

Identification of DNA Methylation Changes in European Beech Seeds during Desiccation and Storage

 , and
, and
Abstract
1. Introduction
2. Results
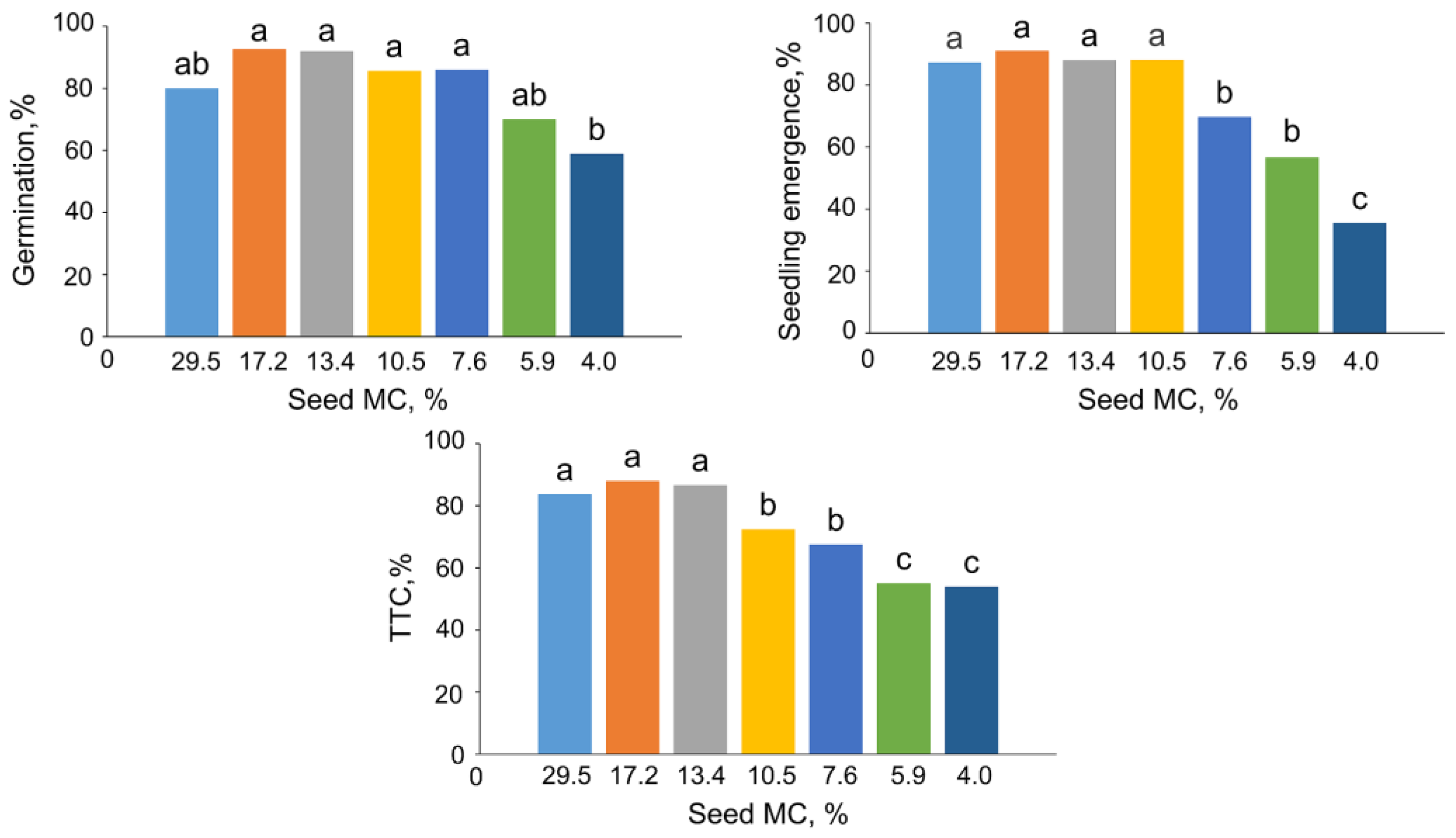
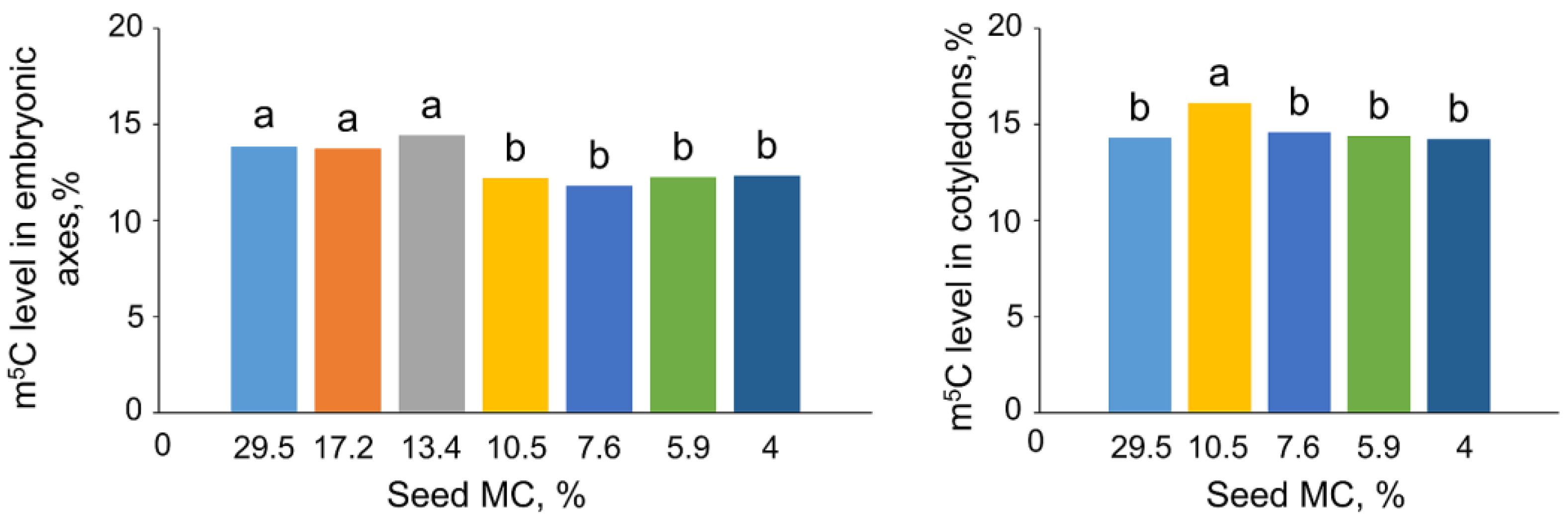
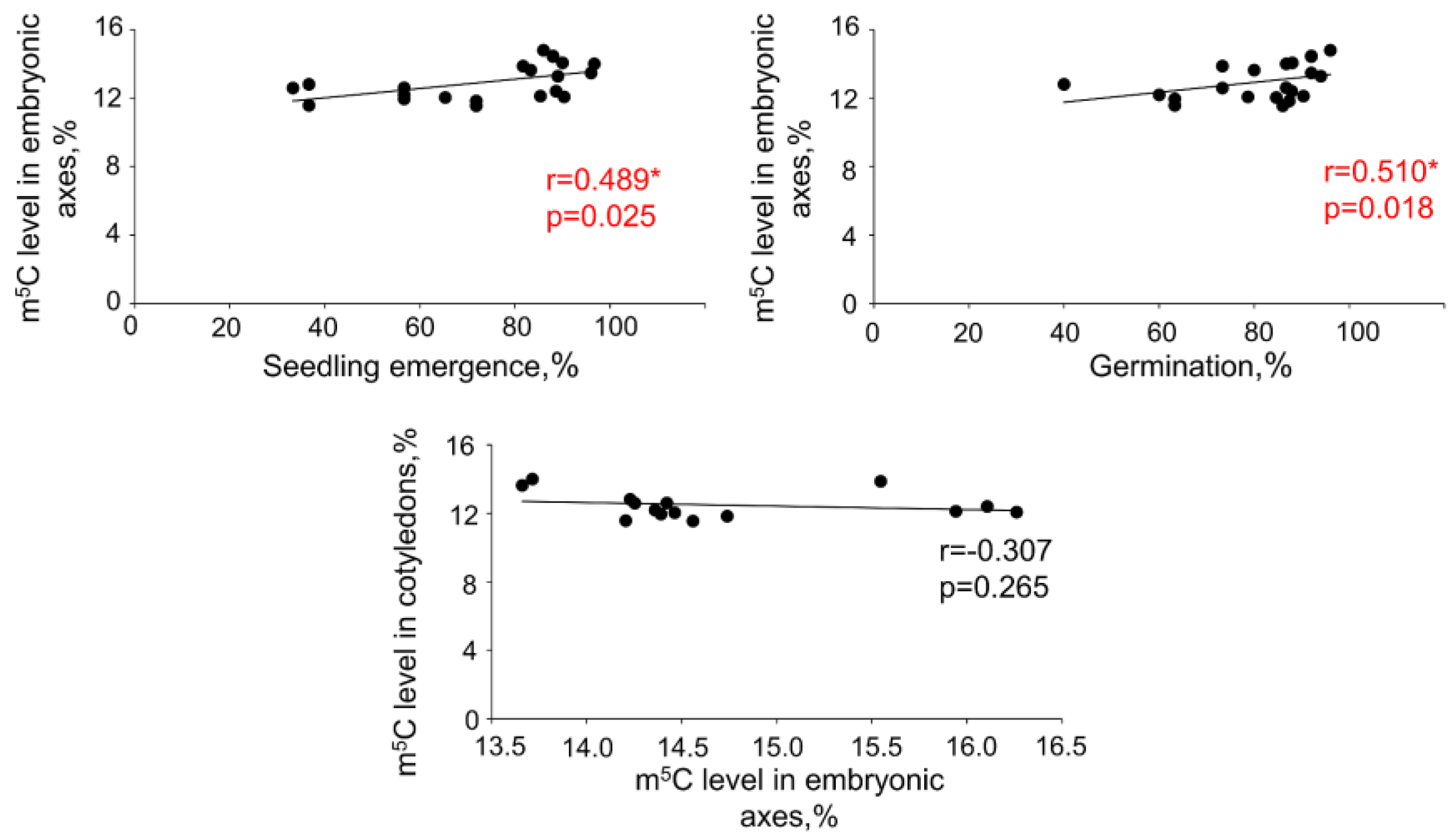
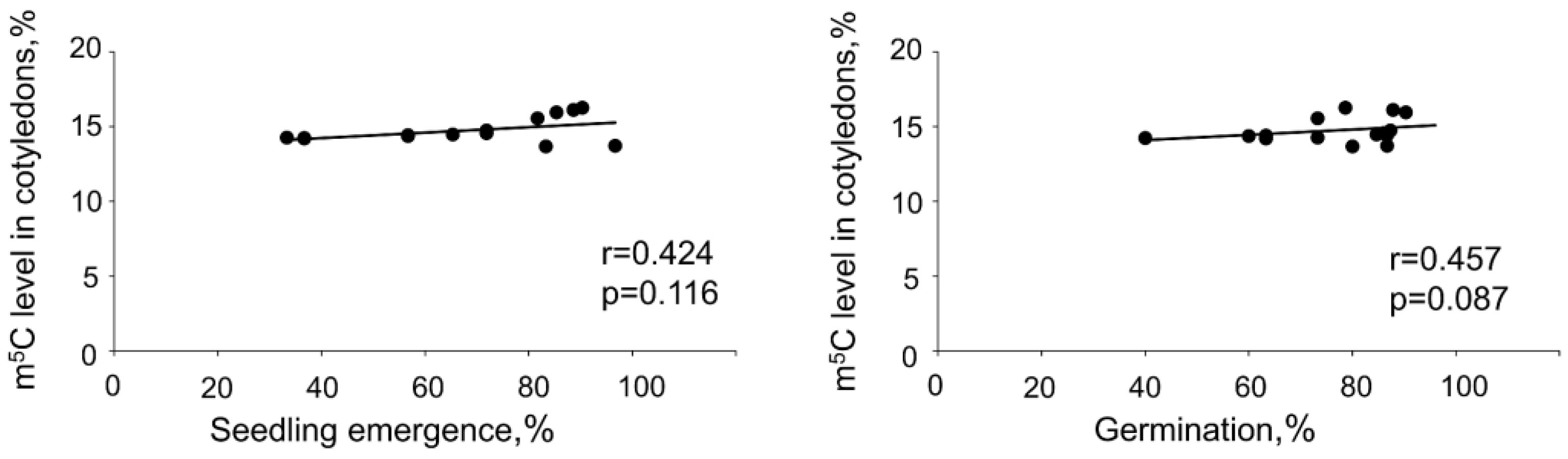
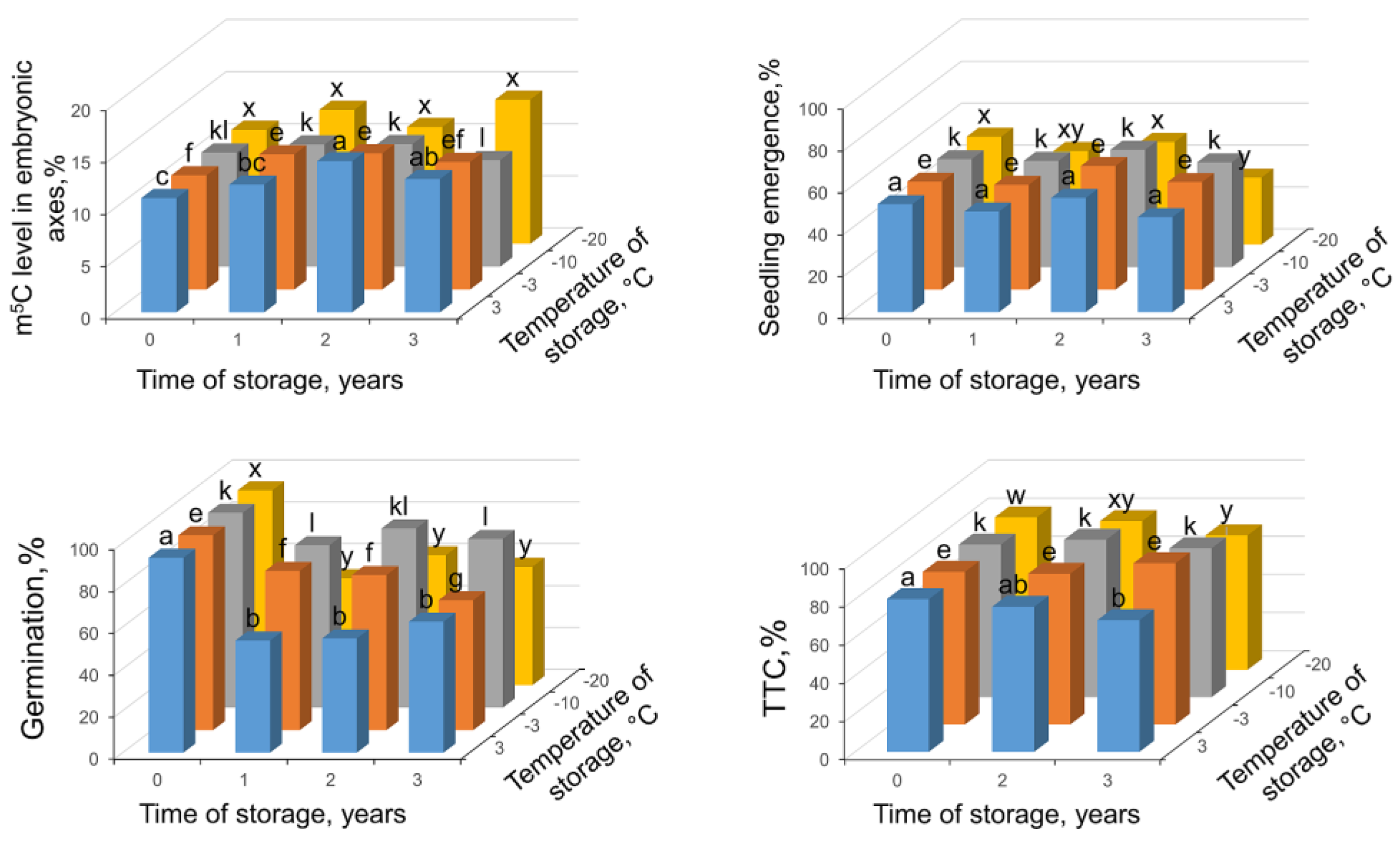
2.1. The Impact of Desiccation on Seed Viability and Global m5C Level
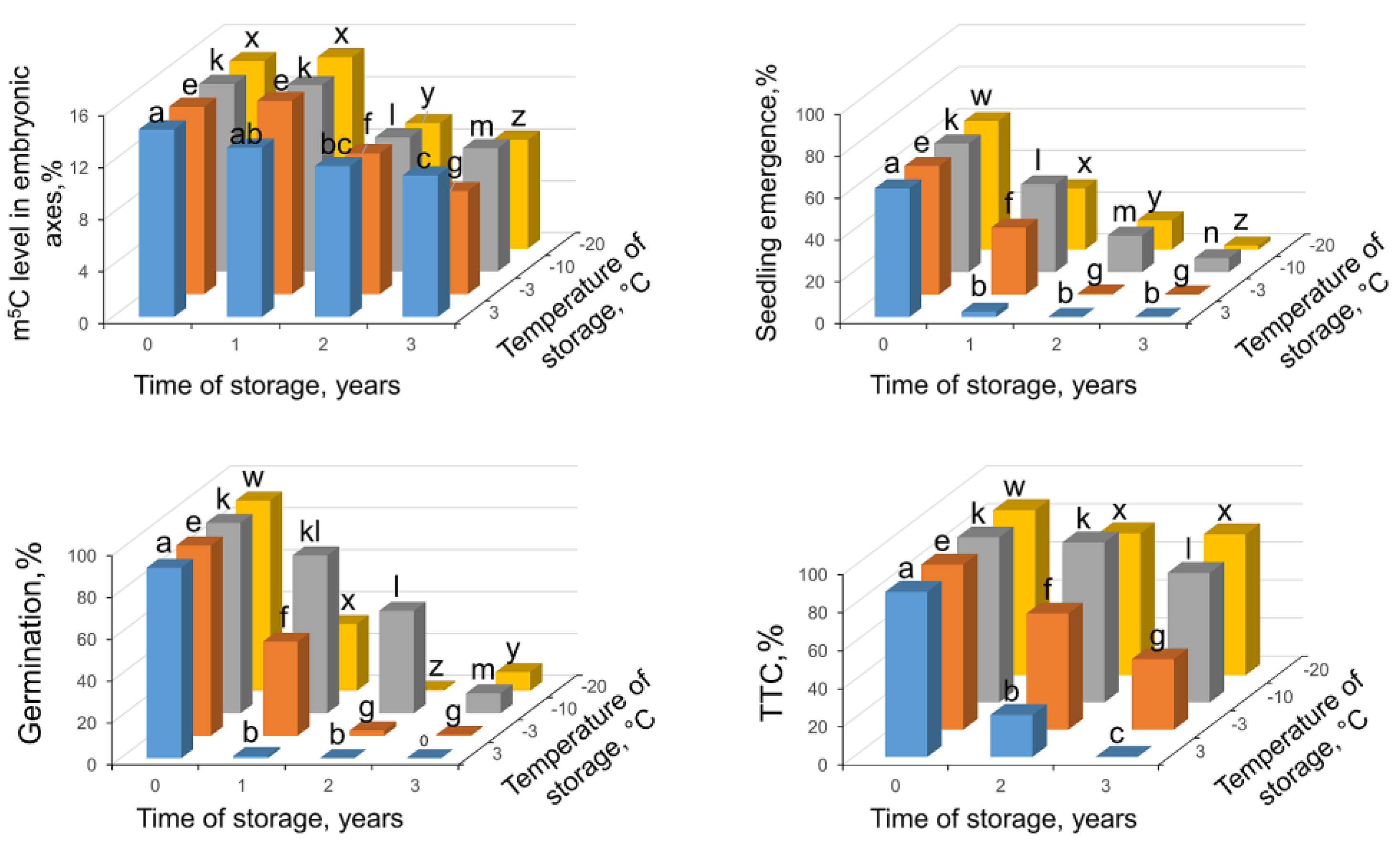
2.2. Seed Viability and Global DNA Methylation Changes in Seeds Stored up to 3 Years at Various Regimes
3. Discussion
4. Materials and Methods
4.1. Plant Material, Assessment of Water Content, and Desiccation of Seeds
4.2. Seed Storage Conditions
4.3. Viability Assessment
4.4. DNA Isolation and Assessment of Global m5C Levels
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bolte, A.; Czajkowski, T.; Kompa, T. The North-Eastern Distribution Range of European Beech—A Review. Forestry 2007, 80, 413–429. [Google Scholar] [CrossRef]
- Del Castillo, E.M.; Zang, C.S.; Buras, A.; Hacket-Pain, A.; Esper, J.; Serrano-Notivoli, R.; Hartl, C.; Weigel, R.; Klesse, S.; Resco de Dios, V.; et al. Climate-Change-Driven Growth Decline of European Beech Forests. Commun. Biol. 2022, 5, 163. [Google Scholar] [CrossRef] [PubMed]
- Dulamsuren, C.; Hauck, M.; Kopp, G.; Ruff, M.; Leuschner, C. European Beech Responds to Climate Change with Growth Decline at Lower, and Growth Increase at Higher Elevations in the Center of Its Distribution Range (SW Germany). Trees-Struct. Funct. 2017, 31, 673–686. [Google Scholar] [CrossRef]
- Roibu, C.-C.; Palaghianu, C.; Nagavciuc, V.; Ionita, M.; Sfecla, V.; Mursa, A.; Crivellaro, A.; Stirbu, M.-I.; Cotos, M.-G.; Popa, A.; et al. The Response of Beech (Fagus sylvatica L.) Populations to Climate in the Easternmost Sites of Its European Distribution. Plants 2022, 11, 3310. [Google Scholar] [CrossRef] [PubMed]
- Bosela, M.; Lukac, M.; Castagneri, D.; Sedmák, R.; Biber, P.; Carrer, M.; Konôpka, B.; Nola, P.; Nagel, T.A.; Popa, I.; et al. Contrasting Effects of Environmental Change on the Radial Growth of Co-Occurring Beech and Fir Trees across Europe. Sci. Total Environ. 2018, 615, 1460–1469. [Google Scholar] [CrossRef]
- Prislan, P.; Čufar, K.; De Luis, M.; Gričar, J. Precipitation Is Not Limiting for Xylem Formation Dynamics and Vessel Development in European Beech from Two Temperate Forest Sites. Tree Physiol. 2018, 38, 186–197. [Google Scholar] [CrossRef]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. ISBN 978-92-9169-143-2. [Google Scholar]
- Alves, S.; Braga, Â.; Parreira, D.; Alhinho, A.T.; Silva, H.; Ramos, M.J.N.; Costa, M.M.R.; Morais-Cecílio, L. Genome-Wide Identification, Phylogeny, and Gene Duplication of the Epigenetic Regulators in Fagaceae. Physiol. Plant. 2022, 174, e13788. [Google Scholar] [CrossRef]
- Tylkowski, T. A Vigour Test for Quality Assessment of Stored Beech (Fagus sylvatica L.) Nuts. Dendrobiology 2002, 47, 43–46. [Google Scholar]
- Vacchiano, G.; Hacket-Pain, A.; Turco, M.; Motta, R.; Maringer, J.; Conedera, M.; Drobyshev, I.; Ascoli, D. Spatial Patterns and Broad-Scale Weather Cues of Beech Mast Seeding in Europe. New Phytol. 2017, 215, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Bogdziewicz, M.; Szymkowiak, J.; Kasprzyk, I.; Grewling, L.; Borowski, Z.; Borycka, K.; Kantorowicz, W.; Myszkowska, D.; Piotrowicz, K.; Ziemianin, M.; et al. Masting in Wind-Pollinated Trees: System-Specific Roles of Weather and Pollination Dynamics in Driving Seed Production. Ecology 2017, 98, 2615–2625. [Google Scholar] [CrossRef]
- Bogdziewicz, M.; Hacket-Pain, A.; Ascoli, D.; Szymkowiak, J. Environmental Variation Drives Continental-Scale Synchrony of European Beech Reproduction. Ecology 2021, 102, e03384. [Google Scholar] [CrossRef]
- Prochazkova, Z.; Bezdeckova, L. Effect of Accelerated Ageing on the Viability and Germination of European Beech (Fagus sylvatica L.) Seeds. Seed Sci. Technol. 2009, 37, 699–712. [Google Scholar] [CrossRef]
- Pukacka, S.; Ratajczak, E. Age-Related Biochemical Changes during Storage of Beech (Fagus sylvatica L.) Seeds. Seed Sci. Res. 2007, 17, 45–53. [Google Scholar] [CrossRef]
- Gosling, P. Raising Trees and Shrubs from Seed: Practice Guide; Forest Research: Bristol, UK, 2007. [Google Scholar]
- Pukacka, S.; Hoffmann, S.K.; Goslar, J.; Pukacki, P.M.; Wójkiewicz, E. Water and Lipid Relations in Beech (Fagus sylvatica L.) Seeds and Its Effect on Storage Behaviour. Biochim. Biophys. Acta 2003, 1621, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Siger, A.; Dwiecki, K.; Borzyszkowski, W.; Turski, M.; Rudzińska, M.; Nogala-Kałucka, M. Physicochemical Characteristics of the Cold-Pressed Oil Obtained from Seeds of Fagus sylvatica L. Food Chem. 2017, 225, 239–245. [Google Scholar] [CrossRef]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. An Intermediate Category of Seed Storage Behaviour? I. COFFEE. J. Exp. Bot. 1990, 41, 1167–1174. [Google Scholar] [CrossRef]
- Volk, G.M.; Crane, J.; Caspersen, A.M.; Hill, L.M.; Gardner, C.; Walters, C. Massive Cellular Disruption Occurs during Early Imbibition of Cuphea Seeds Containing Crystallized Triacylglycerols. Planta 2006, 224, 1415–1426. [Google Scholar] [CrossRef]
- Trusiak, M.; Plitta-Michalak, B.P.; Michalak, M. Choosing the Right Path for the Successful Storage of Seeds. Plants 2023, 12, 72. [Google Scholar] [CrossRef]
- Poulsen, K. Predicting the Storage Life of Beech Nuts. Seed Sci. Technol. 1993, 21, 327–337. [Google Scholar]
- Poulsen, K.M.; Knudsen, H. Viability Constants Based on Eight Years Storage of Beech Nuts (Fagus sylvatica L.). Seed Sci. Technol. 1999, 27, 1037–1039. [Google Scholar]
- Leon-Lobos, P.; Ellis, R.H. Seed Storage Behaviour of Fagus sylvatica and Fagus crenata. Seed Sci. Res. 2002, 12, 31–37. [Google Scholar] [CrossRef]
- Fortes, A.M.; Gallusci, P. Plant Stress Responses and Phenotypic Plasticity in the Epigenomics Era: Perspectives on the Grapevine Scenario, a Model for Perennial Crop Plants. Front. Plant Sci. 2017, 8, 82. [Google Scholar] [CrossRef] [PubMed]
- Gallusci, P.; Agius, D.R.; Moschou, P.N.; Dobránszki, J.; Kaiserli, E.; Martinelli, F. Deep inside the Epigenetic Memories of Stressed Plants. Trends Plant Sci. 2022, 28, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Barciszewska, M.Z.; Barciszewski, J.; Plitta, B.P.; Chmielarz, P. Global Changes in DNA Methylation in Seeds and Seedlings of Pyrus Communis after Seed Desiccation and Storage. PLoS ONE 2013, 8, e70693. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Plitta-Michalak, B.; Naskręt-Barciszewska, M.Z.; Barciszewski, J.; Bujarska-Borkowska, B.; Chmielarz, P. Global 5-Methylcytosine Alterations in DNA during Ageing of Quercus robur Seeds. Ann. Bot. 2015, 116, 369–376. [Google Scholar] [CrossRef]
- Plitta, B.P.; Michalak, M.; Bujarska-Borkowska, B.; Barciszewska, M.Z.; Barciszewski, J.; Chmielarz, P. Effect of Desiccation on the Dynamics of Genome-Wide DNA Methylation in Orthodox Seeds of Acer platanoides L. Plant Physiol. Biochem. 2014, 85, 71–77. [Google Scholar] [CrossRef]
- Plitta-Michalak, B.P.; Naskręt-Barciszewska, M.Z.; Kotlarski, S.; Tomaszewski, D.; Tylkowski, T.; Barciszewski, J.; Chmielarz, P.; Michalak, M. Changes in Genomic 5-Methylcytosine Level Mirror the Response of Orthodox (Acer platanoides L.) and Recalcitrant (Acer pseudoplatanus L.) Seeds to Severe Desiccation. Tree Physiol. 2018, 38, 617–629. [Google Scholar] [CrossRef]
- Michalak, M.; Plitta-Michalak, B.P.; Naskręt-Barciszewska, M.Z.; Barciszewski, J.; Chmielarz, P. DNA Methylation as an Early Indicator of Aging in Stored Seeds of “Exceptional” Species Populus nigra L. Cells 2022, 11, 2080. [Google Scholar] [CrossRef]
- Ballesteros, D.; Pritchard, H.W.; Walters, C. Dry Architecture: Towards the Understanding of the Variation of Longevity in Desiccation-Tolerant Germplasm. Seed Sci. Res. 2020, 30, 142–155. [Google Scholar] [CrossRef]
- Roqueiro, G.; Facorro, G.B.; Huarte, M.G.; Rubín de Celis, E.; García, F.; Maldonado, S.; Maroder, H. Effects of Photooxidation on Membrane Integrity in Salix nigra Seeds. Ann. Bot. 2010, 105, 1027–1034. [Google Scholar] [CrossRef]
- Suszka, J.; Plitta, B.P.; Michalak, M.; Bujarska-Borkowska, B.; Tylkowski, T.; Chmielarz, P. Optimal Seed Water Content and Storage Temperature for Preservation of Populus nigra L. Germplasm. Ann. For. Sci. 2014, 71, 543–549. [Google Scholar] [CrossRef]
- Pathirana, R.; Carimi, F. Management and Utilization of Plant Genetic Resources for a Sustainable Agriculture. Plants 2022, 11, 2038. [Google Scholar] [CrossRef] [PubMed]
- Engels, J.M.M.; Thormann, I. Main Challenges and Actions Needed to Improve Conservation and Sustainable Use of Our Crop Wild Relatives. Plants 2020, 9, 968. [Google Scholar] [CrossRef] [PubMed]
- Trail, P.; Motis, T.; Swartz, S.; Bicksler, A. Low-Cost Seed Storage Technologies for Development Impact of Small-Scale Seed Saving Entities in Tropical Climates. Exp. Agric. 2021, 57, 324–337. [Google Scholar] [CrossRef]
- Niedzielski, M.; Walters, C.; Luzcak, W.; Hill, L.; Wheeler, L.; Puchalski, J. Assessment of Variation in Seed Longevity within Rye, Wheat and the Intergeneric Hybrid Triticale. Seed Sci. Res. 2009, 19, 213–224. [Google Scholar] [CrossRef]
- Walters, C. Understanding the Mechanisms and Kinetics of Seed Aging. Seed Sci. Res. 1998, 8, 223–244. [Google Scholar] [CrossRef]
- Ballesteros, D.; Walters, C. Detailed Characterization of Mechanical Properties and Molecular Mobility within Dry Seed Glasses: Relevance to the Physiology of Dry Biological Systems. Plant J. 2011, 68, 607–619. [Google Scholar] [CrossRef]
- Michalak, M.; Plitta-Michalak, B.P.; Nadarajan, J.; Colville, L. Volatile Signature Indicates Viability of Dormant Orthodox Seeds. Physiol. Plant. 2021, 173, 788–804. [Google Scholar] [CrossRef]
- Ratajczak, E.; Pukacka, S. Decrease in Beech (Fagus sylvatica) Seed Viability Caused by Temperature and Humidity Conditions as Related to Membrane Damage and Lipid Composition. Acta Physiol. Plant. 2005, 27, 3–12. [Google Scholar] [CrossRef]
- Ratajczak, E.; Malecka, A.; Bagniewska-Zadworna, A.; Kalemba, E.M. The Production, Localization and Spreading of Reactive Oxygen Species Contributes to the Low Vitality of Long-Term Stored Common Beech (Fagus sylvatica L.) Seeds. J. Plant Physiol. 2015, 174, 147–156. [Google Scholar] [CrossRef]
- Szczotka, Z.; Pawlowski, T.; Krawiarz, K. Proteins and Polyamines during Dormancy Breaking of European Beech (Fagus sylvatica L.) Seeds. Acta Physiol. Plant. 2003, 25, 423–435. [Google Scholar] [CrossRef]
- Pawlowski, T.A. Proteomics of European Beech (Fagus sylvatica L.) Seed Dormancy Breaking: Influence of Abscisic and Gibberellic Acids. Proteomics 2007, 7, 2246–2257. [Google Scholar] [CrossRef] [PubMed]
- van Zanten, M.; Tessadori, F.; Peeters, A.J.M.; Fransz, P. Environment-Induced Chromatin Reorganisation and Plant Acclimation. In Epigenetic Memory and Control in Plants; Grafi, G., Ohad, N., Eds.; Signaling and Communication in Plants; Springer: Berlin/Heidelberg, Germany, 2013; pp. 21–40. ISBN 978-3-642-35227-0. [Google Scholar]
- van Zanten, M.; Liu, Y.; Soppe, W.J.J. Epigenetic Signalling During the Life of Seeds. In Epigenetic Memory and Control in Plants; Grafi, G., Ohad, N., Eds.; Signaling and Communication in Plants; Springer: Berlin/Heidelberg, Germany, 2013; pp. 127–153. ISBN 978-3-642-35227-0. [Google Scholar]
- Quadrana, L.; Colot, V. Plant Transgenerational Epigenetics. In Annual Review of Genetics; Bonini, N.M., Ed.; Annual Reviews: Palo Alto, CA, USA, 2016; Volume 50, pp. 467–491. ISBN 978-0-8243-1250-3. [Google Scholar]
- Xiao, W.; Custard, K.D.; Brown, R.C.; Lemmon, B.E.; Harada, J.J.; Goldberg, R.B.; Fischer, R.L. DNA Methylation Is Critical for Arabidopsis Embryogenesis and Seed Viability. Plant Cell 2006, 18, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Mira, S.; Pirredda, M.; Martín-Sánchez, M.; Marchessi, J.E.; Martín, C. DNA Methylation and Integrity in Aged Seeds and Regenerated Plants. Seed Sci. Res. 2020, 30, 92–100. [Google Scholar] [CrossRef]
- Ogneva, Z.V.; Dubrovina, A.S.; Kiselev, K.V. Age-Associated Alterations in DNA Methylation and Expression of Methyltransferase and Demethylase Genes in Arabidopsis thaliana. Biol. Plant. 2016, 60, 628–634. [Google Scholar] [CrossRef]
- Pirredda, M.; González-Benito, M.E.; Martín, C.; Mira, S. Genetic and Epigenetic Stability in Rye Seeds under Different Storage Conditions: Ageing and Oxygen Effect. Plants 2020, 9, 393. [Google Scholar] [CrossRef]
- Pukacka, S.; Ratajczak, E. Factors Influencing the Storability of Fagus sylvatica L. Seeds after Release from Dormancy. Plant Growth Regul. 2014, 72, 17–27. [Google Scholar] [CrossRef][Green Version]
- Michalak, M.; Plitta-Michalak, B.P.; Chmielarz, P. A New Insight in Desiccation Tolerance and Cryopreservation of Mazzard Cherry (Prunus avium L.) Seeds. Open Life Sci. 2015, 10, 354–364. [Google Scholar] [CrossRef]
- Sershen; Varghese, B.; Naidoo, C.; Pammenter, N.W. The Use of Plant Stress Biomarkers in Assessing the Effects of Desiccation in Zygotic Embryos from Recalcitrant Seeds: Challenges and Considerations. Plant Biol. 2016, 18, 433–444. [Google Scholar] [CrossRef]
- Plitta-Michalak, B.P.; Litkowiec, M.; Michalak, M. Epigenetic Marks, DNA Damage Markers, or Both? The Impact of Desiccation and Accelerated Aging on Nucleobase Modifications in Plant Genomic DNA. Cells 2022, 11, 1748. [Google Scholar] [CrossRef]
- Plitta-Michalak, B.P.; Naskręt-Barciszewska, M.Z.; Barciszewski, J.; Chmielarz, P.; Michalak, M. Epigenetic Integrity of Orthodox Seeds Stored under Conventional and Cryogenic Conditions. Forests 2021, 12, 288. [Google Scholar] [CrossRef]
- Varga, S.; Soulsbury, C.D. Paternal Arbuscular Mycorrhizal Fungal Status Affects DNA Methylation in Seeds. Biol. Lett. 2017, 13, 20170407. [Google Scholar] [CrossRef]
- Kurek, K.; Plitta-Michalak, B.; Ratajczak, E. Reactive Oxygen Species as Potential Drivers of the Seed Aging Process. Plants 2019, 8, 174. [Google Scholar] [CrossRef]
- Chandra, J.; Parkhey, S.; Keshavkant, S. Ageing-Regulated Changes in Genetic Integrity of Two Recalcitrant Seeded Species Having Contrasting Longevity. Trees-Struct. Funct. 2018, 32, 109–123. [Google Scholar] [CrossRef]
- Suszka, B.; Muller, C.; Bonnet-Masimbert, M. Seeds of Forest Broadleaves: From Harvest to Sowing; Institut National de la Recherche Agronomique: Paris, France, 1996; ISBN 978-2-7380-0659-2. [Google Scholar]
- Wang, W.-Q.; Cheng, H.-Y.; Song, S.-Q. Development of a Threshold Model to Predict Germination of Populus tomentosa Seeds after Harvest and Storage under Ambient Condition. PLoS ONE 2013, 8, e62868. [Google Scholar] [CrossRef] [PubMed]
- ISTA. International Rules for Seed Testing. Available online: https://www.seedtest.org/en/publications/international-rules-seed-testing-1168.html (accessed on 29 April 2022).
- Plitta, B.P.; Michalak, M.; Naskręt-Barciszewska, M.Z.; Barciszewski, J.; Chmielarz, P. DNA Methylation of Quercus robur L. Plumules Following Cryo-Pretreatment and Cryopreservation. Plant Cell Tiss. Organ Cult. 2014, 117, 31–37. [Google Scholar] [CrossRef]
- Bliss, C.I. The Transformation of Percentages for Use in the Analysis of Variance. Ohio J. Sci. 1938, 38, 9–12. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source Variance | DF | F | p |
|---|---|---|---|
| ST | 3 | 119.5 | <0.0001 |
| MC | 1 | 7.3 | 0.0087 |
| STE | 3 | 16.2 | <0.0001 |
| MC * STE | 3 | 5.9 | 0.0012 |
| MC * STE | 3 | 90.6 | <0.0001 |
| ST * STE | 9 | 12.7 | <0.0001 |
| ST * MC * STE | 9 | 3.3 | 0.0024 |
| Source Variance | DF | F | p |
|---|---|---|---|
| ST | 3 | 29.05 | <0.0001 |
| MC | 1 | 671.52 | <0.0001 |
| STE | 3 | 598.91 | <0.0001 |
| MC * STE | 3 | 34.52 | <0.0001 |
| MC *STE | 3 | 296.25 | <0.0001 |
| ST * STE | 9 | 7.33 | <0.0001 |
| ST * MC * STE | 9 | 8.14 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michalak, M.; Plitta-Michalak, B.P.; Suszka, J.; Naskręt-Barciszewska, M.Z.; Kotlarski, S.; Barciszewski, J.; Chmielarz, P. Identification of DNA Methylation Changes in European Beech Seeds during Desiccation and Storage. Int. J. Mol. Sci. 2023, 24, 3557. https://doi.org/10.3390/ijms24043557
Michalak M, Plitta-Michalak BP, Suszka J, Naskręt-Barciszewska MZ, Kotlarski S, Barciszewski J, Chmielarz P. Identification of DNA Methylation Changes in European Beech Seeds during Desiccation and Storage. International Journal of Molecular Sciences. 2023; 24(4):3557. https://doi.org/10.3390/ijms24043557
Chicago/Turabian StyleMichalak, Marcin, Beata Patrycja Plitta-Michalak, Jan Suszka, Mirosława Zofia Naskręt-Barciszewska, Szymon Kotlarski, Jan Barciszewski, and Paweł Chmielarz. 2023. "Identification of DNA Methylation Changes in European Beech Seeds during Desiccation and Storage" International Journal of Molecular Sciences 24, no. 4: 3557. https://doi.org/10.3390/ijms24043557
APA StyleMichalak, M., Plitta-Michalak, B. P., Suszka, J., Naskręt-Barciszewska, M. Z., Kotlarski, S., Barciszewski, J., & Chmielarz, P. (2023). Identification of DNA Methylation Changes in European Beech Seeds during Desiccation and Storage. International Journal of Molecular Sciences, 24(4), 3557. https://doi.org/10.3390/ijms24043557