
The Angiogenesis Inhibitor Isthmin-1 (ISM1) Is Overexpressed in Experimental Models of Glomerulopathy and Impairs the Viability of Podocytes

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
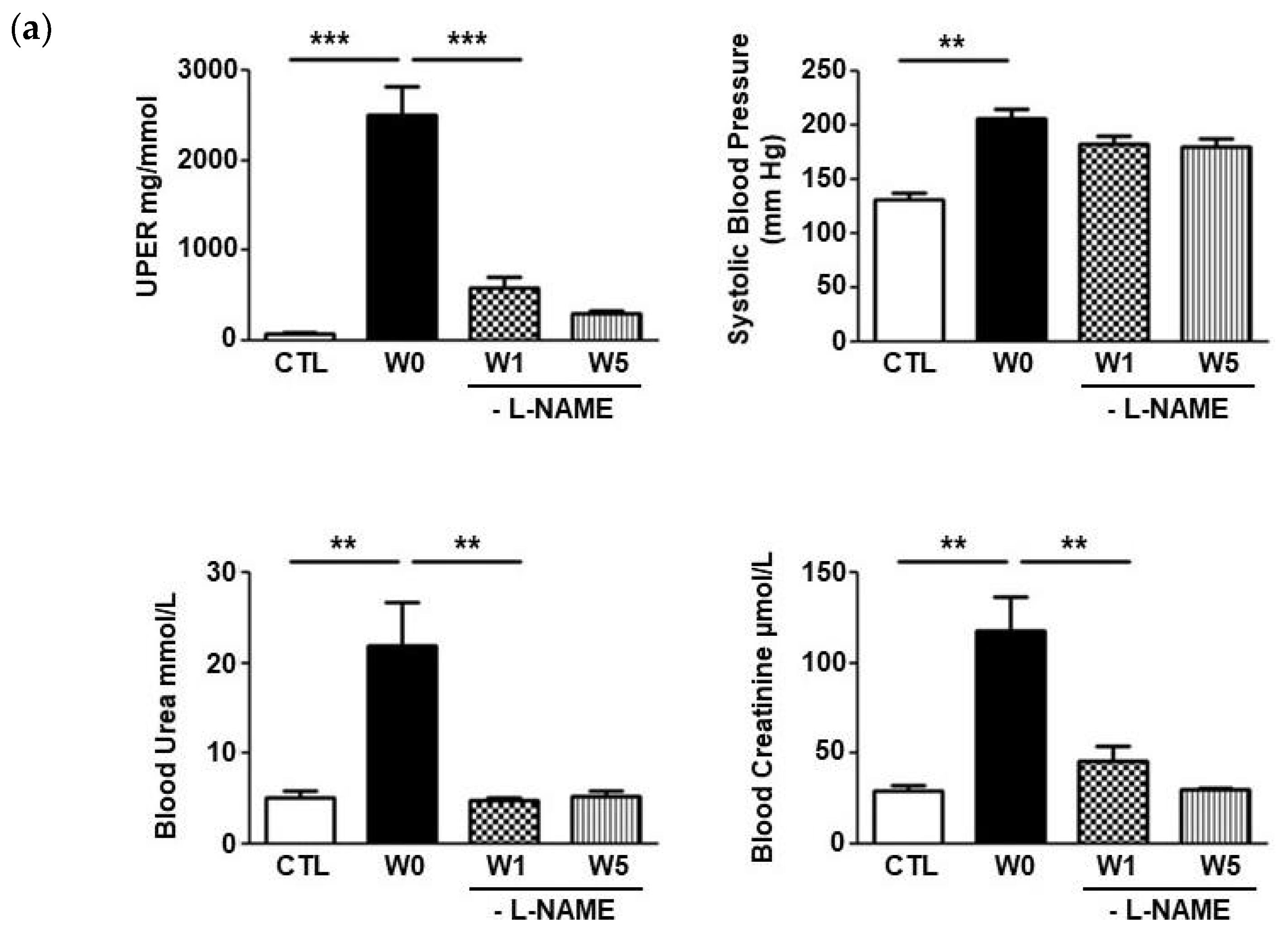
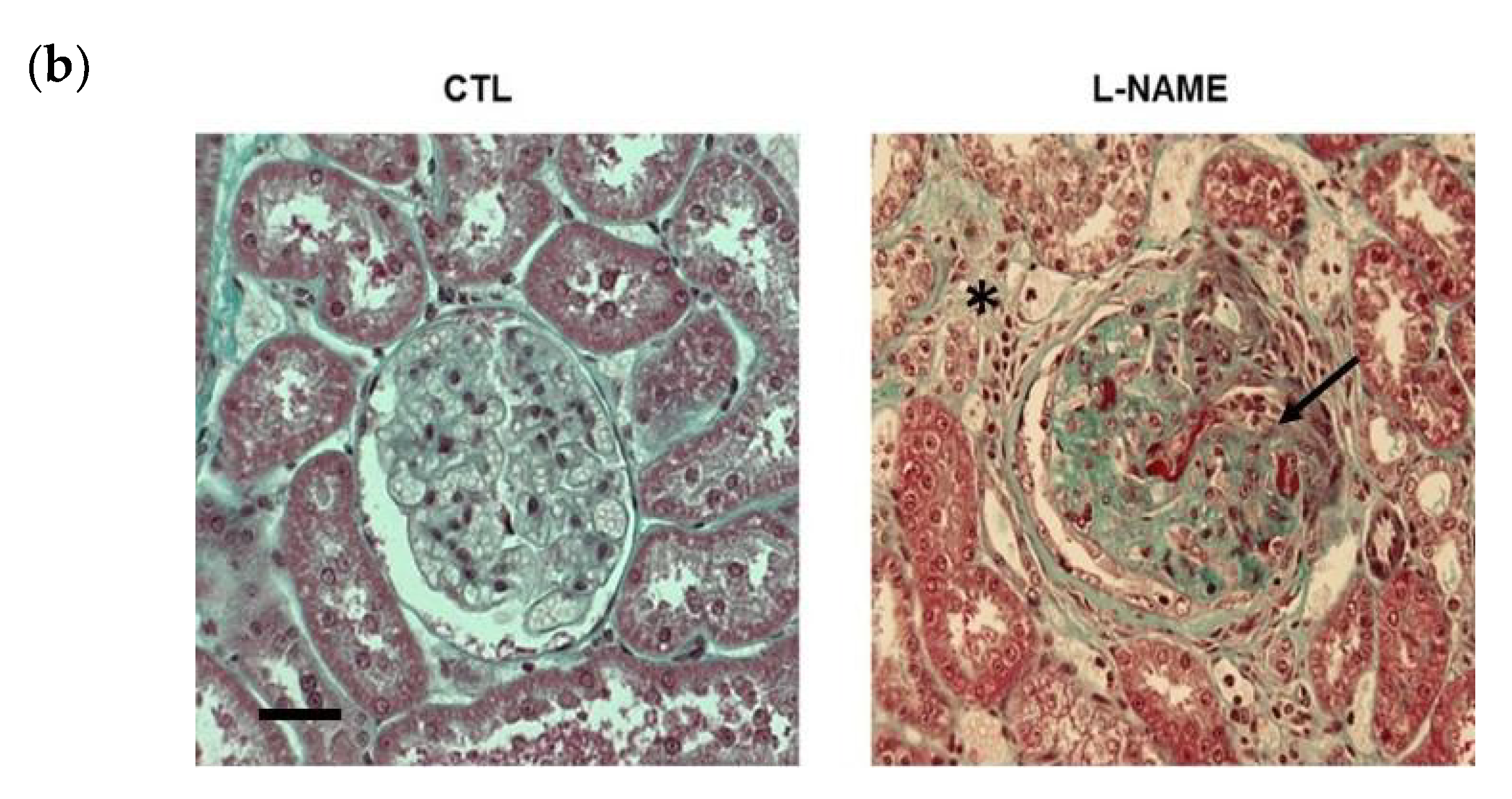
2.1. L-NAME Induced Nephroangiosclerosis with Focal Segmental Glomerulosclerosis
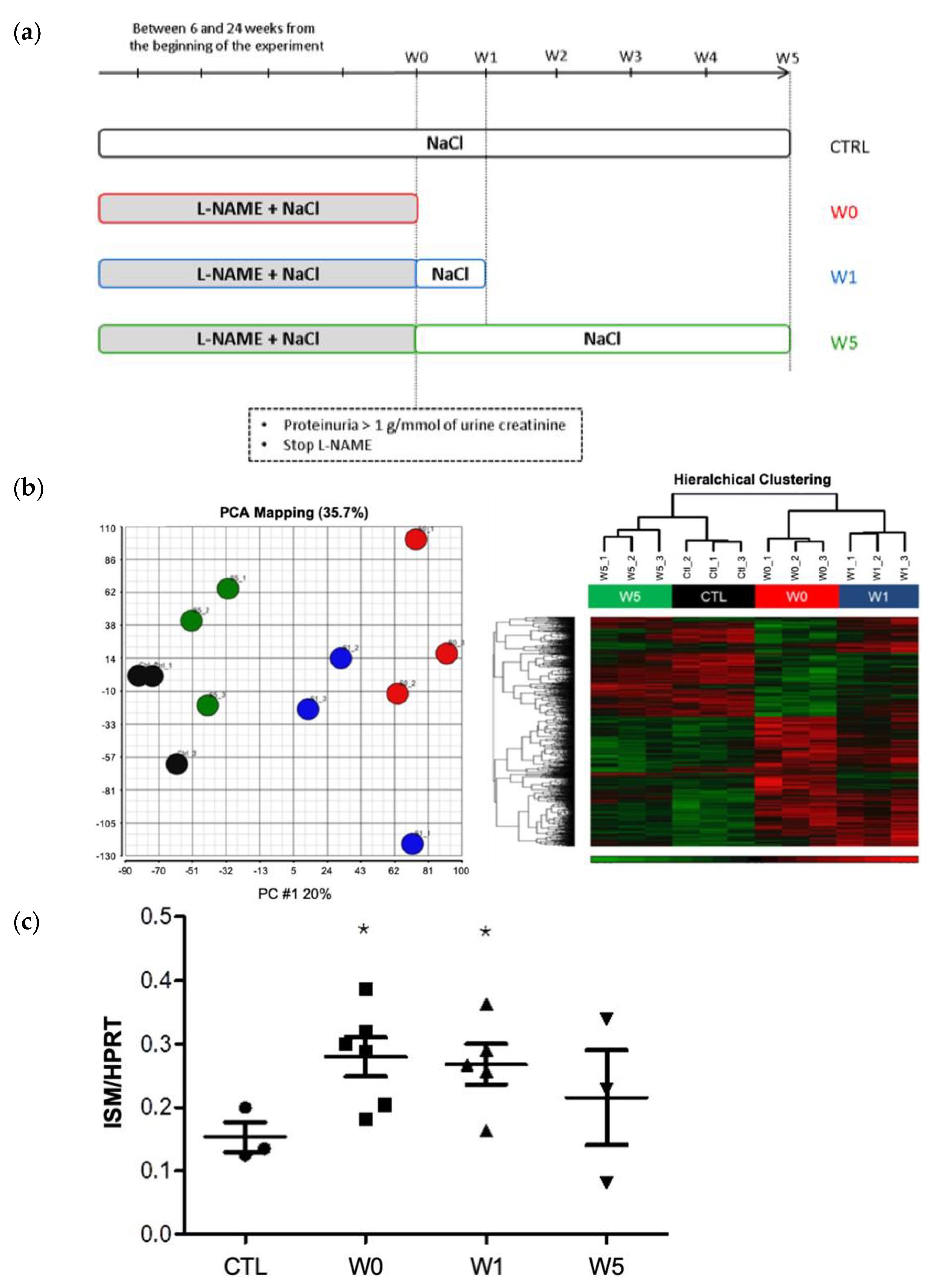
2.2. ISM1 Gene Expression Is Associated to the Intensity of Renal Injury in L-NAME-Induced Nephropathy and Its Repair
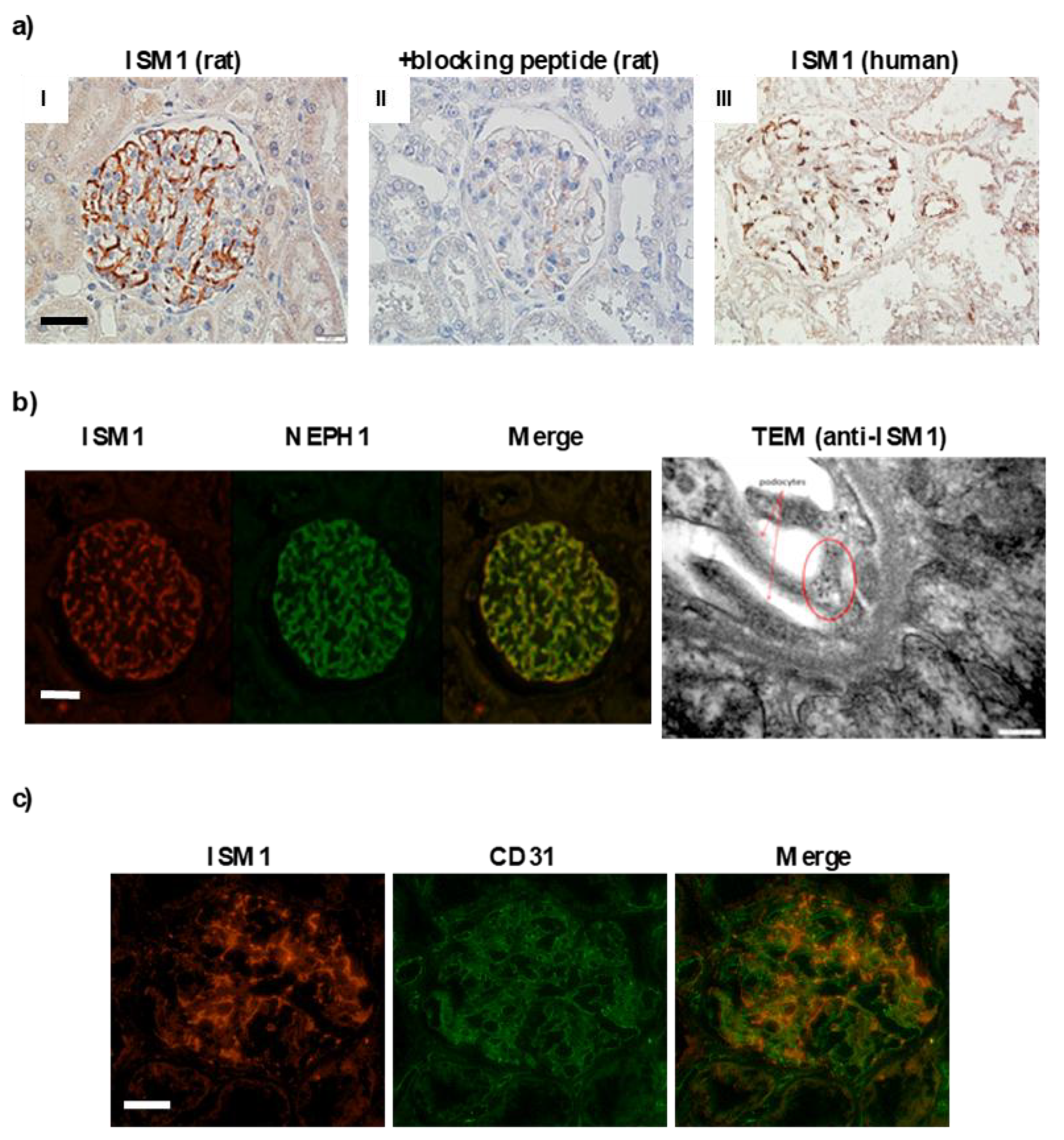
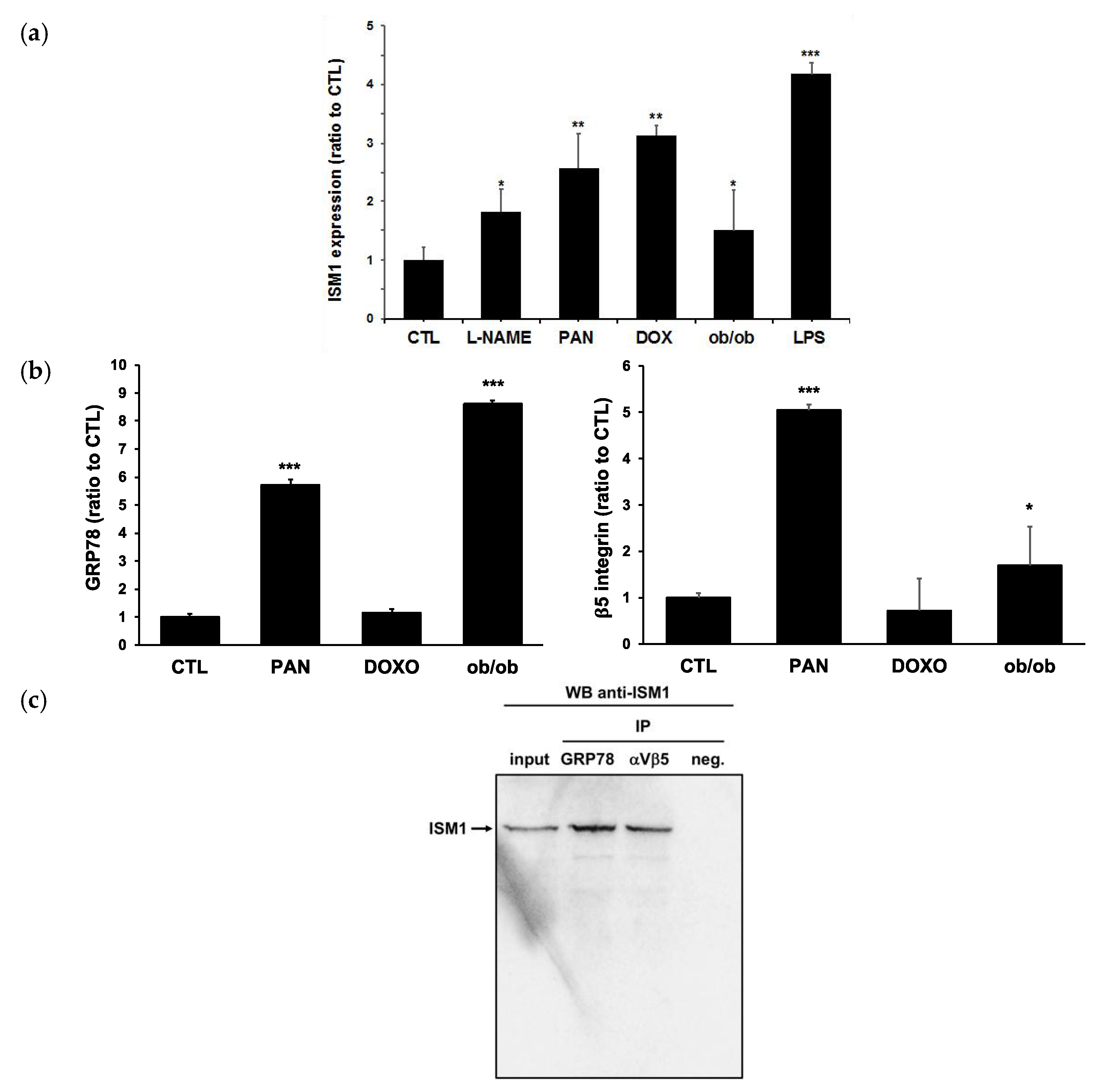
2.3. ISM1 and Receptors Are Expressed in Podocytes of Different Models of Glomerulopathy
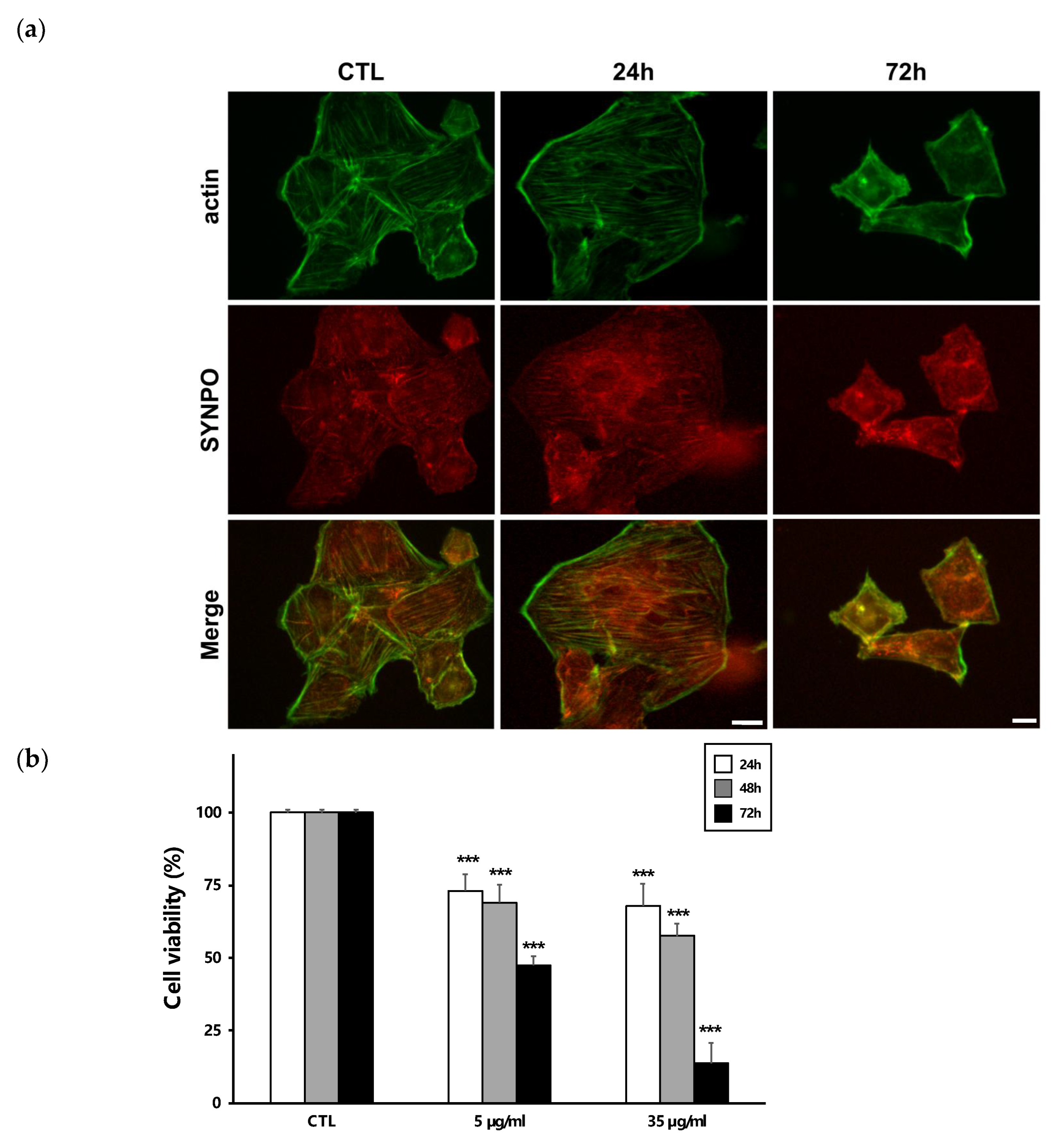
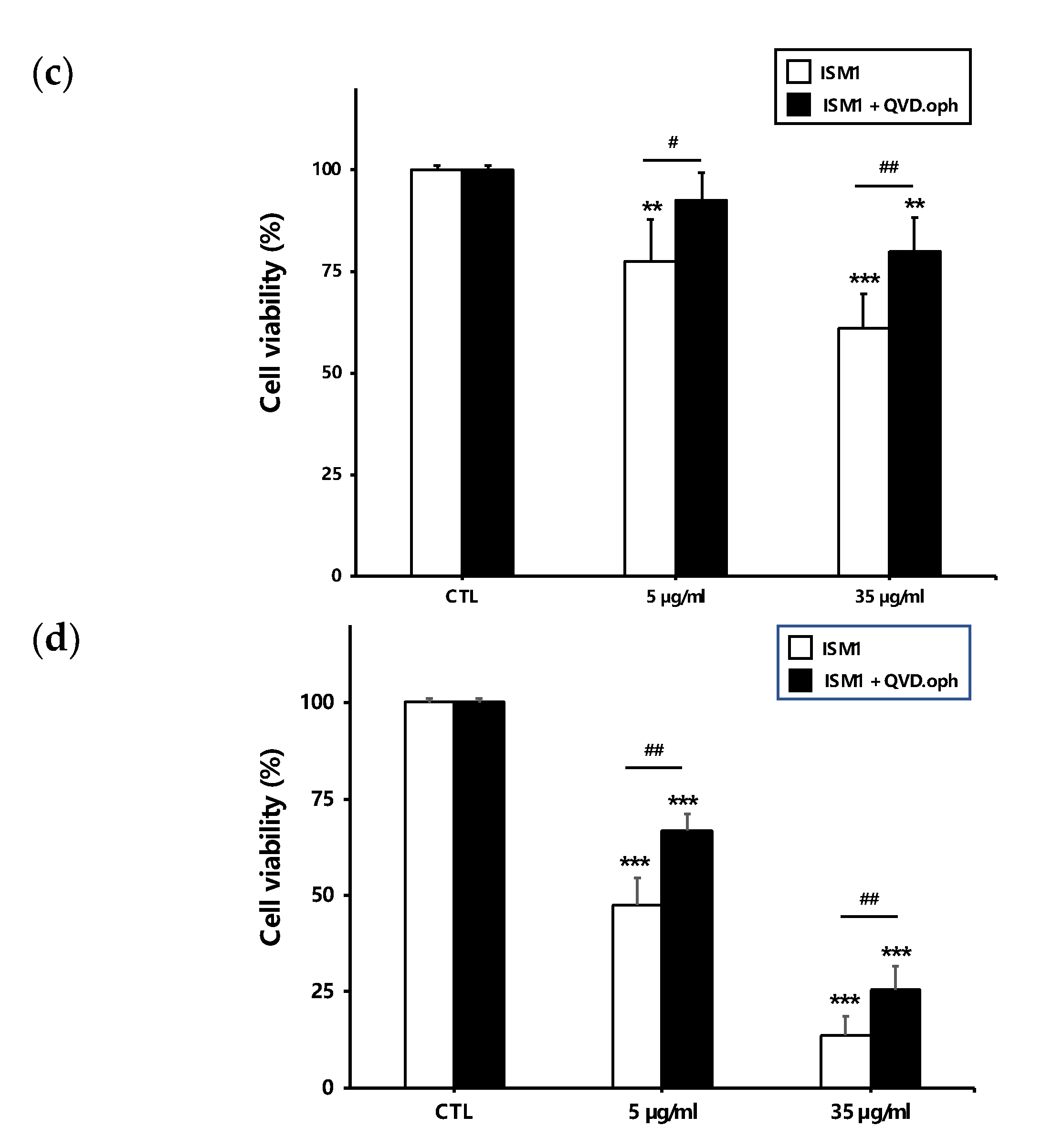
2.4. Recombinant ISM1 Induced Podocyte Injury In Vitro
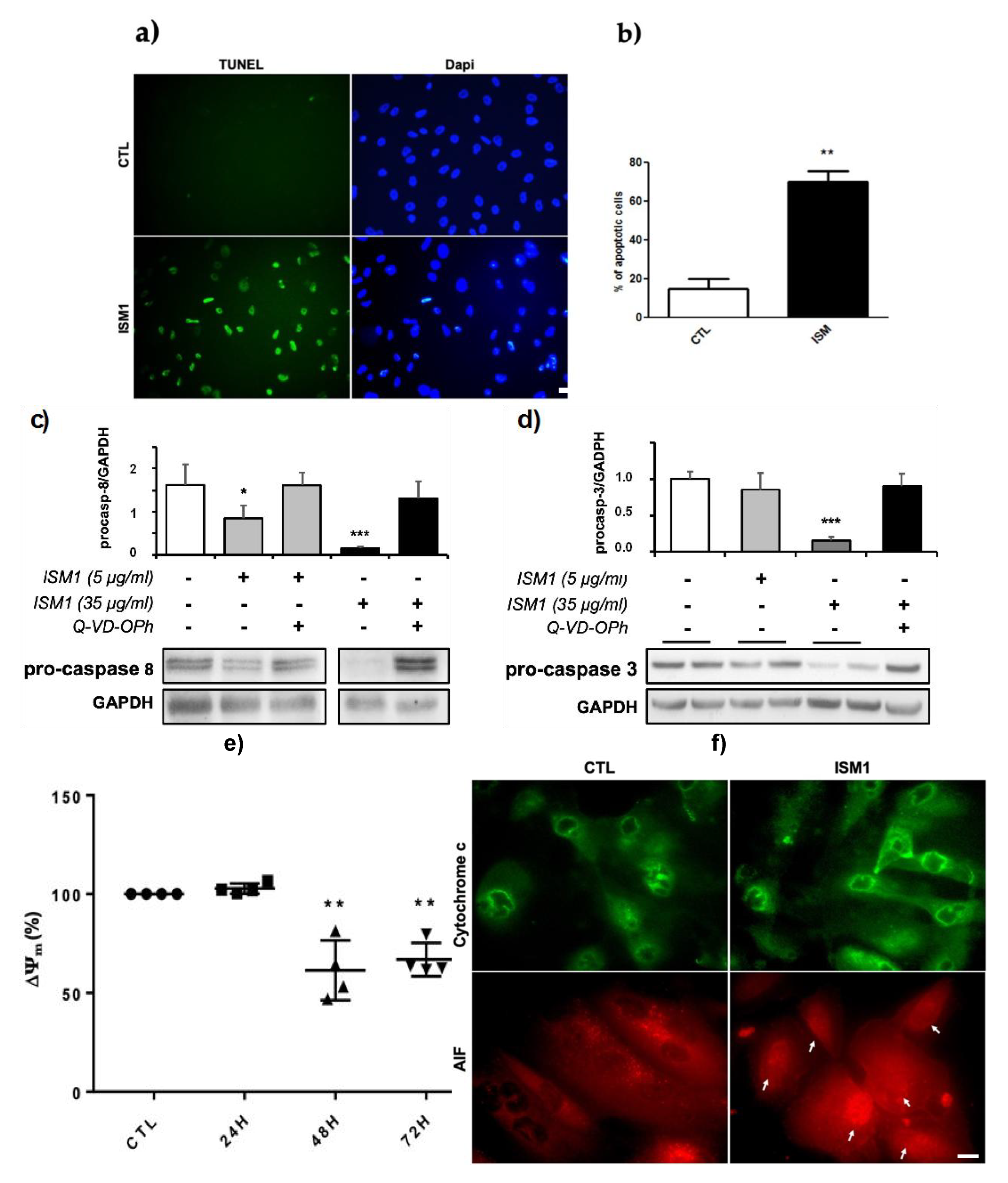
2.5. Recombinant ISM1 Induced Podocyte Apoptosis, Mitochondrial Membrane Depolarization and Release of Pro-Apoptotic Proteins
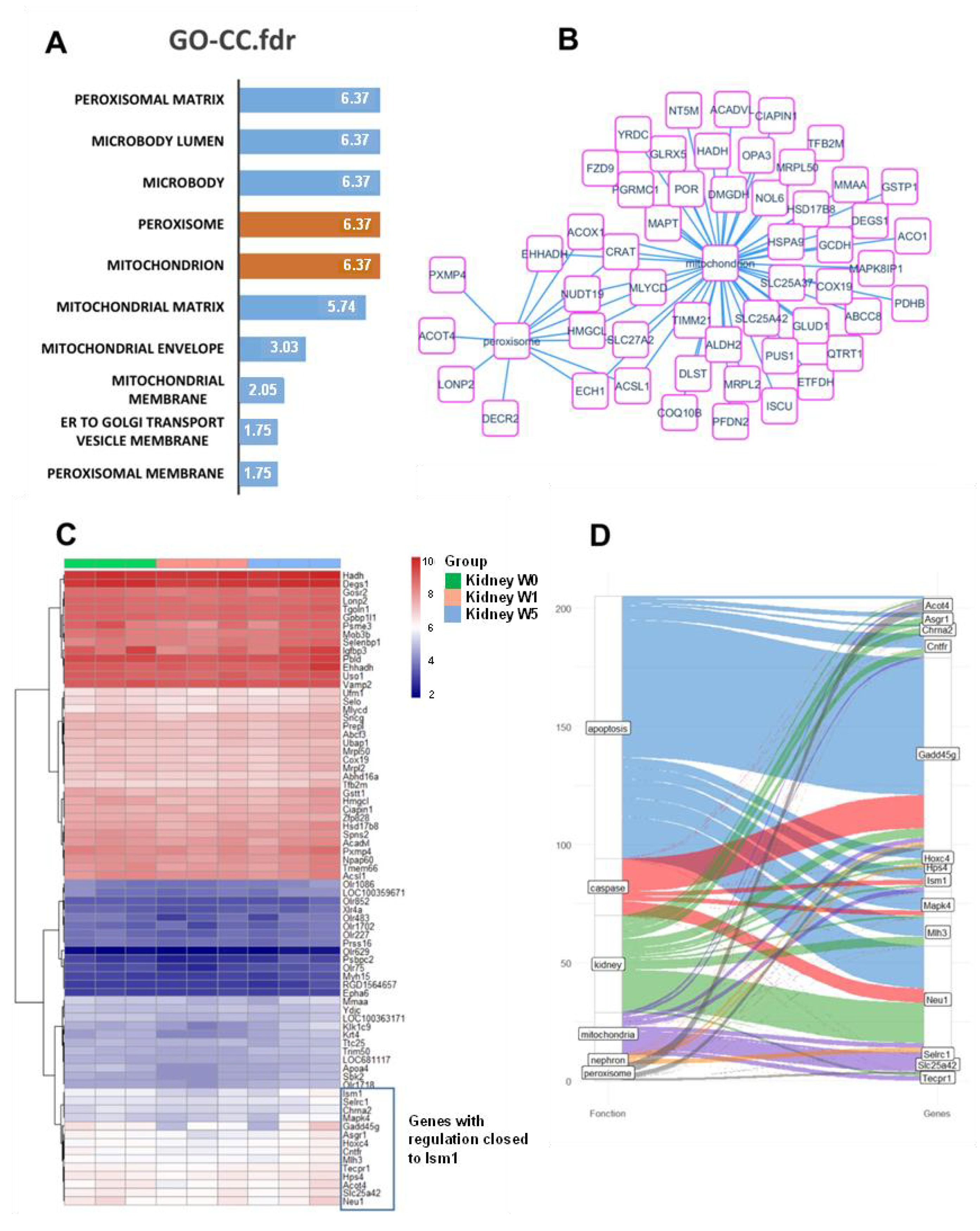
2.6. Proposal Roles of Mitochondria: In Silico Analysis of ISM1 Regulation in the L-NAME Model
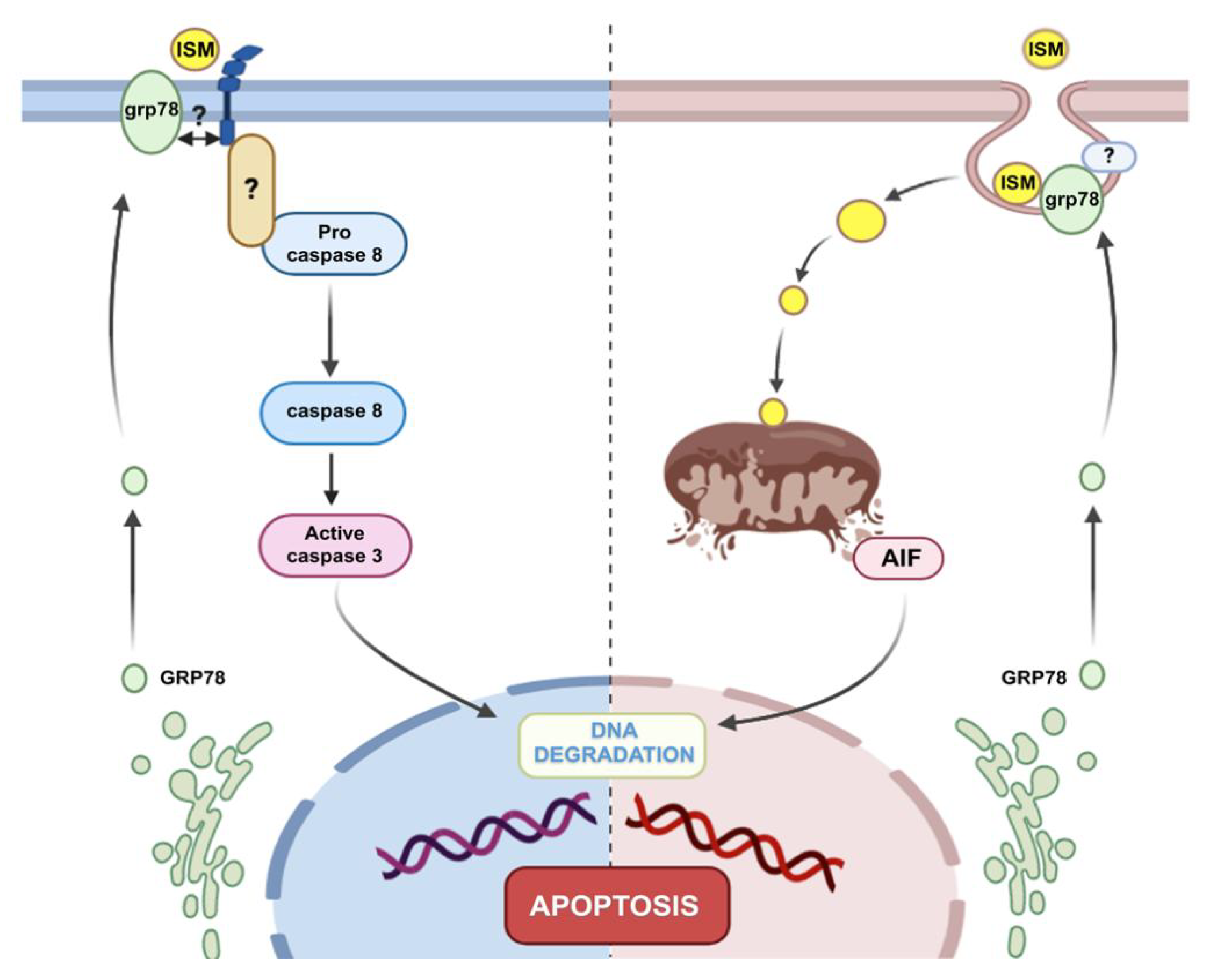
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Animal Models of Proteinuria
4.2.1. Hypertensive Nephropathy with FSGS Lesions (L-NAME Model)
- -
- Group W0, sacrificed at the peak of renal damage (UPER > 1 g/mmol) (n = 6).
- -
- Group W1, animals with UPER >1 g/mmol were sacrificed one week after the interruption of L-NAME (n = 5).
- -
- Group W5, animals with UPER >1 g/mmol were sacrificed 5 weeks after the interruption of L-NAME (n = 4).
- -
- -CTL group, which did not receive L-NAME and was sacrificed alongside other groups to obtain animals of the same age (n = 4).
4.2.2. Puromycin Model
4.2.3. Doxorubicin Model
4.2.4. Diabetic Nephropathy Model
4.2.5. Lipopolysaccharide (LPS) Model
4.3. DNA Microarrays Analysis
4.4. Immunohistochemistry Analysis
4.5. Isolation of Glomeruli and qPCR Analysis
4.6. Podocytes Cell Culture
4.7. Cell Viability and TUNEL Assay
4.8. Immunofluorescence
4.9. Fluorimetric Determination of Mitochondrial Membrane Potential (ΔΨm)
4.10. Western Blotting and Immunoprecipitation Assays
4.11. Statistical Analysis
4.12. In Silico Analysis of ISM1-Related Pathways of the L-NAME Model
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosenberg, A.Z.; Kopp, J.B. Focal Segmental Glomerulosclerosis. Clin. J. Am. Soc. Nephrol. CJASN 2017, 12, 502–517. [Google Scholar] [CrossRef] [PubMed]
- Costantino, V.V.; Gil Lorenzo, A.F.; Bocanegra, V.; Vallés, P.G. Molecular Mechanisms of Hypertensive Nephropathy: Renoprotective Effect of Losartan through Hsp70. Cells 2021, 10, 3146. [Google Scholar] [CrossRef]
- Cai, A.; Chatziantoniou, C.; Calmont, A. Vascular Permeability: Regulation Pathways and Role in Kidney Diseases. Nephron 2021, 145, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Pera, E.M.; Kim, J.I.; Martinez, S.L.; Brechner, M.; Li, S.Y.; Wessely, O.; De Robertis, E.M. Isthmin Is a Novel Secreted Protein Expressed as Part of the Fgf-8 Synexpression Group in the Xenopus Midbrain-Hindbrain Organizer. Mech. Dev. 2002, 116, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Valle-Rios, R.; Maravillas-Montero, J.L.; Burkhardt, A.M.; Martinez, C.; Buhren, B.A.; Homey, B.; Gerber, P.A.; Robinson, O.; Hevezi, P.; Zlotnik, A. Isthmin 1 Is a Secreted Protein Expressed in Skin, Mucosal Tissues, and NK, NKT, and Th17 Cells. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 2014, 34, 795–801. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhao, M.; Voilquin, L.; Jung, Y.; Aikio, M.A.; Sahai, T.; Dou, F.Y.; Roche, A.M.; Carcamo-Orive, I.; Knowles, J.W.; et al. Isthmin-1 Is an Adipokine That Promotes Glucose Uptake and Improves Glucose Tolerance and Hepatic Steatosis. Cell Metab. 2021, 33, 1836–1852.e11. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Ke, Z.; Zhang, Y.; Cheng, G.H.-Y.; Irwan, I.D.; Sulochana, K.N.; Potturi, P.; Wang, Z.; Yang, H.; Wang, J.; et al. Isthmin Is a Novel Secreted Angiogenesis Inhibitor That Inhibits Tumour Growth in Mice. J. Cell. Mol. Med. 2011, 15, 359–374. [Google Scholar] [CrossRef]
- Venugopal, S.; Chen, M.; Liao, W.; Er, S.Y.; Wong, W.-S.F.; Ge, R. Isthmin Is a Novel Vascular Permeability Inducer That Functions through Cell-Surface GRP78-Mediated Src Activation. Cardiovasc. Res. 2015, 107, 131–142. [Google Scholar] [CrossRef]
- Li, J.; Xia, Y.; Huang, Z.; Zhao, Y.; Xiong, R.; Li, X.; Huang, Q.; Shan, F. Novel HIF-1-Target Gene Isthmin1 Contributes to Hypoxia-Induced Hyperpermeability of Pulmonary Microvascular Endothelial Cells Monolayers. Am. J. Physiol. Cell Physiol. 2021, 321, C671–C680. [Google Scholar] [CrossRef]
- Min, S.-Y.; Ha, D.-S.; Ha, T.-S. Puromycin Aminonucleoside Triggers Apoptosis in Podocytes by Inducing Endoplasmic Reticulum Stress. Kidney Res. Clin. Pract. 2018, 37, 210–221. [Google Scholar] [CrossRef]
- Inagi, R.; Kumagai, T.; Nishi, H.; Kawakami, T.; Miyata, T.; Fujita, T.; Nangaku, M. Preconditioning with Endoplasmic Reticulum Stress Ameliorates Mesangioproliferative Glomerulonephritis. J. Am. Soc. Nephrol. JASN 2008, 19, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Van Krieken, R.; Mehta, N.; Wang, T.; Zheng, M.; Li, R.; Gao, B.; Ayaub, E.; Ask, K.; Paton, J.C.; Paton, A.W.; et al. Cell Surface Expression of 78-KDa Glucose-Regulated Protein (GRP78) Mediates Diabetic Nephropathy. J. Biol. Chem. 2019, 294, 7755–7768. [Google Scholar] [CrossRef]
- Henderson, N.C.; Arnold, T.D.; Katamura, Y.; Giacomini, M.M.; Rodriguez, J.D.; McCarty, J.H.; Pellicoro, A.; Raschperger, E.; Betsholtz, C.; Ruminski, P.G.; et al. Targeting of Av Integrin Identifies a Core Molecular Pathway That Regulates Fibrosis in Several Organs. Nat. Med. 2013, 19, 1617–1624. [Google Scholar] [CrossRef] [PubMed]
- McCurley, A.; Alimperti, S.; Campos-Bilderback, S.B.; Sandoval, R.M.; Calvino, J.E.; Reynolds, T.L.; Quigley, C.; Mugford, J.W.; Polacheck, W.J.; Gomez, I.G.; et al. Inhibition of Avβ5 Integrin Attenuates Vascular Permeability and Protects against Renal Ischemia-Reperfusion Injury. J. Am. Soc. Nephrol. JASN 2017, 28, 1741–1752. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Atakilit, A.; Li, J.T.; Wu, N.; Luong, J.; Chen, R.; Bhattacharya, M.; Sheppard, D. Effective Treatment of Mouse Sepsis with an Inhibitory Antibody Targeting Integrin Avβ5. Crit. Care Med. 2013, 41, 546–553. [Google Scholar] [CrossRef]
- Caron, J.; Michel, P.-A.; Dussaule, J.-C.; Chatziantoniou, C.; Ronco, P.; Boffa, J.-J. Extracorporeal Shock Wave Therapy Does Not Improve Hypertensive Nephropathy. Physiol. Rep. 2016, 4, e12699. [Google Scholar] [CrossRef] [PubMed]
- Boffa, J.-J.; Lu, Y.; Placier, S.; Stefanski, A.; Dussaule, J.-C.; Chatziantoniou, C. Regression of Renal Vascular and Glomerular Fibrosis: Role of Angiotensin II Receptor Antagonism and Matrix Metalloproteinases. J. Am. Soc. Nephrol. JASN 2003, 14, 1132–1144. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, C.K.; Michellazzo, S.M.; de Nucci, G.; Zatz, R. Sodium Excess Aggravates Hypertension and Renal Parenchymal Injury in Rats with Chronic NO Inhibition. Am. J. Physiol. 1994, 266, F697–F705. [Google Scholar] [CrossRef] [PubMed]
- Guerrot, D.; Dussaule, J.-C.; Mael-Ainin, M.; Xu-Dubois, Y.-C.; Rondeau, E.; Chatziantoniou, C.; Placier, S. Identification of Periostin as a Critical Marker of Progression/Reversal of Hypertensive Nephropathy. PLoS ONE 2012, 7, e31974. [Google Scholar] [CrossRef]
- Faul, C.; Donnelly, M.; Merscher-Gomez, S.; Chang, Y.H.; Franz, S.; Delfgaauw, J.; Chang, J.-M.; Choi, H.Y.; Campbell, K.N.; Kim, K.; et al. The Actin Cytoskeleton of Kidney Podocytes Is a Direct Target of the Antiproteinuric Effect of Cyclosporine A. Nat. Med. 2008, 14, 931–938. [Google Scholar] [CrossRef]
- Chen, M.; Qiu, T.; Wu, J.; Yang, Y.; Wright, G.D.; Wu, M.; Ge, R. Extracellular Anti-Angiogenic Proteins Augment an Endosomal Protein Trafficking Pathway to Reach Mitochondria and Execute Apoptosis in HUVECs. Cell Death Differ. 2018, 25, 1905–1920. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Youle, R.J. The Role of Mitochondria in Apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Osório, L.; Wu, X.; Wang, L.; Jiang, Z.; Neideck, C.; Sheng, G.; Zhou, Z. ISM1 Regulates NODAL Signaling and Asymmetric Organ Morphogenesis during Development. J. Cell Biol. 2019, 218, 2388–2402. [Google Scholar] [CrossRef]
- Berrun, A.; Harris, E.; Stachura, D.L. Isthmin 1 (Ism1) Is Required for Normal Hematopoiesis in Developing Zebrafish. PLoS ONE 2018, 13, e0196872. [Google Scholar] [CrossRef] [PubMed]
- Hosoe-Nagai, Y.; Hidaka, T.; Sonoda, A.; Sasaki, Y.; Yamamoto-Nonaka, K.; Seki, T.; Asao, R.; Tanaka, E.; Trejo, J.A.O.; Kodama, F.; et al. Re-Expression of Sall1 in Podocytes Protects against Adriamycin-Induced Nephrosis. Lab. Investig. J. Techcol. Methods Pathol. 2017, 97, 1306–1320. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, M.; Venugopal, S.; Zhou, Y.; Xiang, W.; Li, Y.-H.; Lin, Q.; Kini, R.M.; Chong, Y.-S.; Ge, R. Isthmin Exerts Pro-Survival and Death-Promoting Effect on Endothelial Cells through Alphavbeta5 Integrin Depending on Its Physical State. Cell Death Dis. 2011, 2, e153. [Google Scholar] [CrossRef]
- Kachurina, N.; Chung, C.-F.; Benderoff, E.; Babayeva, S.; Bitzan, M.; Goodyer, P.; Kitzler, T.; Matar, D.; Cybulsky, A.V.; Alachkar, N.; et al. Novel Unbiased Assay for Circulating Podocyte-Toxic Factors Associated with Recurrent Focal Segmental Glomerulosclerosis. Am. J. Physiol. Renal Physiol. 2016, 310, F1148–F1156. [Google Scholar] [CrossRef]
- Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Silachev, D.N.; Pevzner, I.B.; Jankauskas, S.S.; Babenko, V.A.; Zorov, S.D.; Balakireva, A.V.; Juhaszova, M.; et al. Mitochondrial Membrane Potential. Anal. Biochem. 2018, 552, 50–59. [Google Scholar] [CrossRef]
- Povea-Cabello, S.; Oropesa-Ávila, M.; de la Cruz-Ojeda, P.; Villanueva-Paz, M.; de la Mata, M.; Suárez-Rivero, J.M.; Álvarez-Córdoba, M.; Villalón-García, I.; Cotán, D.; Ybot-González, P.; et al. Dynamic Reorganization of the Cytoskeleton during Apoptosis: The Two Coffins Hypothesis. Int. J. Mol. Sci. 2017, 18, 2393. [Google Scholar] [CrossRef]
- Lorenzo, H.K.; Susin, S.A. Mitochondrial Effectors in Caspase-Independent Cell Death. FEBS Lett. 2004, 557, 14–20. [Google Scholar] [CrossRef]
- Susin, S.A.; Lorenzo, H.K.; Zamzami, N.; Marzo, I.; Snow, B.E.; Brothers, G.M.; Mangion, J.; Jacotot, E.; Costantini, P.; Loeffler, M.; et al. Molecular Characterization of Mitochondrial Apoptosis-Inducing Factor. Nature 1999, 397, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Ow, Y.-L.P.; Green, D.R.; Hao, Z.; Mak, T.W. Cytochrome c: Functions beyond Respiration. Nat. Rev. Mol. Cell Biol. 2008, 9, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Pinedo, C.; Guío-Carrión, A.; Goldstein, J.C.; Fitzgerald, P.; Newmeyer, D.D.; Green, D.R. Different Mitochondrial Intermembrane Space Proteins Are Released during Apoptosis in a Manner That Is Coordinately Initiated but Can Vary in Duration. Proc. Natl. Acad. Sci. USA 2006, 103, 11573–11578. [Google Scholar] [CrossRef] [PubMed]
- Fransen, M.; Lismont, C.; Walton, P. The Peroxisome-Mitochondria Connection: How and Why? Int. J. Mol. Sci. 2017, 18, 1126. [Google Scholar] [CrossRef]
- Placier, S.; Boffa, J.-J.; Dussaule, J.-C.; Chatziantoniou, C. Reversal of Renal Lesions Following Interruption of Nitric Oxide Synthesis Inhibition in Transgenic Mice. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc.-Eur. Ren. Assoc. 2006, 21, 881–888. [Google Scholar] [CrossRef]
- Bolstad, B.M.; Irizarry, R.A.; Astrand, M.; Speed, T.P. A Comparison of Normalization Methods for High Density Oligonucleotide Array Data Based on Variance and Bias. Bioinforma. Oxf. Engl. 2003, 19, 185–193. [Google Scholar] [CrossRef]
- Regele, H.M.; Fillipovic, E.; Langer, B.; Poczewki, H.; Kraxberger, I.; Bittner, R.E.; Kerjaschki, D. Glomerular Expression of Dystroglycans Is Reduced in Minimal Change Nephrosis but Not in Focal Segmental Glomerulosclerosis. J. Am. Soc. Nephrol. JASN 2000, 11, 403–412. [Google Scholar] [CrossRef]
- Chatziantoniou, C.; Boffa, J.J.; Ardaillou, R.; Dussaule, J.C. Nitric Oxide Inhibition Induces Early Activation of Type I Collagen Gene in Renal Resistance Vessels and Glomeruli in Transgenic Mice. Role of Endothelin. J. Clin. Investig. 1998, 101, 2780–2789. [Google Scholar] [CrossRef]
- Saleem, M.A.; O’Hare, M.J.; Reiser, J.; Coward, R.J.; Inward, C.D.; Farren, T.; Xing, C.Y.; Ni, L.; Mathieson, P.W.; Mundel, P. A Conditionally Immortalized Human Podocyte Cell Line Demonstrating Nephrin and Podocin Expression. J. Am. Soc. Nephrol. JASN 2002, 13, 630–638. [Google Scholar] [CrossRef]
- Pavlidis, P.; Noble, W.S. Analysis of Strain and Regional Variation in Gene Expression in Mouse Brain. Genome Biol. 2001, 2, research0042.1. [Google Scholar] [CrossRef]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for Gene List Enrichment Analysis and Candidate Gene Prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef] [PubMed]
- Cline, M.S.; Smoot, M.; Cerami, E.; Kuchinsky, A.; Landys, N.; Workman, C.; Christmas, R.; Avila-Campilo, I.; Creech, M.; Gross, B.; et al. Integration of Biological Networks and Gene Expression Data Using Cytoscape. Nat. Protoc. 2007, 2, 2366–2382. [Google Scholar] [CrossRef] [PubMed]
- Brancotte, B.; Biton, A.; Bernard-Pierrot, I.; Radvanyi, F.; Reyal, F.; Cohen-Boulakia, S. Gene List Significance At-a-Glance with GeneValorization. Bioinforma. Oxf. Engl. 2011, 27, 1187–1189. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahiri, V.; Caron, J.; Roger, E.; Desterke, C.; Ghachem, K.; Mohamadou, I.; Serre, J.; Prakoura, N.; Fellahi, S.; Placier, S.; et al. The Angiogenesis Inhibitor Isthmin-1 (ISM1) Is Overexpressed in Experimental Models of Glomerulopathy and Impairs the Viability of Podocytes. Int. J. Mol. Sci. 2023, 24, 2723. https://doi.org/10.3390/ijms24032723
Sahiri V, Caron J, Roger E, Desterke C, Ghachem K, Mohamadou I, Serre J, Prakoura N, Fellahi S, Placier S, et al. The Angiogenesis Inhibitor Isthmin-1 (ISM1) Is Overexpressed in Experimental Models of Glomerulopathy and Impairs the Viability of Podocytes. International Journal of Molecular Sciences. 2023; 24(3):2723. https://doi.org/10.3390/ijms24032723
Chicago/Turabian StyleSahiri, Virgilia, Jonathan Caron, Elena Roger, Christophe Desterke, Khalil Ghachem, Inna Mohamadou, Justine Serre, Niki Prakoura, Soraya Fellahi, Sandrine Placier, and et al. 2023. "The Angiogenesis Inhibitor Isthmin-1 (ISM1) Is Overexpressed in Experimental Models of Glomerulopathy and Impairs the Viability of Podocytes" International Journal of Molecular Sciences 24, no. 3: 2723. https://doi.org/10.3390/ijms24032723
APA StyleSahiri, V., Caron, J., Roger, E., Desterke, C., Ghachem, K., Mohamadou, I., Serre, J., Prakoura, N., Fellahi, S., Placier, S., Adriouch, S., Zhang, L., Chadjichristos, C. E., Chatziantoniou, C., Lorenzo, H. K., & Boffa, J.-J. (2023). The Angiogenesis Inhibitor Isthmin-1 (ISM1) Is Overexpressed in Experimental Models of Glomerulopathy and Impairs the Viability of Podocytes. International Journal of Molecular Sciences, 24(3), 2723. https://doi.org/10.3390/ijms24032723