


Transient Receptor Potential (TRP) Channels in Pain, Neuropsychiatric Disorders, and Epilepsy



Abstract
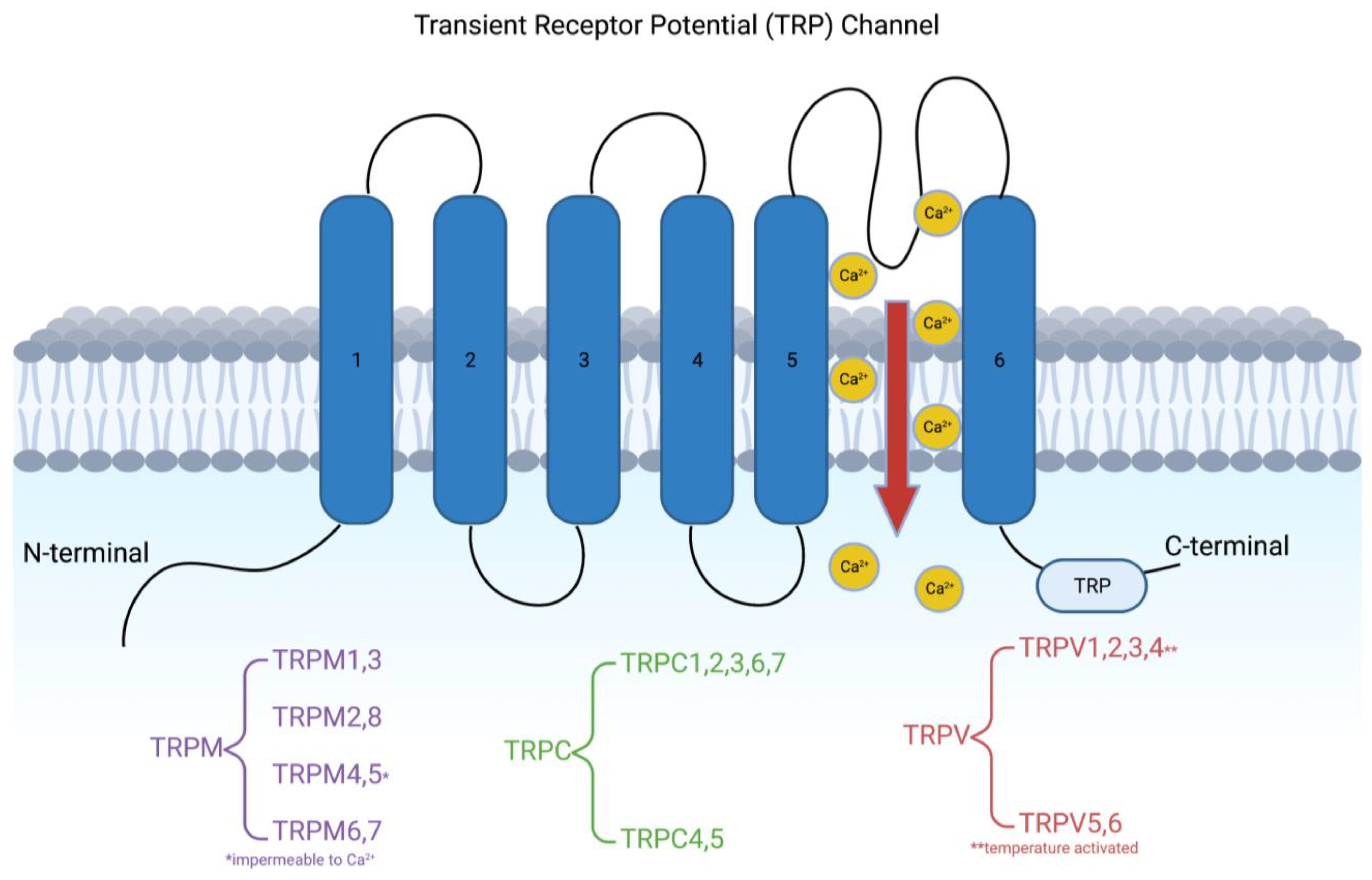
1. Introduction

{kind=link}
| TRP Channel Subfamily | Pain Disorders | Psychiatric Disorders | Epileptic Disorders |
|---|---|---|---|
| TRPM |
| ||
| TRPV |
|
| |
| TRPC |
2. TRPM
2.1. Pain Disorders
2.2. Psychiatric Disorders
2.3. Epileptic Disorders
3. TRPV
3.1. Pain Disorders
3.2. Psychiatric Disorders
3.3. Epileptic Disorders
4. TRPC
4.1. Pain Disorders
4.2. Psychiatric Disorders
4.3. Epileptic Disorders
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fallah, H.P.; Ahuja, E.; Lin, H.; Qi, J.; He, Q.; Gao, S.; An, H.; Zhang, J.; Xie, Y.; Liang, D. A Review on the Role of TRP Channels and Their Potential as Drug Targets_An Insight into the TRP Channel Drug Discovery Methodologies. Front. Pharmacol. 2022, 13, 914499. [Google Scholar] [CrossRef] [PubMed]
- Cosens, D.J.; Manning, A. Abnormal Electroretinogram from a Drosophila Mutant. Nature 1969, 224, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Samanta, A.; Hughes, T.E.T.; Moiseenkova-Bell, V.Y. Transient Receptor Potential (TRP) Channels. Subcell. Biochem. 2018, 87, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Tu, S.; Zhang, J.; Shao, A. Roles of TRP Channels in Neurological Diseases. Oxidative Med. Cell. Longev. 2020, 2020, 7289194. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Fliegert, R.; Guse, A.H.; Lü, W.; Du, J. A Structural Overview of the Ion Channels of the TRPM Family. Cell Calcium 2020, 85, 102111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Papadopoulos, P.; Hamel, E. Endothelial TRPV4 Channels Mediate Dilation of Cerebral Arteries: Impairment and Recovery in Cerebrovascular Pathologies Related to Alzheimer’s Disease. Br. J. Pharmacol. 2013, 170, 661–670. [Google Scholar] [CrossRef]
- Takayama, Y.; Derouiche, S.; Maruyama, K.; Tominaga, M. Emerging Perspectives on Pain Management by Modulation of TRP Channels and ANO1. Int. J. Mol. Sci. 2019, 20, 3411. [Google Scholar] [CrossRef]
- García-Rodríguez, C.; Bravo-Tobar, I.D.; Duarte, Y.; Barrio, L.C.; Sáez, J.C. Contribution of Non-Selective Membrane Channels and Receptors in Epilepsy. Pharmacol. Ther. 2022, 231, 107980. [Google Scholar] [CrossRef]
- Derbenev, A.V.; Zsombok, A. Potential Therapeutic Value of TRPV1 and TRPA1 in Diabetes Mellitus and Obesity. Semin. Immunopathol. 2016, 38, 397–406. [Google Scholar] [CrossRef]
- Morelli, M.B.; Amantini, C. Transient Receptor Potential (TRP) Channels: Markers and Therapeutic Targets for Cancer? Biomolecules 2022, 12, 547. [Google Scholar] [CrossRef]
- Souza Bomfim, G.H.; Niemeyer, B.A.; Lacruz, R.S.; Lis, A. On the Connections between TRPM Channels and SOCE. Cells 2022, 11, 1190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tóth, B.; Szollosi, A.; Chen, J.; Csanády, L. Structure of a TRPM2 Channel in Complex with Ca2+ Explains Unique Gating Regulation. eLife 2018, 7, e36409. [Google Scholar] [CrossRef]
- Yin, Y.; Le, S.C.; Hsu, A.L.; Borgnia, M.J.; Yang, H.; Lee, S.-Y. Structural Basis of Cooling Agent and Lipid Sensing by the Cold-Activated TRPM8 Channel. Science 2019, 363, eaav9334. [Google Scholar] [CrossRef] [PubMed]
- Winkler, P.A.; Huang, Y.; Sun, W.; Du, J.; Lü, W. Electron Cryo-Microscopy Structure of a Human TRPM4 Channel. Nature 2017, 552, 200–204. [Google Scholar] [CrossRef]
- Montell, C.; Birnbaumer, L.; Flockerzi, V.; Bindels, R.J.; Bruford, E.A.; Caterina, M.J.; Clapham, D.E.; Harteneck, C.; Heller, S.; Julius, D.; et al. A unifed nomenclature for the superfamily of TRP cation channels. Mol. Cell 2002, 9, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Du, W.; Yao, H.; Wang, Y. TRPC Channels in Neuronal Survival. In TRP Channels; Zhu, M.X., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2011; ISBN 978-1-4398-1860-2. [Google Scholar]
- Liu, X.; Yao, X.; Tsang, S.Y. Post-Translational Modification and Natural Mutation of TRPC Channels. Cells 2020, 9, 135. [Google Scholar] [CrossRef]
- Wang, H.; Cheng, X.; Tian, J.; Xiao, Y.; Tian, T.; Xu, F.; Hong, X.; Zhu, M.X. TRPC Channels: Structure, Function, Regulation and Recent Advances in Small Molecular Probes. Pharmacol. Ther. 2020, 209, 107497. [Google Scholar] [CrossRef]
- Martín-Bórnez, M.; Galeano-Otero, I.; Del Toro, R.; Smani, T. TRPC and TRPV Channels’ Role in Vascular Remodeling and Disease. Int. J. Mol. Sci. 2020, 21, 6125. [Google Scholar] [CrossRef]
- Hall, G.; Wang, L.; Spurney, R.F. TRPC Channels in Proteinuric Kidney Diseases. Cells 2019, 9, 44. [Google Scholar] [CrossRef]
- Wen, H.; Gwathmey, J.K.; Xie, L.-H. Role of Transient Receptor Potential Canonical Channels in Heart Physiology and Pathophysiology. Front. Cardiovasc. Med. 2020, 7, 24. [Google Scholar] [CrossRef]
- TRPC Channels as Emerging Targets for Seizure Disorders: Trends in Pharmacological Sciences. Available online: https://www.cell.com/trends/pharmacological-sciences/fulltext/S0165-6147(22)00134-1#t0010 (accessed on 10 November 2022).
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 Structures in Distinct Conformations Reveal Activation Mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The Capsaicin Receptor: A Heat-Activated Ion Channel in the Pain Pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Yelshanskaya, M.V.; Sobolevsky, A.I. Ligand-Binding Sites in Vanilloid-Subtype TRP Channels. Front. Pharmacol. 2022, 13, 900623. [Google Scholar] [CrossRef]
- Xie, Q.; Ma, R.; Li, H.; Wang, J.; Guo, X.; Chen, H. Advancement in Research on the Role of the Transient Receptor Potential Vanilloid Channel in Cerebral Ischemic Injury (Review). Exp. Ther. Med. 2021, 22, 881. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, P.K.; Jaggi, A.S. A Review on Potential Involvement of TRPV1 Channels in Ischemia-Reperfusion Injury. J. Cardiovasc. Pharmacol. Ther. 2018, 23, 38–45. [Google Scholar] [CrossRef] [PubMed]
- De Caro, C.; Cristiano, C.; Avagliano, C.; Bertamino, A.; Ostacolo, C.; Campiglia, P.; Gomez-Monterrey, I.; La Rana, G.; Gualillo, O.; Calignano, A.; et al. Characterization of New TRPM8 Modulators in Pain Perception. Int. J. Mol. Sci. 2019, 20, 5544. [Google Scholar] [CrossRef]
- Xu, D.H.; Cullen, B.D.; Tang, M.; Fang, Y. The Effectiveness of Topical Cannabidiol Oil in Symptomatic Relief of Peripheral Neuropathy of the Lower Extremities. Curr. Pharm. Biotechnol. 2020, 21, 390–402. [Google Scholar] [CrossRef]
- Kashio, M.; Sokabe, T.; Shintaku, K.; Uematsu, T.; Fukuta, N.; Kobayashi, N.; Mori, Y.; Tominaga, M. Redox signal-mediated sensitization of transient receptor potential melastatin 2 (TRPM2) to temperature affects macrophage functions. Proc. Natl. Acad. Sci. USA 2012, 109, 6745–6750. [Google Scholar] [CrossRef]
- Jang, Y.; Cho, P.S.; Yang, Y.D.; Hwang, S.W. Nociceptive Roles of TRPM2 Ion Channel in Pathologic Pain. Mol. Neurobiol. 2018, 55, 6589–6600. [Google Scholar] [CrossRef]
- Haraguchi, K.; Kawamoto, A.; Isami, K.; Maeda, S.; Kusano, A.; Asakura, K.; Shirakawa, H.; Mori, Y.; Nakagawa, T.; Kaneko, S. TRPM2 Contributes to Inflammatory and Neuropathic Pain through the Aggravation of Pronociceptive Inflammatory Responses in Mice. J. Neurosci. 2012, 32, 3931–3941. [Google Scholar] [CrossRef]
- Straub, I.; Krügel, U.; Mohr, F.; Teichert, J.; Rizun, O.; Konrad, M.; Oberwinkler, J.; Schaefer, M. Flavanones That Selectively Inhibit TRPM3 Attenuate Thermal Nociception in Vivo. Mol. Pharmacol. 2013, 84, 736–750. [Google Scholar] [CrossRef]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef]
- Lee, C.R.; Machold, R.P.; Witkovsky, P.; Rice, M.E. TRPM2 Channels Are Required for NMDA-Induced Burst Firing and Contribute to H(2)O(2)-Dependent Modulation in Substantia Nigra Pars Reticulata GABAergic Neurons. J. Neurosci. 2013, 33, 1157–1168. [Google Scholar] [CrossRef]
- McQuillin, A.; Bass, N.J.; Kalsi, G.; Lawrence, J.; Puri, V.; Choudhury, K.; Detera-Wadleigh, S.D.; Curtis, D.; Gurling, H.M.D. Fine Mapping of a Susceptibility Locus for Bipolar and Genetically Related Unipolar Affective Disorders, to a Region Containing the C21ORF29 and TRPM2 Genes on Chromosome 21q22.3. Mol. Psychiatry 2006, 11, 134–142. [Google Scholar] [CrossRef]
- Jang, Y.; Lee, S.H.; Lee, B.; Jung, S.; Khalid, A.; Uchida, K.; Tominaga, M.; Jeon, D.; Oh, U. TRPM2, a Susceptibility Gene for Bipolar Disorder, Regulates Glycogen Synthase Kinase-3 Activity in the Brain. J. Neurosci. 2015, 35, 11811–11823. [Google Scholar] [CrossRef]
- Andoh, C.; Nishitani, N.; Hashimoto, E.; Nagai, Y.; Takao, K.; Miyakawa, T.; Nakagawa, T.; Mori, Y.; Nagayasu, K.; Shirakawa, H.; et al. TRPM2 Confers Susceptibility to Social Stress but Is Essential for Behavioral Flexibility. Brain Res. 2019, 1704, 68–77. [Google Scholar] [CrossRef]
- Nazıroğlu, M. TRPV1 Channel: A Potential Drug Target for Treating Epilepsy. Curr. Neuropharmacol. 2015, 13, 239–247. [Google Scholar] [CrossRef]
- Katano, M.; Numata, T.; Aguan, K.; Hara, Y.; Kiyonaka, S.; Yamamoto, S.; Miki, T.; Sawamura, S.; Suzuki, T.; Yamakawa, K.; et al. The Juvenile Myoclonic Epilepsy-Related Protein EFHC1 Interacts with the Redox-Sensitive TRPM2 Channel Linked to Cell Death. Cell Calcium 2012, 51, 179–185. [Google Scholar] [CrossRef]
- Zhao, S.; Rohacs, T. The Newest TRP Channelopathy: Gain of Function TRPM3 Mutations Cause Epilepsy and Intellectual Disability. Channels 2021, 15, 386–397. [Google Scholar] [CrossRef]
- Grimm, C.; Kraft, R.; Sauerbruch, S.; Schultz, G.; Harteneck, C. Molecular and Functional Characterization of the Melastatin-Related Cation Channel TRPM3. J. Biol. Chem. 2003, 278, 21493–21501. [Google Scholar] [CrossRef]
- Aarts, M.M.; Tymianski, M. TRPMs and Neuronal Cell Death. Pflüg. Arch. 2005, 451, 243–249. [Google Scholar] [CrossRef]
- McGaraughty, S.; Chu, K.L.; Faltynek, C.R.; Jarvis, M.F. Systemic and Site-Specific Effects of A-425619, a Selective TRPV1 Receptor Antagonist, on Wide Dynamic Range Neurons in CFA-Treated and Uninjured Rats. J. Neurophysiol. 2006, 95, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Chizh, B.A.; O’Donnell, M.B.; Napolitano, A.; Wang, J.; Brooke, A.C.; Aylott, M.C.; Bullman, J.N.; Gray, E.J.; Lai, R.Y.; Williams, P.M.; et al. The Effects of the TRPV1 Antagonist SB-705498 on TRPV1 Receptor-Mediated Activity and Inflammatory Hyperalgesia in Humans. Pain 2007, 132, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.; Chapman, R.J.; Woodhams, S.; Sagar, D.R.; Turner, J.; Burston, J.J.; Bullock, C.; Paton, K.; Huang, J.; Wong, A.; et al. Increased Function of Pronociceptive TRPV1 at the Level of the Joint in a Rat Model of Osteoarthritis Pain. Ann. Rheum. Dis. 2015, 74, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Anand, U.; Jones, B.; Korchev, Y.; Bloom, S.R.; Pacchetti, B.; Anand, P.; Sodergren, M.H. CBD Effects on TRPV1 Signaling Pathways in Cultured DRG Neurons. J. Pain Res. 2020, 13, 2269–2278. [Google Scholar] [CrossRef] [PubMed]
- Nazıroğlu, M.; Demirdaş, A. Psychiatric Disorders and TRP Channels: Focus on Psychotropic Drugs. Curr. Neuropharmacol. 2015, 13, 248–257. [Google Scholar] [CrossRef]
- Marsch, R.; Foeller, E.; Rammes, G.; Bunck, M.; Kössl, M.; Holsboer, F.; Zieglgänsberger, W.; Landgraf, R.; Lutz, B.; Wotjak, C.T. Reduced Anxiety, Conditioned Fear, and Hippocampal Long-Term Potentiation in Transient Receptor Potential Vanilloid Type 1 Receptor-Deficient Mice. J. Neurosci. 2007, 27, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Kasckow, J.W.; Mulchahey, J.J.; Geracioti, T.D. Effects of the Vanilloid Agonist Olvanil and Antagonist Capsazepine on Rat Behaviors. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2004, 28, 291–295. [Google Scholar] [CrossRef]
- Santos, C.J.P.A.; Stern, C.A.J.; Bertoglio, L.J. Attenuation of Anxiety-Related Behaviour after the Antagonism of Transient Receptor Potential Vanilloid Type 1 Channels in the Rat Ventral Hippocampus. Behav. Pharmacol. 2008, 19, 357–360. [Google Scholar] [CrossRef]
- Aguiar, D.C.; Almeida-Santos, A.F.; Moreira, F.A.; Guimarães, F.S. Involvement of TRPV1 Channels in the Periaqueductal Grey on the Modulation of Innate Fear Responses. Acta Neuropsychiatr. 2015, 27, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Mezey, É.; Tóth, Z.E.; Cortright, D.N.; Arzubi, M.K.; Krause, J.E.; Elde, R.; Guo, A.; Blumberg, P.M.; Szallasi, A. Distribution of MRNA for Vanilloid Receptor Subtype 1 (VR1), and VR1-like Immunoreactivity, in the Central Nervous System of the Rat and Human. Proc. Natl. Acad. Sci. USA 2000, 97, 3655–3660. [Google Scholar] [CrossRef]
- Jia, Y.-F.; Li, Y.-C.; Tang, Y.-P.; Cao, J.; Wang, L.-P.; Yang, Y.-X.; Xu, L.; Mao, R.-R. Interference of TRPV1 Function Altered the Susceptibility of PTZ-Induced Seizures. Front. Cell. Neurosci. 2015, 9, 20. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.-J.; Guo, W.; Zheng, D.-H.; Zhang, C.-Q.; Li, S.; Liu, S.-Y.; Yin, Q.; Yang, H.; Shu, H.-F. Increased Expression of TRPV1 in the Cortex and Hippocampus from Patients with Mesial Temporal Lobe Epilepsy. J. Mol. Neurosci. 2013, 49, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Alawi, K.M.; Russell, F.A.; Aubdool, A.A.; Srivastava, S.; Riffo-Vasquez, Y.; Baldissera, L.; Thakore, P.; Saleque, N.; Fernandes, E.S.; Walsh, D.A.; et al. Transient Receptor Potential Canonical 5 (TRPC5) Protects against Pain and Vascular Inflammation in Arthritis and Joint Inflammation. Ann. Rheum. Dis. 2017, 76, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Lawhorn, B.G.; Brnardic, E.J.; Behm, D.J. Recent Advances in TRPV4 Agonists and Antagonists. Bioorg. Med. Chem. Lett. 2020, 30, 127022. [Google Scholar] [CrossRef]
- Sadler, K.E.; Moehring, F.; Shiers, S.I.; Laskowski, L.J.; Mikesell, A.R.; Plautz, Z.R.; Brezinski, A.N.; Mecca, C.M.; Dussor, G.; Price, T.J.; et al. Transient Receptor Potential Canonical 5 Mediates Inflammatory Mechanical and Spontaneous Pain in Mice. Sci. Transl. Med. 2021, 13, eabd7702. [Google Scholar] [CrossRef]
- Yang, L.; Liu, C.; Li, W.; Ma, Y.; Huo, S.; Ozathaley, A.; Ren, J.; Yuan, W.; Ni, H.; Li, D.; et al. Depression-like Behavior Associated with E/I Imbalance of MPFC and Amygdala without TRPC Channels in Mice of Knockout IL-10 from Microglia. Brain Behav. Immun. 2021, 97, 68–78. [Google Scholar] [CrossRef]
- Just, S.; Chenard, B.L.; Ceci, A.; Strassmaier, T.; Chong, J.A.; Blair, N.T.; Gallaschun, R.J.; Del Camino, D.; Cantin, S.; D’Amours, M.; et al. Treatment with HC-070, a Potent Inhibitor of TRPC4 and TRPC5, Leads to Anxiolytic and Antidepressant Effects in Mice. PLoS ONE 2018, 13, e0191225. [Google Scholar] [CrossRef]
- Nagib, M.M.; Zhang, S.; Yasmen, N.; Li, L.; Hou, R.; Yu, Y.; Boda, V.K.; Wu, Z.; Li, W.; Jiang, J. Inhibition of TRPC3 Channels by a Novel Pyrazole Compound Confers Antiseizure Effects. Epilepsia 2022, 63, 1003–1015. [Google Scholar] [CrossRef]
- Lee, K.; Jo, Y.Y.; Chung, G.; Jung, J.H.; Kim, Y.H.; Park, C.-K. Functional Importance of Transient Receptor Potential (TRP) Channels in Neurological Disorders. Front. Cell Dev. Biol. 2021, 9, 611773. [Google Scholar] [CrossRef]
- Zang, Z.; Li, S.; Zhang, W.; Chen, X.; Zheng, D.; Shu, H.; Guo, W.; Zhao, B.; Shen, K.; Wei, Y.; et al. Expression Patterns of TRPC1 in Cortical Lesions from Patients with Focal Cortical Dysplasia. J. Mol. Neurosci. 2015, 57, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Yam, M.F.; Loh, Y.C.; Tan, C.S.; Khadijah Adam, S.; Abdul Manan, N.; Basir, R. General Pathways of Pain Sensation and the Major Neurotransmitters Involved in Pain Regulation. Int. J. Mol. Sci. 2018, 19, 2164. [Google Scholar] [CrossRef] [PubMed]
- Pitake, S.; Middleton, L.J.; Abdus-Saboor, I.; Mishra, S.K. Inflammation Induced Sensory Nerve Growth and Pain Hypersensitivity Requires the N-Type Calcium Channel Cav2.2. Front. Neurosci. 2019, 13, 1009. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, I.; Prado, Y.; Marchant, F.; Otero, C.; Eltit, F.; Cabello-Verrugio, C.; Cerda, O.; Simon, F. TRPM Channels in Human Diseases. Cells 2020, 9, 2604. [Google Scholar] [CrossRef]
- Behrendt, M. Transient Receptor Potential Channels in the Context of Nociception and Pain—Recent Insights into TRPM3 Properties and Function. Biol. Chem. 2019, 400, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Owsianik, G.; Hofmann, T.; Philipp, S.E.; Stab, J.; Chen, X.; Benoit, M.; Xue, F.; Janssens, A.; Kerselaers, S.; et al. TRPM3 Is a Nociceptor Channel Involved in the Detection of Noxious Heat. Neuron 2011, 70, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Descoeur, J.; Pereira, V.; Pizzoccaro, A.; Francois, A.; Ling, B.; Maffre, V.; Couette, B.; Busserolles, J.; Courteix, C.; Noel, J.; et al. Oxaliplatin-Induced Cold Hypersensitivity Is Due to Remodelling of Ion Channel Expression in Nociceptors. EMBO Mol. Med. 2011, 3, 266–278. [Google Scholar] [CrossRef]
- Burgos-Vega, C.C.; Ahn, D.D.-U.; Bischoff, C.; Wang, W.; Horne, D.; Wang, J.; Gavva, N.; Dussor, G. Meningeal Transient Receptor Potential Channel M8 Activation Causes Cutaneous Facial and Hindpaw Allodynia in a Preclinical Rodent Model of Headache. Cephalalgia 2016, 36, 185–193. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, J.; Zhou, F.; Zhang, P. Increased Transient Receptor Potential Melastatin 8 Expression in the Development of Bladder Pain in Patients with Interstitial Cystitis/Bladder Pain Syndrome. Urology 2020, 146, 301.e1–301.e6. [Google Scholar] [CrossRef]
- Mukerji, G.; Yiangou, Y.; Corcoran, S.L.; Selmer, I.S.; Smith, G.D.; Benham, C.D.; Bountra, C.; Agarwal, S.K.; Anand, P. Cool and Menthol Receptor TRPM8 in Human Urinary Bladder Disorders and Clinical Correlations. BMC Urol. 2006, 6, 6. [Google Scholar] [CrossRef]
- Nakao, A.; Matsunaga, Y.; Hayashida, K.; Takahashi, N. Role of Oxidative Stress and Ca2+ Signaling in Psychiatric Disorders. Front. Cell Dev. Biol. 2021, 9, 615569. [Google Scholar] [CrossRef]
- Rossetti, A.C.; Paladini, M.S.; Riva, M.A.; Molteni, R. Oxidation-Reduction Mechanisms in Psychiatric Disorders: A Novel Target for Pharmacological Intervention. Pharmacol. Ther. 2020, 210, 107520. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, B.S.; Dean, O.M.; Dodd, S.; Malhi, G.S.; Berk, M. N-Acetylcysteine in Depressive Symptoms and Functionality: A Systematic Review and Meta-Analysis. J. Clin. Psychiatry 2016, 77, e457–e466. [Google Scholar] [CrossRef] [PubMed]
- Sepehrmanesh, Z.; Heidary, M.; Akasheh, N.; Akbari, H.; Heidary, M. Therapeutic Effect of Adjunctive N-Acetyl Cysteine (NAC) on Symptoms of Chronic Schizophrenia: A Double-Blind, Randomized Clinical Trial. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2018, 82, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, R.; Mori, Y. Transient Receptor Potential (TRP) Channels: Biosensors for Redox Environmental Stimuli and Cellular Status. Free Radic. Biol. Med. 2020, 146, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Olah, M.E.; Jackson, M.F.; Li, H.; Perez, Y.; Sun, H.-S.; Kiyonaka, S.; Mori, Y.; Tymianski, M.; MacDonald, J.F. Ca2+-Dependent Induction of TRPM2 Currents in Hippocampal Neurons. J. Physiol. 2009, 587, 965–979. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.-F.; Belrose, J.C.; Lei, G.; Tymianski, M.; Mori, Y.; Macdonald, J.F.; Jackson, M.F. Dependence of NMDA/GSK-3β Mediated Metaplasticity on TRPM2 Channels at Hippocampal CA3-CA1 Synapses. Mol. Brain 2011, 4, 44. [Google Scholar] [CrossRef] [PubMed]
- Thippeswamy, H.; Davies, W. A New Molecular Risk Pathway for Postpartum Mood Disorders: Clues from Steroid Sulfatase-Deficient Individuals. Arch. Women’s Ment. Health 2021, 24, 391–401. [Google Scholar] [CrossRef]
- Maddaloni, G.; Migliarini, S.; Napolitano, F.; Giorgi, A.; Nazzi, S.; Biasci, D.; De Felice, A.; Gritti, M.; Cavaccini, A.; Galbusera, A.; et al. Serotonin Depletion Causes Valproate-Responsive Manic-like Condition and Increased Hippocampal Neuroplasticity That Are Reversed by Stress. Sci. Rep. 2018, 8, 11847. [Google Scholar] [CrossRef]
- Steinlein, O.K. Calcium Signaling and Epilepsy. Cell Tissue Res. 2014, 357, 385–393. [Google Scholar] [CrossRef]
- Tucholski, J.; Roth, K.A.; Johnson, G.V.W. Tissue Transglutaminase Overexpression in the Brain Potentiates Calcium-Induced Hippocampal Damage. J. Neurochem. 2006, 97, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Arundine, M.; Tymianski, M. Molecular Mechanisms of Calcium-Dependent Neurodegeneration in Excitotoxicity. Cell Calcium 2003, 34, 325–337. [Google Scholar] [CrossRef]
- Muona, M.; Berkovic, S.F.; Dibbens, L.M.; Oliver, K.L.; Maljevic, S.; Bayly, M.A.; Joensuu, T.; Canafoglia, L.; Franceschetti, S.; Michelucci, R.; et al. A Recurrent de Novo Mutation in KCNC1 Causes Progressive Myoclonus Epilepsy. Nat. Genet. 2015, 47, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Wengert, E.R.; Patel, M.K. The Role of the Persistent Sodium Current in Epilepsy. Epilepsy Curr. 2020, 21, 40–47. [Google Scholar] [CrossRef]
- Jeong, J.H.; Lee, S.H.; Kho, A.R.; Hong, D.K.; Kang, D.H.; Kang, B.S.; Park, M.K.; Choi, B.Y.; Choi, H.C.; Lim, M.-S.; et al. The Transient Receptor Potential Melastatin 7 (TRPM7) Inhibitors Suppress Seizure-Induced Neuron Death by Inhibiting Zinc Neurotoxicity. Int. J. Mol. Sci. 2020, 21, 7897. [Google Scholar] [CrossRef]
- Stawicki, T.M.; Zhou, K.; Yochem, J.; Chen, L.; Jin, Y. TRPM Channels Modulate Epileptic-like Convulsions via Systemic Ion Homeostasis. Curr. Biol. 2011, 21, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Morgans, C.W.; Zhang, J.; Jeffrey, B.G.; Nelson, S.M.; Burke, N.S.; Duvoisin, R.M.; Brown, R.L. TRPM1 Is Required for the Depolarizing Light Response in Retinal ON-Bipolar Cells. Proc. Natl. Acad. Sci. USA 2009, 106, 19174–19178. [Google Scholar] [CrossRef]
- Jara-Oseguera, A.; Simon, S.A.; Rosenbaum, T. TRPV1: On the Road to Pain Relief. Curr. Mol. Pharmacol. 2008, 1, 255–269. [Google Scholar] [CrossRef]
- Pickering, G.; Creveaux, I.; Macian, N.; Pereira, B. Paracetamol and Pain Modulation by TRPV1, UGT2B15, SULT1A1 Genotypes: A Randomized Clinical Trial in Healthy Volunteers. Pain Med. 2020, 21, 661–669. [Google Scholar] [CrossRef]
- Marrone, M.C.; Morabito, A.; Giustizieri, M.; Chiurchiù, V.; Leuti, A.; Mattioli, M.; Marinelli, S.; Riganti, L.; Lombardi, M.; Murana, E.; et al. TRPV1 Channels Are Critical Brain Inflammation Detectors and Neuropathic Pain Biomarkers in Mice. Nat. Commun. 2017, 8, 15292. [Google Scholar] [CrossRef]
- Kim, Y.H.; Back, S.K.; Davies, A.J.; Jeong, H.; Jo, H.J.; Chung, G.; Na, H.S.; Bae, Y.C.; Kim, S.J.; Kim, J.S.; et al. TRPV1 in GABAergic Interneurons Mediates Neuropathic Mechanical Allodynia and Disinhibition of the Nociceptive Circuitry in the Spinal Cord. Neuron 2012, 74, 640–647. [Google Scholar] [CrossRef]
- Kim, Y.S.; Chu, Y.; Han, L.; Li, M.; Li, Z.; LaVinka, P.C.; Sun, S.; Tang, Z.; Park, K.; Caterina, M.J.; et al. Central Terminal Sensitization of TRPV1 by Descending Serotonergic Facilitation Modulates Chronic Pain. Neuron 2014, 81, 873–887. [Google Scholar] [CrossRef]
- Schilling, J.M.; Hughes, C.G.; Wallace, M.S.; Sexton, M.; Backonja, M.; Moeller-Bertram, T. Cannabidiol as a Treatment for Chronic Pain: A Survey of Patients’ Perspectives and Attitudes. J. Pain Res. 2021, 14, 1241–1250. [Google Scholar] [CrossRef]
- Mnich, K.; Finn, D.P.; Dowd, E.; Gorman, A.M. Inhibition by Anandamide of 6-Hydroxydopamine-Induced Cell Death in PC12 Cells. Int. J. Cell Biol. 2010, 2010, 818497. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Lee, S.-H.; Kim, K.-T.; Zeng, X.; Han, I. TRPV4: A Sensor for Homeostasis and Pathological Events in the CNS. Mol. Neurobiol. 2018, 55, 8695–8708. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Meng, H.; Inamdar, S.; Das, B.; Gupta, H.; Wang, W.; Thompson, C.L.; Knight, M.M. Activation of TRPV4 by Mechanical, Osmotic or Pharmaceutical Stimulation Is Anti-Inflammatory Blocking IL-1β Mediated Articular Cartilage Matrix Destruction. Osteoarthr. Cartil. 2021, 29, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.-L.; Wang, Y.-H.; Ning, L.-P.; Zhang, Y.; Ge, H.-Y.; Jiang, H.; Wang, R.; Yue, S.-W. Involvement of TRPV4-NO-CGMP-PKG Pathways in the Development of Thermal Hyperalgesia Following Chronic Compression of the Dorsal Root Ganglion in Rats. Behav. Brain Res. 2010, 208, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.-Y.; Yang, J.; Li, H.-M.; Hu, S.-J.; Xing, J.-L. Dorsal Root Ganglion Compression as an Animal Model of Sciatica and Low Back Pain. Neurosci. Bull. 2012, 28, 618–630. [Google Scholar] [CrossRef]
- Brooks, C.A.; Barton, L.S.; Behm, D.J.; Brnardic, E.J.; Costell, M.H.; Holt, D.A.; Jolivette, L.J.; Matthews, J.M.; McAtee, J.J.; McCleland, B.W.; et al. Discovery of GSK3527497: A Candidate for the Inhibition of Transient Receptor Potential Vanilloid-4 (TRPV4). J. Med. Chem. 2019, 62, 9270–9280. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.-H.; Lee, S.-Y.; Kim, H.-C.; Jang, C.-G. Repeated Methamphetamine Treatment Increases Expression of TRPV1 MRNA in the Frontal Cortex but Not in the Striatum or Hippocampus of Mice. Neurosci. Lett. 2010, 472, 61–64. [Google Scholar] [CrossRef]
- Blednov, Y.A.; Harris, R.A. Deletion of Vanilloid Receptor (TRPV1) in Mice Alters Behavioral Effects of Ethanol. Neuropharmacology 2009, 56, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.S.S.; Umathe, S.N. Involvement of Transient Receptor Potential Vanilloid Type 1 Channels in the Pro-Convulsant Effect of Anandamide in Pentylenetetrazole-Induced Seizures. Epilepsy Res. 2012, 100, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Nazıroğlu, M.; Övey, İ.S. Involvement of Apoptosis and Calcium Accumulation through TRPV1 Channels in Neurobiology of Epilepsy. Neuroscience 2015, 293, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Neeper, M.P.; Liu, Y.; Hutchinson, T.L.; Lubin, M.L.; Flores, C.M. TRPV2 Is Activated by Cannabidiol and Mediates CGRP Release in Cultured Rat Dorsal Root Ganglion Neurons. J. Neurosci. 2008, 28, 6231–6238. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.A.; Hill, A.J.; Smith, I.; Bevan, S.A.; Williams, C.M.; Whalley, B.J.; Stephens, G.J. Cannabidiol Displays Antiepileptiform and Antiseizure Properties In Vitro and In Vivo. J. Pharmacol. Exp. Ther. 2010, 332, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.; Limjunyawong, N.; Narang, C.; Jamaldeen, H.; Yu, S.; Patiram, S.; Nie, H.; Caterina, M.J.; Dong, X.; et al. Sensory Neuron–Expressed TRPC3 Mediates Acute and Chronic Itch. Pain 2023, 164, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Alkhani, H.; Ase, A.R.; Grant, R.; O’Donnell, D.; Groschner, K.; Séguéla, P. Contribution of TRPC3 to Store-Operated Calcium Entry and Inflammatory Transductions in Primary Nociceptors. Mol. Pain 2014, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Kress, M.; Karasek, J.; Ferrer-Montiel, A.V.; Scherbakov, N.; Haberberger, R.V. TRPC Channels and Diacylglycerol Dependent Calcium Signaling in Rat Sensory Neurons. Histochem. Cell Biol. 2008, 130, 655–667. [Google Scholar] [CrossRef]
- De Sousa Valente, J.; Alawi, K.M.; Keringer, P.; Bharde, S.; Ayaz, F.; Saleque, N.; Kodji, X.; Thapa, D.; Argunhan, F.; Brain, S.D. Examining the Role of Transient Receptor Potential Canonical 5 (TRPC5) in Osteoarthritis. Osteoarthr. Cartil. Open 2020, 2, 100119. [Google Scholar] [CrossRef]
- Riccio, A.; Li, Y.; Moon, J.; Kim, K.-S.; Smith, K.S.; Rudolph, U.; Gapon, S.; Yao, G.L.; Tsvetkov, E.; Rodig, S.J.; et al. Essential Role for TRPC5 in Amygdala Function and Fear-Related Behavior. Cell 2009, 137, 761–772. [Google Scholar] [CrossRef]
- Roedding, A.S.; Gao, A.F.; Au-Yeung, W.; Scarcelli, T.; Li, P.P.; Warsh, J.J. Effect of Oxidative Stress on TRPM2 and TRPC3 Channels in B Lymphoblast Cells in Bipolar Disorder. Bipolar Disord. 2012, 14, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Andreopoulos, S.; Wasserman, M.; Woo, K.; Li, P.P.; Warsh, J.J. Chronic Lithium Treatment of B Lymphoblasts from Bipolar Disorder Patients Reduces Transient Receptor Potential Channel 3 Levels. Pharm. J. 2004, 4, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yu, H.; Xu, B.; Xu, H.; Zhang, Z.; Ren, X.; Yuan, J.; Liu, J.; Guo, Y.; Spencer, P.S.; et al. TRPC1 Deletion Causes Striatal Neuronal Cell Apoptosis and Proteomic Alterations in Mice. Front. Aging Neurosci. 2018, 10, 72. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.-P.; Jiang, F.-J.; Wu, G.-S.; Deng, K.; Wen, M.; Zhou, X.; Hong, X.; Zhu, M.X.; Luo, H.-R. Acute Treatment with a Novel TRPC4/C5 Channel Inhibitor Produces Antidepressant and Anxiolytic-Like Effects in Mice. PLoS ONE 2015, 10, e0136255. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.-W.; Roper, S.N. TRPC3 Mediates Hyperexcitability and Epileptiform Activity in Immature Cortex and Experimental Cortical Dysplasia. J. Neurophysiol. 2014, 111, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Li, W.; Jiang, J. TRPC Channels as Emerging Targets for Seizure Disorders. Trends Pharmacol. Sci. 2022, 43, 787–798. [Google Scholar] [CrossRef]
- Sun, D.; Ma, H.; Ma, J.; Wang, J.; Deng, X.; Hu, C.; Deng, X. Canonical Transient Receptor Potential Channel 3 Contributes to Febrile Seizure Inducing Neuronal Cell Death and Neuroinflammation. Cell. Mol. Neurobiol. 2018, 38, 1215–1226. [Google Scholar] [CrossRef]
- Kim, J.-E.; Park, H.; Choi, S.-H.; Kong, M.-J.; Kang, T.-C. TRPC6-Mediated ERK1/2 Activation Increases Dentate Granule Cell Resistance to Status Epilepticus via Regulating Lon Protease-1 Expression and Mitochondrial Dynamics. Cells 2019, 8, 1376. [Google Scholar] [CrossRef]
- Jardín, I.; López, J.J.; Diez, R.; Sánchez-Collado, J.; Cantonero, C.; Albarrán, L.; Woodard, G.E.; Redondo, P.C.; Salido, G.M.; Smani, T.; et al. TRPs in Pain Sensation. Front. Physiol. 2017, 8, 392. [Google Scholar] [CrossRef]
- Julius, D. TRP Channels and Pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef]
- Vowles, K.E.; McEntee, M.L.; Julnes, P.S.; Frohe, T.; Ney, J.P.; van der Goes, D.N. Rates of Opioid Misuse, Abuse, and Addiction in Chronic Pain: A Systematic Review and Data Synthesis. Pain 2015, 156, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Krebs, E.E.; Gravely, A.; Nugent, S.; Jensen, A.C.; DeRonne, B.; Goldsmith, E.S.; Kroenke, K.; Bair, M.J.; Noorbaloochi, S. Effect of Opioid vs Nonopioid Medications on Pain-Related Function in Patients with Chronic Back Pain or Hip or Knee Osteoarthritis Pain: The SPACE Randomized Clinical Trial. JAMA 2018, 319, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, C.; Martín-Martínez, M.; Gómez-Monterrey, I.; González-Muñiz, R. TRPM8 Channels: Advances in Structural Studies and Pharmacological Modulation. Int. J. Mol. Sci. 2021, 22, 8502. [Google Scholar] [CrossRef]
- Rosenberger, D.C.; Binzen, U.; Treede, R.-D.; Greffrath, W. The Capsaicin Receptor TRPV1 Is the First Line Defense Protecting from Acute Non Damaging Heat: A Translational Approach. J. Transl. Med. 2020, 18, 28. [Google Scholar] [CrossRef] [PubMed]
- Moreau, C.; Kirchberger, T.; Swarbrick, J.M.; Bartlett, S.J.; Fliegert, R.; Yorgan, T.; Bauche, A.; Harneit, A.; Guse, A.H.; Potter, B.V.L. Structure-Activity Relationship of Adenosine 5′-Diphosphoribose at the Transient Receptor Potential Melastatin 2 (TRPM2) Channel: Rational Design of Antagonists. J. Med. Chem. 2013, 56, 10079–10102. [Google Scholar] [CrossRef]
- Hayase, T. Differential Effects of TRPV1 Receptor Ligands against Nicotine-Induced Depression-like Behaviors. BMC Pharmacol. 2011, 11, 6. [Google Scholar] [CrossRef]
- Ho, K.W.; Ward, N.J.; Calkins, D.J. TRPV1: A Stress Response Protein in the Central Nervous System. Am. J. Neurodegener. Dis. 2012, 1, 1–14. [Google Scholar]
- Barker-Haliski, M.; White, H.S. Glutamatergic Mechanisms Associated with Seizures and Epilepsy. Cold Spring Harb. Perspect. Med. 2015, 5, a022863. [Google Scholar] [CrossRef]
- Krügel, U.; Straub, I.; Beckmann, H.; Schaefer, M. Primidone Inhibits TRPM3 and Attenuates Thermal Nociception in Vivo. Pain 2017, 158, 856–867. [Google Scholar] [CrossRef]
- Menigoz, A.; Boudes, M.; Yamada, K.; Cirrito, J.R.; Stewart, F.R.; Jiang, H.; Finn, M.B.; Holmes, B.B.; Binder, L.I.; Mandelkow, E.-M.; et al. The Expression Pattern of TRPV1 in Brain. J. Neurosci. 2011, 31, 13025–13027. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; Sivils, A.; Cegielski, V.; Singh, S.; Chu, X.-P. Transient Receptor Potential (TRP) Channels in Pain, Neuropsychiatric Disorders, and Epilepsy. Int. J. Mol. Sci. 2023, 24, 4714. https://doi.org/10.3390/ijms24054714
Yang F, Sivils A, Cegielski V, Singh S, Chu X-P. Transient Receptor Potential (TRP) Channels in Pain, Neuropsychiatric Disorders, and Epilepsy. International Journal of Molecular Sciences. 2023; 24(5):4714. https://doi.org/10.3390/ijms24054714
Chicago/Turabian StyleYang, Felix, Andy Sivils, Victoria Cegielski, Som Singh, and Xiang-Ping Chu. 2023. "Transient Receptor Potential (TRP) Channels in Pain, Neuropsychiatric Disorders, and Epilepsy" International Journal of Molecular Sciences 24, no. 5: 4714. https://doi.org/10.3390/ijms24054714
APA StyleYang, F., Sivils, A., Cegielski, V., Singh, S., & Chu, X.-P. (2023). Transient Receptor Potential (TRP) Channels in Pain, Neuropsychiatric Disorders, and Epilepsy. International Journal of Molecular Sciences, 24(5), 4714. https://doi.org/10.3390/ijms24054714