
MRL/MpJ Mice Resist to Age-Related and Long-Term Ovariectomy-Induced Bone Loss: Implications for Bone Regeneration and Repair

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
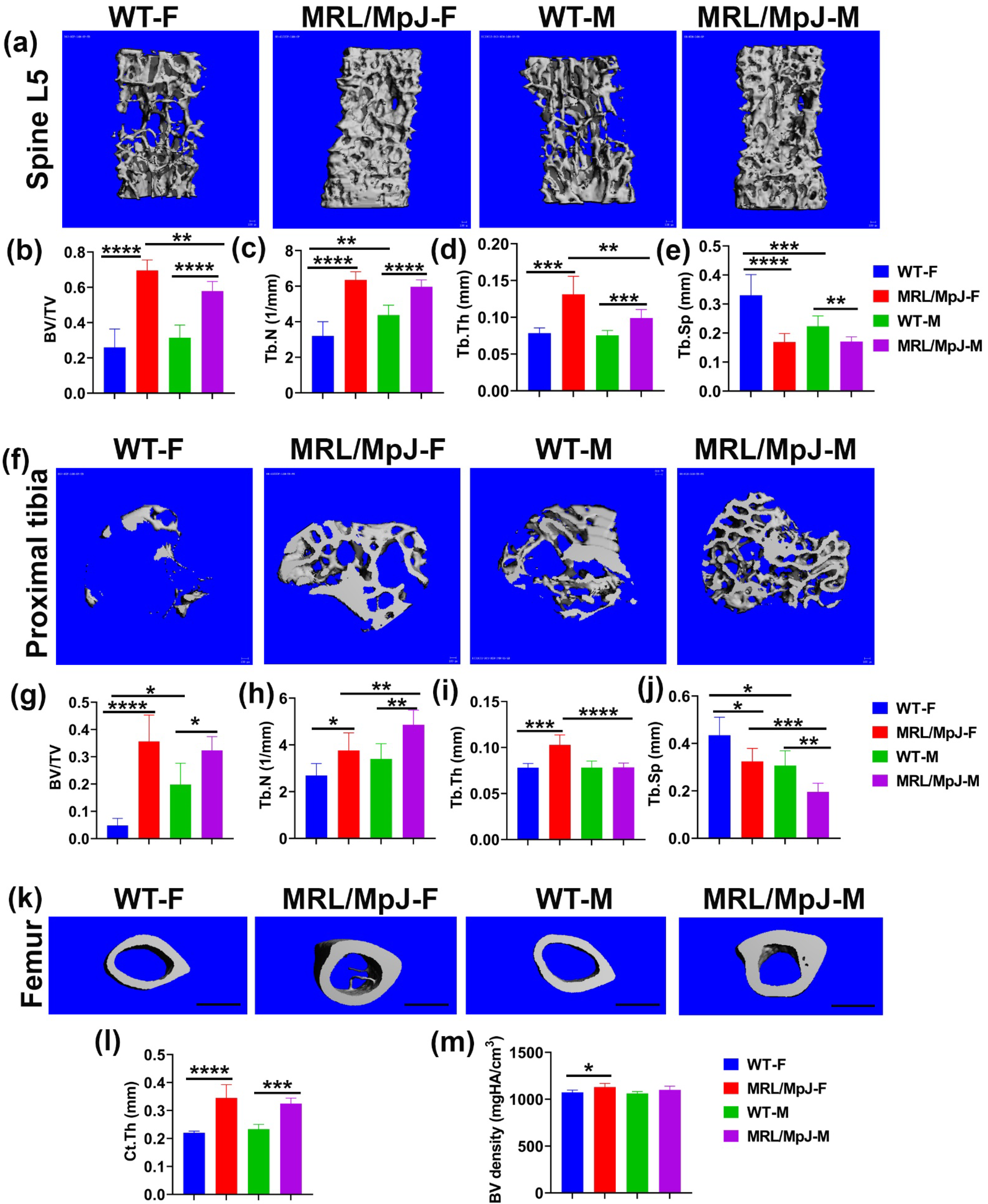
2.1. MRL/MpJ Mice Maintained Better Bone Microarchitecture at 14 Months Old (14 M) Compared to WT Mice
2.2. MRL/MpJ Mice Demonstrated Significantly Greater Bone Matrix Architecture Compared to WT Mice at 14 M
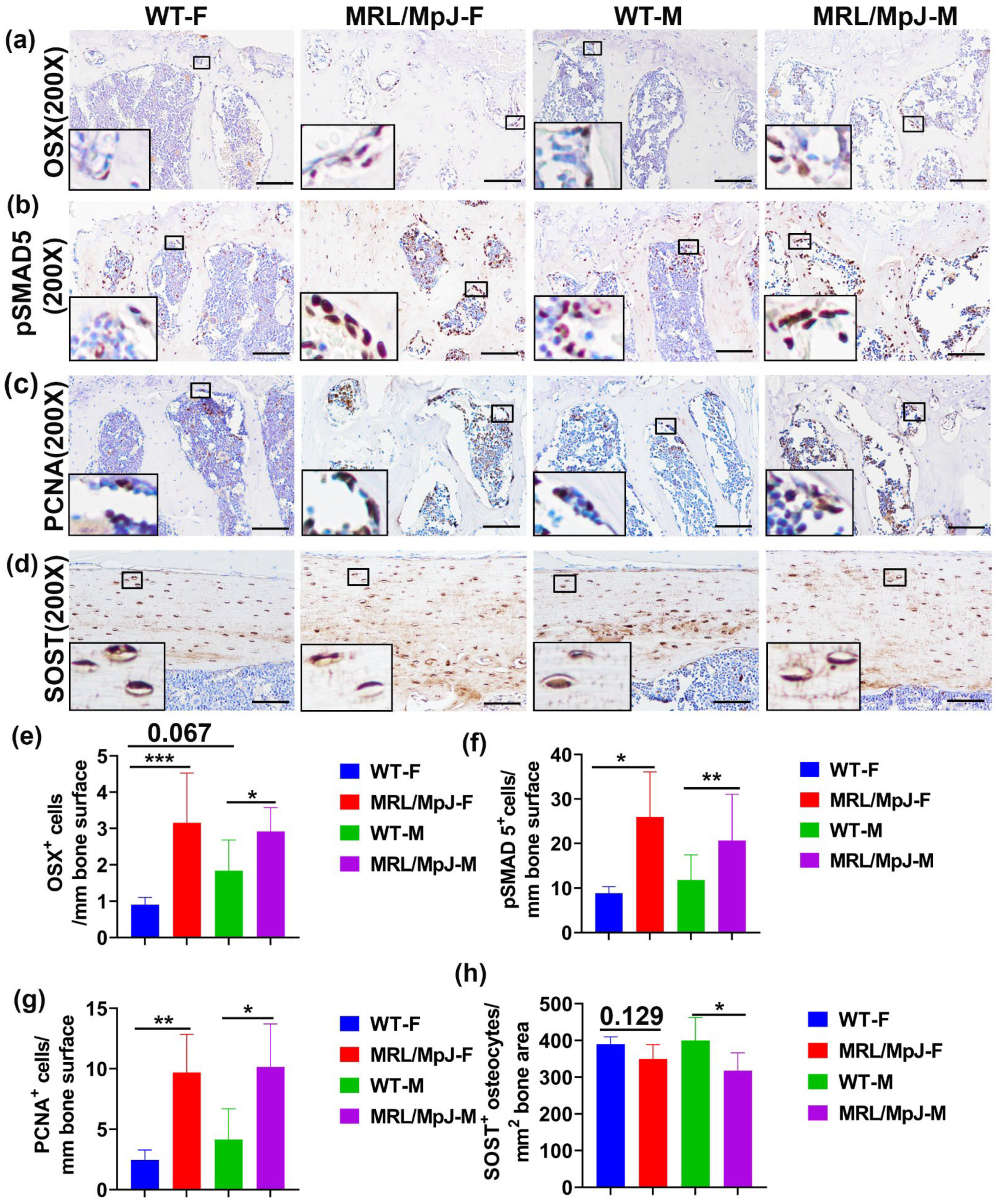
2.3. High Number of Osterix (OSX)+ Osteogenic Progenitor Cells with the Activation of pSMAD5, and Increased Osteoblasts Proliferation, Is Observed in the 14 M MRL/MpJ Mice Compared to WT Mice
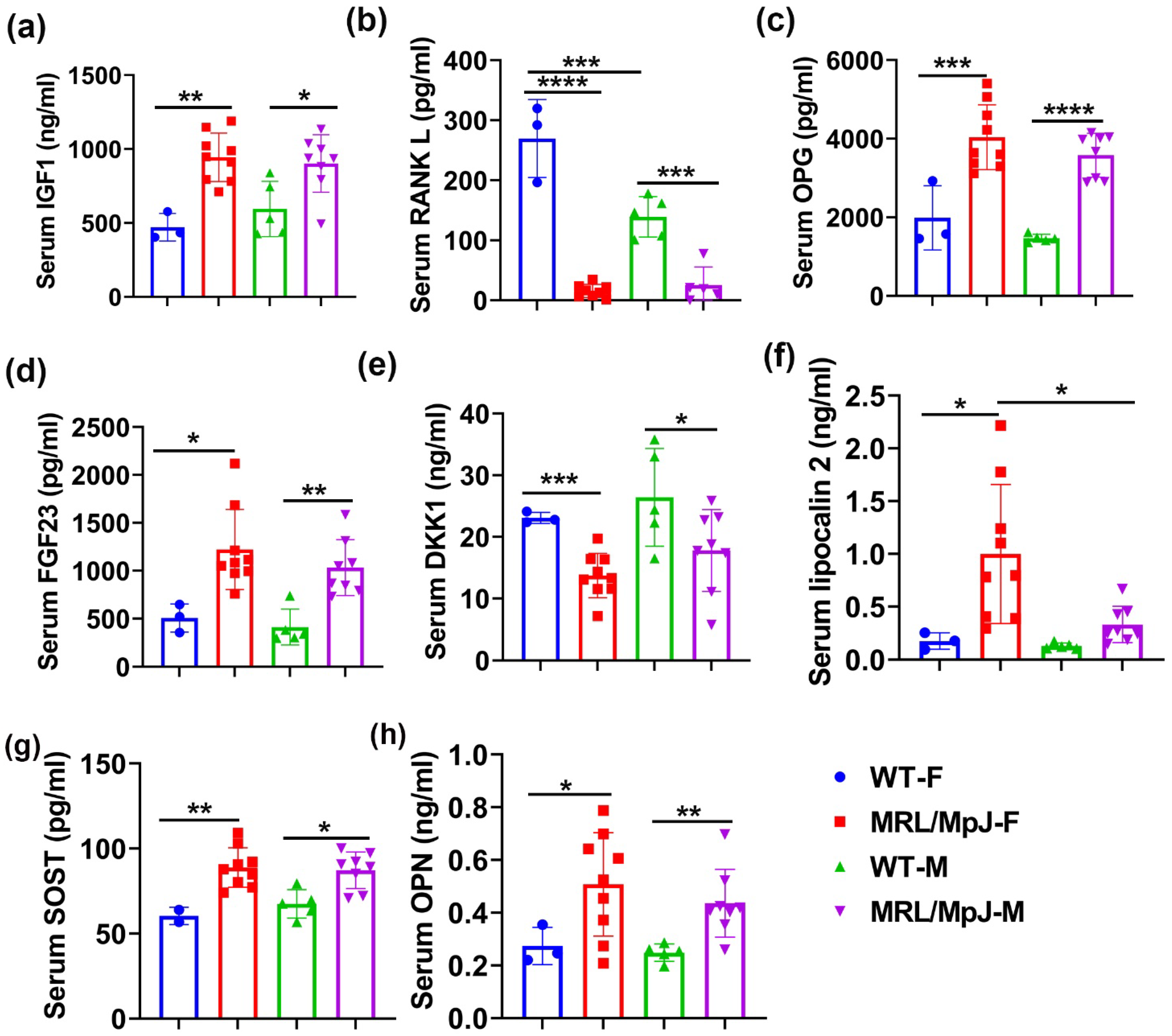
2.4. Serum Bone Marker Quantification in MRL/MpJ and WT Mice at 14 M
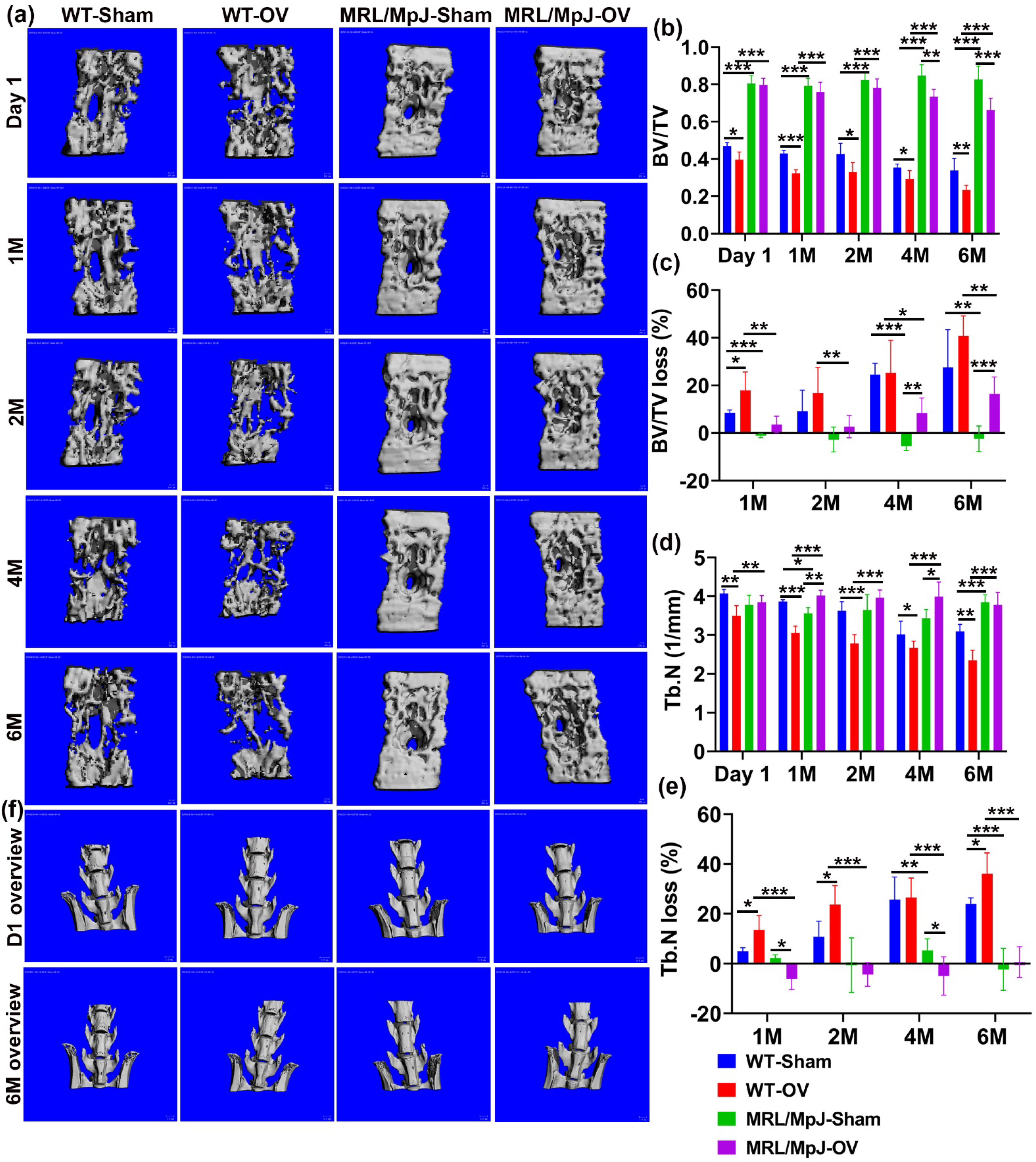
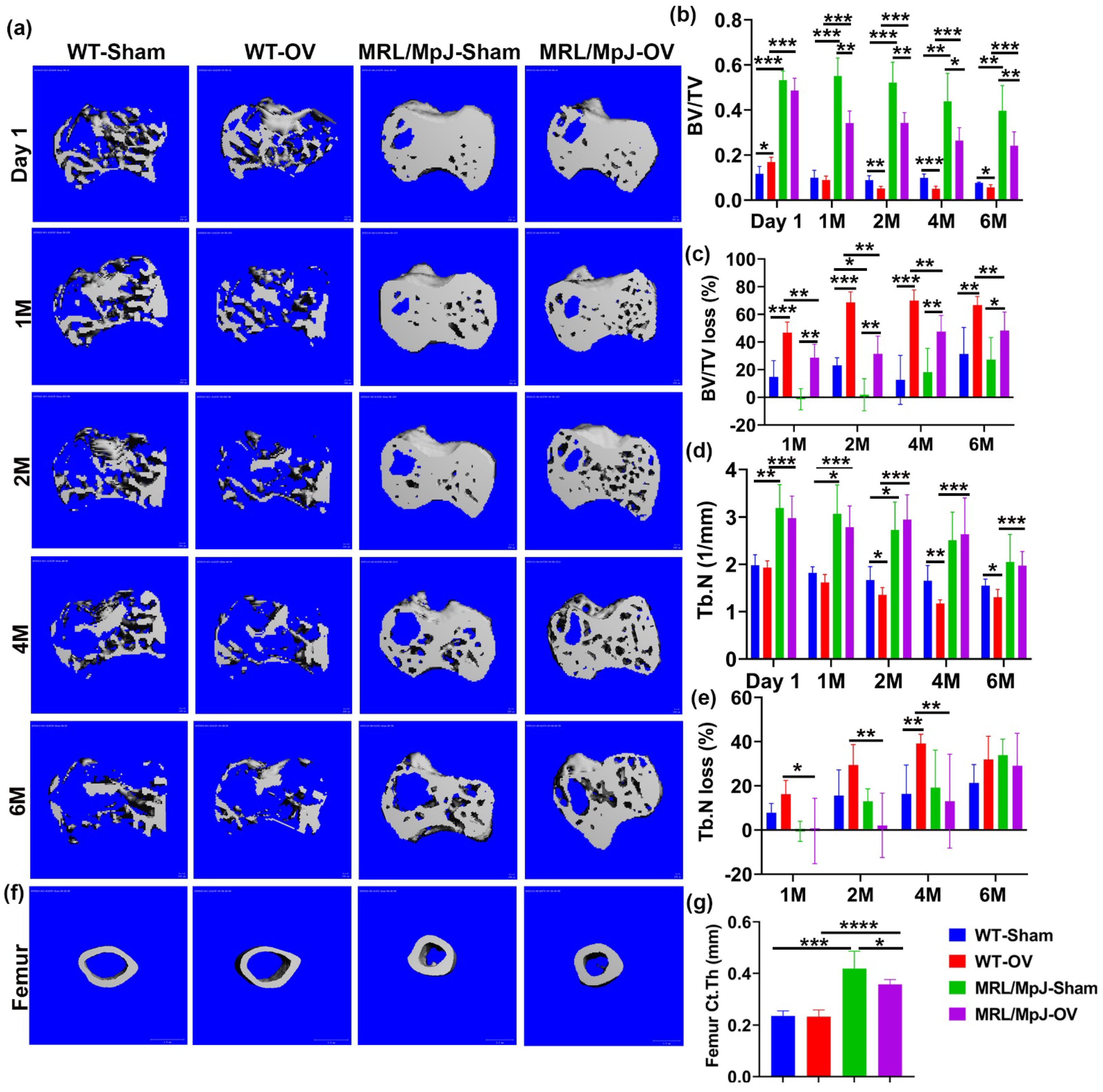
2.5. MRL/MpJ Mice Demonstrated Less Severe Bone Loss after Long-Term Ovariectomy
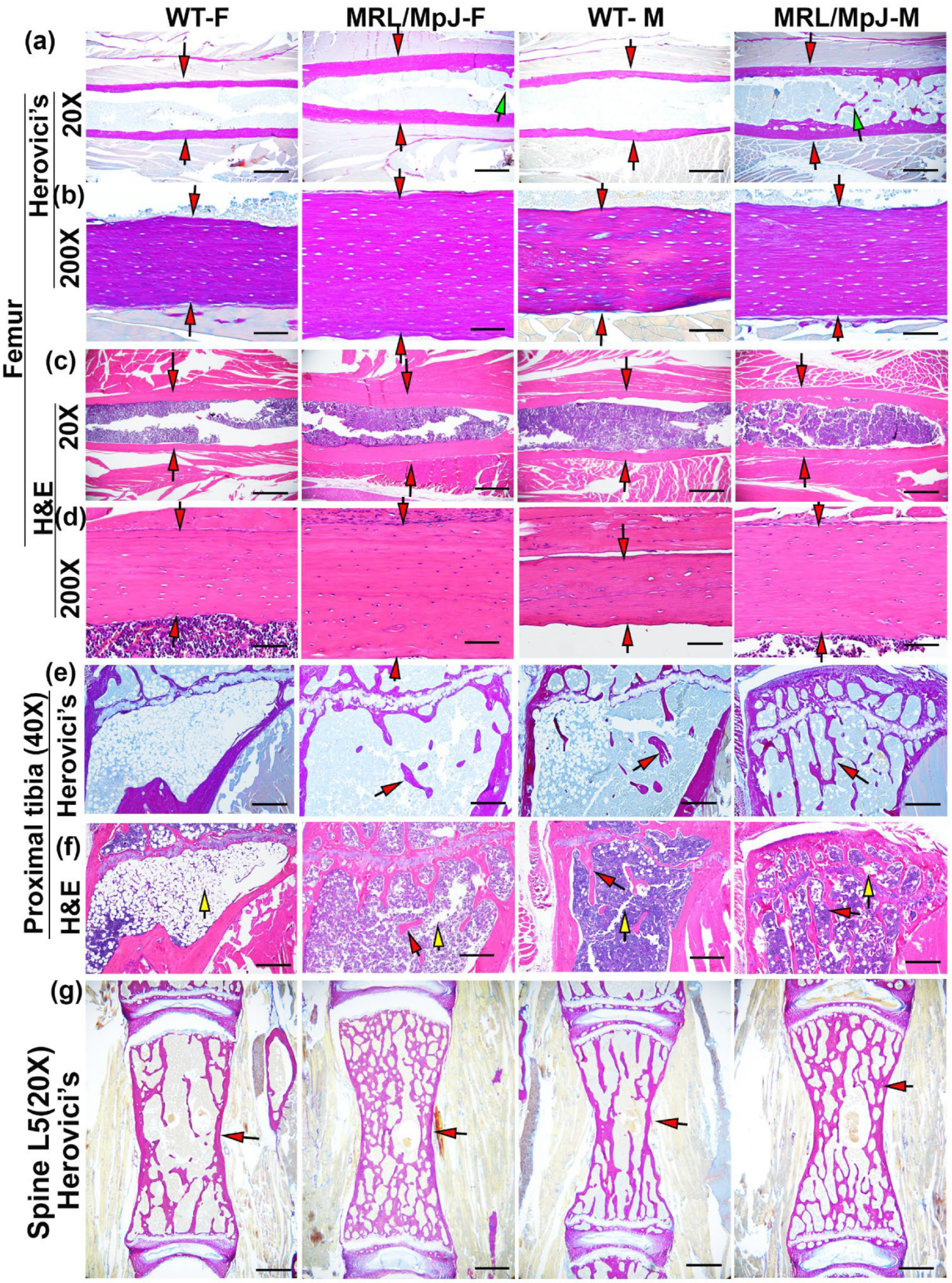
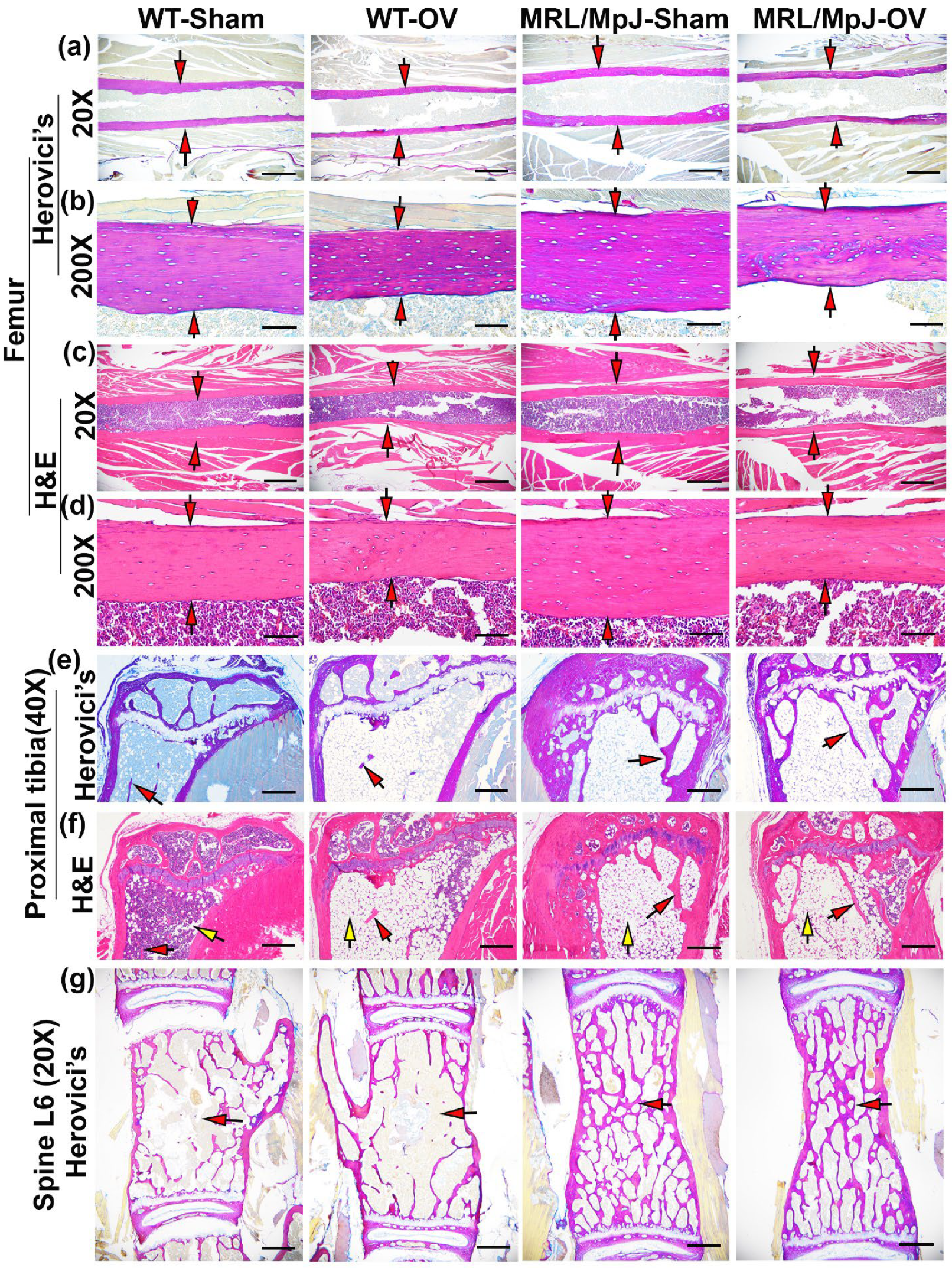
2.6. Histology of Bone Tissues 6 M Following Ovariectomy
2.7. MRL/MpJ Mice Maintained Higher OSX+ Cells, High pSMAD5+ and PCNA+ Cells on Bone Surface, and Lower TRAP+ Osteoclasts after Ovariectomy
2.8. Serum Protein ELISA and Multiplex Measurements after Ovariectomy
3. Discussion
4. Materials and Methods
4.1. Animal Use Ethics
4.2. Characterization of the Bone Microarchitecture of MRL/MpJ and WT Mice at 14 M
4.3. Micro-CT Scanning and Analysis of 14 M Mice Bone Specimens
4.4. Ovariectomy Surgeries
4.5. Micro-CT Scanning and Analysis of Mice Undergoing Ovariectomy and Sham Surgery
4.6. Histology
4.7. Immunohistochemistry
4.8. Serum Bone-Related Protein Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, D.; Nakamura, M.; Miclau, T.; Marcucio, R. Effects of Aging on Fracture Healing. Curr. Osteoporos. Rep. 2017, 15, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Kostenuik, P.; Mirza, F.M. Fracture healing physiology and the quest for therapies for delayed healing and nonunion. J. Orthop. Res. 2017, 35, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Burge, R.; Dawson-Hughes, B.; Solomon, D.H.; Wong, J.B.; King, A.; Tosteson, A. Incidence and economic burden of osteoporosis-related fractures in the United States, 2005–2025. J. Bone Miner. Res. 2007, 22, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Machida, M.; Weinstein, S.L.; Yamada, T.; Kimura, J.; Toriyama, S. Dissociation of muscle action potentials and spinal somatosensory evoked potentials after ischemic damage of spinal cord. Spine 1988, 13, 1119–1124. [Google Scholar] [CrossRef]
- Clark, L.D.; Clark, R.K.; Heber-Katz, E. A new murine model for mammalian wound repair and regeneration. Clin. Immunol. Immunopathol. 1998, 88, 35–45. [Google Scholar] [CrossRef]
- Buckley, G.; Metcalfe, A.D.; Ferguson, M.W. Peripheral nerve regeneration in the MRL/MpJ ear wound model. J. Anat. 2011, 218, 163–172. [Google Scholar] [CrossRef]
- Thuret, S.; Thallmair, M.; Horky, L.L.; Gage, F.H. Enhanced functional recovery in MRL/MpJ mice after spinal cord dorsal hemisection. PLoS ONE 2012, 7, e30904. [Google Scholar] [CrossRef]
- Rai, M.F.; Sandell, L.J. Regeneration of articular cartilage in healer and non-healer mice. Matrix Biol. 2014, 39, 50–55. [Google Scholar] [CrossRef]
- Zins, S.R.; Amare, M.F.; Anam, K.; Elster, E.A.; Davis, T.A. Wound trauma mediated inflammatory signaling attenuates a tissue regenerative response in MRL/MpJ mice. J. Inflamm. 2010, 7, 25. [Google Scholar] [CrossRef]
- Buckley, G.; Wong, J.; Metcalfe, A.D.; Ferguson, M.W. Denervation affects regenerative responses in MRL/MpJ and repair in C57BL/6 ear wounds. J. Anat. 2012, 220, 3–12. [Google Scholar] [CrossRef]
- Mak, J.; Leonard, C.; Foniok, T.; Rushforth, D.; Dunn, J.F.; Krawetz, R. Evaluating endogenous repair of focal cartilage defects in C57BL/6 and MRL/MpJ mice using 9.4T magnetic resonance imaging: A pilot study. Magn. Reason. Imaging 2015, 33, 690–694. [Google Scholar] [CrossRef]
- Fitzgerald, J. Enhanced cartilage repair in ‘healer’ mice-New leads in the search for better clinical options for cartilage repair. Semin. Cell Dev. Biol. 2017, 62, 78–85. [Google Scholar] [CrossRef]
- Heydemann, A. The super super-healing MRL mouse strain. Front. Biol. 2012, 7, 522–538. [Google Scholar] [CrossRef]
- Paredes, J.; Marvin, J.C.; Vaughn, B.; Andarawis-Puri, N. Innate tissue properties drive improved tendon healing in MRL/MpJ and harness cues that enhance behavior of canonical healing cells. FASEB J. 2020, 34, 8341–8356. [Google Scholar] [CrossRef]
- Deng, Z.; Gao, X.; Sun, X.; Amra, S.; Lu, A.; Cui, Y.; Eltzschig, H.K.; Lei, G.; Huard, J. Characterization of articular cartilage homeostasis and the mechanism of superior cartilage regeneration of MRL/MpJ mice. FASEB J. 2019, 33, 8809–8821. [Google Scholar] [CrossRef]
- Ward, B.D.; Furman, B.D.; Huebner, J.L.; Kraus, V.B.; Guilak, F.; Olson, S.A. Absence of posttraumatic arthritis following intraarticular fracture in the MRL/MpJ mouse. Arthritis. Rheum. 2008, 58, 744–753. [Google Scholar] [CrossRef]
- Ueno, M.; Lyons, B.L.; Burzenski, L.M.; Gott, B.; Shaffer, D.J.; Roopenian, D.C.; Shultz, L.D. Accelerated wound healing of alkali-burned corneas in MRL mice is associated with a reduced inflammatory signature. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4097–4106. [Google Scholar] [CrossRef]
- Naseem, R.H.; Meeson, A.P.; Michael Dimaio, J.; White, M.D.; Kallhoff, J.; Humphries, C.; Goetsch, S.C.; De Windt, L.J.; Williams, M.A.; Garry, M.G.; et al. Reparative myocardial mechanisms in adult C57BL/6 and MRL mice following injury. Physiol. Genom. 2007, 30, 44–52. [Google Scholar] [CrossRef]
- Leferovich, J.M.; Bedelbaeva, K.; Samulewicz, S.; Zhang, X.M.; Zwas, D.; Lankford, E.B.; Heber-Katz, E. Heart regeneration in adult MRL mice. Proc. Natl. Acad. Sci. USA 2001, 98, 9830–9835. [Google Scholar] [CrossRef]
- Velasco, C.; Dunn, C.; Sturdy, C.; Izda, V.; Martin, J.; Rivas, A.; McNaughton, J.; Jeffries, M.A. Ear wound healing in MRL/MpJ mice is associated with gut microbiome composition and is transferable to non-healer mice via microbiome transplantation. PLoS ONE 2021, 16, e0248322. [Google Scholar] [CrossRef]
- Wang, R.; Jiang, W.; Zhang, L.; Xie, S.; Zhang, S.; Yuan, S.; Jin, Y.; Zhou, G. Intra-articular delivery of extracellular vesicles secreted by chondrogenic progenitor cells from MRL/MpJ superhealer mice enhances articular cartilage repair in a mouse injury model. Stem. Cell Res. Ther. 2020, 11, 93. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.; Sinha, K.; Pan, H.; Cui, Y.; Guo, P.; Lin, C.Y.; Yang, F.; Deng, Z.; Eltzschig, H.K.; Lu, A.; et al. Markers of Accelerated Skeletal Muscle Regenerative Response in Murphy Roths Large Mice: Characteristics of Muscle Progenitor Cells and Circulating Factors. Stem. Cells 2019, 37, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Sinha, K.M.; Tseng, C.; Guo, P.; Lu, A.; Pan, H.; Gao, X.; Andrews, R.; Eltzschig, H.; Huard, J. Hypoxia-inducible factor 1alpha (HIF-1alpha) is a major determinant in the enhanced function of muscle-derived progenitors from MRL/MpJ mice. FASEB J. 2019, 33, 8321–8334. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Gao, X.; Deng, Z.; Zhang, L.; McGilvray, K.; Gadomski, B.C.; Amra, S.; Bao, G.; Huard, J. High bone microarchitecture, strength, and resistance to bone loss in MRL/MpJ mice correlates with activation of different signaling pathways and systemic factors. FASEB J. 2020, 34, 789–806. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Hwang, M.P.; Wright, N.; Lu, A.; Ruzbarsky, J.J.; Huard, M.; Cheng, H.; Mullen, M.; Ravuri, S.; Wang, B.; et al. The use of heparin/polycation coacervate sustain release system to compare the bone regenerative potentials of 5 BMPs using a critical sized calvarial bone defect model. Biomaterials 2022, 288, 121708. [Google Scholar] [CrossRef]
- Kelman, Z. PCNA: Structure, functions and interactions. Oncogene 1997, 14, 629–640. [Google Scholar] [CrossRef]
- Lee, H.; Lee, K.; Lee, S.; Lee, J.; Jeong, W.T.; Lim, H.B.; Hyun, T.K.; Yi, S.J.; Kim, K. Ethyl Acetate Fraction of Aqueous Extract of Lentinula edodes Inhibits Osteoclastogenesis by Suppressing NFATc1 Expression. Int. J. Mol. Sci. 2020, 21, 1347. [Google Scholar] [CrossRef]
- Cordoba, A.; Manzanaro-Moreno, N.; Colom, C.; Ronold, H.J.; Lyngstadaas, S.P.; Monjo, M.; Ramis, J.M. Quercitrin Nanocoated Implant Surfaces Reduce Osteoclast Activity In Vitro and In Vivo. Int. J. Mol. Sci. 2018, 19, 3319. [Google Scholar] [CrossRef]
- Anderson, P.H.; Sawyer, R.K.; Moore, A.J.; May, B.K.; O’Loughlin, P.D.; Morris, H.A. Vitamin D depletion induces RANKL-mediated osteoclastogenesis and bone loss in a rodent model. J. Bone Miner. Res. 2008, 23, 1789–1797. [Google Scholar] [CrossRef]
- Chen, X.; Zhi, X.; Pan, P.; Cui, J.; Cao, L.; Weng, W.; Zhou, Q.; Wang, L.; Zhai, X.; Zhao, Q.; et al. Matrine prevents bone loss in ovariectomized mice by inhibiting RANKL-induced osteoclastogenesis. FASEB J. 2017, 31, 4855–4865. [Google Scholar] [CrossRef]
- Tarantino, U.; Celi, M.; Feola, M.; Liuni, F.M.; Resmini, G.; Iolascon, G. A new antiresorptive approach to the treatment of fragility fractures: Long-term efficacy and safety of denosumab. Aging Clin. Exp. Res. 2013, 25 (Suppl. 1), S65–S69. [Google Scholar] [CrossRef]
- Simic, P.; Kim, W.; Zhou, W.; Pierce, K.A.; Chang, W.; Sykes, D.B.; Aziz, N.B.; Elmariah, S.; Ngo, D.; Pajevic, P.D.; et al. Glycerol-3-phosphate is an FGF23 regulator derived from the injured kidney. J. Clin. Investig. 2020, 130, 1513–1526. [Google Scholar] [CrossRef]
- Kroon, T.; Bhadouria, N.; Niziolek, P.; Edwards, D., 3rd; Choi, R.; Clinkenbeard, E.L.; Robling, A.; Holguin, N. Suppression of Sost/Sclerostin and Dickkopf-1 Augment Intervertebral Disc Structure in Mice. J. Bone Miner. Res. 2022, 37, 1156–1169. [Google Scholar] [CrossRef]
- Lewiecki, E.M.; Chastek, B.; Sundquist, K.; Williams, S.A.; Weiss, R.J.; Wang, Y.; Fitzpatrick, L.A.; Curtis, J.R. Osteoporotic fracture trends in a population of US managed care enrollees from 2007 to 2017. Osteoporos. Int. 2020, 31, 1299–1304. [Google Scholar] [CrossRef]
- Yakar, S.; Rosen, C.J.; Beamer, W.G.; Ackert-Bicknell, C.L.; Wu, Y.; Liu, J.L.; Ooi, G.T.; Setser, J.; Frystyk, J.; Boisclair, Y.R.; et al. Circulating levels of IGF-1 directly regulate bone growth and density. J. Clin. Investig. 2002, 110, 771–781. [Google Scholar] [CrossRef]
- Xian, L.; Wu, X.; Pang, L.; Lou, M.; Rosen, C.J.; Qiu, T.; Crane, J.; Frassica, F.; Zhang, L.; Rodriguez, J.P.; et al. Matrix IGF-1 maintains bone mass by activation of mTOR in mesenchymal stem cells. Nat. Med. 2012, 18, 1095–1101. [Google Scholar] [CrossRef]
- Celiker, R.; Arslan, S. Comparison of serum insulin-like growth factor-1 and growth hormone levels in osteoporotic and non-osteoporotic postmenopausal women. Rheumatol. Int. 2000, 19, 205–208. [Google Scholar] [CrossRef]
- Garnero, P.; Sornay-Rendu, E.; Delmas, P.D. Low serum IGF-1 and occurrence of osteoporotic fractures in postmenopausal women. Lancet 2000, 355, 898–899. [Google Scholar] [CrossRef]
- Sun, L.; Su, J.; Wang, M. Changes of serum IGF-1 and ET-1 levels in patients with osteoporosis and its clinical significance. Pak. J. Med. Sci. 2019, 35, 691–695. [Google Scholar] [CrossRef]
- Paccou, J.; Dewailly, J.; Cortet, B. Reduced levels of serum IGF-1 is related to the presence of osteoporotic fractures in male idiopathic osteoporosis. Joint Bone Spine 2012, 79, 78–82. [Google Scholar] [CrossRef]
- Ohlsson, C.; Mellstrom, D.; Carlzon, D.; Orwoll, E.; Ljunggren, O.; Karlsson, M.K.; Vandenput, L. Older men with low serum IGF-1 have an increased risk of incident fractures: The MrOS Sweden study. J. Bone Miner. Res. 2011, 26, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.M.; Zhao, H.Y.; Ning, G.; Chen, Y.; Zhang, L.Z.; Sun, L.H.; Zhao, Y.J.; Xu, M.Y.; Chen, J.L. IGF-1 as an early marker for low bone mass or osteoporosis in premenopausal and postmenopausal women. J. Bone Miner. Metab. 2008, 26, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Ardawi, M.S.; Akhbar, D.H.; Alshaikh, A.; Ahmed, M.M.; Qari, M.H.; Rouzi, A.A.; Ali, A.Y.; Abdulrafee, A.A.; Saeda, M.Y. Increased serum sclerostin and decreased serum IGF-1 are associated with vertebral fractures among postmenopausal women with type-2 diabetes. Bone 2013, 56, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Goetz, T.G.; Nair, N.; Shiau, S.; Recker, R.R.; Lappe, J.M.; Dempster, D.W.; Zhou, H.; Zhao, B.; Guo, X.; Shen, W.; et al. In premenopausal women with idiopathic osteoporosis, lower bone formation rate is associated with higher body fat and higher IGF-1. Osteoporos. Int. 2022, 33, 659–672. [Google Scholar] [CrossRef]
- Zhang, W.B.; Ye, K.; Barzilai, N.; Milman, S. The antagonistic pleiotropy of insulin-like growth factor 1. Aging Cell 2021, 20, e13443. [Google Scholar] [CrossRef]
- Edwards, J.R.; Mundy, G.R. Advances in osteoclast biology: Old findings and new insights from mouse models. Nat. Rev. Rheumatol. 2011, 7, 235–243. [Google Scholar] [CrossRef]
- Reid, I.R.; Billington, E.O. Drug therapy for osteoporosis in older adults. Lancet 2022, 399, 1080–1092. [Google Scholar] [CrossRef]
- Miller, P.D. A review of the efficacy and safety of denosumab in postmenopausal women with osteoporosis. Ther. Adv. Musculoskelet. Dis. 2011, 3, 271–282. [Google Scholar] [CrossRef]
- Fassio, A.; Adami, G.; Benini, C.; Vantaggiato, E.; Saag, K.G.; Giollo, A.; Lippolis, I.; Viapiana, O.; Idolazzi, L.; Orsolini, G.; et al. Changes in Dkk-1, sclerostin, and RANKL serum levels following discontinuation of long-term denosumab treatment in postmenopausal women. Bone 2019, 123, 191–195. [Google Scholar] [CrossRef]
- Ahmed, S.F.; Fouda, N.; Abbas, A.A. Serum dickkopf-1 level in postmenopausal females: Correlation with bone mineral density and serum biochemical markers. J. Osteoporos. 2013, 2013, 460210. [Google Scholar] [CrossRef]
- Tian, J.; Xu, X.J.; Shen, L.; Yang, Y.P.; Zhu, R.; Shuai, B.; Zhu, X.W.; Li, C.G.; Ma, C.; Lv, L. Association of serum Dkk-1 levels with beta-catenin in patients with postmenopausal osteoporosis. J. Huazhong Univ. Sci. Technolog. Med. Sci. 2015, 35, 212–218. [Google Scholar] [CrossRef]
- Long, L.; Liu, Y.; Wang, S.; Zhao, Y.; Guo, J.; Yu, P.; Li, Z. Dickkopf-1 as potential biomarker to evaluate bone erosion in systemic lupus erythematosus. J. Clin. Immunol. 2010, 30, 669–675. [Google Scholar] [CrossRef]
- van Lierop, A.H.; Moester, M.J.; Hamdy, N.A.; Papapoulos, S.E. Serum Dickkopf 1 levels in sclerostin deficiency. J. Clin. Endocrinol. Metab. 2014, 99, E252–E256. [Google Scholar] [CrossRef]
- Wanby, P.; Nobin, R.; Von, S.P.; Brudin, L.; Carlsson, M. Serum levels of the bone turnover markers dickkopf-1, sclerostin, osteoprotegerin, osteopontin, osteocalcin and 25-hydroxyvitamin D in Swedish geriatric patients aged 75 years or older with a fresh hip fracture and in healthy controls. J. Endocrinol. Invest. 2016, 39, 855–863. [Google Scholar] [CrossRef]
- Lim, Y.; Kim, C.H.; Lee, S.Y.; Kim, H.; Ahn, S.H.; Lee, S.H.; Koh, J.M.; Rhee, Y.; Baek, K.H.; Min, Y.K.; et al. Decreased Plasma Levels of Sclerostin But Not Dickkopf-1 are Associated with an Increased Prevalence of Osteoporotic Fracture and Lower Bone Mineral Density in Postmenopausal Korean Women. Calcif. Tissue Int. 2016, 99, 350–359. [Google Scholar] [CrossRef]
- Chen, Y.; Aiken, A.; Saw, S.; Weiss, A.; Fang, H.; Khokha, R. TIMP Loss Activates Metalloproteinase-TNFalpha-DKK1 Axis To Compromise Wnt Signaling and Bone Mass. J. Bone Miner. Res. 2019, 34, 182–194. [Google Scholar] [CrossRef]
- Ueland, T.; Stilgren, L.; Bollerslev, J. Bone Matrix Levels of Dickkopf and Sclerostin are Positively Correlated with Bone Mass and Strength in Postmenopausal Osteoporosis. Int. J. Mol. Sci. 2019, 20, 2896. [Google Scholar] [CrossRef]
- Jo, S.; Yoon, S.; Lee, S.Y.; Kim, S.Y.; Park, H.; Han, J.; Choi, S.H.; Han, J.S.; Yang, J.H.; Kim, T.H. DKK1 Induced by 1,25D3 Is Required for the Mineralization of Osteoblasts. Cells 2020, 9, 236. [Google Scholar] [CrossRef]
- Colditz, J.; Thiele, S.; Baschant, U.; Garbe, A.I.; Niehrs, C.; Hofbauer, L.C.; Rauner, M. Osteogenic Dkk1 Mediates Glucocorticoid-Induced but Not Arthritis-Induced Bone Loss. J. Bone Miner. Res. 2019, 34, 1314–1323. [Google Scholar] [CrossRef]
- Lehmann, J.; Thiele, S.; Baschant, U.; Rachner, T.D.; Niehrs, C.; Hofbauer, L.C.; Rauner, M. Mice lacking DKK1 in T cells exhibit high bone mass and are protected from estrogen-deficiency-induced bone loss. iScience 2021, 24, 102224. [Google Scholar] [CrossRef]
- Peng, J.; Dong, Z.; Hui, Z.; Aifei, W.; Lianfu, D.; Youjia, X. Bone Sclerostin and Dickkopf-related protein-1 are positively correlated with bone mineral density, bone microarchitecture, and bone strength in postmenopausal osteoporosis. BMC Musculoskelet. Disord. 2021, 22, 480. [Google Scholar] [CrossRef] [PubMed]
- Li, C.W.; Liang, B.; Shi, X.L.; Wang, H. Opg/Rankl mRNA dynamic expression in the bone tissue of ovariectomized rats with osteoporosis. Genet Mol. Res. 2015, 14, 9215–9224. [Google Scholar] [CrossRef] [PubMed]
- Idris, A.I. Ovariectomy/orchidectomy in rodents. Methods Mol. Biol. 2012, 816, 545–551. [Google Scholar] [PubMed]
- Turner, N.J.; Pezzone, M.A.; Brown, B.N.; Badylak, S.F. Quantitative multispectral imaging of Herovici’s polychrome for the assessment of collagen content and tissue remodelling. J. Tissue Eng. Regen. Med. 2013, 7, 139–148. [Google Scholar] [CrossRef]
- Gao, X.; Tang, Y.; Amra, S.; Sun, X.; Cui, Y.; Cheng, H.; Wang, B.; Huard, J. Systemic investigation of bone and muscle abnormalities in dystrophin/utrophin double knockout mice during postnatal development and the mechanisms. Hum. Mol. Genet. 2019, 28, 1738–1751. [Google Scholar] [CrossRef]
- Gao, X.; Lu, A.; Tang, Y.; Schneppendahl, J.; Liebowitz, A.B.; Scibetta, A.C.; Morris, E.R.; Cheng, H.; Huard, C.; Amra, S.; et al. Influences of donor and host age on human muscle-derived stem cell-mediated bone regeneration. Stem. Cell Res. Ther. 2018, 9, 316. [Google Scholar] [CrossRef]
- Gao, X.; Usas, A.; Lu, A.; Kozemchak, A.; Tang, Y.; Poddar, M.; Sun, X.; Cummins, J.H.; Huard, J. Cyclooxygenase-2 deficiency impairs muscle-derived stem cell-mediated bone regeneration via cellular autonomous and non-autonomous mechanisms. Hum. Mol. Genet. 2016, 25, 3216–3231. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Sun, X.; Cheng, H.; Ruzbarsky, J.J.; Mullen, M.; Huard, M.; Huard, J. MRL/MpJ Mice Resist to Age-Related and Long-Term Ovariectomy-Induced Bone Loss: Implications for Bone Regeneration and Repair. Int. J. Mol. Sci. 2023, 24, 2396. https://doi.org/10.3390/ijms24032396
Gao X, Sun X, Cheng H, Ruzbarsky JJ, Mullen M, Huard M, Huard J. MRL/MpJ Mice Resist to Age-Related and Long-Term Ovariectomy-Induced Bone Loss: Implications for Bone Regeneration and Repair. International Journal of Molecular Sciences. 2023; 24(3):2396. https://doi.org/10.3390/ijms24032396
Chicago/Turabian StyleGao, Xueqin, Xuying Sun, Haizi Cheng, Joseph J. Ruzbarsky, Michael Mullen, Matthieu Huard, and Johnny Huard. 2023. "MRL/MpJ Mice Resist to Age-Related and Long-Term Ovariectomy-Induced Bone Loss: Implications for Bone Regeneration and Repair" International Journal of Molecular Sciences 24, no. 3: 2396. https://doi.org/10.3390/ijms24032396
APA StyleGao, X., Sun, X., Cheng, H., Ruzbarsky, J. J., Mullen, M., Huard, M., & Huard, J. (2023). MRL/MpJ Mice Resist to Age-Related and Long-Term Ovariectomy-Induced Bone Loss: Implications for Bone Regeneration and Repair. International Journal of Molecular Sciences, 24(3), 2396. https://doi.org/10.3390/ijms24032396