

Inflammatory Treatment Used to Mimic Osteoarthritis and Patients’ Synovial Fluid Have Divergent Molecular Impact on Chondrocytes In Vitro



Abstract
:1. Introduction
2. Results
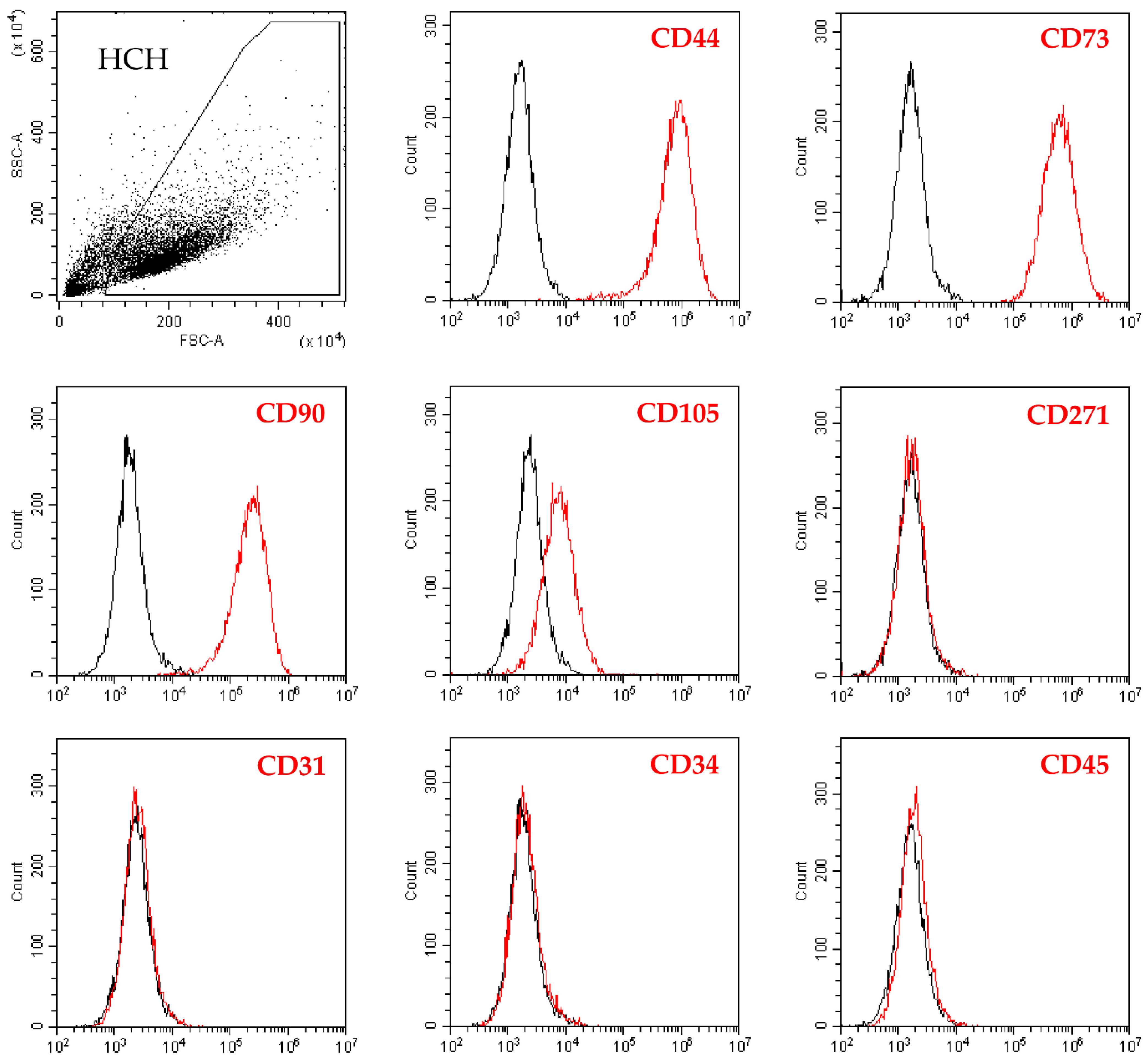
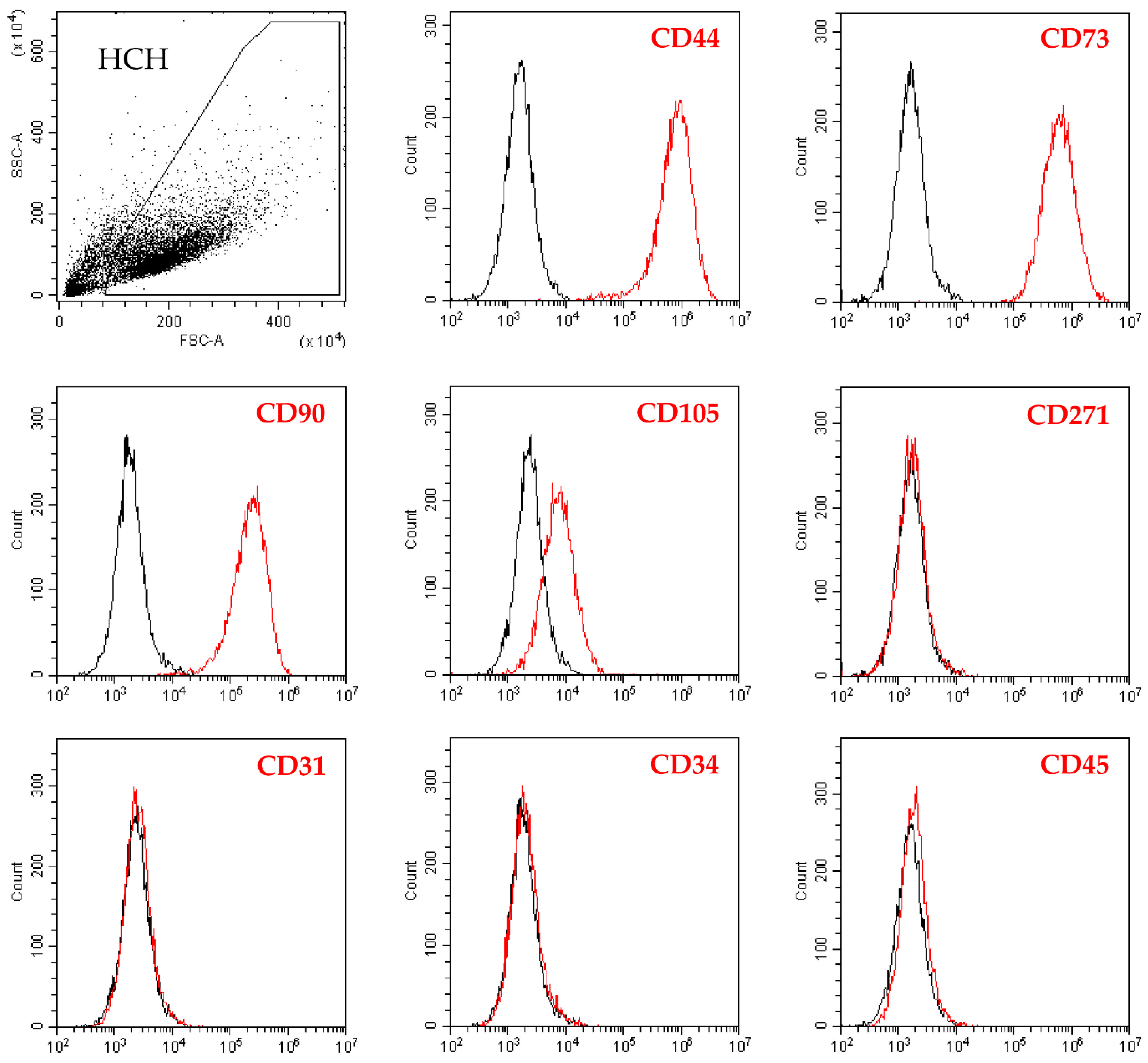
2.1. Flow Cytometry Analysis of Isolated Chondrocytes
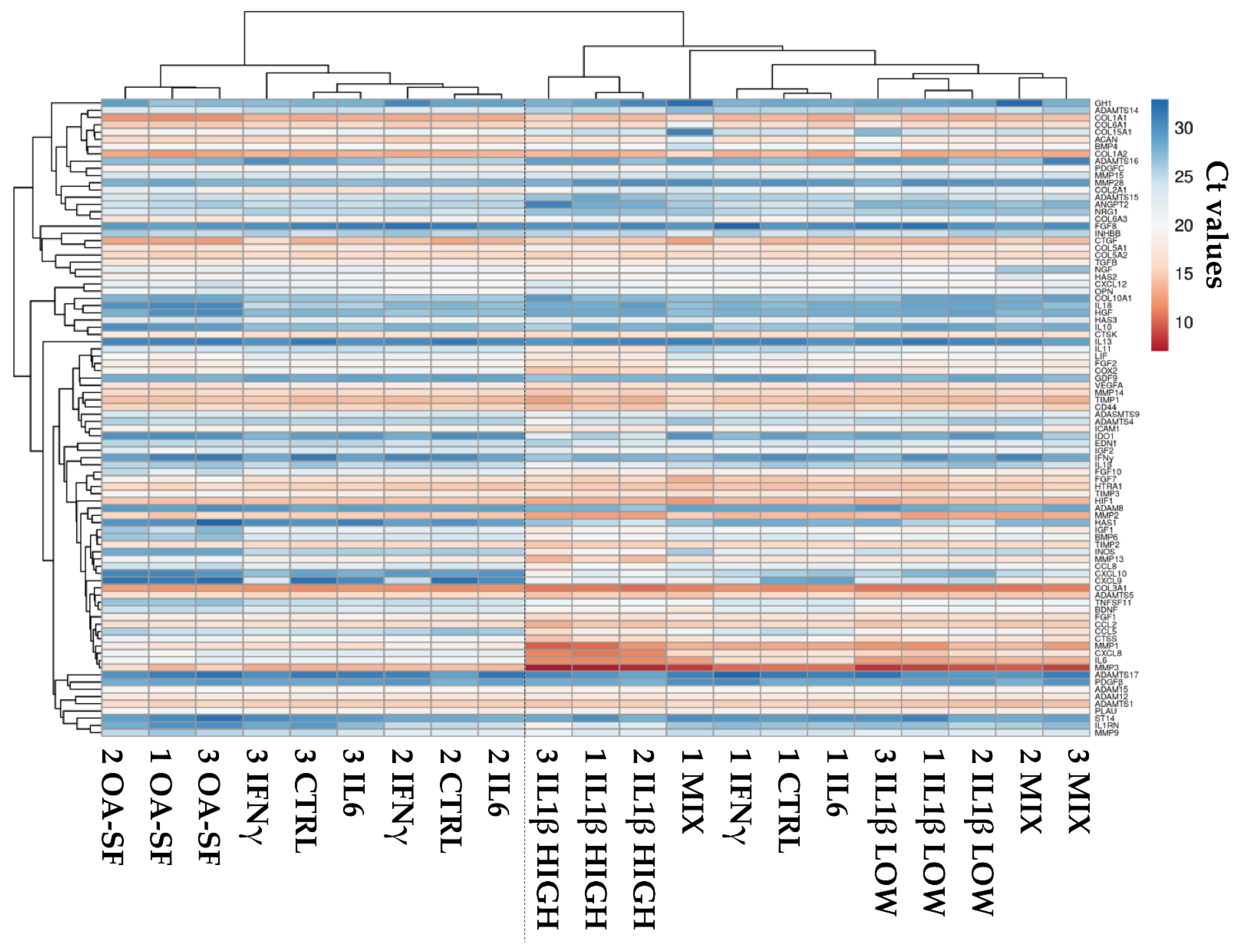
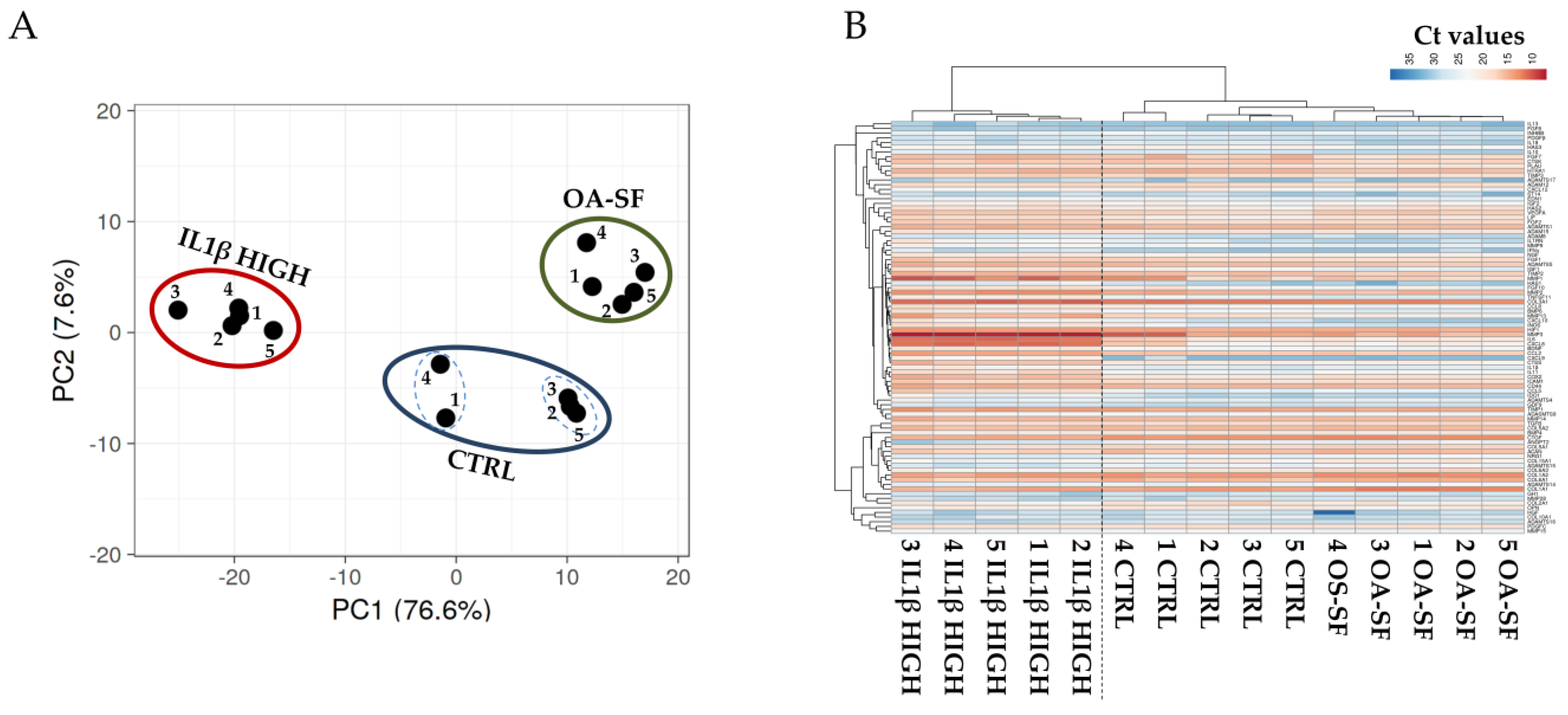
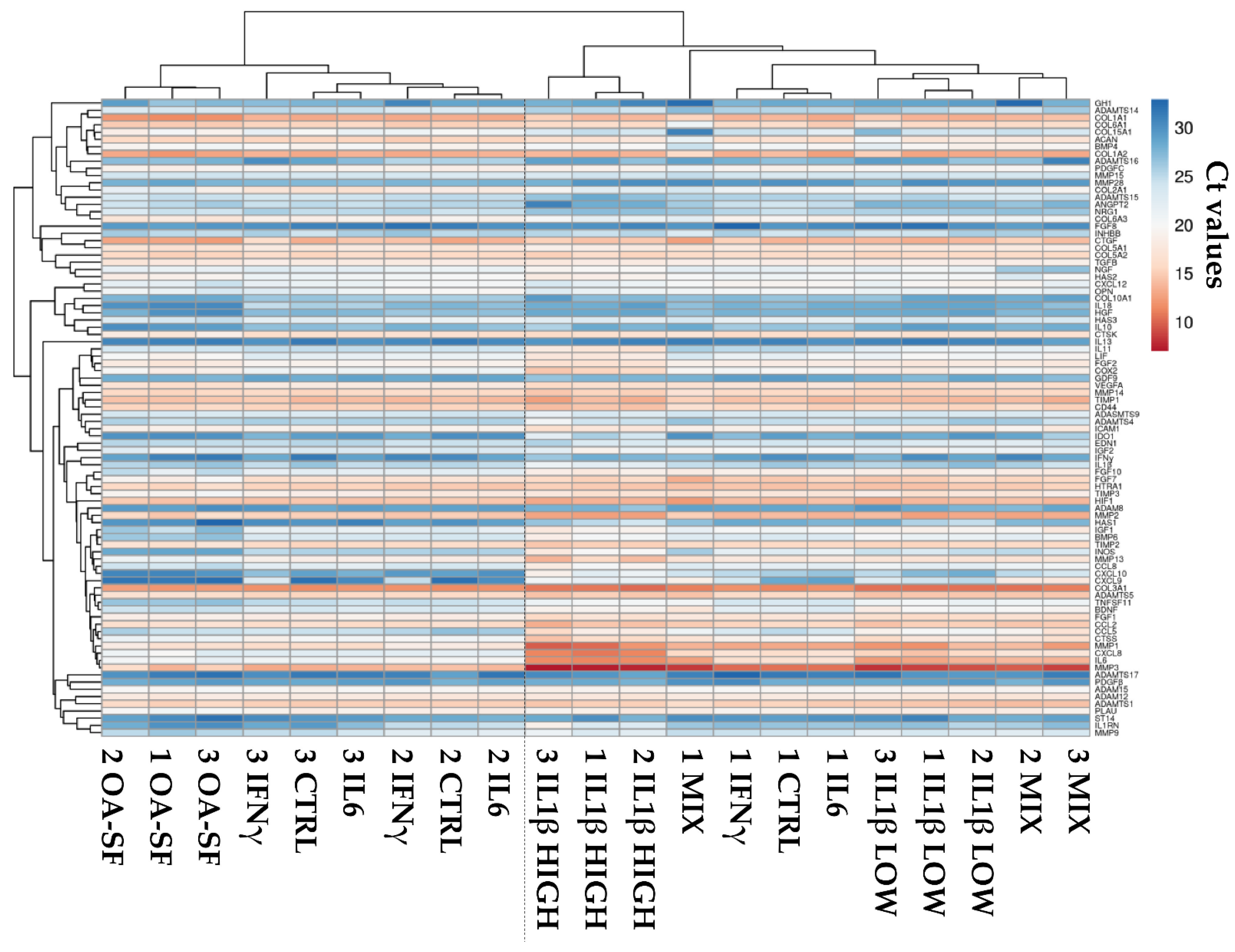
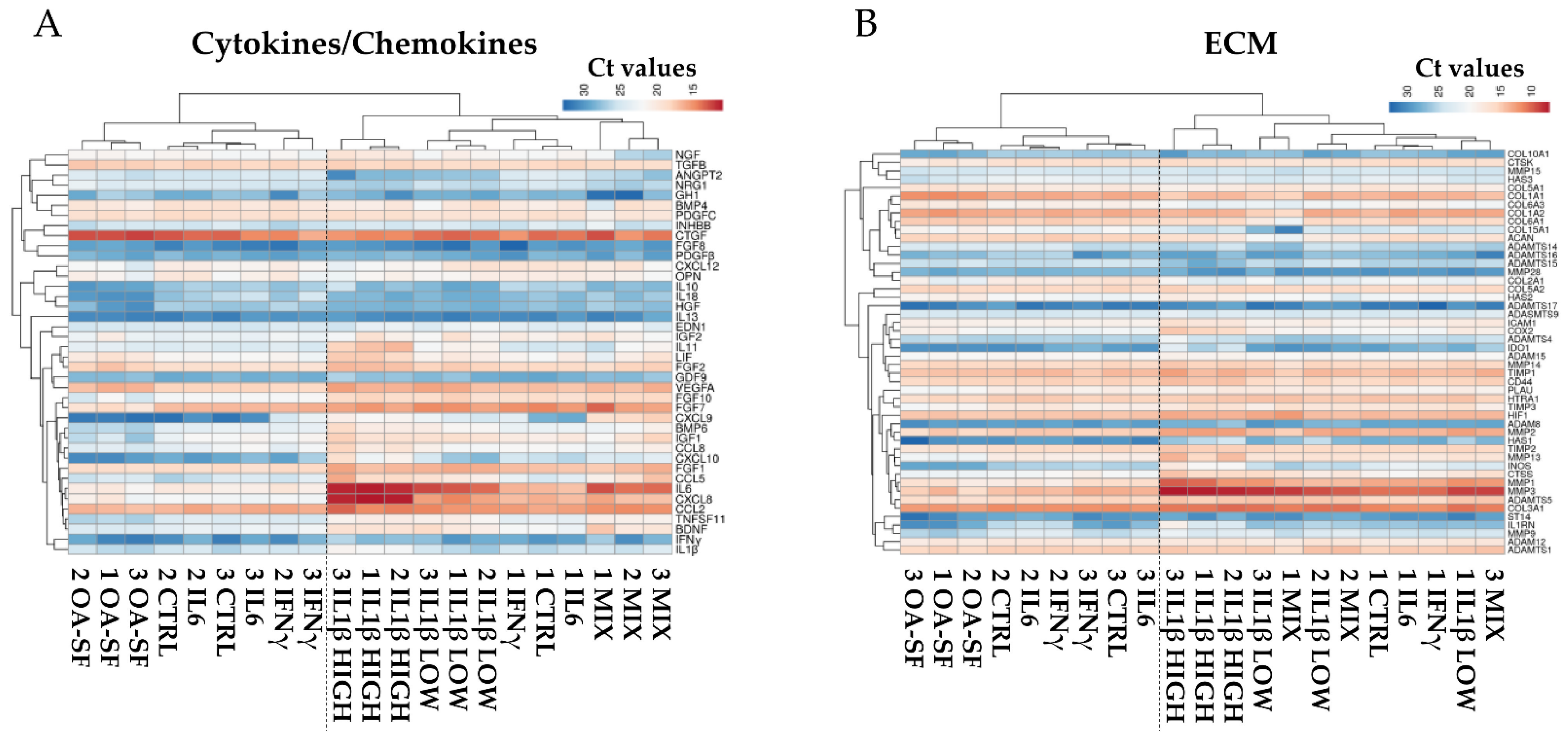
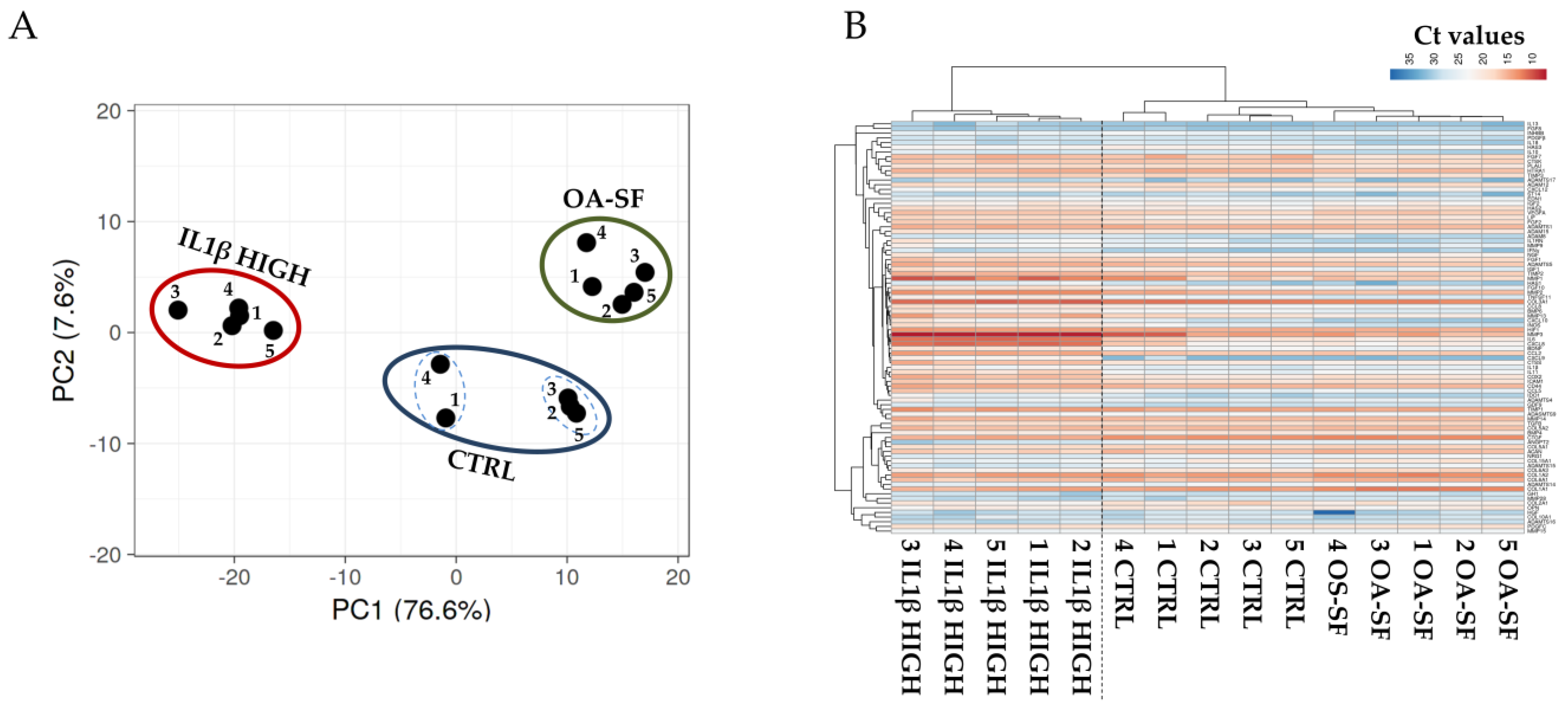
2.2. Donors and Treatments Drive Gene Expression Profile
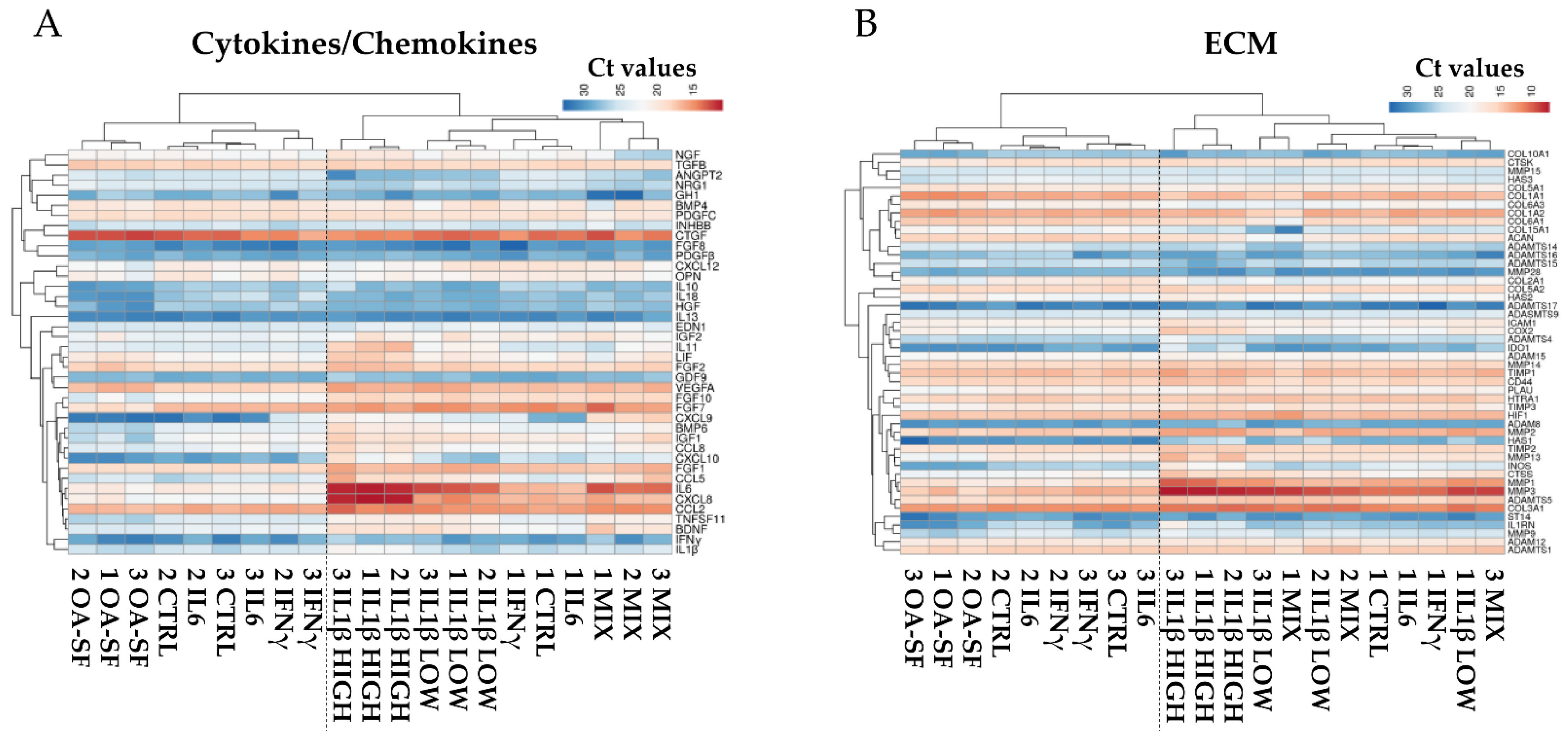
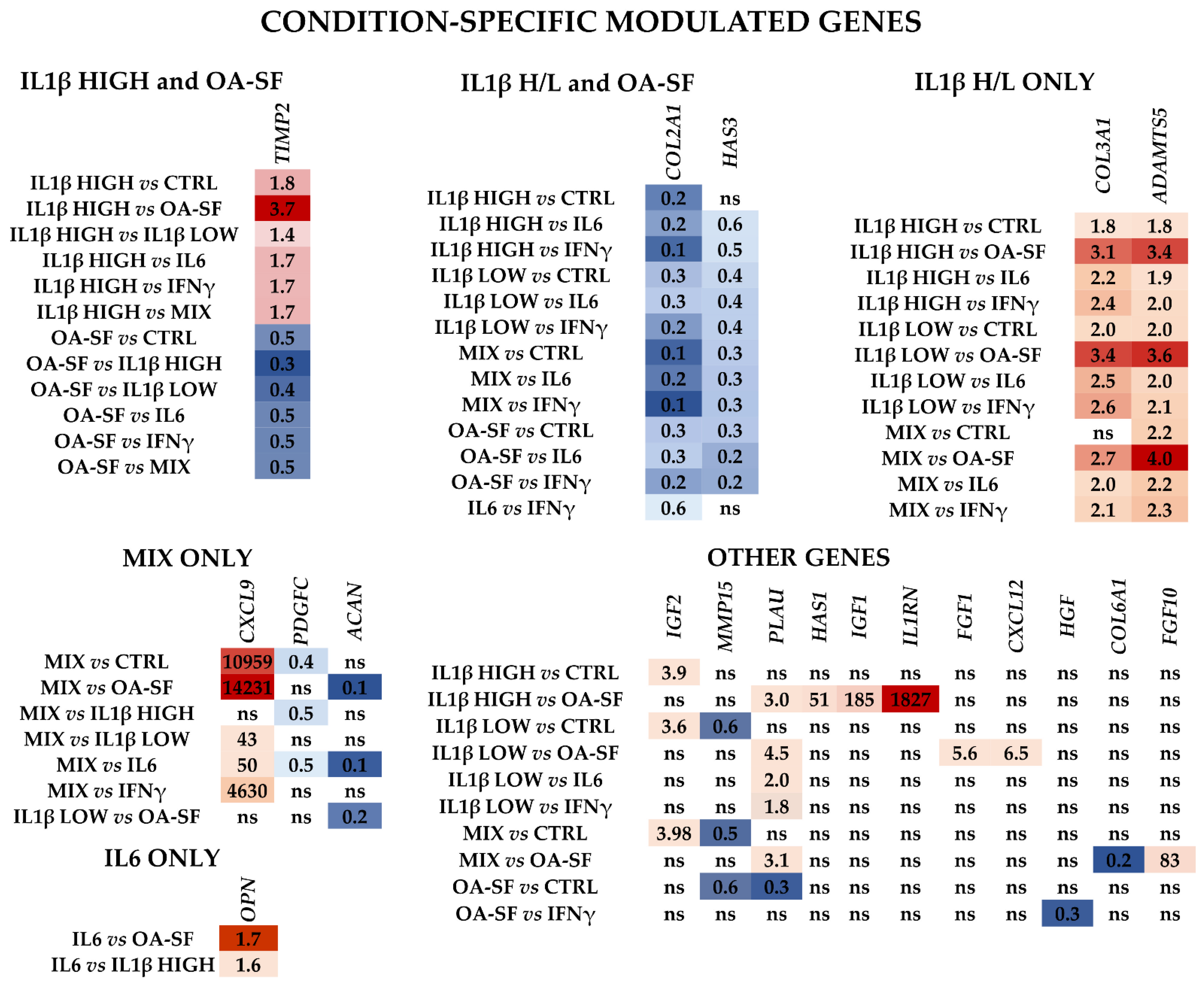
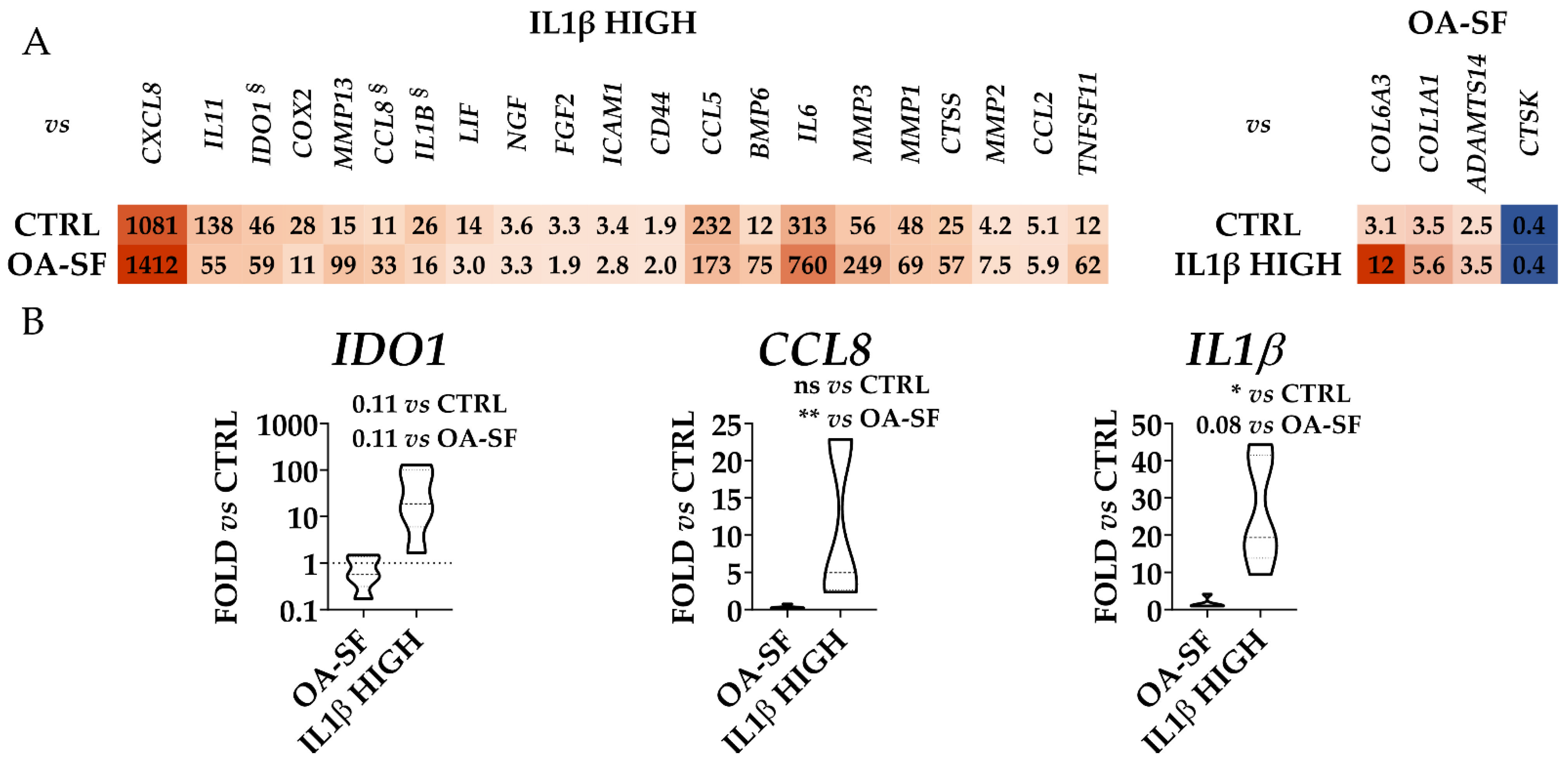
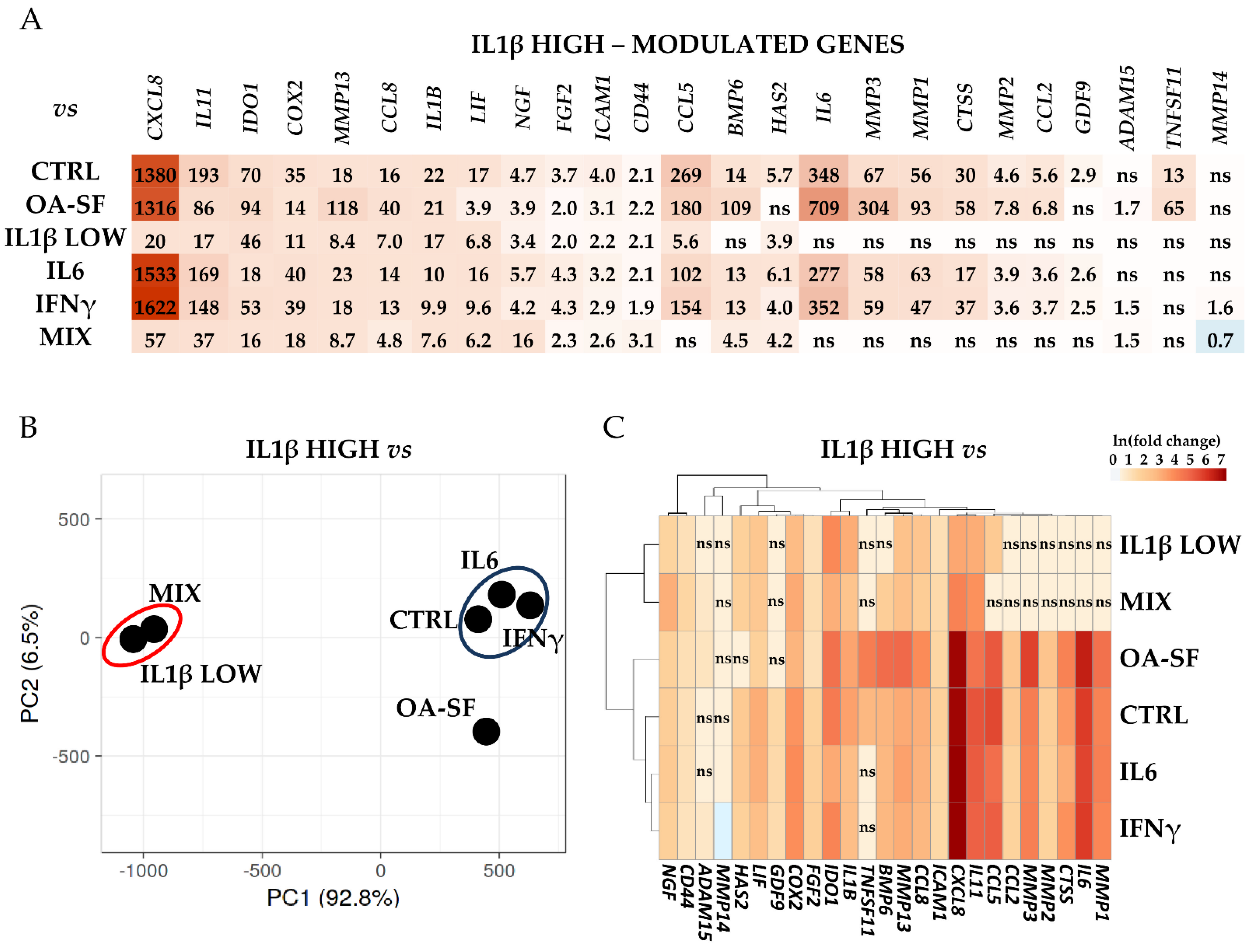
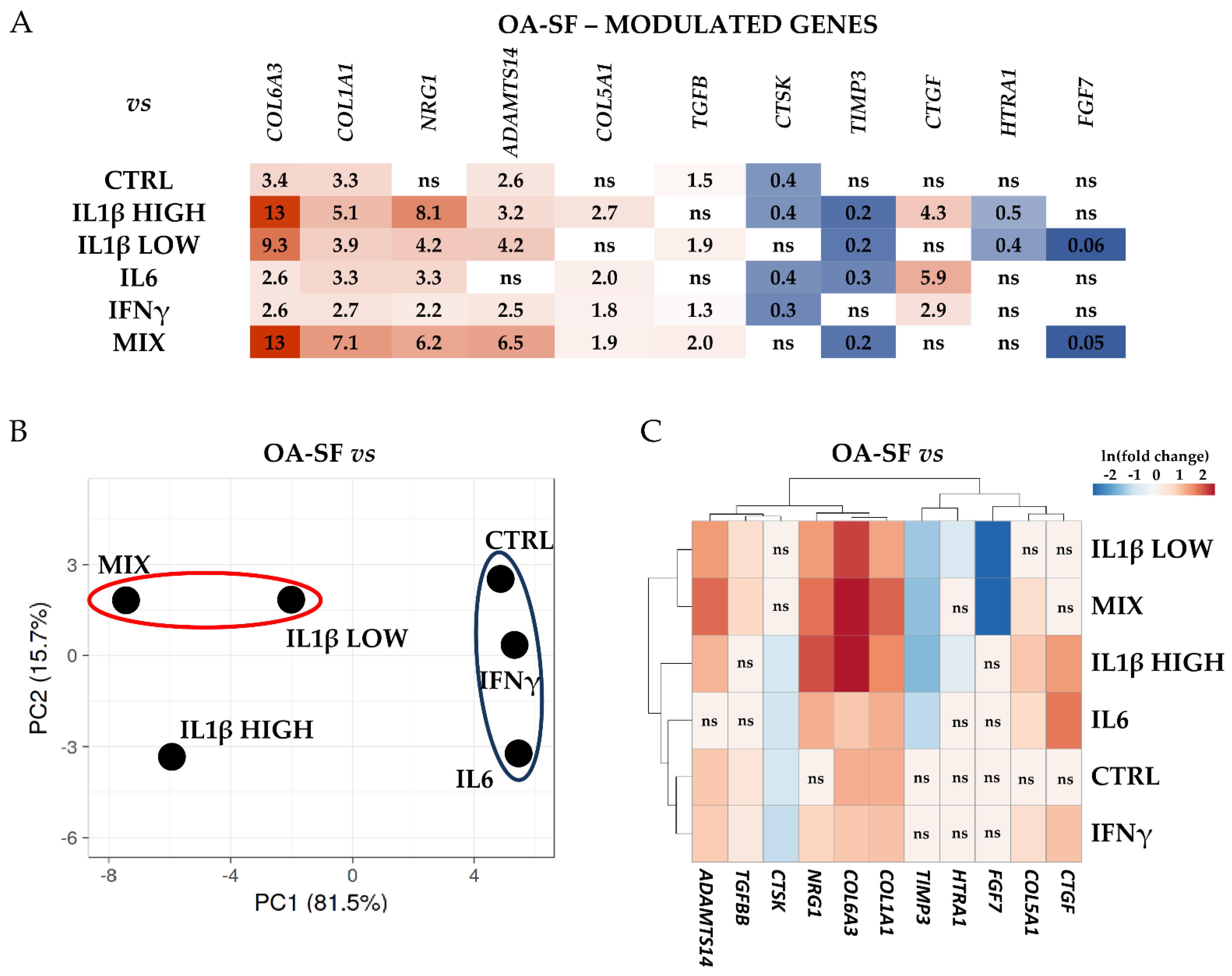
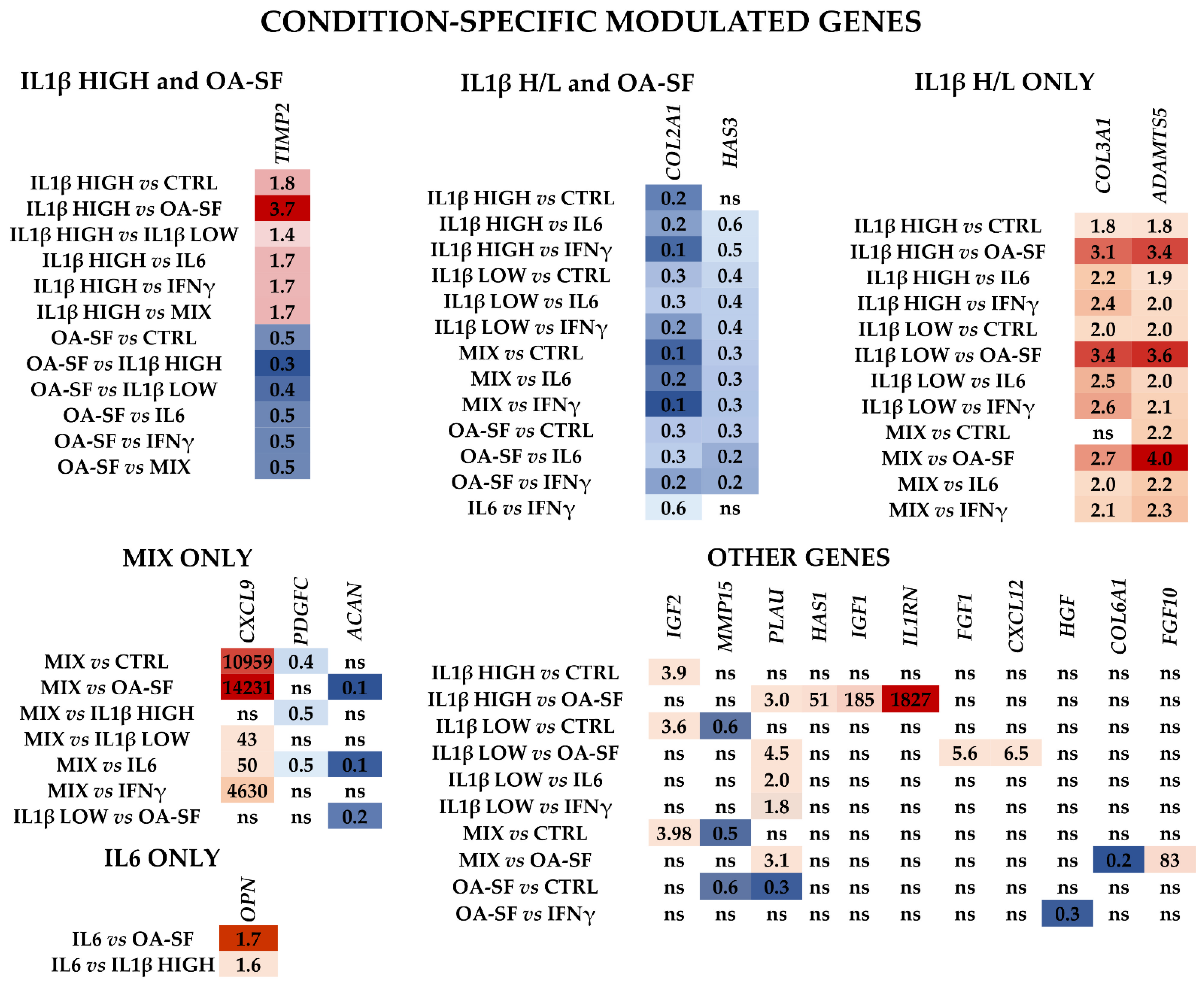
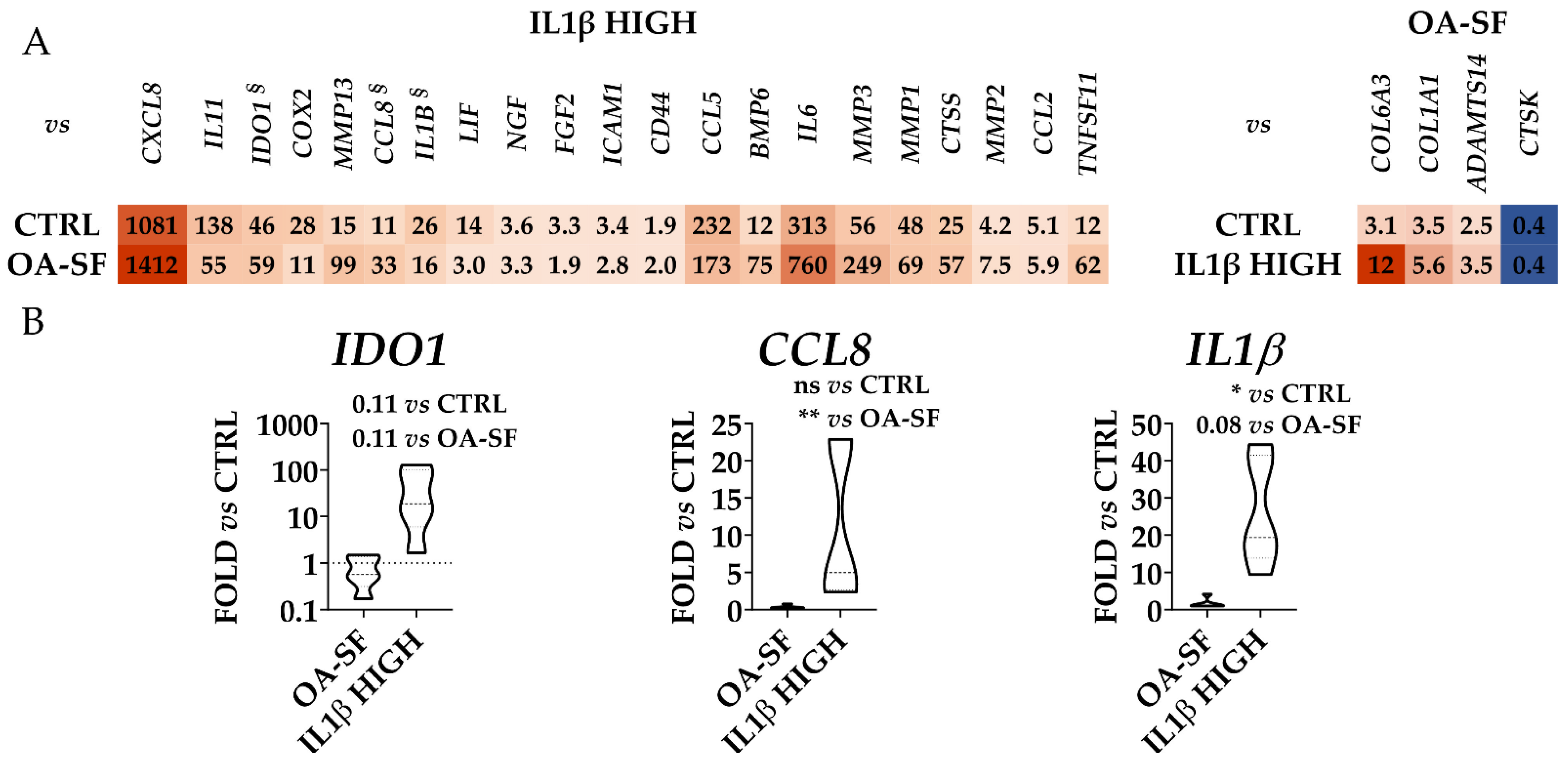
2.3. IL1β and OA-SF Account for the Strongest Modulation of OA-Related Factors
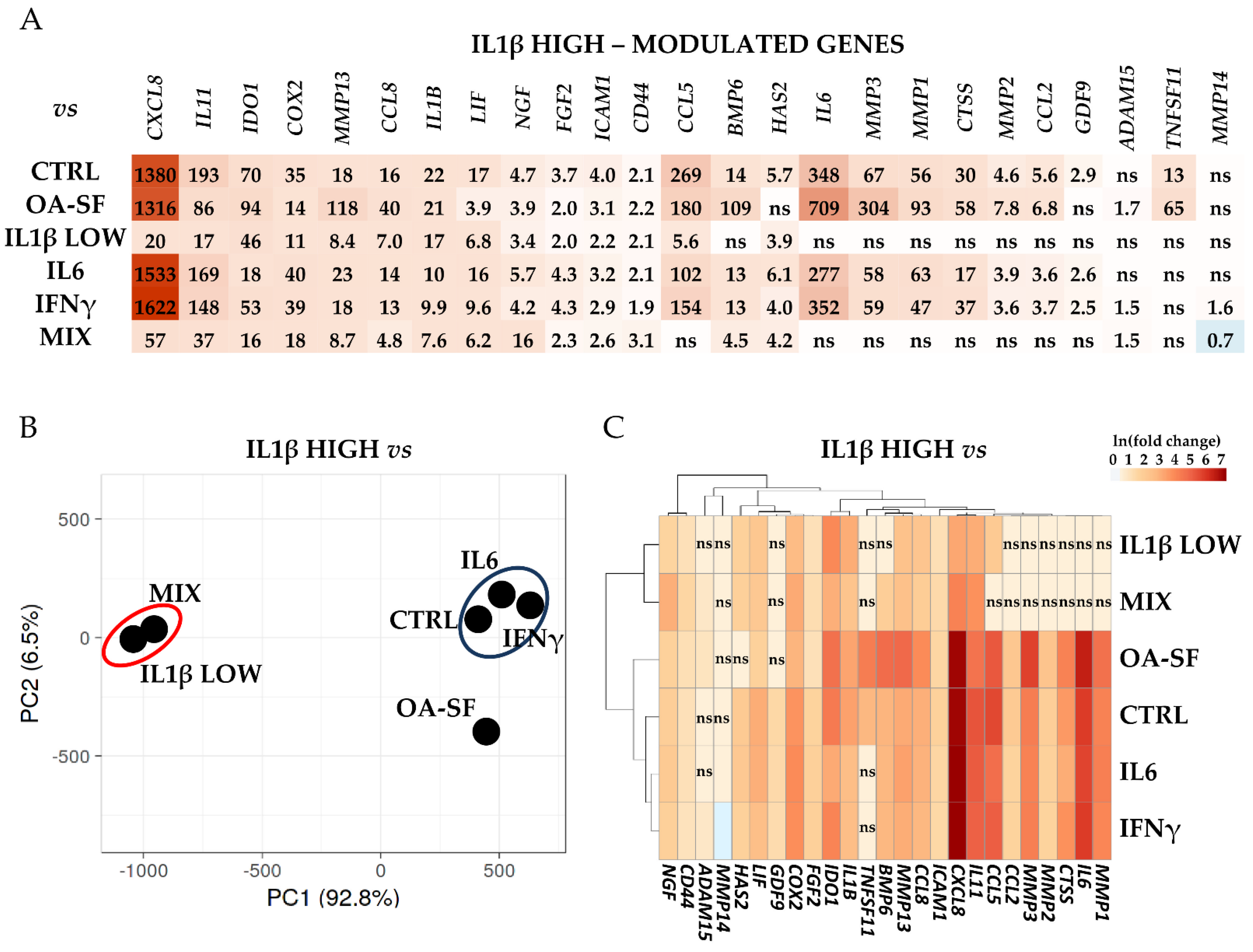
2.4. Validation of Identified IL1β High and OA-SF-Modulated Genes
3. Discussion
4. Materials and Methods
4.1. Isolation and Expansion of Human Articular Chondrocytes
4.2. Flow Cytometry
4.3. Cell Treatments
4.4. RNA Isolation and cDNA Synthesis
4.5. Gene Expression
4.6. Computational Anlyses
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J. Osteoarthritis. Nat. Rev. Dis. Prim. 2016, 2, 16072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeser, R.F.; Collins, J.A.; Diekman, B.O. Ageing and the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 412–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathiessen, A.; Conaghan, P.G. Synovitis in osteoarthritis: Current understanding with therapeutic implications. Arthritis Res. Ther. 2017, 19, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nees, T.A.; Rosshirt, N.; Zhang, J.A.; Reiner, T.; Sorbi, R.; Tripel, E.; Walker, T.; Schiltenwolf, M.; Hagmann, S.; Moradi, B. Synovial Cytokines Significantly Correlate with Osteoarthritis-Related Knee Pain and Disability: Inflammatory Mediators of Potential Clinical Relevance. J. Clin. Med. 2019, 8, 1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragni, E.; Colombini, A.; Viganò, M.; Libonati, F.; Perucca Orfei, C.; Zagra, L.; de Girolamo, L. Cartilage Protective and Immunomodulatory Features of Osteoarthritis Synovial Fluid-Treated Adipose-Derived Mesenchymal Stem Cells Secreted Factors and Extracellular Vesicles-Embedded miRNAs. Cells 2021, 10, 1072. [Google Scholar] [CrossRef]
- Watt, F.E.; Gulati, M. New Drug Treatments for Osteoarthritis: What is on the Horizon? Eur. Med. J. Rheumatol. 2017, 2, 50–58. [Google Scholar] [CrossRef]
- Cotter, E.J.; Frank, R.M.; Mandelbaum, B. Management of osteoarthritis—Biological approaches: Current concepts. Curr. Concepts Rev. 2020, 5, 27–31. [Google Scholar] [CrossRef]
- Samvelyan, H.J.; Hughes, D.; Stevens, C.; Staines, K.A. Models of Osteoarthritis: Relevance and New Insights. Calcif. Tissue Int. 2021, 109, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ma, S.; Su, H.; Cheng, J. Isoliquiritigenin Inhibits IL-1β-Induced Production of Matrix Metalloproteinase in Articular Chondrocytes. Mol. Ther.-Methods Clin. Dev. 2018, 9, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Stöckl, S.; Lukas, C.; Götz, J.; Herrmann, M.; Federlin, M.; Grässel, S. hBMSC-Derived Extracellular Vesicles Attenuate IL-1β-Induced Catabolic Effects on OA-Chondrocytes by Regulating Pro-inflammatory Signaling Pathways. Front. Bioeng. Biotechnol. 2020, 8, 603598. [Google Scholar] [CrossRef]
- Stokes, D.G.; Liu, G.; Coimbra, I.B.; Piera-Velazquez, S.; Crowl, R.M. Sergio A Jiménez Assessment of the gene expression profile of differentiated and dedifferentiated human fetal chondrocytes by microarray analysis. Arthritis Rheumatol. 2002, 46, 404–419. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Deng, Z.; Chen, K.; Jian, S.; Zhou, F.; Yang, Y.; Fu, Z.; Xie, H.; Xiong, J.; Zhu, W. Cartilage tissue engineering: From proinflammatory and anti-inflammatory cytokines to osteoarthritis treatments (Review). Mol. Med. Rep. 2022, 25, 99. [Google Scholar] [CrossRef] [PubMed]
- Tarricone, E.; Mattiuzzo, E.; Belluzzi, E.; Elia, R.; Benetti, A.; Venerando, R.; Vindigni, V.; Ruggieri, P.; Brun, P. Anti-Inflammatory Performance of Lactose-Modified Chitosan and Hyaluronic Acid Mixtures in an In Vitro Macrophage-Mediated Inflammation Osteoarthritis Model. Cells 2020, 9, 1328. [Google Scholar] [CrossRef]
- Kokebie, R.; Aggarwal, R.; Lidder, S.; Hakimiyan, A.A.; Rueger, D.C.; Block, J.A.; Chubinskaya, S. The role of synovial fluid markers of catabolism and anabolism in osteoarthritis, rheumatoid arthritis and asymptomatic organ donors. Arthritis Res. Ther. 2011, 13, R50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, M.; Zhou, Y.; Polson, S.W.; Wan, L.Q.; Wang, M.; Han, L.; Wang, L.; Lu, X.L. Identification of Chondrocyte Genes and Signaling Pathways in Response to Acute Joint Inflammation. Sci. Rep. 2019, 9, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Fu, N.; Cai, L.; Gong, T.; Li, G.; Peng, Q.; Cai, X. The effects of interleukin-1β in modulating osteoclast-conditioned medium’s influence on gelatinases in chondrocytes through mitogen-activated protein kinases. Int. J. Oral Sci. 2015, 7, 220–231. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Söder, S.; Oehler, S.; Fundel, K.; Aigner, T. Activation of interleukin-1 signaling cascades in normal and osteoarthritic articular cartilage. Am. J. Pathol. 2007, 171, 938–946. [Google Scholar] [CrossRef] [Green Version]
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, W.; Yong, H.; He, M.; Yang, Y.; Deng, Z.; Li, Y. Macrophages in osteoarthritis: Pathophysiology and therapeutics. Am. J. Transl. Res. 2020, 12, 261–268. [Google Scholar]
- Rahmati, M.; Mobasheri, A.; Mozafari, M. Inflammatory mediators in osteoarthritis: A critical review of the state-of-the-art, current prospects, and future challenges. Bone 2016, 85, 81–90. [Google Scholar] [CrossRef]
- Lambert, C.; Zappia, J.; Sanchez, C.; Florin, A.; Dubuc, J.; Henrotin, Y. The Damage-Associated Molecular Patterns (DAMPs) as Potential Targets to Treat Osteoarthritis: Perspectives From a Review of the Literature. Front. Med. 2021, 7, 607186. [Google Scholar] [CrossRef] [PubMed]
- Ragni, E.; Perucca Orfei, C.; Valli, F.; Zagra, L.; de Girolamo, L. Molecular Characterization of Secreted Factors and Extracellular Vesicles-Embedded miRNAs from Bone Marrow-Derived Mesenchymal Stromal Cells in Presence of Synovial Fluid from Osteoarthritis Patients. Biology 2022, 11, 1632. [Google Scholar] [CrossRef] [PubMed]
- Manicourt, D.H.; Poilvache, P.; Van Egeren, A.; Devogelaer, J.P.; Lenz, M.E.; Thonar, E.J. Synovial fluid levels of tumor necrosis factor alpha and oncostatin M correlate with levels of markers of the degradation of crosslinked collagen and cartilage aggrecan in rheumatoid arthritis but not in osteoarthritis. Arthritis Rheumatol. 2000, 3, 281–288. [Google Scholar] [CrossRef]
- Bartolotti, I.; Roseti, L.; Petretta, M.; Grigolo, B.; Desando, G. A Roadmap of In Vitro Models in Osteoarthritis: A Focus on Their Biological Relevance in Regenerative Medicine. J. Clin. Med. 2021, 10, 1920. [Google Scholar] [CrossRef] [PubMed]
- Bunning, R.A.; Russell, R.G.; Van Damme, J. Independent induction of interleukin 6 and prostaglandin E2 by interleukin 1 in human articular chondrocytes. Biochem. Biophys. Res. Commun. 1990, 166, 1163–1170. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, M.; Saas, J.; Sohler, F.; Haag, J.; Söder, S.; Pieper, M.; Bartnik, E.; Beninga, J.; Zimmer, R.; Aigner, T. Comparison of the chondrosarcoma cell line SW1353 with primary human adult articular chondrocytes with regard to their gene expression profile and reactivity to IL-1beta. Osteoarthr. Cartil. 2005, 13, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Löfgren, M.; Svala, E.; Lindahl, A.; Skiöldebrand, E.; Ekman, S. Time-dependent changes in gene expression induced in vitro by interleukin-1β in equine articular cartilage. Res. Vet. Sci. 2018, 118, 466–476. [Google Scholar] [CrossRef]
- Toegel, S.; Wu, S.Q.; Piana, C.; Unger, F.M.; Wirth, M.; Goldring, M.B.; Gabor, F.; Viernstein, H. Comparison between chondroprotective effects of glucosamine, curcumin, and diacerein in IL-1beta-stimulated C-28/I2 chondrocytes. Osteoarthr. Cartil. 2008, 16, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhou, L.; Wang, Z.; Gao, W.; Chen, W.; Zhang, H.; Jing, B.; Zhu, X.; Chen, L.; Zheng, C.; et al. Eradication of specific donor-dependent variations of mesenchymal stem cells in immunomodulation to enhance therapeutic values. Cell Death Dis. 2021, 12, 357. [Google Scholar] [CrossRef]
- Hoff, P.; Buttgereit, F.; Burmester, G.; Jakstadt, M.; Gaber, T.; Andreas, K.; Matziolis, G.; Perka, C.; Röhner, E. Osteoarthritis synovial fluid activates pro-inflammatory cytokines in primary human chondrocytes. Int. Orthop. 2013, 37, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Housmans, B.A.C.; Neefjes, M.; Surtel, D.A.M.; Vitík, M.; Cremers, A.; van Rhijn, L.W.; van der Kraan, P.M.; van den Akker, G.G.H.; Welting, T.J.M. Synovial fluid from end-stage osteoarthritis induces proliferation and fibrosis of articular chondrocytes via MAPK and RhoGTPase signaling. Osteoarthr. Cartil. 2022, 30, 862–874. [Google Scholar] [CrossRef]
- Fraser, J.R.; Clarris, B.J.; Baxter, E. Patterns of induced variation in the morphology, hyaluronic acid secretion, and lysosomal enzyme activity of cultured human synovial cells. Ann. Rheum. Dis. 1979, 38, 287–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollander, A.P.; Pidoux, I.; Reiner, A.; Rorabeck, C.; Bourne, R.; Poole, A.R. Damage to type II collagen in aging and osteoarthritis starts at the articular surface, originates around chondrocytes, and extends into the cartilage with progressive degeneration. J. Clin. Investig. 1995, 96, 2859–2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayram, B.; Limberg, A.K.; Salib, C.G.; Bettencourt, J.W.; Trousdale, W.H.; Lewallen, E.A.; Reina, N.; Paradise, C.R.; Thaler, R.; Morrey, M.E.; et al. Molecular pathology of human knee arthrofibrosis defined by RNA sequencing. Genomics 2020, 112, 2703–2712. [Google Scholar] [CrossRef]
- Li, C.; Luo, J.; Xu, X.; Zhou, Z.; Ying, S.; Liao, X.; Wu, K. Single cell sequencing revealed the underlying pathogenesis of the development of osteoarthritis. Genomics 2020, 757, 144939. [Google Scholar] [CrossRef] [PubMed]
- Chen, C. COX-2′s new role in inflammation. Nat. Chem. Biol. 2010, 6, 401–402. [Google Scholar] [CrossRef]
- Ogbechi, J.; Clanchy, F.I.; Huang, Y.; Topping, L.M.; Stone, T.W.; Williams, R.O. IDO activation, inflammation and musculoskeletal disease. Exp. Gerontol. 2020, 131, 110820. [Google Scholar] [CrossRef] [PubMed]
- Banner, L.R.; Patterson, P.H.; Allchorne, A.; Poole, S.; Woolf, C.J. Leukemia inhibitory factor is an anti-inflammatory and analgesic cytokine. J. Neurosci. 1998, 18, 5456–5462. [Google Scholar] [CrossRef] [Green Version]
- Viganò, M.; Ragni, E.; Di Matteo, B.; Manlio Gambaro, F.; Perucca Orfei, C.; Spinelli, G.; Colombini, A.; de Girolamo, L.; Kon, E. A single step, centrifuge-free method to harvest bone marrow highly concentrated in mesenchymal stem cells: Results of a pilot trial. Int. Orthop. 2022, 46, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTRL | OA-SF | IL1β HIGH | IL1β LOW | IL6 | IFNγ | MIX | |
|---|---|---|---|---|---|---|---|
| CTRL | 0.922 ± 0.026 | 0.727 ± 0.091 | 0.858 ± 0.060 | 0.984 ± 0.002 | 0.959 ± 0.011 | 0.799 ± 0.039 | |
| OA-SF | 0.657 ± 0.049 | 0.787 ± 0.049 | 0.920 ± 0.009 | 0.882 ± 0.023 | 0.702 ± 0.019 | ||
| IL1β HIGH | 0.908 ± 0.030 | 0.746 ± 0.080 | 0.779 ± 0.076 | 0.879 ± 0.013 | |||
| IL1β LOW | 0.874 ± 0.051 | 0.891 ± 0.043 | 0.927 ± 0.025 | ||||
| IL6 | 0.968 ± 0.008 | 0.798 ± 0.007 | |||||
| IFNγ | 0.859 ± 0.009 | ||||||
| MIX |
| Rank | CTRL | OA-SF | IL1β HIGH | IL1β LOW | IL6 | IFNγ | MIX | All Samples |
|---|---|---|---|---|---|---|---|---|
| 1 | COL3A1 | TIMP1 | ADAMTS5 | MMP14 | CCL2 | COL3A1 | CTSK | ADAM15 |
| 2 | COL5A1 | MMP15 | TGFB | COL3A1 | CD44 | ADAMTS1 | COL5A2 | MMP14 |
| 3 | TGFB | MMP14 | NRG1 | CD44 | MMP14 | ICAM1 | TIMP2 | TIMP2 |
| 4 | NRG1 | ADAM15 | ADAMTS1 | TIMP1 | ADAM15 | NRG1 | CCL2 | COL5A2 |
| 5 | CTSK | TNFSF11 | CD44 | PDGFC | GDF9 | TIMP2 | HAS3 | TIMP1 |
| 6 | PLAU | NGF | ADAM15 | ANGPT2 | COL1A1 | ADAM15 | TGFB | ADAM12 |
| 7 | ADAMTS5 | FGF1 | HIF1 | TIMP2 | COL6A1 | COL5A1 | ADAMTS5 | ADAMTS1 |
| 8 | CCL2 | CTSS | PDGFC | TGFB | TNFSF11 | COL5A2 | FGF10 | COL3A1 |
| 9 | CD44 | COL6A1 | BDNF | MMP13 | TIMP2 | PLAU | MMP13 | HTRA1 |
| 10 | IL13 | BMP6 | TIMP2 | FGF1 | TIMP1 | HIF1 | ADAM15 | CD44 |
| 79 | MMP28 | MMP3 | HAS1 | ADAMTS16 | HAS1 | IL1RN | HAS2 | MMP1 |
| 80 | CXCL9 | HGF | COL2A1 | IGF2 | IL18 | MMP3 | IDO1 | IL11 |
| 81 | FGF10 | CXCL8 | EDN1 | COL10A1 | IGF2 | COL15A1 | ADAMTS16 | INOS |
| 82 | MMP3 | GH1 | COL10A1 | COL6A1 | CXCL10 | IL6 | ACAN | MMP3 |
| 83 | COL2A1 | IGF2 | IDO1 | ST14 | MMP3 | GH1 | IL11 | CXCL10 |
| 84 | IL6 | CXCL12 | IGF2 | ACAN | COL2A1 | COL2A1 | GH1 | COL15A1 |
| 85 | IL1RN | ST14 | CXCL10 | CXCL10 | IL6 | CXCL10 | COL6A1 | CCL5 |
| 86 | MMP1 | HAS1 | ANGPT2 | HAS1 | MMP1 | ADAMTS16 | NGF | IL6 |
| 87 | COL15A1 | FGF10 | ADAMTS4 | BMP4 | INOS | MMP1 | BMP4 | CXCL8 |
| 88 | CXCL8 | COL2A1 | IL1RN | COL15A1 | CXCL8 | CXCL8 | COL15A1 | CXCL9 |
| CTRL | OA-SF | IL1β HIGH | IL1β LOW | IL6 | IFNγ | MIX | |
|---|---|---|---|---|---|---|---|
| CTRL | 0.198 | 0.223 | 0.361 | 0.414 | 0.610 | 0.124 | |
| OA-SF | 0.178 | 0.111 | 0.354 | 0.293 | 0.029 | ||
| IL1β HIGH | 0.229 | 0.260 | 0.283 | 0.100 | |||
| IL1β LOW | 0.177 | 0.295 | 0.179 | ||||
| IL6 | 0.543 | 0.100 | |||||
| IFNγ | 0.155 | ||||||
| MIX |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragni, E.; De Luca, P.; Valli, F.; Zagra, L.; de Girolamo, L. Inflammatory Treatment Used to Mimic Osteoarthritis and Patients’ Synovial Fluid Have Divergent Molecular Impact on Chondrocytes In Vitro. Int. J. Mol. Sci. 2023, 24, 2625. https://doi.org/10.3390/ijms24032625
Ragni E, De Luca P, Valli F, Zagra L, de Girolamo L. Inflammatory Treatment Used to Mimic Osteoarthritis and Patients’ Synovial Fluid Have Divergent Molecular Impact on Chondrocytes In Vitro. International Journal of Molecular Sciences. 2023; 24(3):2625. https://doi.org/10.3390/ijms24032625
Chicago/Turabian StyleRagni, Enrico, Paola De Luca, Federico Valli, Luigi Zagra, and Laura de Girolamo. 2023. "Inflammatory Treatment Used to Mimic Osteoarthritis and Patients’ Synovial Fluid Have Divergent Molecular Impact on Chondrocytes In Vitro" International Journal of Molecular Sciences 24, no. 3: 2625. https://doi.org/10.3390/ijms24032625
APA StyleRagni, E., De Luca, P., Valli, F., Zagra, L., & de Girolamo, L. (2023). Inflammatory Treatment Used to Mimic Osteoarthritis and Patients’ Synovial Fluid Have Divergent Molecular Impact on Chondrocytes In Vitro. International Journal of Molecular Sciences, 24(3), 2625. https://doi.org/10.3390/ijms24032625