
Polysaccharides from Holothuria leucospilota Relieve Loperamide-Induced Constipation Symptoms in Mice
 and
and
Abstract
1. Introduction
2. Results and Discussion
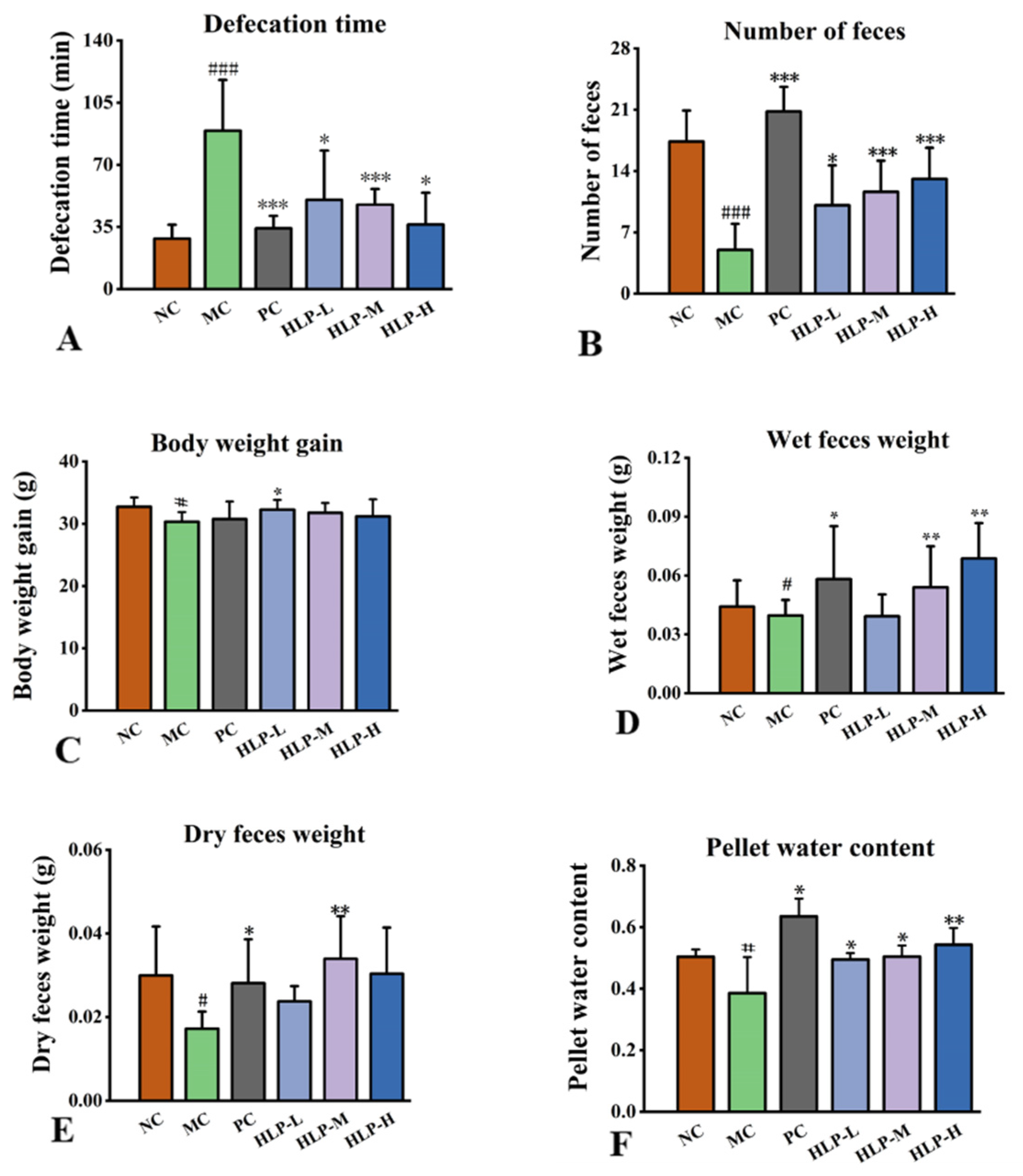
2.1. Effects of HLP on the Parameters Related to Defecation in Constipated Mice
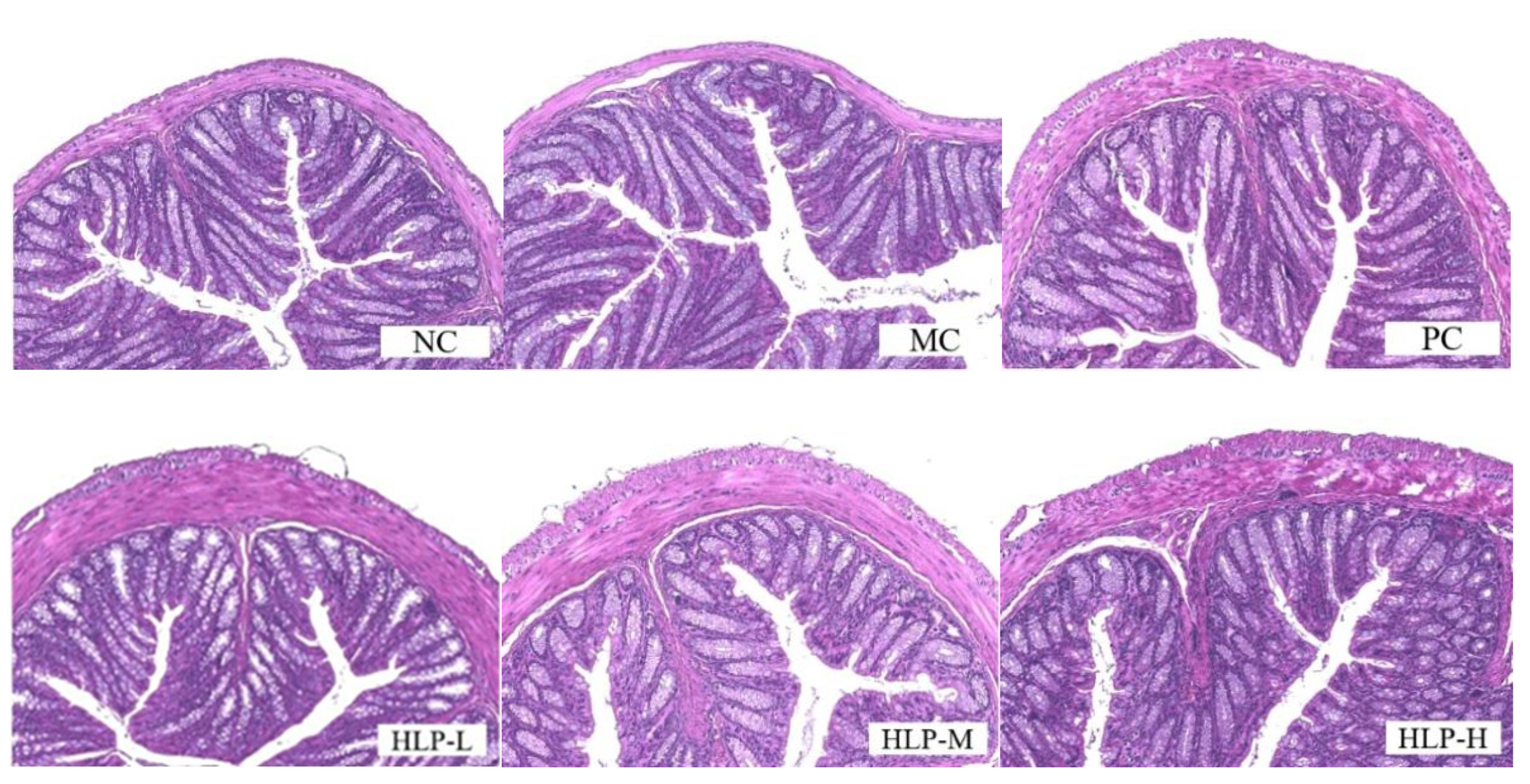
2.2. Effects of HLP on the Morphology of Colon Tissue in Constipated Mice
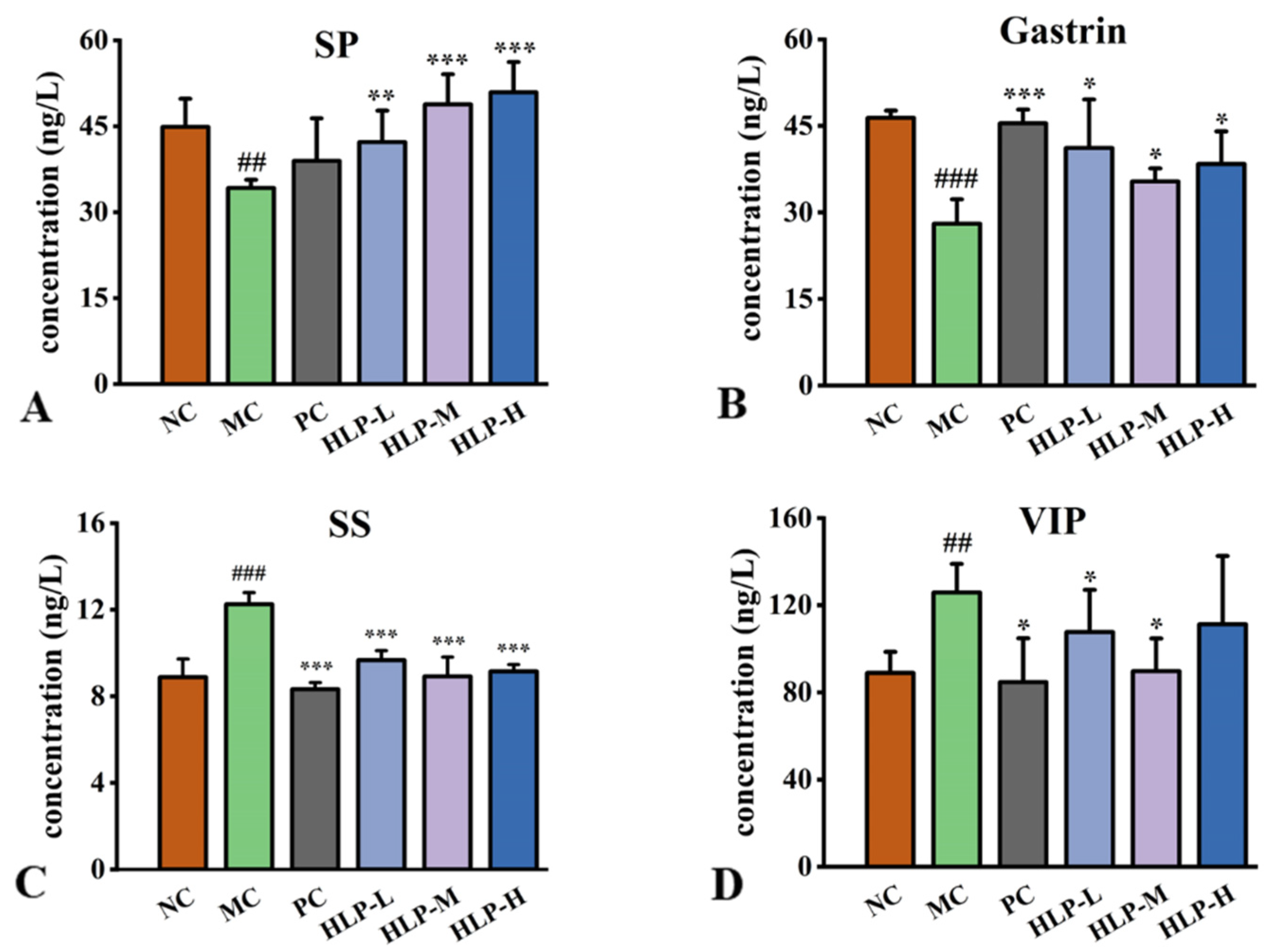
2.3. Effects of HLP on Regulatory Peptides Related to the Gastrointestinal Tract in Constipation Mice
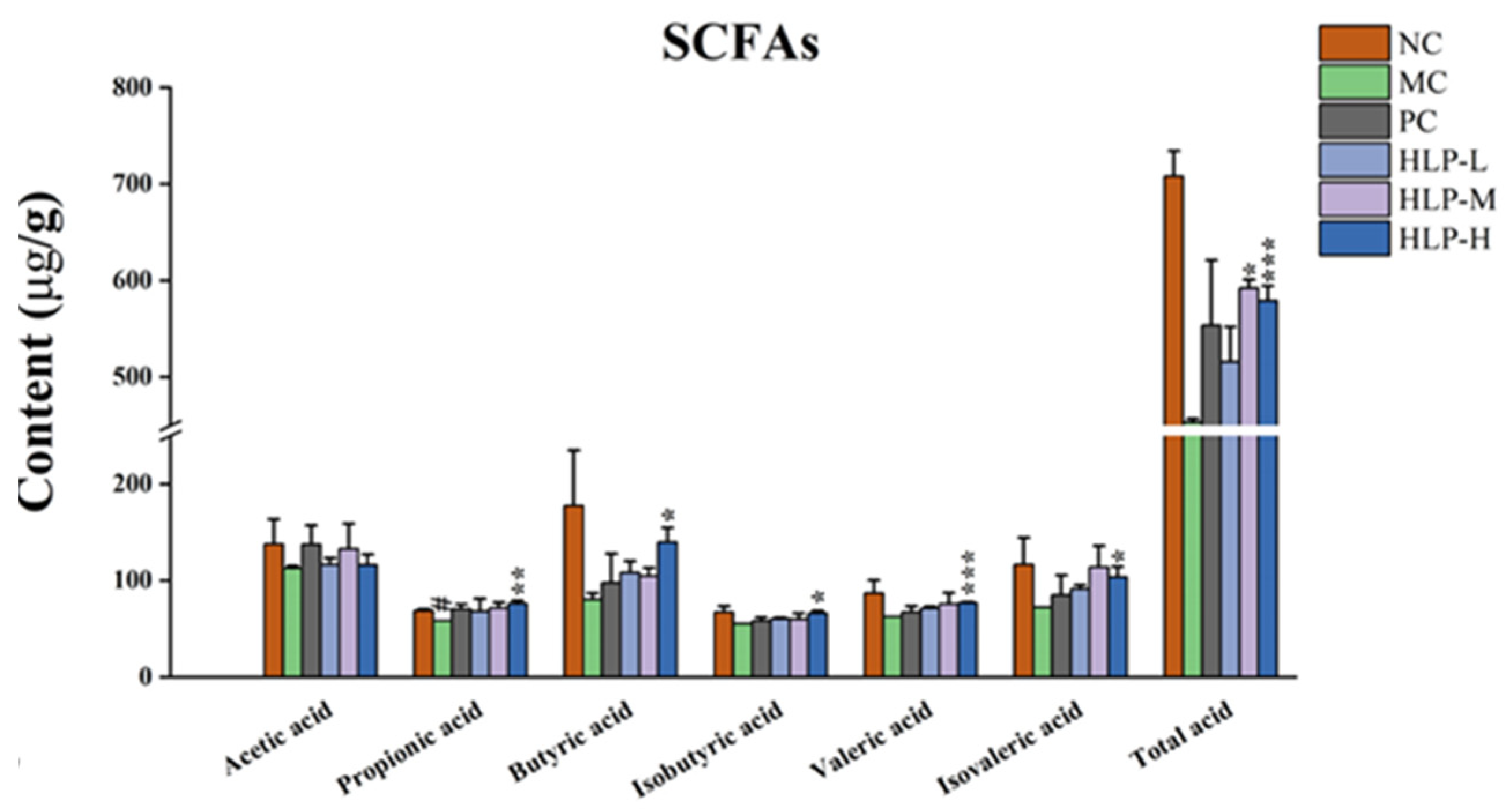
2.4. Effects of HLP on the Intestinal Metabolites of Constipated Mice
2.5. Effects of HLP on the Expression of Water Metabolism and Motility of Intestinal Tract-Related Genes and Proteins in Constipated Mice
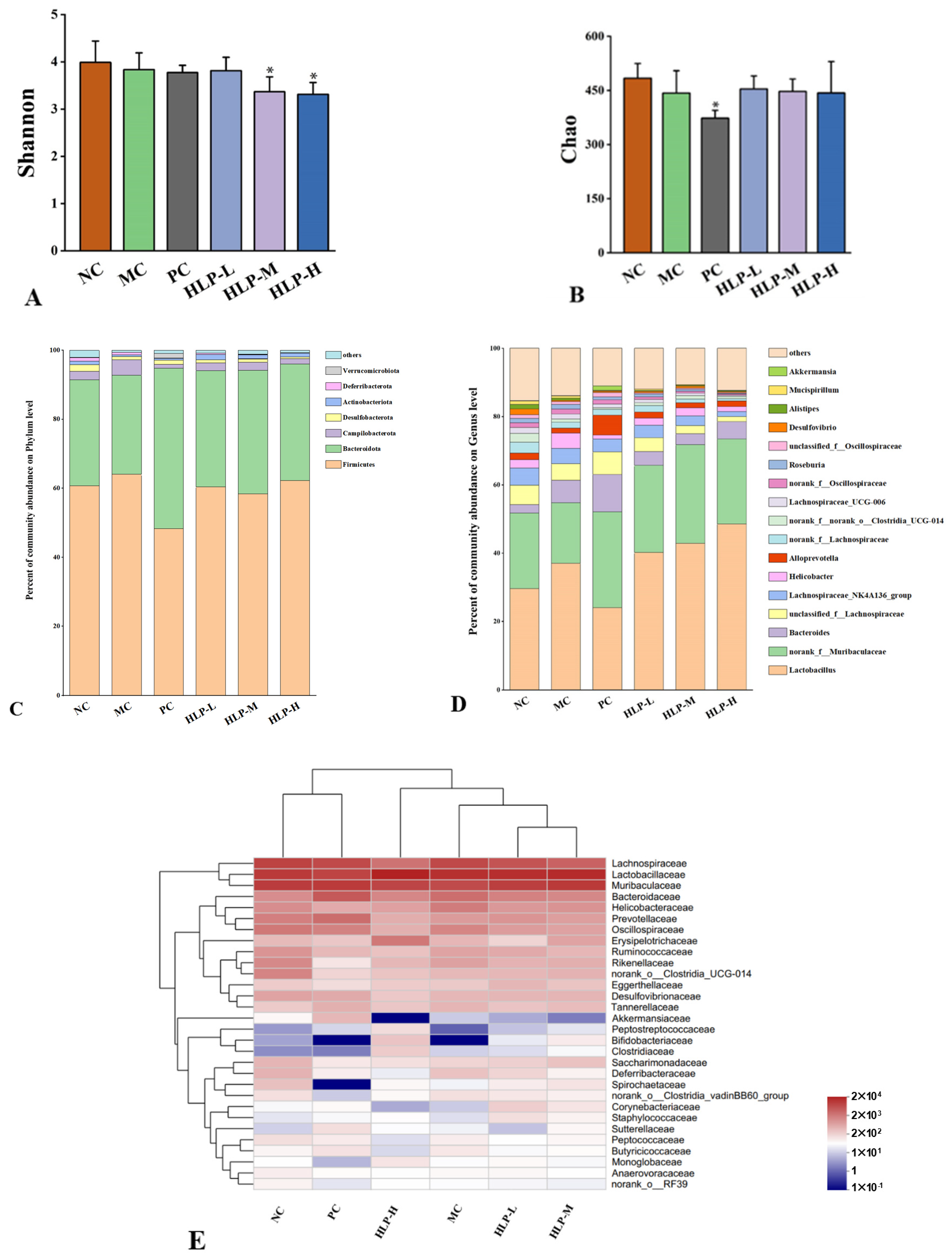
2.6. Effects of HLP on the Diversity and Community Structure of Gut Microbiota in Constipated Mice
3. Materials and Methods
3.1. Material and Reagents
3.2. Extraction and Purification of HLP
3.3. Animals and Experimental Design
3.4. Analyses of Constipation-Related Parameters
3.5. Histopathology and Immunohistochemistry
3.6. Enzyme-Linked Immunosorbent Assay (ELISA)
3.7. RNA Extraction and Real-Time qPCR (RT-qPCR)
3.8. Quantitative Assay of Short-Chain Fatty Acids (SCFAs)
3.9. Analysis of Microbial Diversity
3.10. Statistical Analysis
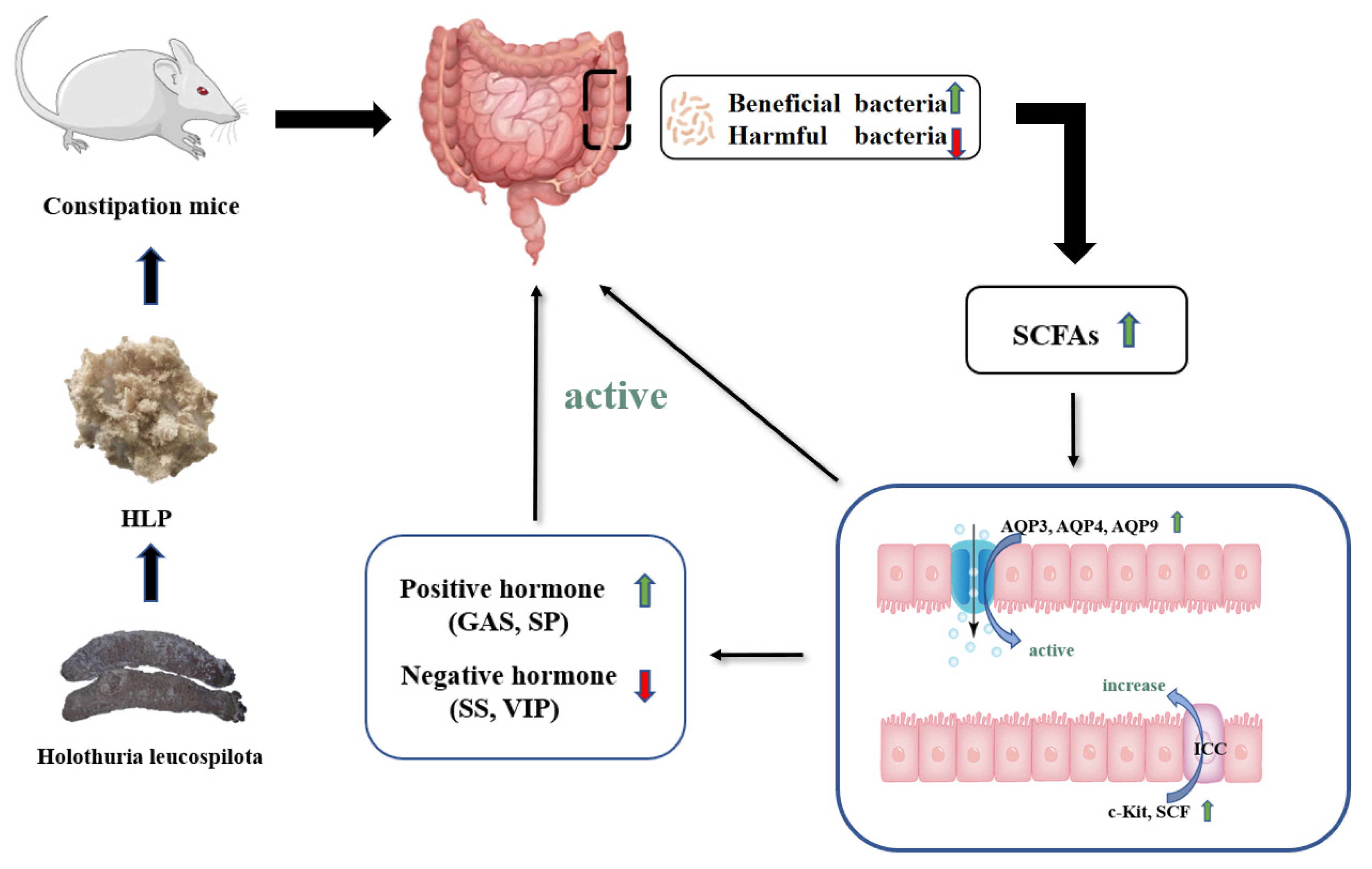
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bharucha, A.E.; Lacy, B.E. Mechanisms, Evaluation, and Management of Chronic Constipation. Gastroenterology 2020, 158, 1232–1249.e1233. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.S.; Rattanakovit, K.; Patcharatrakul, T. Diagnosis and management of chronic constipation in adults. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 295–305. [Google Scholar] [CrossRef]
- Sumida, K.; Molnar, M.Z.; Potukuchi, P.K.; Thomas, F.; Lu, J.L.; Yamagata, K.; Kalantar-Zadeh, K.; Kovesdy, C.P. Constipation and risk of death and cardiovascular events. Atherosclerosis 2019, 281, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Wald, A. Update on the Management of Constipation. JAMA 2019, 322, 2239–2240. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Natividad, J.M.; Verdu, E.F. Modulation of intestinal barrier by intestinal microbiota: Pathological and therapeutic implications. Pharmacol. Res. 2013, 69, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Icaza-Chávez, M.E. Gut microbiota in health and disease. Rev. Gastroenterol. Méx. Engl. Ed. 2013, 78, 240–248. [Google Scholar] [CrossRef]
- Ohkusa, T.; Koido, S.; Nishikawa, Y.; Sato, N. Gut Microbiota and Chronic Constipation: A Review and Update. Front. Med. 2019, 6, 19. [Google Scholar] [CrossRef]
- Heymen, S.; Jones, K.R.; Scarlett, Y.; Whitehead, W.E. Biofeedback treatment of constipation. Dis. Colon Rectum 2003, 46, 1208–1217. [Google Scholar] [CrossRef]
- Hsieh, C. Treatment of constipation in older adults. Am. Fam. Physician 2005, 72, 2277–2284. [Google Scholar] [CrossRef]
- Knowles, C.H.; Dinning, P.G.; Pescatori, M.; Rintala, R.; Rosen, H. Surgical management of constipation. Neurogastroenterol. Motil. 2009, 21 (Suppl. S2), 62–71. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Chen, F.; Wang, Z.; Cao, J.; Li, C. Effect and mechanism of functional compound fruit drink on gut microbiota in constipation mice. Food Chem. 2022, 401, 134210. [Google Scholar] [CrossRef] [PubMed]
- Khotimchenko, Y. Pharmacological potential of sea cucumbers. Int. J. Mol. Sci. 2018, 19, 1342. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zhu, K.; Zhao, Q.; Liu, Q.; Cao, J.; Xia, G.; Liu, Z.; Li, C. Holothuria leucospilota polysaccharides alleviate liver injury via AMPK and NF-κB signaling pathways in type 2 diabetic rats. J. Funct. Foods 2021, 85, 104657. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, Q.; Zhao, F.; Cao, J.; Shen, X.; Li, C. Holothuria Leucospilota Polysaccharides Ameliorate Hyperlipidemia in High-Fat Diet-Induced Rats via Short-Chain Fatty Acids Production and Lipid Metabolism Regulation. Int. J. Mol. Sci. 2019, 20, 4738. [Google Scholar] [CrossRef]
- Wang, W.; Yuan, Y.; Cao, J.; Shen, X.; Li, C. Beneficial Effects of Holothuria leucospilota Polysaccharides on Fermentability In Vivo and In Vitro. Foods 2021, 10, 1884. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Liu, Q.; Cao, J.; Xu, Y.; Pei, Z.; Fan, H.; Yuan, Y.; Shen, X.; Li, C. A sea cucumber (Holothuria leucospilota) polysaccharide improves the gut microbiome to alleviate the symptoms of type 2 diabetes mellitus in Goto-Kakizaki rats. Food Chem. Toxicol. 2020, 135, 110886. [Google Scholar] [CrossRef]
- Zhao, F.; Ma, T.; Zhang, X.; Zhao, Q.; Zhu, K.; Cao, J.; Liu, Z.; Shen, X.; Li, C. Holothuria Leucospilota Polysaccharides Improve Immunity and the Gut Microbiota in Cyclophosphamide-Treated Immunosuppressed Mice. Mol. Nutr. Food Res. 2022, 2022, 2200317. [Google Scholar] [CrossRef]
- Lu, S.; Yuan, Y.; Chen, F.; Zheng, Y.; Li, C.; Cao, J.; Xia, G.; Liu, Z.; Shen, X.; He, Y. Holothuria Leucospilota polysaccharides alleviate hyperlipidemia via alteration of lipid metabolism and inflammation-related gene expression. J. Food Biochem. 2022, 46, e14392. [Google Scholar] [CrossRef]
- Jiang, H.; Dong, J.; Jiang, S.; Liang, Q.; Zhang, Y.; Liu, Z.; Ma, C.; Wang, J.; Kang, W. Effect of Durio zibethinus rind polysaccharide on functional constipation and intestinal microbiota in rats. Food Res. Int. 2020, 136, 109316. [Google Scholar] [CrossRef]
- Wang, J.; Liang, Q.; Zhao, Q.; Tang, Q.; Ahmed, A.F.; Zhang, Y.; Kang, W. The effect of microbial composition and proteomic on improvement of functional constipation by Chrysanthemum morifolium polysaccharide. Food Chem. Toxicol. 2021, 153, 112305. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, S.; Yan, Q.; Li, Y.; Jiang, Z. Effect of Konjac mannan oligosaccharides on diphenoxylate-induced constipation in mice. J. Funct. Foods 2019, 57, 399–407. [Google Scholar] [CrossRef]
- Eor, J.Y.; Tan, P.L.; Lim, S.M.; Choi, D.H.; Yoon, S.M.; Yang, S.Y.; Kim, S.H. Laxative effect of probiotic chocolate on loperamide-induced constipation in rats. Food Res. Int. 2019, 116, 1173–1182. [Google Scholar] [CrossRef]
- Yang, Z.; Ye, S.; Xu, Z.; Su, H.; Tian, X.; Han, B.; Shen, B.; Liao, Q.; Xie, Z.; Hong, Y. Dietary synbiotic ameliorates constipation through the modulation of gut microbiota and its metabolic function. Food Res. Int. 2021, 147, 110569. [Google Scholar] [CrossRef]
- Krausova, A.; Buresova, P.; Sarnova, L.; Oyman-Eyrilmez, G.; Skarda, J.; Wohl, P.; Bajer, L.; Sticova, E.; Bartonova, L.; Pacha, J. Plectin ensures intestinal epithelial integrity and protects colon against colitis. Mucosal Immunol. 2021, 14, 691–702. [Google Scholar] [CrossRef]
- Hu, T.G.; Wen, P.; Fu, H.Z.; Lin, G.Y.; Liao, S.T.; Zou, Y.X. Protective effect of mulberry (Morus atropurpurea) fruit against diphenoxylate-induced constipation in mice through the modulation of gut microbiota. Food Funct. 2019, 10, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.Y.; Kim, S.; Hong, H.S. Substance-P Ameliorates Dextran Sodium Sulfate-Induced Intestinal Damage by Preserving Tissue Barrier Function. Tissue Eng. Regen. Med. 2018, 15, 63–73. [Google Scholar] [CrossRef]
- Hellmich, M.R.; Rui, X.L.; Hellmich, H.L.; Fleming, R.Y.; Evers, B.M.; Townsend, C.M., Jr. Human colorectal cancers express a constitutively active cholecystokinin-B/gastrin receptor that stimulates cell growth. J. Biol. Chem. 2000, 275, 32122–32128. [Google Scholar] [CrossRef]
- Vinolo, M.A.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhong, D.; Sun, R.; Zhang, Y.; Pegg, R.B.; Zhong, G. Prevention of loperamide induced constipation in mice by KGM and the mechanisms of different gastrointestinal tract microbiota regulation. Carbohydr. Polym. 2021, 256, 117418. [Google Scholar] [CrossRef]
- Silberstein, C.; Kierbel, A.; Amodeo, G.; Zotta, E.; Bigi, F.; Berkowski, D.; Ibarra, C. Functional characterization and localization of AQP3 in the human colon. Braz. J. Med. Biol. Res. 1999, 32, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Burns, A.J.; Herbert, T.M.; Ward, S.M.; Sanders, K.M. Interstitial cells of Cajal in the guinea-pig gastrointestinal tract as revealed by c-Kit immunohistochemistry. Cell Tissue Res. 1997, 290, 11–20. [Google Scholar] [CrossRef]
- Takaki, M. Gut pacemaker cells: The interstitial cells of Cajal (ICC). J. Smooth Muscle Res. 2003, 39, 137–161. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Watabe, K.; Nakahara, M.; Ogiyama, H.; Kiyohara, T.; Tsutsui, S.; Tamura, S.; Shinomura, Y.; Hayashi, N. Disturbed gastrointestinal motility and decreased interstitial cells of Cajal in diabetic db/db mice. J. Gastroenterol. Hepatol. 2008, 23, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Huang, Y.; Tang, H.; Tu, X.; He, J.; Wang, T.; Zhang, Q.; Xiong, F.; Li, D.; Qiu, Z. Role of stem cell growth factor/c-Kit in the pathogenesis of irritable bowel syndrome. Exp. Ther. Med. 2017, 13, 1187–1193. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, M.; Ouyang, D.; Tong, H.; Wu, M.; Su, L. Research Progress on the Protective Effect of Brown Algae-Derived Polysaccharides on Metabolic Diseases and Intestinal Barrier Injury. Int. J. Mol. Sci. 2022, 23, 10784. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A.; Strukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Zhao, L.; Huang, Y.; Lu, L.; Yang, W.; Huang, T.; Lin, Z.; Lin, C.; Kwan, H.; Wong, H.L.X.; Chen, Y.; et al. Saturated long-chain fatty acid-producing bacteria contribute to enhanced colonic motility in rats. Microbiome 2018, 6, 107. [Google Scholar] [CrossRef]
- Sun, Z.; Yu, Z.; Wang, B. Perilla frutescens Leaf Alters the Rumen Microbial Community of Lactating Dairy Cows. Microorganisms 2019, 7, 562. [Google Scholar] [CrossRef]
- Luo, Y.; Fang, Q.; Lai, Y.; Niu, H.; Wang, R.; Song, C. High-throughput sequencing technology reveals polysaccharides from Angelica dahurica that affect gut microbiota in mice. Biotechnol. Biotechnol. Equip. 2021, 35, 1934–1940. [Google Scholar] [CrossRef]
- Sibai, M.; Altuntas, E.; Yildirim, B.; Ozturk, G.; Yildirim, S.; Demircan, T. Microbiome and Longevity: High Abundance of Longevity-Linked Muribaculaceae in the Gut of the Long-Living Rodent Spalax leucodon. OMICS 2020, 24, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Bohr, U.R.; Primus, A.; Zagoura, A.; Glasbrenner, B.; Wex, T.; Malfertheiner, P. A group-specific PCR assay for the detection of Helicobacteraceae in human gut. Helicobacter 2002, 7, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Li, C.; Zheng, Q.; Wu, J.; Zhu, K.; Shen, X.; Cao, J. Effect of simulated gastrointestinal digestion in vitro on the antioxidant activity, molecular weight and microstructure of polysaccharides from a tropical sea cucumber (Holothuria leucospilota). Food Hydrocoll. 2019, 89, 735–741. [Google Scholar] [CrossRef]
- Li, C.; Nie, S.-P.; Zhu, K.-X.; Xiong, T.; Li, C.; Gong, J.; Xie, M.-Y. Effect of Lactobacillus plantarum NCU116 on loperamide-induced constipation in mice. Int. J. Food Sci. Nutr. 2015, 66, 533–538. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Group | |||||
|---|---|---|---|---|---|---|
| NC | MC | PC | HLP-L | HLP-M | HLP-H | |
| AQP3 | 1.20 ± 0.35 | 0.35 ± 0.13 # | 3.19 ± 0.95 ** | 0.85 ± 0.33 | 1.15 ± 0.38 * | 0.54 ± 0.26 |
| AQP4 | 1.19 ± 0.36 | 0.32 ± 0.15 # | 1.62 ± 0.80 * | 0.75 ± 0.11 * | 1.38 ± 1.06 | 1.12 ± 0.69 |
| AQP9 | 0.44 ± 0.00 | 0.16 ± 0.09 # | 1.40 ± 0.42 * | 0.69 ± 0.11 * | 0.57 ± 0.11 * | 0.43 ± 0.00 * |
| SCF | 1.03 ± 0.14 | 0.89 ± 0.05 # | 2.32 ± 1.55 * | 0.92 ± 0.40 * | 4.14 ± 0.58 | 2.06 ± 0.20 |
| c-Kit | 1.02 ± 0.50 | 0.25 ± 0.05 # | 1.36 ± 0.40 * | 1.03 ± 0.10 | 1.12 ± 0.07 *** | 1.40 ± 0.38 * |
| Gene Primer | Primer Sequence |
|---|---|
| AQP3 | Forward 5′-GCTGTCACCCTTGGCATCTTGG-3′ Reverse 5′-AGGAAGCACATTGCGAAGGTCAC-3′ |
| AQP4 | Forward 5′-CAGCATCGCTAAGTCCGTCTTCTAC-3′ Reverse 5′-ACCGTGGTGACTCCCAATCCTC-3′ |
| AQP9 | Forward 5′-GCTGTCCTGGGAGGTCTCATCTATG-3′ Reverse 5′-GCTGGTTCTGCCTTCACTTCTGG-3′ |
| SCF | Forward 5′-TGCGGGAATCCTGTGACTGATAATG-3′ Reverse 5′-CCGGCGACATAGTTGAGGGTTATC-3′ |
| c-Kit | Forward 5′-GATCTGCTCTGCGTCCTGTTGG-3′ Reverse 5′-AACTCTGATTGTGCTGGATGGATGG-3′ |
| GADPH | Forward 5′-GGTTGTCTCCTGCGACTTCA-3′ Reverse 5′-TGGTCCAGGGTTTCTTACTCC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Shi, Y.; Zeng, S.; Zheng, Y.; Wang, H.; Liao, H.; Song, J.; Zhang, X.; Cao, J.; Li, C. Polysaccharides from Holothuria leucospilota Relieve Loperamide-Induced Constipation Symptoms in Mice. Int. J. Mol. Sci. 2023, 24, 2553. https://doi.org/10.3390/ijms24032553
Wang Z, Shi Y, Zeng S, Zheng Y, Wang H, Liao H, Song J, Zhang X, Cao J, Li C. Polysaccharides from Holothuria leucospilota Relieve Loperamide-Induced Constipation Symptoms in Mice. International Journal of Molecular Sciences. 2023; 24(3):2553. https://doi.org/10.3390/ijms24032553
Chicago/Turabian StyleWang, Ziqi, Yali Shi, Shiyu Zeng, Yuanping Zheng, Huaijie Wang, Haihui Liao, Jie Song, Xinyue Zhang, Jun Cao, and Chuan Li. 2023. "Polysaccharides from Holothuria leucospilota Relieve Loperamide-Induced Constipation Symptoms in Mice" International Journal of Molecular Sciences 24, no. 3: 2553. https://doi.org/10.3390/ijms24032553
APA StyleWang, Z., Shi, Y., Zeng, S., Zheng, Y., Wang, H., Liao, H., Song, J., Zhang, X., Cao, J., & Li, C. (2023). Polysaccharides from Holothuria leucospilota Relieve Loperamide-Induced Constipation Symptoms in Mice. International Journal of Molecular Sciences, 24(3), 2553. https://doi.org/10.3390/ijms24032553