 | 1.05 | 1.07 | 2.48 ** | 1.40 |
| SP3/MG | 1.69 ** | 0.96 | 1.67 ** | 1.16 | 1.49 | 1.74 | 1.06 | 0.95 | 2.52 ** | 1.18 |
| SP4/MG | 0.75 | 1.05 | 1.23 | 1.31 | 1.56 | 0.93 | 0.66 | 0.82 | 1.10 | 1.23 |
| SP5/MG | 2.21 *** | 0.78 | 2.63 *** | 1.22 | 1.34 | 2.14 * | 1.13 | 0.81 | 4.65 *** | 1.38 |
| Coating | SYN | SYN2 | SYN3 | |||
|---|---|---|---|---|---|---|
| IDN | DN | IDN | DN | IDN | DN | |
| SP1/MG | 1.14 | 1.10 | 2.58 *** | 1.03 | 1.31 | 0.75 |
| SP2/MG | 1.21 | 1.37 | 2.00 ** | 1.00 | 1.88 | 0.96 |
| SP3/MG | 1.14 | 1.21 | 2.08 ** | 1.18 | 2.25 * | 0.83 |
| SP4/MG | 0.90 | 1.14 | 1.33 | 1.15 | 1.00 | 0.93 |
| SP5/MG | 1.38 | 1.07 | 3.22 *** | 1.13 | 1.75 | 0.69 * |
| Coating | SNAP 25 | STX1A | SNPT | SYP | PSD95 | GSG1L | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Docking of Synaptic Vesicles with the Membrane | Synaptic Protein Vesicle | Post Synaptic Sites | Post Synaptic Density | |||||||||
| IDN | DN | IDN | DN | IDN | DN | IDN | DN | IDN | DN | IDN | DN | |
| SP3/SP4 | 2.24 ** | 0.91 | 1.36 | 0.89 | 0.95 | 1.87 * | 1.22 | 1.05 | 1.59 | 1.15 * | 2.29 ** | 0.96 |
| SP1/SP2 | 1.11 | 0.93 | 1.19 | 1.05 | 1.10 | 1.46 | 0.79 | 0.94 | 0.99 | 0.85 | 1.57 * | 0.88 |
| SP5/SP2 | 1.19 | 0.84 | 1.87 *** | 1.09 | 1.06 | 1.25 | 0.80 | 0.83 | 1.08 | 0.75 | 1.87 ** | 0.98 |
| Coating | SYN | SYN2 | SYN3 | |||
|---|---|---|---|---|---|---|
| IDN | DN | IDN | DN | IDN | DN | |
| SP3/SP4 | 1.27 | 1.06 | 1.50 * | 1.03 | 2.22 | 0.89 |
| SP1/SP2 | 0.87 | 0.80 | 1.33 * | 1.02 | 0.68 | 0.78 |
| SP5/SP2 | 1.11 | 0.78 | 1.78 ** | 1.12 | 0.88 | 0.72 |
| Coating | BDNF | NT3 | GDNF | NGF | ||||
|---|---|---|---|---|---|---|---|---|
| IDN | DN | IDN | DN | IDN | DN | IDN | DN | |
| SP1/MG | 3.85 *** | 0.87 | 0.54 *** | 0.77 | 11.92 *** | 1.61 | 3.38 *** | 1.60 |
| SP2/MG | 2.34 ** | 0.98 | 0.64 ** | 0.93 | 4.86 | 1.70 | 1.95 * | 1.36 |
| SP3/MG | 3.04 ** | 0.83 | 0.61 ** | 0.63 | 5.05 * | 2.06 * | 2.83 *** | 1.46 |
| SP4/MG | 1.33 | 0.85 | 0.50 ** | 0.69 | 3.20 | 1.34 | 1.89 * | 1.12 |
| SP5/MG | 4.34 *** | 0.63 | 0.26 *** | 0.81 | 5.98 * | 2.16 ** | 2.96 *** | 0.98 |
| Marker | Antibodies | Dilution | Company, Cat # |
|---|---|---|---|
| Primary antibodies | Mouse anti—NeuN | 1:500 | Abcam, #ab 104224 |
| Rabbit anti—TH Mouse anti—βIII Tubulin | 1:1000 1:2000 | Abcam, #ab 112 Abcam, #ab 7751 | |
| Rabbit anti—Sox2 | 1:1000 | Abcam, # 97959 | |
| Rabbit anti—S100 | 1:2 | Agilent Dako, #GA50461-2 | |
| Secondary antibodies | Goat anti-Rabbit IgG (H + L), AF546 | 1:1000 | ThermoFisher, #A11010 |
| Goat anti- Mouse IgG (H + L), AF488 | 1:1000 | ThermoFisher, #A11008 |
| Gene | Forward | Reverse |
|---|---|---|
| PAX6 | CCGAGAGTAGCGACTCCAG | CTTCCGGTCTGCCCGTTC |
| SOX2 | TCCTGATTCCAGTTTGCCTC | GCTTAGCCTCGTCGATGAAC |
| NESTIN | CAGCTGGCGCACCTCAAGATG | AGGGAAGTTGGGCTCAGGACTGG |
| TUBB3 | CTCAGGGGCCTTTGGACATC | CAGGCAGTCGCAGTTTTCAC |
| TH | GTCCCCTGGTTCCCAAGAAAAGT | TCCAGCTGGGGGATATTGTCTTC |
| AADC | CTCGGACCAAAGTGATCCAT | GGGTGGCAACCATAAAGAAA |
| BDNF | ATTGGCTGGCGATTCATAAG | GTTTCCCTTCTGGTCATGGA |
| NT3 | AACTGCTGCGACAACAGAGA | CCAGCCCACGAGTTTATTGT |
| TRKC | CCGACACTGTGGTCATTGGCAT | CAGTTCTCGCTTCAGCACGATG |
| GDNF | ACCTGGAGTTAATGTCCAACC | GGCATATTTGAGTCACTGCT |
| GFRα | GCCTGTGTGCTCCTATGAAG | CTGGCTGGCAGTTGGTAAA |
| NGF | TAAAAAGCGGCGACTCCGTT | ATTCGCCCCTGTGGAAGATG |
| P75NTR | AAGGAGGGGAGGTGATAGGA | GTGGGACAGAGTCTGGGTGT |
| SNAP25 | CGTCGTATGCTGCAACTGGTTG | GGTTCATGCCTTCTTCGACACG |
| STX1A | TGGAGAACAGCATCCGTGAGCT | CCTCTCCACATAGTCTACCGCG |
| SNPT1 | GCTGACTGTTGTCATTCTGGAGG | CTTCAGCCTCTTACCATTCTGCA |
| SYP | TCGGCTTTGTGAAGGTGCTGCA | TCACTCTCGGTCTTGTTGGCAC |
| PSD95 | TCCACTCTGACAGTGAGACCGA | CGTCACTGTCTCGTAGCTCAGA |
| SYN3 | GTGGACATGCAGGTCGTGAGAA | ATGACCAGGCTGCGGTAGTCTT |
| 18S | CGGCTACCACATCCAAGGAA | GCTGGAATTACCGCGGCT |
| GSG1L | CTCCTACACCAAGACGGTCATTG | GGCAGTCTAAGTGAAAGTCCTCC |
| SYN | CGATGCCAAATATGACGTGCGTG | AGCATCGCAGAGCCAGTATTGG |
| SYN2 | ACCTGCTCTGAGATGTTTGGCG | GTTCGGTGATGAGTTGCCTGTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novosadova, E.V.; Dolotov, O.V.; Novosadova, L.V.; Davydova, L.I.; Sidoruk, K.V.; Arsenyeva, E.L.; Shimchenko, D.M.; Debabov, V.G.; Bogush, V.G.; Tarantul, V.Z. Composite Coatings Based on Recombinant Spidroins and Peptides with Motifs of the Extracellular Matrix Proteins Enhance Neuronal Differentiation of Neural Precursor Cells Derived from Human Induced Pluripotent Stem Cells. Int. J. Mol. Sci. 2023, 24, 4871. https://doi.org/10.3390/ijms24054871
Novosadova EV, Dolotov OV, Novosadova LV, Davydova LI, Sidoruk KV, Arsenyeva EL, Shimchenko DM, Debabov VG, Bogush VG, Tarantul VZ. Composite Coatings Based on Recombinant Spidroins and Peptides with Motifs of the Extracellular Matrix Proteins Enhance Neuronal Differentiation of Neural Precursor Cells Derived from Human Induced Pluripotent Stem Cells. International Journal of Molecular Sciences. 2023; 24(5):4871. https://doi.org/10.3390/ijms24054871
Chicago/Turabian StyleNovosadova, Ekaterina V., Oleg V. Dolotov, Lyudmila V. Novosadova, Lubov I. Davydova, Konstantin V. Sidoruk, Elena L. Arsenyeva, Darya M. Shimchenko, Vladimir G. Debabov, Vladimir G. Bogush, and Vyacheslav Z. Tarantul. 2023. "Composite Coatings Based on Recombinant Spidroins and Peptides with Motifs of the Extracellular Matrix Proteins Enhance Neuronal Differentiation of Neural Precursor Cells Derived from Human Induced Pluripotent Stem Cells" International Journal of Molecular Sciences 24, no. 5: 4871. https://doi.org/10.3390/ijms24054871
APA StyleNovosadova, E. V., Dolotov, O. V., Novosadova, L. V., Davydova, L. I., Sidoruk, K. V., Arsenyeva, E. L., Shimchenko, D. M., Debabov, V. G., Bogush, V. G., & Tarantul, V. Z. (2023). Composite Coatings Based on Recombinant Spidroins and Peptides with Motifs of the Extracellular Matrix Proteins Enhance Neuronal Differentiation of Neural Precursor Cells Derived from Human Induced Pluripotent Stem Cells. International Journal of Molecular Sciences, 24(5), 4871. https://doi.org/10.3390/ijms24054871