
Characterization of Different Types of Epiretinal Proliferations by Synchrotron Radiation-Based Fourier Transform Infrared Micro-Spectroscopy




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Lipid Region
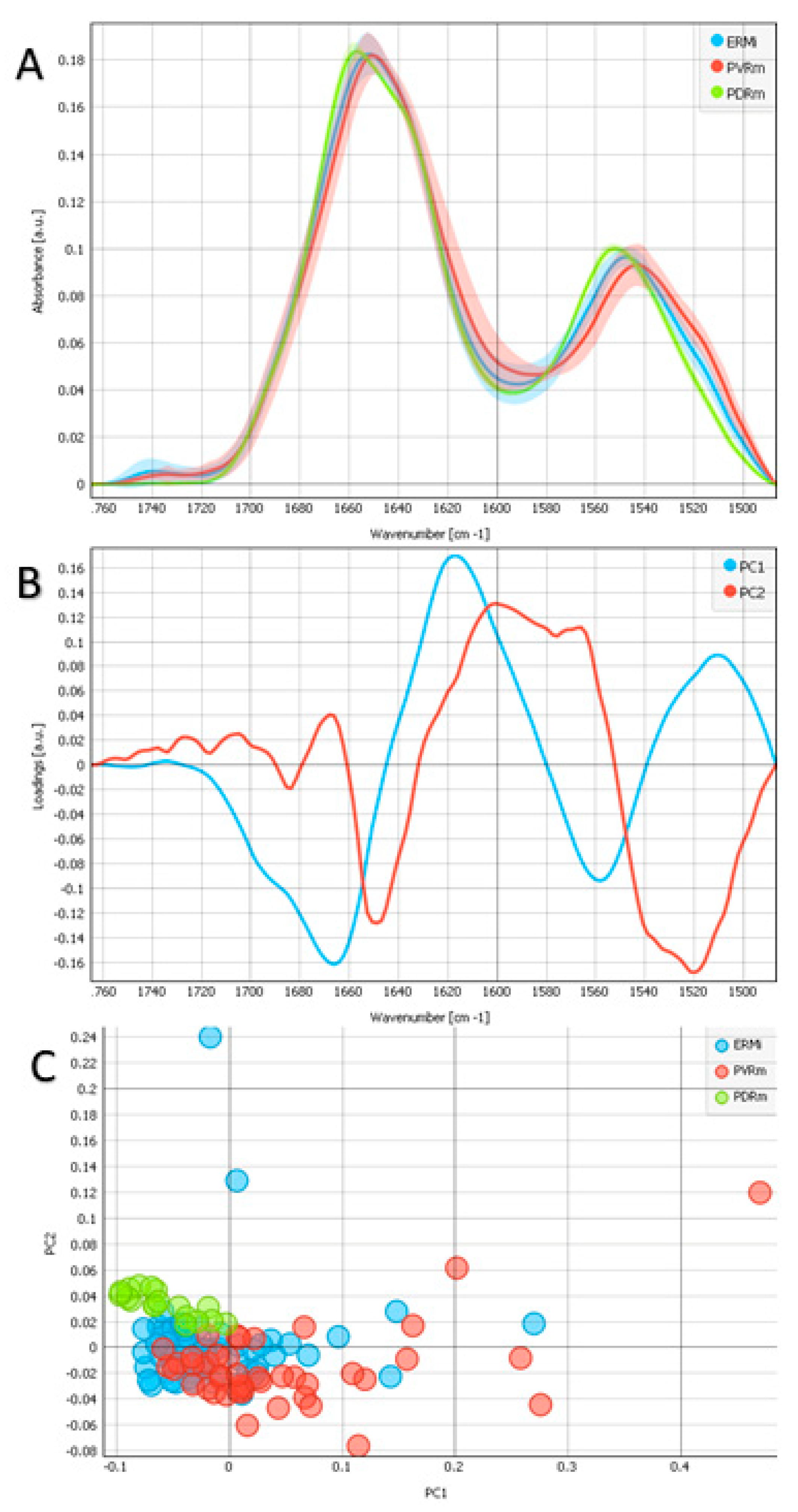
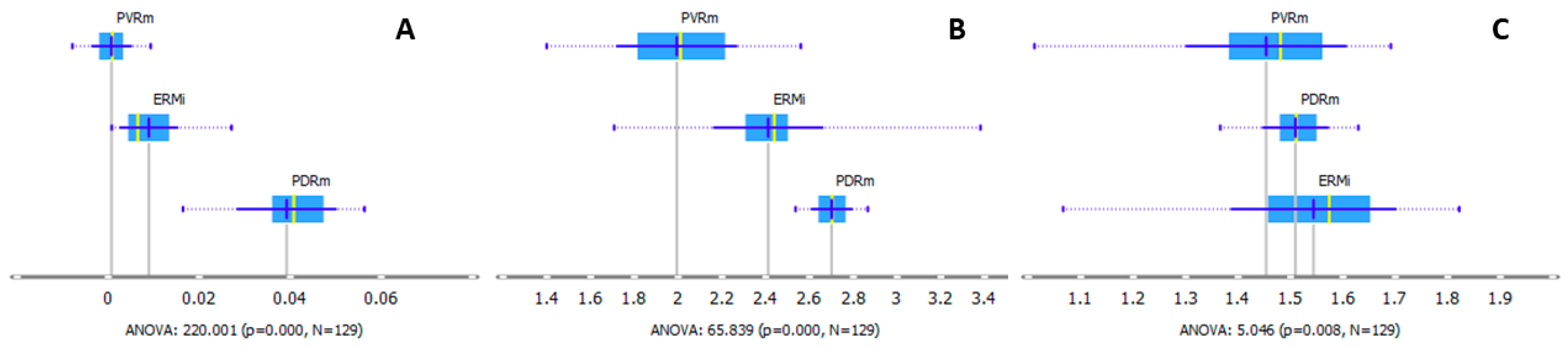
2.2. Protein Region
2.3. Protein Region Band Deconvolution
2.4. Nucleic Acids and Carbohydrates Regions
2.5. Additional Protein Analysis
3. Discussion
4. Materials & Methods
4.1. Sample Preparation
4.2. Synchrotron Radiation-Based FTIR Micro-Spectroscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- George, B.; Chen, S.; Chaudhary, V.; Gonder, J.; Chakrabarti, S. Extracellular matrix proteins in epiretinal membranes and in diabetic retinopathy. Curr. Eye Res. 2009, 34, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Tsotridou, E.; Loukovitis, E.; Zapsalis, K.; Pentara, I.; Asteriadis, S.; Tranos, P.; Zachariadis, Z.; Anogeianakis, G. A Review of Last Decade Developments on Epiretinal Membrane Pathogenesis. Med. Hypothesis Discov. Innov. Ophthalmol. 2020, 9, 91–110. [Google Scholar] [PubMed]
- Fung, A.; Galvin, J.; Tran, T. Epiretinal membrane: A review. Clin. Exp. Ophthalmol. 2021, 49, 289–308. [Google Scholar] [CrossRef] [PubMed]
- Smiddy, W.E.; Maguire, A.M.; Green, W.R.; Michels, R.G.; de la Cruz, Z.; Enger, C.; Jaeger, M.; Rice, T.A. Idiopathic epiretinal membranes: Ultrastructural characteristics and clinicopathologic correlation. Retina 1989, 25 (Suppl. 5), 811–820. [Google Scholar] [CrossRef] [PubMed]
- Andjelić, S.; Lumi, X.; Yan, X.; Graw, J.; Moe, M.C.; Facskó, A.; Hawlina, M.; Petrovski, G. Characterization of ex vivo cultured neuronal- and glial- like cells from human idiopathic epiretinal membranes. BMC Ophthalmol. 2014, 23, 165. [Google Scholar] [CrossRef]
- Bu, S.C.; Kuijer, R.; van der Worp, R.J.; Huiskamp, E.A.; Renardel de Lavalette, V.W.; Li, X.R.; Hooymans, J.M.; Los, L.I. Glial cells and collagens in epiretinal membranes associated with idiopathic macularholes. Retina 2014, 34, 897–906. [Google Scholar] [CrossRef]
- Schumann, R.G.; Eibl, K.H.; Zhao, F.; Scheerbaum, M.; Scheler, R.; Schaumberger, M.M.; Wehnes, H.; Walch, A.K.; Haritoglou, C.; Kampik, A.; et al. Immunocytochemical and ultrastructural evidence of glial cells and hyalocytes in internal limiting membrane specimens of idiopathic macular holes. Investig. Ophthalmol. Vis. Sci. 2011, 52, 7822–7834. [Google Scholar] [CrossRef]
- Bringmann, A.; Pannicke, T.; Grosche, J.; Francke, M.; Wiedemann, P.; Skatchkov, S.N.; Osborne, N.N.; Reichenbach, A. Muller cells in the healthy and diseased retina. Prog. Retin. Eye Res. 2006, 25, 397–424. [Google Scholar] [CrossRef]
- Ioachim, E.; Stefaniotou, M.; Gorezis, S.; Tsanou, E.; Psilas, K.; Agnantis, N.J. Immunohistochemical study of extracellular matrix components in epiretinal membranes of vitreoproliferative retinopathy and proliferative diabetic retinopathy. Eur. J. Ophthalmol. 2005, 15, 384–391. [Google Scholar] [CrossRef]
- Campochiaro, P. The Pathogenesis of Proliferative Vitreoretinopathy; Elsevier Mosby: Philadelphia, PA, USA, 2006. [Google Scholar]
- Veréb, Z.; Lumi, X.; Andjelic, S.; Globocnik-Petrovic, M.; Urbancic, M.; Hawlina, M.; Facskó, A.; Petrovski, G. Functional and molecular characterization of ex vivo cultured epiretinal membrane cells from human proliferative diabetic retinopathy. Biomed. Res. Int. 2013, 2013, 492376. [Google Scholar] [CrossRef]
- Tamaki, K.; Usui-Ouchi, A.; Murakami, A.; Ebihara, N. Fibrocytes and Fibrovascular Membrane Formation in Proliferative Diabetic Retinopathy. Investig. Ophthalmol. Vis. Sci. 2016, 57, 4999–5005. [Google Scholar] [CrossRef]
- Leisser, C.; Paschalis, E.; Rokidi, S.; Behanova, M.; Ruiss, M.; Burgmüller, W.; Findl, O. Fourier-Transform Infrared Spectroscopy of Epiretinal Membranes and Internal Limiting Membranes after Pars Plana Vitrectomy with Membrane Peeling. Ophthalmic Res. 2021, 64, 793–797. [Google Scholar] [CrossRef]
- Surewicz, W.K.; Mantsch, H.H. New insight into protein secondary structure from resolution-enhanced infrared spectra. Biochim. Biophys. Acta Protein Struct. Mol. 1988, 952, 115–130. [Google Scholar] [CrossRef]
- Mantsch, H.H.; McElhaney, R.N. Phospholipid phase transitions in model and biological membranes as studied by infrared spectroscopy. Chem. Phys. Lipids 1991, 57, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Taillandier, E.; Liquier, J. Infrared spectroscopy of DNA. Methods Enzymol. 1992, 211, 307–335. [Google Scholar] [CrossRef] [PubMed]
- Baranska, M. (Ed.) Optical Spectroscopy and Computational Methods in Biology and Medicine; Springer Science & Business Media: Dordrecht, The Netherlands, 2013; Volume 14. [Google Scholar]
- Miller, L.M.; Dumas, P. Chemical imaging of biological tissue with synchrotron infrared light. Biochim. Biophys. Acta Biomembr. 2006, 1758, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer, M.; Dučić, T.; Hawlina, M.; Andjelic, S. Synchrotron based FTIR microspectroscopy of protein aggregation and lipids peroxidation changes in human cataractous lens epithelial cells. Sci. Rep. 2020, 10, 15489. [Google Scholar] [CrossRef]
- Bhatia, N.K.; Srivastava, A.; Katyal, N.; Jain, N.; Khan, M.A.; Kundu, B.; Deep, S. Curcumin binds to the pre-fibrillar aggregates of Cu/Zn superoxide dismutase (SOD1) and alters its amyloidogenic pathway resulting in reduced cytotoxicity. Biochim. Biophys. Acta Proteins Proteom. 2015, 1854, 426–436. [Google Scholar] [CrossRef]
- Sarroukh, R.; Goormaghtigh, E.; Ruysschaert, J.M.; Raussens, V. ATR-FTIR: A ‘rejuvenated’ tool to investigate amyloid proteins. Biochim. Biophys. Acta Biomembr. 2013, 1828, 2328–2338. [Google Scholar] [CrossRef]
- Olsztyńska-Janus, S.; Pietruszka, A.; Kiełbowicz, Z.; Czarnecki, M.A. ATR-IR study of skin components: Lipids, proteins and water. Part I: Temperature effect. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2018, 188, 37–49. [Google Scholar] [CrossRef]
- Shahzad, M.I.; Giorcelli, M.; Shahzad, N.; Guastella, S.; Castellino, M.; Jagdale, P.; Tagliaferro, A. Study of carbon nanotubes based Polydimethylsiloxane composite films. J. Phys. Conf. Ser. 2013, 439, 012010. [Google Scholar] [CrossRef]
- Sanden, K.W.; Böcker, U.; Ofstad, R.; Pedersen, M.E.; Høst, V.; Afseth, N.K.; Rønning, S.B.; Pleshko, N. Characterization of Collagen Structure in Normal, Wooden Breast and Spaghetti Meat Chicken Fillets by FTIR Microspectroscopy and Histology. Foods 2021, 10, 548. [Google Scholar] [CrossRef]
- Cheheltani, R.; McGoverin, C.M.; Rao, J.; Vorp, D.A.; Kiani, M.F.; Pleshko, N. Fourier transform infrared spectroscopy to quantify collagen and elastin in an in vitro model of extracellular matrix degradation in aorta. Analyst 2014, 139, 3039–3047. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.Z.; Man, A.; Shaw, R.A.; Liang, B.; Xu, Z.; Gong, Y. Molecular determination of liver fibrosis by synchrotron infrared microspectroscopy. Biochim. Biophys. Acta 2006, 1758, 960–967. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Querido, W.; Kandel, S.; Pleshko, N. Applications of vibrational spectroscopy for analysis of connective tissues. Molecules 2021, 26, 922. [Google Scholar] [CrossRef] [PubMed]
- Suryadevara, V.; Nazeer, S.S.; Sreedhar, H.; Adelaja, O.; Kajdacsy-Balla, A.; Natarajan, V.; Walsh, M.J. Infrared spectral microscopy as a tool to monitor lung fibrosis development in a model system. Biomed. Opt. Express. 2020, 11, 3996–4007. [Google Scholar] [CrossRef] [PubMed]
- Mantsch, H.H.; Jackson, M. Molecular spectroscopy in biodiagnostics (from hippocrates to herschel and beyond). J. Mol. Struct. 1995, 347, 187–206. [Google Scholar] [CrossRef]
- Riaza, T.; Zeeshana, R.; Zarifa, F.; Ilyasa, K.; Muhammada, N.; Safia, S.Z.; Rahima, A.; Rizvib, S.A.A.; Rehman, I.U. FTIR analysis of natural and synthetic collagen. Appl. Spectrosc. Rev. 2018, 53, 703–746. [Google Scholar] [CrossRef]
- Lasch, P.; Boese, M.; Pacifico, A.; Diem, M. Ft–IR spectroscopic investigations of single cells on the subcellular level. Vib. Spectrosc. 2002, 28, 147–157. [Google Scholar] [CrossRef]
- Wood, B.R.; Chernenko, T.; Matthäus, C.; Diem, M.; Chong, C.; Bernhard, U.; Jene, C.; Brandli, A.A.; McNaughton, D.; Tobin, M.J.; et al. Shedding new light on the molecular architecture of oocytes using a combination of synchrotron Fourier transform-infrared and Raman spectroscopic imaging. Anal. Chem. 2008, 80, 9065–9072. [Google Scholar] [CrossRef]
- Ozek, N.S.; Tuna, S.; Erson-Bensan, A.E.; Severcan, F. Characterization of microRNA-125b expression in mCF7 breast cancer cells by AtR-FtIR spectroscopy. Analyst 2010, 135, 3094–3102. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kearns, V.R.; Zhou, L.; Sandinha, T.; Lam, W.C.; Steel, D.H.; Chan, Y.K. Silicone oil in vitreoretinal surgery: Indications, complications, new developments and alternative long-term tamponade agents. Acta Ophthalmol. 2021, 99, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Barca, F.; Caporossi, T.; Rizzo, S. Silicone oil: Different physical proprieties and clinical applications. Biomed. Res. Int. 2014, 2014, 502143. [Google Scholar] [CrossRef] [PubMed]
- Faulborn, J.; Bowald, S. Silikonöl in epiretinalen Membranen 3-4 Monate nach Implantation: Mikroskopische Befunde in 2 Fällen [Silicone oil in epiretinal membranes 3-4 months following implantation: Microscopic findings in 2 cases]. Klin Monbl. Augenheilkd. 1986, 188, 133–134. [Google Scholar] [CrossRef]
- Bornfeld, N.; E1-Hifnawi, E.; Laqua, H. Ultrastructural characteristics of preretinal membranes from human eyes filled with silicone oil. Am. J. Ophthalmol. 1987, 103, 770–775. [Google Scholar] [CrossRef]
- Kirchhof, B.; Tavakolian, U.; Paulmann, H.; Heimann, K. Histopathological findings in eyes after silicone oil injection. Graefes Arch. Clin. Exp. Ophthalmol. 1986, 224, 34–37. [Google Scholar] [CrossRef]
- Ni, C.; Wang, W.J.; Albert, D.M.; Schepens, C.L. Intravitreous silicone injection. Histopathologic findings in a human eye after 12 years. Arch. Ophthalmol. 1983, 101, 1399–1401. [Google Scholar] [CrossRef]
- Leaver, P.K.; Grey, R.H.; Garner, A. Silicone oil injection in the treatment of massive preretinal retraction: II. Late com plications in 93 eyes. Br. J. Ophthalmol. 1979, 63, 361–367. [Google Scholar] [CrossRef]
- Altera, A.; Tosi, G.M.; Regoli, M.; De Benedetto, E.; Bertelli, E. The extracellular matrix complexity of idiopathic epiretinal membranes and the bilaminar arrangement of the associated internal limiting membrane in the posterior retina. Graefes Arch. Clin. Exp. Ophthalmol. 2021, 259, 2559–2571. [Google Scholar] [CrossRef]
- Yang, Y.; Sulé-Suso, J.; Sockalingum, G.D.; Kegelaer, G.; Manfait, M.; El Haj, A.J. Study of tumor cell invasion by Fourier transform infrared microspectroscopy. Biopolymers 2005, 78, 311–317. [Google Scholar] [CrossRef]
- Wood, B.R.; Quinn, M.A.; Tait, B.; Ashdown, M.; Hislop, T.; Romeo, M.; McNaughton, D. FTIR microspectroscopic study of cell types and potential confounding variables in screening for cervical malignancies. Biospectroscopy 1998, 4, 75–91. [Google Scholar] [CrossRef]
- Dovbeshko, G.; Gridina, N.Y.; Kruglova, E.B.; Pashchuk, O.P. FTIR spectroscopy studies of nucleic acid damage. Talanta 1997, 53, 233–246. [Google Scholar] [CrossRef]
- Zohdi, V.; Whelan, D.R.; Wood, B.R.; Pearson, J.T.; Bambery, K.R.; Black, M.J. Importance of Tissue Preparation Methods in FTIR Micro-Spectroscopical Analysis of Biological Tissues: ‘Traps for New Users’. PLoS ONE 2015, 10, e0116491. [Google Scholar] [CrossRef] [PubMed]
- Yousef, I.; Ribó, L.; Crisol, A.; Šics, I.; Ellis, G.; Ducic, T.; Kreuzer, M.; Benseny-Cases, N.; Quispe, M.; Dumas, P.; et al. MIRAS: The Infrared Synchrotron Radiation Beamline at ALBA. Synchrotron Radiat News 2017, 30, 4–6. [Google Scholar] [CrossRef]
- Toplak, M.; Birarda, G.; Read, S.; Sandt, C.; Rosendahl, S.M.; Vaccari, L.; Demšar, J.; Borondics, F. Infrared Orange: Connecting Hyperspectral Data with Machine Learning. Synchrotron Radiat News 2017, 30, 40–45. [Google Scholar] [CrossRef]
- Lumi, X.; Dučić, T.; Kreuzer, M.; Hawlina, M.; Andjelic, S. UV Effect on Human Anterior Lens Capsule Macro-Molecular Composition Studied by Synchrotron-Based FTIR Micro-Spectroscopy. Int. J. Mol. Sci. 2021, 22, 5249. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andjelic, S.; Kreuzer, M.; Hawlina, M.; Lumi, X. Characterization of Different Types of Epiretinal Proliferations by Synchrotron Radiation-Based Fourier Transform Infrared Micro-Spectroscopy. Int. J. Mol. Sci. 2023, 24, 4834. https://doi.org/10.3390/ijms24054834
Andjelic S, Kreuzer M, Hawlina M, Lumi X. Characterization of Different Types of Epiretinal Proliferations by Synchrotron Radiation-Based Fourier Transform Infrared Micro-Spectroscopy. International Journal of Molecular Sciences. 2023; 24(5):4834. https://doi.org/10.3390/ijms24054834
Chicago/Turabian StyleAndjelic, Sofija, Martin Kreuzer, Marko Hawlina, and Xhevat Lumi. 2023. "Characterization of Different Types of Epiretinal Proliferations by Synchrotron Radiation-Based Fourier Transform Infrared Micro-Spectroscopy" International Journal of Molecular Sciences 24, no. 5: 4834. https://doi.org/10.3390/ijms24054834
APA StyleAndjelic, S., Kreuzer, M., Hawlina, M., & Lumi, X. (2023). Characterization of Different Types of Epiretinal Proliferations by Synchrotron Radiation-Based Fourier Transform Infrared Micro-Spectroscopy. International Journal of Molecular Sciences, 24(5), 4834. https://doi.org/10.3390/ijms24054834