
Protective Effect of Uridine on Structural and Functional Rearrangements in Heart Mitochondria after a High-Dose Isoprenaline Exposure Modelling Stress-Induced Cardiomyopathy in Rats

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
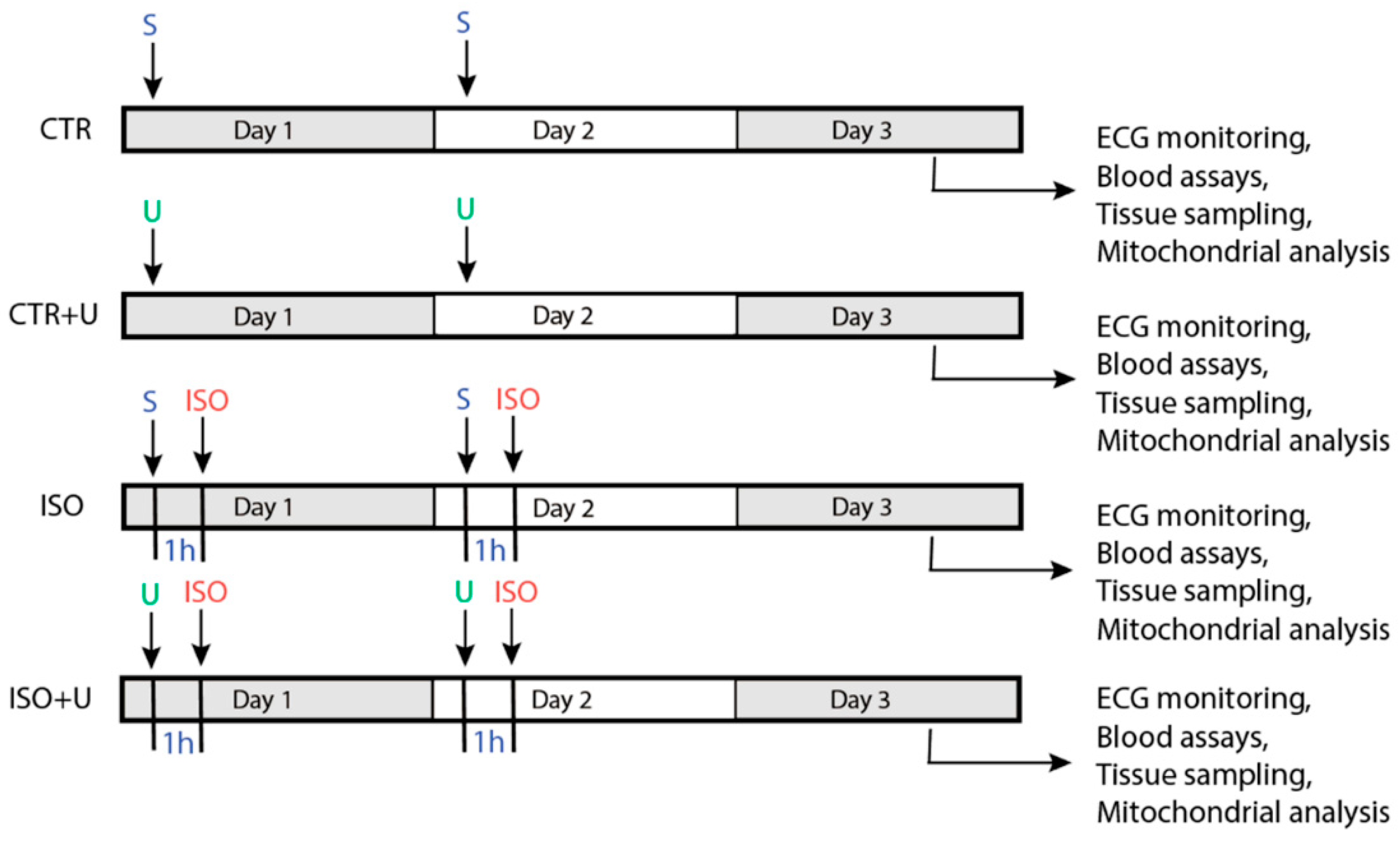
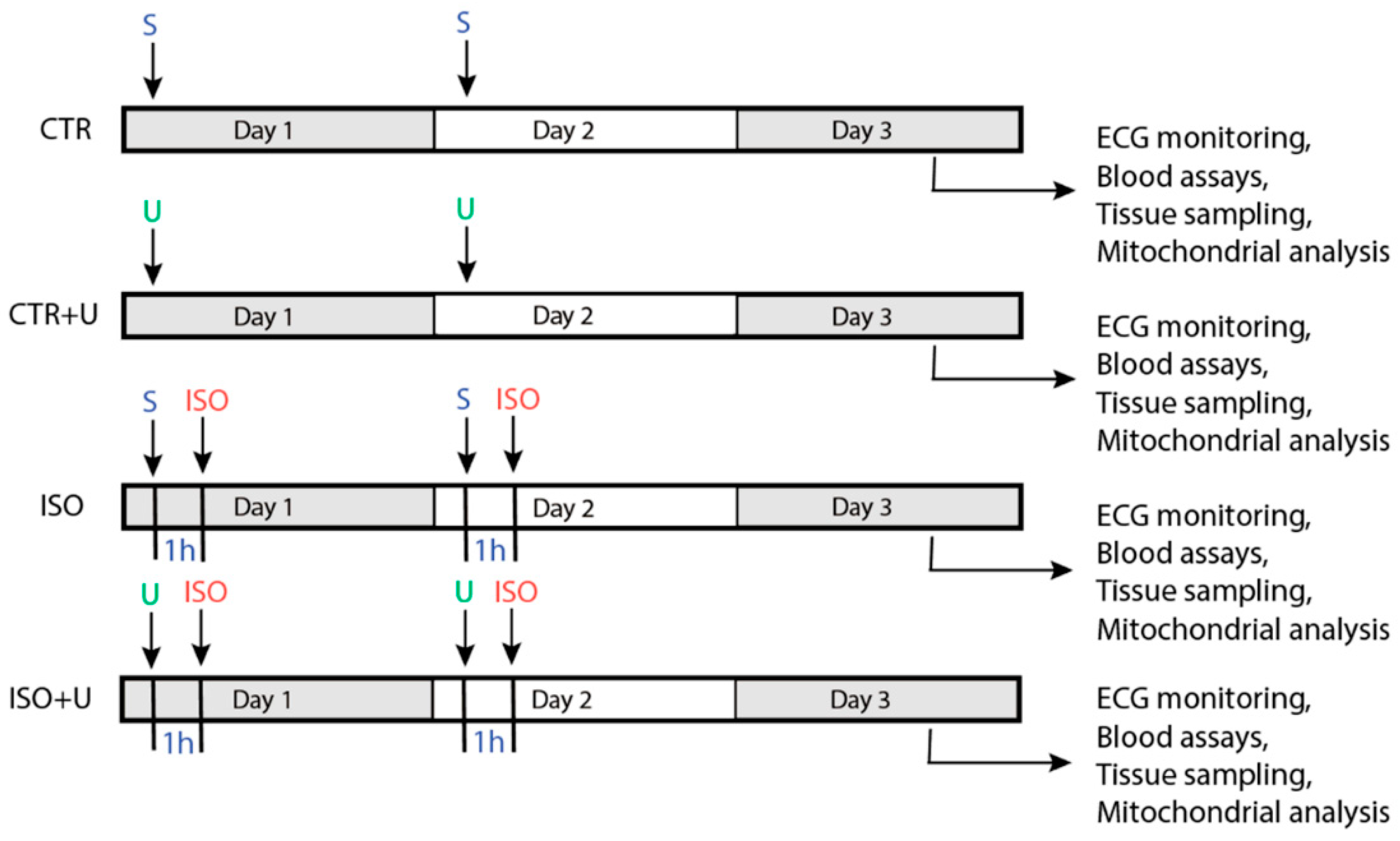
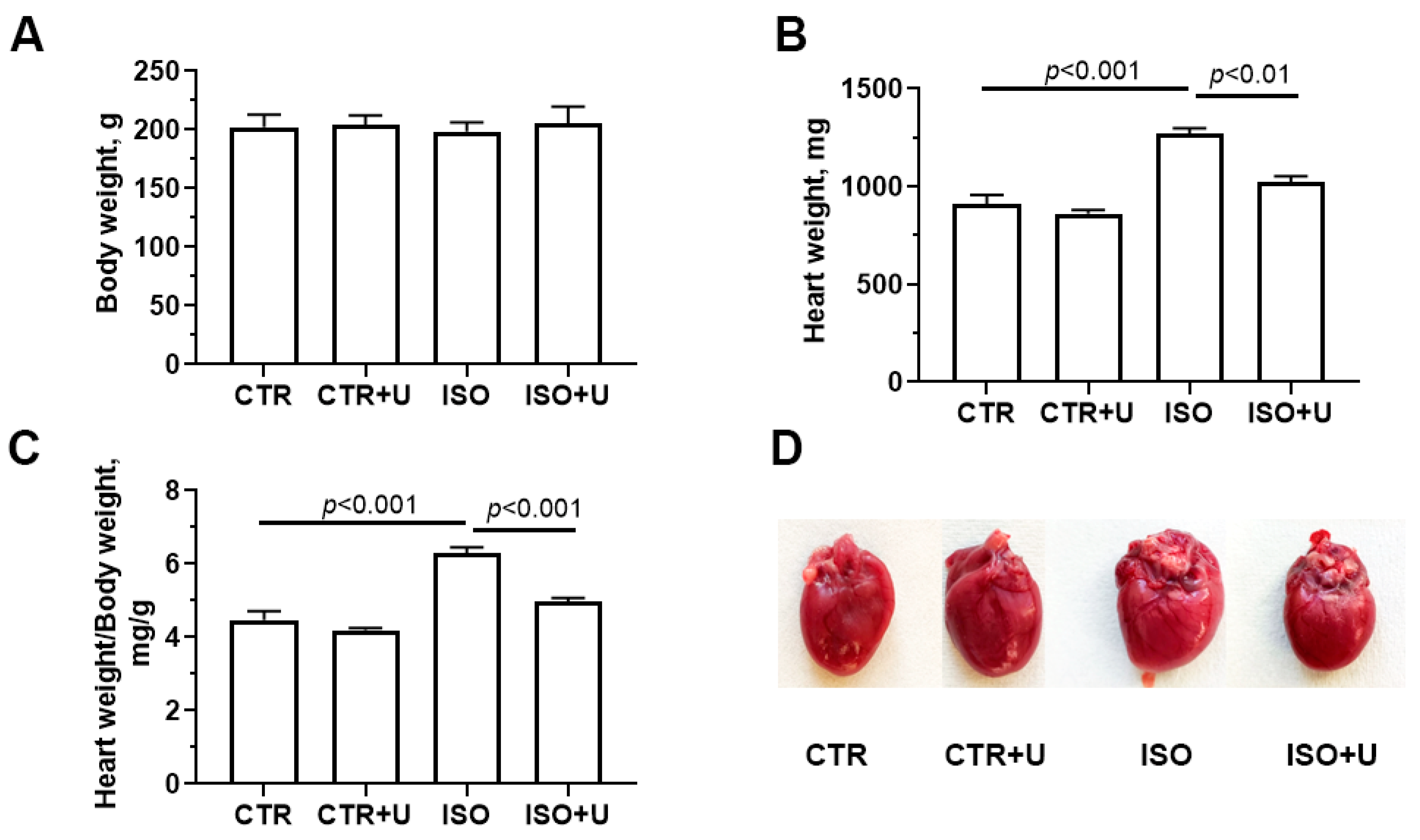
2.1. Study Design and Baseline Characteristics of Experimental Animals
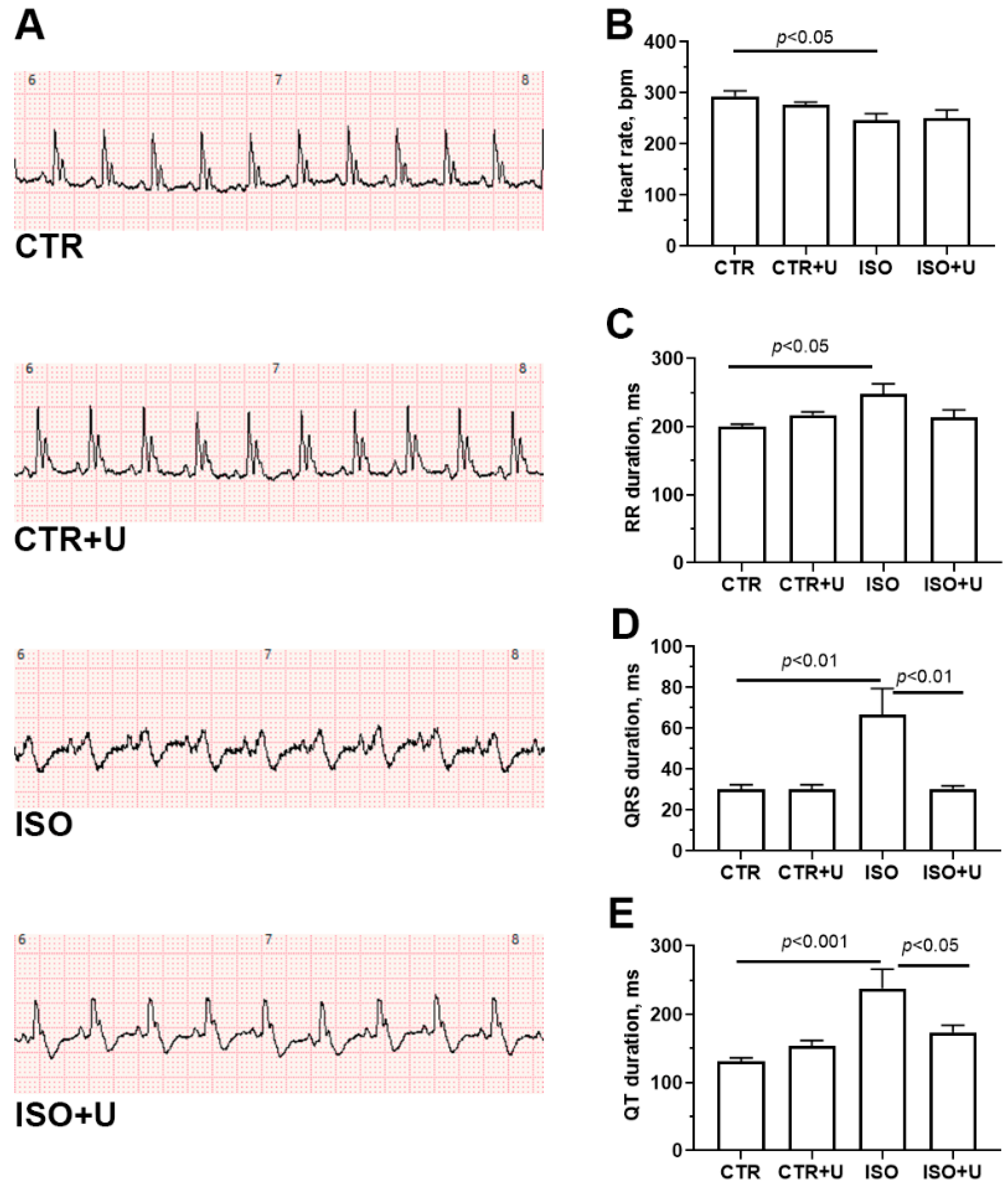
2.2. Electrophysiological Changes in the Hearts of Experimental Rats
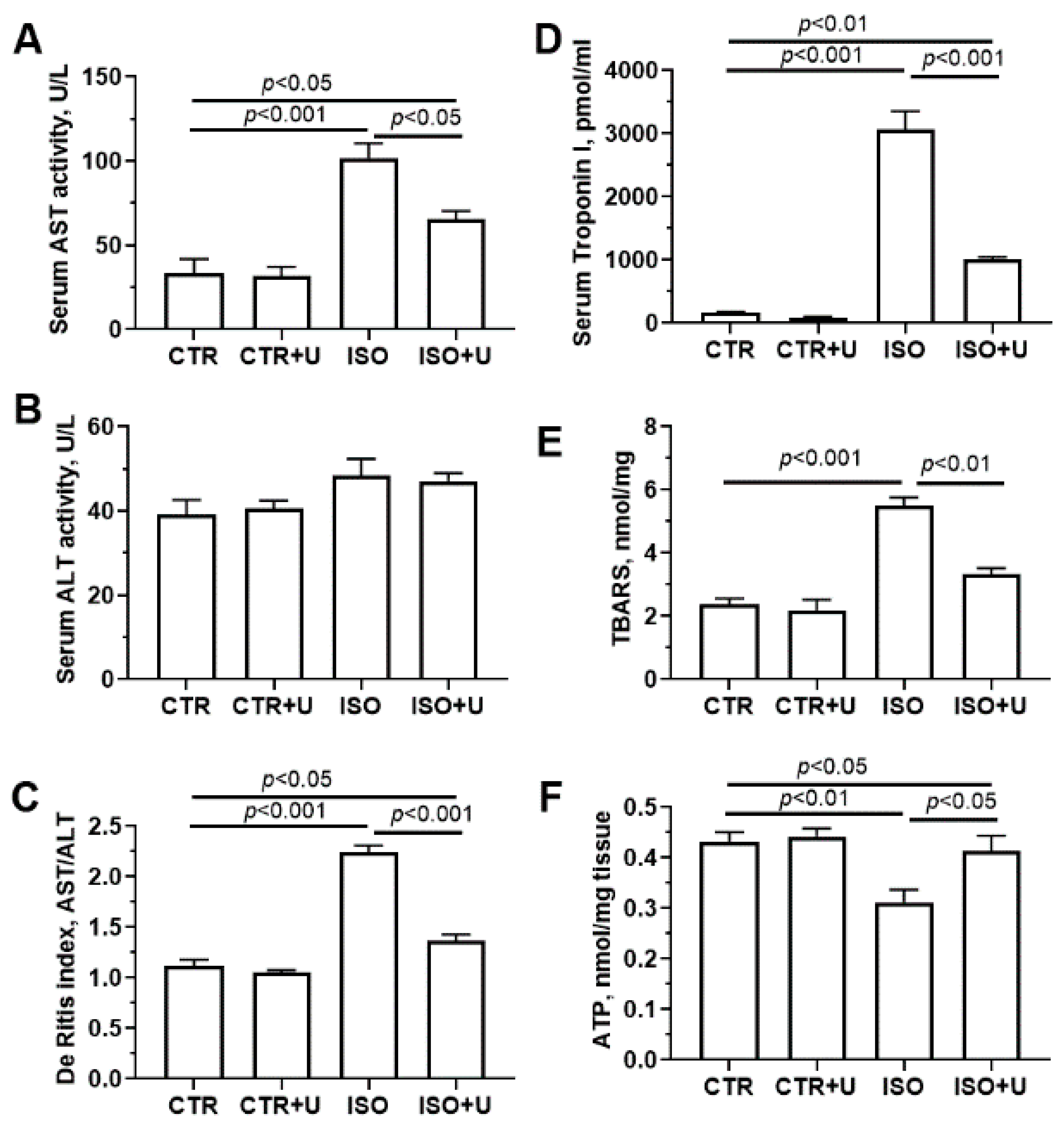
2.3. Effect of Uridine Pretreatment on Serum Cardiac Biomarkers and ATP Content in Heart Tissue Homogenates of Rats after Acute Exposure to ISO
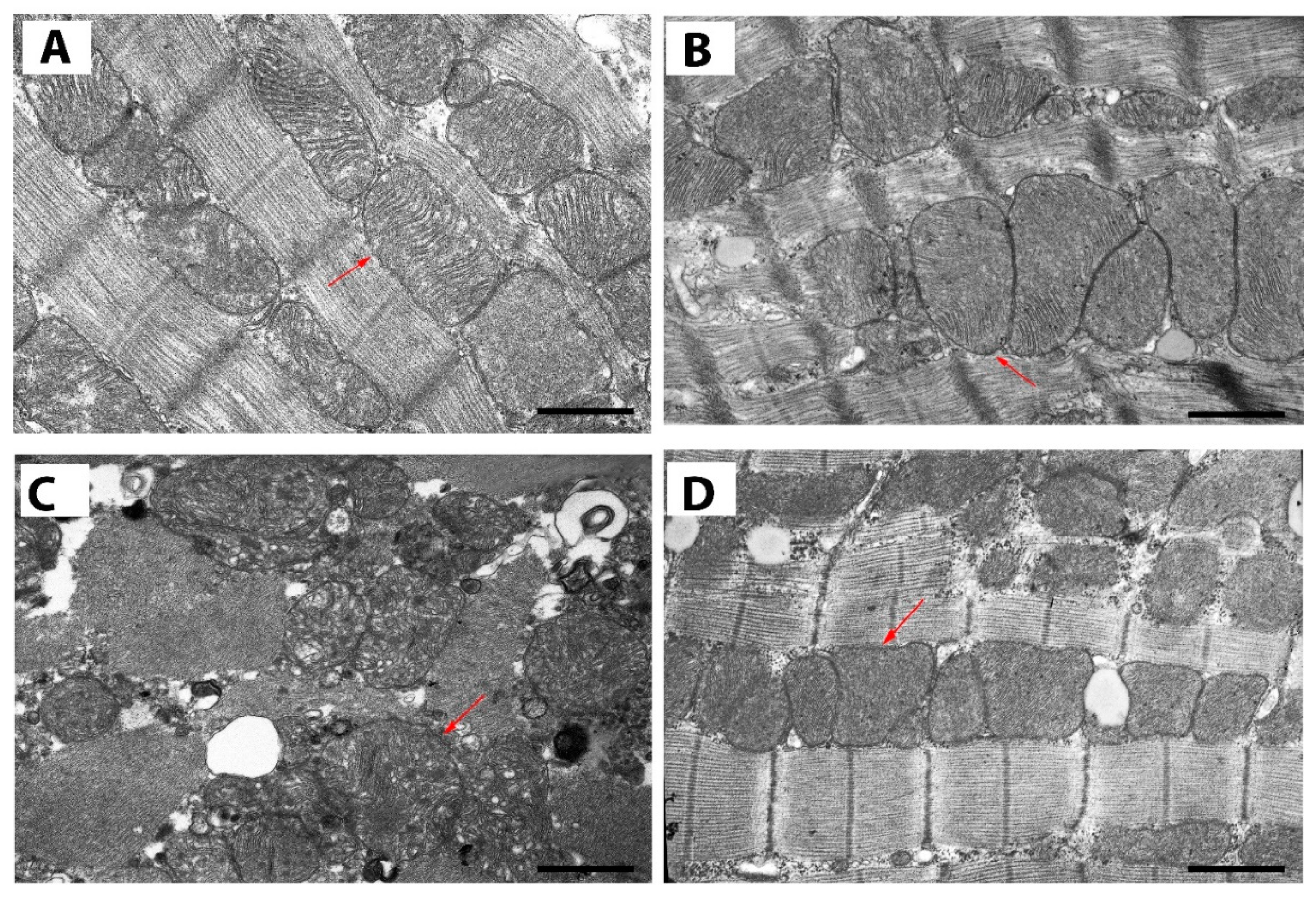
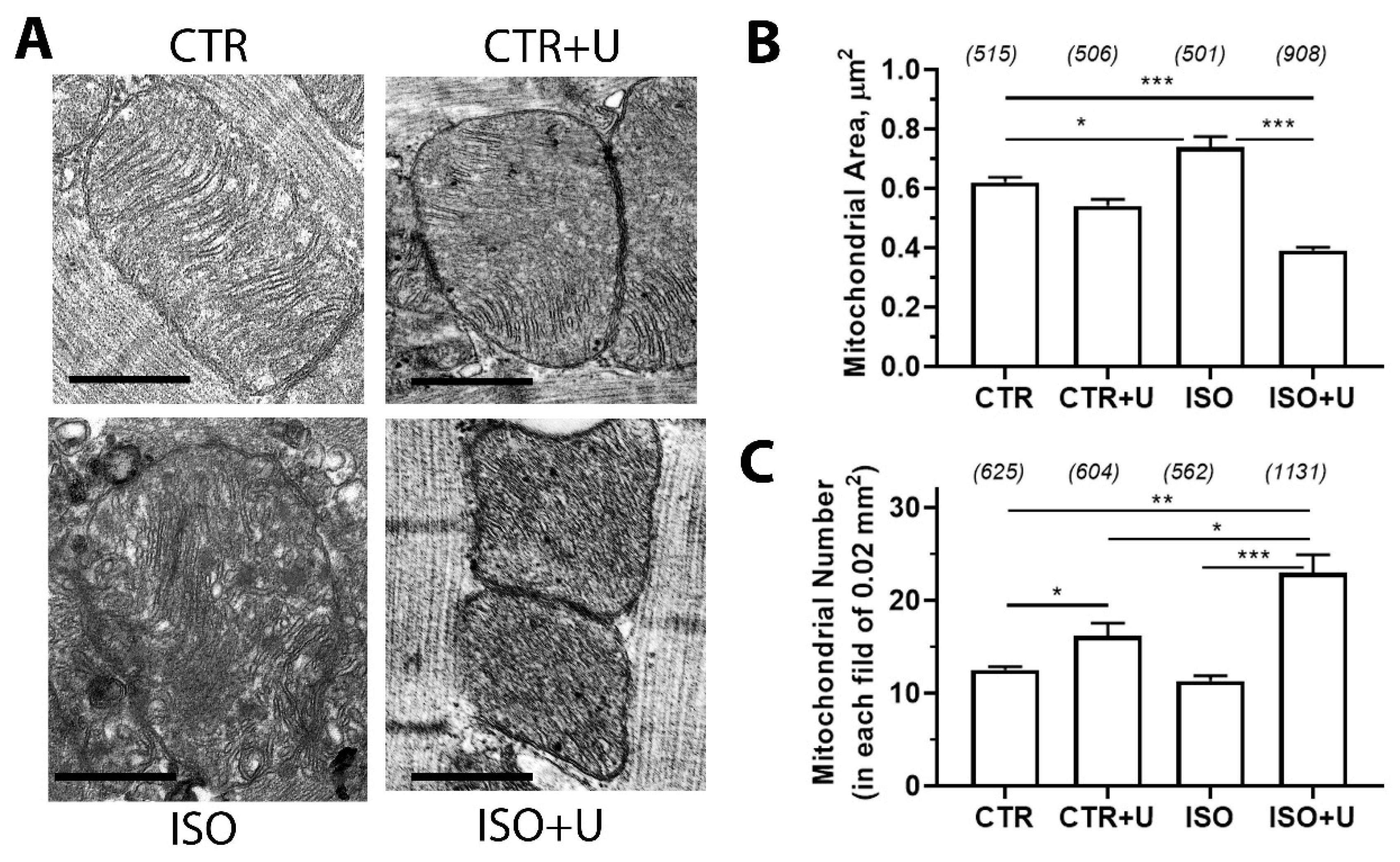
2.4. Effect of Uridine Administration on Ultrastructural Alterations in Mitochondria of Left Ventricular Cardiomyocytes from ISO-Treated Rats
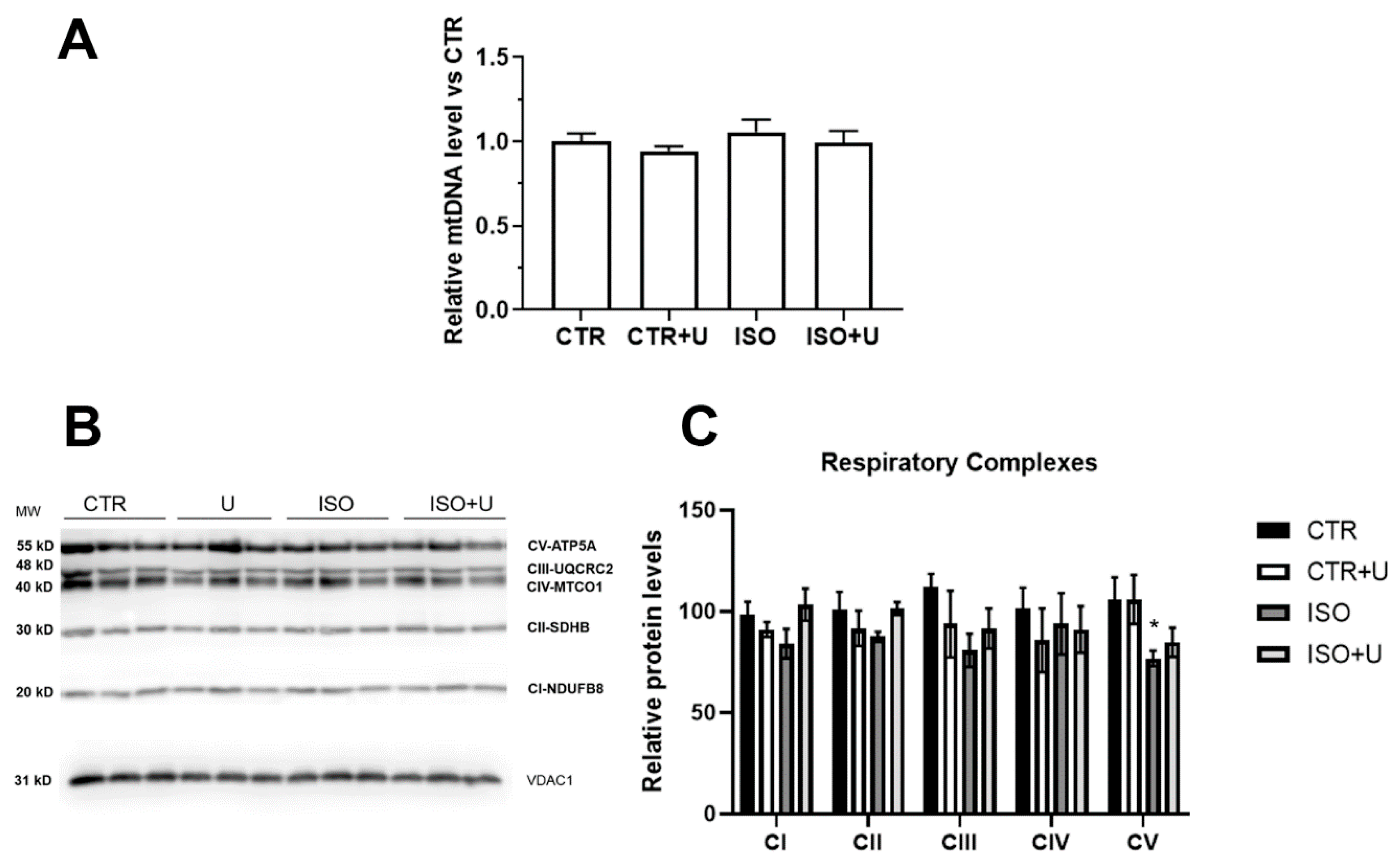
2.5. Effect of Uridine Administration on Mitochondrial Respiration, Oxidative Phosphorylation, and mtDNA Level in Experimental Rats
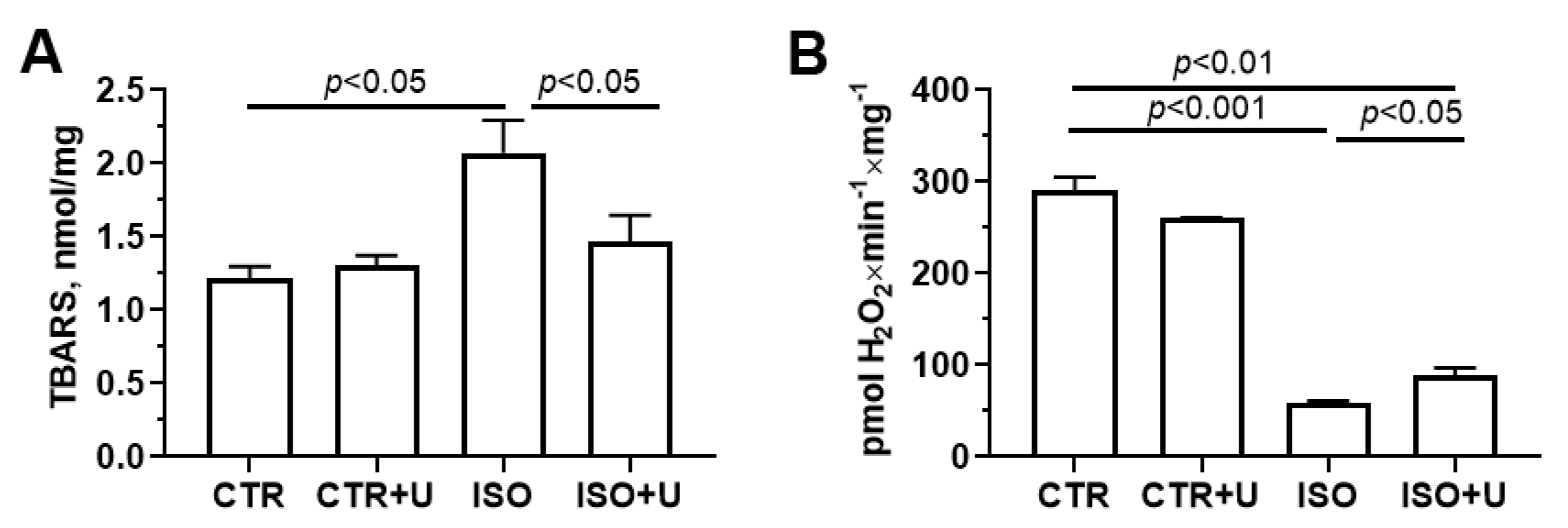
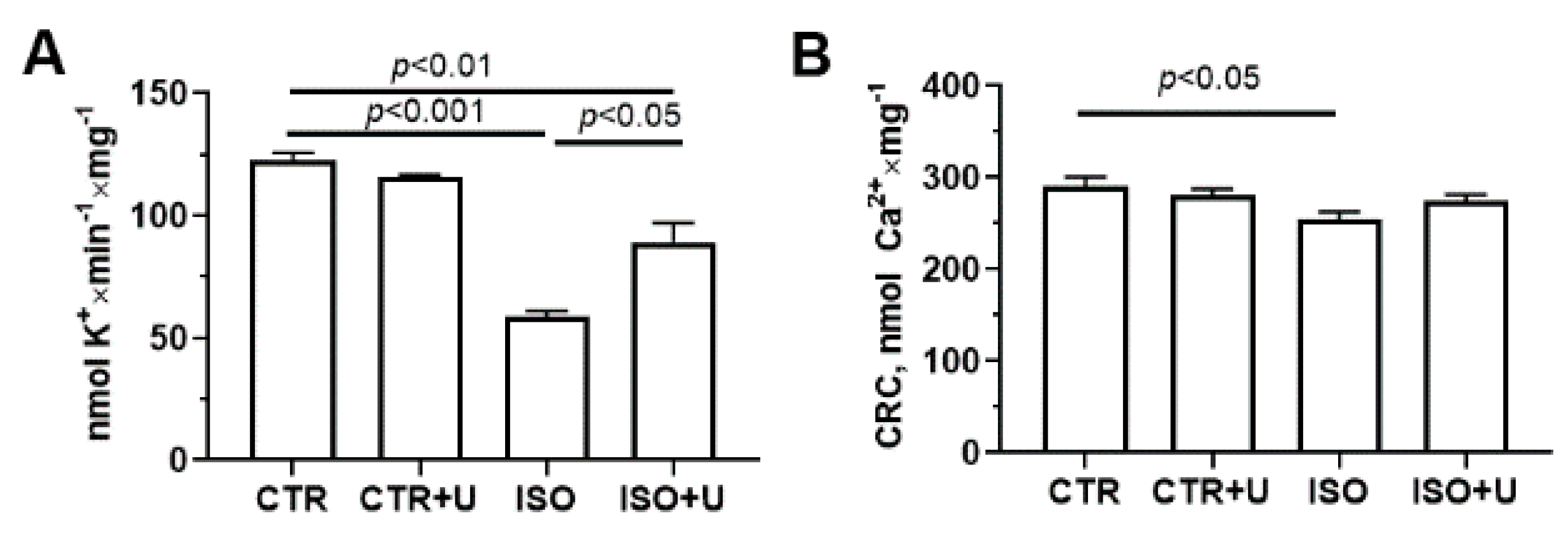
2.6. Effect of Uridine Pretreatment on Oxidative Stress Markers and Ion Transport Systems of Isolated Heart Mitochondria in Experimental Animal Groups
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Experimental Rats and Housing
4.3. Serum Cardiac Markers and Lipid Peroxidation Index
4.4. Electrocardiographic Analysis
4.5. Heart Tissue Homogenization and ATP Assay
4.6. Transmission Electron Microscopy Analysis
4.7. Isolation of Rat Heart Mitochondria
4.8. Evaluation of Mitochondrial Function
4.9. Mitochondrial DNA Copy Number
4.10. Electrophoresis and Immunoblotting
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Heart Report 2023: Confronting the World’s Number One Killer; World Heart Federation: Geneva, Switzerland, 2023; Available online: https://world-heart-federation.org/wp-content/uploads/World-Heart-Report-2023.pdf (accessed on 10 May 2023).
- Lindstrom, M.; DeCleene, N.; Dorsey, H.; Fuster, V.; Johnson, C.O.; LeGrand, K.E.; Mensah, G.A.; Razo, C.; Stark, B.; Varieur Turco, J.; et al. Global burden of cardiovascular diseases and risks collaboration, 1990–2021. J. Am. Coll. Cardiol. 2022, 80, 2372–2425. [Google Scholar] [CrossRef]
- Sharkey, S.W.; Windenburg, D.C.; Lesser, J.R.; Maron, M.S.; Hauser, R.G.; Lesser, J.N.; Haas, T.S.; Hodges, J.S.; Maron, B.J. Natural history and expansive clinical profile of stress (tako-tsubo) cardiomyopathy. J. Am. Coll. Cardiol. 2010, 55, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Nef, H.M.; Möllmann, H.; Akashi, Y.J.; Hamm, C.W. Mechanisms of stress (Takotsubo) cardiomyopathy. Nat. Rev. Cardiol. 2010, 7, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Redfors, B.; Shao, Y.; Omerovic, E. Stress-induced cardiomyopathy in a patient with chronic spinal cord transection at the level of c5: Endocrinologically mediated catecholamine toxicity. Int. J. Cardiol. 2012, 159, e61–e62. [Google Scholar] [CrossRef] [PubMed]
- Burgdorf, C.; Kurowski, V.; Bonnemeier, H.; Schunkert, H.; Radke, P.W. Long-term prognosis of the transient left ventricular dysfunction syndrome (Tako-Tsubo cardiomyopathy): Focus on malignancies. Eur. J. Heart Fail. 2008, 10, 1015–1019. [Google Scholar] [CrossRef] [PubMed]
- Fineschi, V.; Michalodimitrakis, M.; D’Errico, S.; Neri, M.; Pomara, C.; Riezzo, I.; Turillazzi, E. Insight into stress-induced cardiomyopathy and sudden cardiac death due to stress. A forensic cardio-pathologist point of view. Forensic Sci. Int. 2010, 194, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Redfors, B.; Täng, M.S.; Möllmann, H.; Troidl, C.; Szardien, S.; Hamm, C.; Nef, H.; Borén, J.; Omerovic, E. Novel rat model reveals important roles of β-adrenoreceptors in stress-induced cardiomyopathy. Int. J. Cardiol. 2013, 168, 1943–1950. [Google Scholar] [CrossRef] [PubMed]
- Yoganathan, T.; Perez-Liva, M.; Balvay, D.; Le Gall, M.; Lallemand, A.; Certain, A.; Autret, G.; Mokrani, Y.; Guillonneau, F.; Bruce, J.; et al. Acute stress induces long-term metabolic, functional, and structural remodeling of the heart. Nat. Commun. 2023, 14, 3835. [Google Scholar] [CrossRef]
- Wu, W.; Lu, Q.; Ma, S.; Du, J.C.; Huynh, K.; Duong, T.; Pang, Z.D.; Donner, D.; Meikle, P.J.; Deng, X.L.; et al. Mitochondrial damage in a Takotsubo syndrome-like mouse model mediated by activation of β-adrenoceptor-Hippo signaling pathway. Am. J. Physiol. Heart Circ. Physiol. 2023, 324, H528–H541. [Google Scholar] [CrossRef]
- Kołodzińska, A.; Czarzasta, K.; Szczepankiewicz, B.; Budnik, M.; Główczyńska, R.; Fojt, A.; Ilczuk, T.; Krasuski, K.; Borodzicz, S.; Cudnoch-Jędrzejewska, A.; et al. Isoprenaline induced Takotsubo syndrome: Histopathological analyses of female rat hearts. Cardiol. J. 2022, 29, 105–114. [Google Scholar] [CrossRef]
- Willis, B.C.; Salazar-Cantú, A.; Silva-Platas, C.; Fernández-Sada, E.; Villegas, C.A.; Rios-Argaiz, E.; González-Serrano, P.; Sánchez, L.A.; Guerrero-Beltrán, C.E.; García, N.; et al. Impaired oxidative metabolism and calcium mishandling underlie cardiac dysfunction in a rat model of post-acute isoproterenol-induced cardiomyopathy. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H467–H477. [Google Scholar] [CrossRef] [PubMed]
- Du, X. Sympatho-adrenergic mechanisms in heart failure: New insights into pathophysiology. Med. Rev. 2021, 1, 47–77. [Google Scholar] [CrossRef] [PubMed]
- Lyon, A.R.; Bossone, E.; Schneider, B.; Sechtem, U.; Citro, R.; Underwood, S.R.; Sheppard, M.N.; Figtree, G.A.; Parodi, G.; Akashi, Y.J.; et al. Current state of knowledge on Takotsubo syndrome: A Position Statement from the Taskforce on Takotsubo Syndrome of the Heart Failure Association of the European Society of Cardiology. Eur. J. Heart Fail. 2016, 18, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Krestinin, R.; Baburina, Y.; Odinokova, I.; Kruglov, A.; Sotnikova, L.; Krestinina, O. The effect of astaxanthin on mitochondrial dynamics in rat heart mitochondria under ISO-induced injury. Antioxidants 2023, 12, 1247. [Google Scholar] [CrossRef]
- Godsman, N.; Kohlhaas, M.; Nickel, A.; Cheyne, L.; Mingarelli, M.; Schweiger, L.; Hepburn, C.; Munts, C.; Welch, A.; Delibegovic, M.; et al. Metabolic alterations in a rat model of takotsubo syndrome. Cardiovasc. Res. 2022, 118, 1932–1946. [Google Scholar] [CrossRef]
- Lopaschuk, G.D.; Karwi, Q.G.; Tian, R.; Wende, A.R.; Abel, E.D. Cardiac energy metabolism in heart failure. Circ. Res. 2021, 128, 1487–1513. [Google Scholar] [CrossRef] [PubMed]
- Muntean, D.M.; Sturza, A.; Dănilă, M.D.; Borza, C.; Duicu, O.M.; Mornoș, C. The role of mitochondrial reactive oxygen species in cardiovascular injury and protective strategies. Oxidative Med. Cell. Longev. 2016, 2016, 8254942. [Google Scholar] [CrossRef]
- Zhou, B.; Tian, R. Mitochondrial dysfunction in pathophysiology of heart failure. J. Clin. Investig. 2018, 128, 3716–3726. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Kiyuna, L.A.; Albuquerque, R.P.E.; Chen, C.H.; Mochly-Rosen, D.; Ferreira, J.C.B. Targeting mitochondrial dysfunction and oxidative stress in heart failure: Challenges and opportunities. Free Radic. Biol. Med. 2018, 129, 155–168. [Google Scholar] [CrossRef]
- Monzel, A.S.; Enríquez, J.A.; Picard, M. Multifaceted mitochondria: Moving mitochondrial science beyond function and dysfunction. Nat. Metab. 2023, 5, 546–562. [Google Scholar] [CrossRef]
- Casanova, A.; Wevers, A.; Navarro-Ledesma, S.; Pruimboom, L. Mitochondria: It is all about energy. Front. Physiol. 2023, 14, 1114231. [Google Scholar] [CrossRef]
- Mannella, C.A. The relevance of mitochondrial membrane topology to mitochondrial function. Biochim. Biophys. Acta 2006, 1762, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, R.; Rigoux, L.; Albus, K.; Kretschmer, A.C.; Edwin Thanarajah, S.; Chen, W.; Hinze, Y.; Giavalisco, P.; Steculorum, S.M.; Cornely, O.A.; et al. Circulating uridine dynamically and adaptively regulates food intake in humans. Cell Rep. Med. 2023, 4, 100897. [Google Scholar] [CrossRef]
- Adant, I.; Bird, M.; Decru, B.; Windmolders, P.; Wallays, M.; de Witte, P.; Rymen, D.; Witters, P.; Vermeersch, P.; Cassiman, D.; et al. Pyruvate and uridine rescue the metabolic profile of OXPHOS dysfunction. Mol. Metab. 2022, 63, 101537. [Google Scholar] [CrossRef] [PubMed]
- Urasaki, Y.; Pizzorno, G.; Le, T.T. Uridine affects liver protein glycosylation, insulin signaling, and heme biosynthesis. PLoS ONE 2014, 9, e99728. [Google Scholar] [CrossRef]
- Aussedat, J. Effect of uridine supply on glycogen resynthesis after ischemia in the isolated perfused rat heart. Cardiovasc. Res. 1983, 17, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Bul’on, V.V.; Krylova, I.B.; Rodionova, O.M.; Selina, E.N.; Evdokimova, N.R.; Sapronov, N.S. Comparative study of cardioprotective effects of uridine-5′-monophosphate and uridine-5′-triphosphate during the early periods of acute myocardial ischemia. Bull. Exp. Biol. Med. 2007, 144, 322–325. [Google Scholar] [CrossRef]
- Krylova, I.B.; Selina, E.N.; Bulion, V.V.; Rodionova, O.M.; Evdokimova, N.R.; Belosludtseva, N.V.; Shigaeva, M.I.; Mironova, G.D. Uridine treatment prevents myocardial injury in rat models of acute ischemia and ischemia/reperfusion by activating the mitochondrial ATP-dependent potassium channel. Sci. Rep. 2021, 11, 16999. [Google Scholar] [CrossRef]
- Li, S.; Yokota, T.; Wang, P.; Ten Hoeve, J.; Ma, F.; Le, T.M.; Abt, E.R.; Zhou, Y.; Wu, R.; Nanthavongdouangsy, M.; et al. Cardiomyocytes disrupt pyrimidine biosynthesis in nonmyocytes to regulate heart repair. J. Clin. Investig. 2022, 132, e149711. [Google Scholar] [CrossRef]
- Liu, Z.; Li, W.; Geng, L.; Sun, L.; Wang, Q.; Yu, Y.; Yan, P.; Liang, C.; Ren, J.; Song, M.; et al. Cross-species metabolomic analysis identifies uridine as a potent regeneration promoting factor. Cell Discov. 2022, 8, 6. [Google Scholar] [CrossRef]
- Mironova, G.D.; Khrenov, M.O.; Talanov, E.Y.; Glushkova, O.V.; Parfenyuk, S.B.; Novoselova, T.V.; Lunin, S.M.; Belosludtseva, N.V.; Novoselova, E.G.; Lemasters, J.J. The role of mitochondrial KATP channel in anti-inflammatory effects of uridine in endotoxemic mice. Arch. Biochem. Biophys. 2018, 654, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Rozova, E.V.; Mankovskaya, I.N.; Belosludtseva, N.V.; Khmil, N.V.; Mironova, G.D. Uridine as a protector against hypoxia-induced lung injury. Sci. Rep. 2019, 9, 9418. [Google Scholar] [CrossRef] [PubMed]
- Belosludtseva, N.V.; Starinets, V.S.; Mikheeva, I.B.; Belosludtsev, M.N.; Dubinin, M.V.; Mironova, G.D.; Belosludtsev, K.N. Effect of chronic treatment with uridine on cardiac mitochondrial dysfunction in the C57BL/6 mouse model of high-fat diet-streptozotocin-induced diabetes. Int. J. Mol. Sci. 2022, 23, 10633. [Google Scholar] [CrossRef] [PubMed]
- Spitkovsky, D.; Sasse, P.; Kolossov, E.; Böttinger, C.; Fleischmann, B.K.; Hescheler, J.; Wiesner, R.J. Activity of complex III of the mitochondrial electron transport chain is essential for early heart muscle cell differentiation. FASEB J. 2004, 18, 1300–1302. [Google Scholar] [CrossRef] [PubMed]
- Titov, D.V.; Cracan, V.; Goodman, R.P.; Peng, J.; Grabarek, Z.; Mootha, V.K. Complementation of mitochondrial electron transport chain by manipulation of the NAD+/NADH ratio. Science 2016, 352, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Carreño, J.E.; Apablaza, F.; Ocaranza, M.P.; Jalil, J.E. Cardiac hypertrophy: Molecular and cellular events. Rev. Esp. Cardiol. 2006, 59, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, H.G. Catecholamine-induced cardiac hypertrophy: Significance of proto-oncogene expression. J. Mol. Med. 1997, 75, 849–859. [Google Scholar] [CrossRef]
- Nichtova, Z.; Novotova, M.; Kralova, E.; Stankovicova, T. Morphological and functional characteristics of models of experimental myocardial injury induced by isoproterenol. Gen. Physiol. Biophys. 2012, 31, 141–151. [Google Scholar] [CrossRef]
- Holanda, T.M.; Rocha, D.G.; Silveira, J.A.M.; Costa, P.P.C.; Maia, P.H.F.; Ingram, C.; Moraes, M.E.A.; Fechine, F.V.; Moraes Filho, M.O. Effect of essential oil of Alpinia zerumbet on cardiovascular and autonomic function in rats with isoproterenol induced acute myocardial infarction. An. Acad. Bras. Cienc. 2023, 95, e20201878. [Google Scholar] [CrossRef]
- Van, A.P.; Ha, T.T.; Thanh, P.M.; Lien, H.N.; Van, H.N.; Thang, H.N.; Son, S.B.; An, V.V.; Ha, T.D.; Quang, V.T. Myocardial infarction model induced by isoproterenol in rats and potential cardiovascular protective effect of a nattokinase-containing hard capsule. Phytomed. Plus 2023, 3, 100472. [Google Scholar] [CrossRef]
- Feng, W.; Li, W. The study of ISO induced heart failure rat model. Exp. Mol. Pathol. 2010, 88, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.N.; Czajka, A. Is mitochondrial DNA content a potential biomarker of mitochondrial dysfunction? Mitochondrion 2013, 13, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.S.; Longchamps, R.J.; Zhao, D.; Castellani, C.A.; Loehr, L.R.; Chang, P.P.; Matsushita, K.; Grove, M.L.; Boerwinkle, E.; Arking, D.E.; et al. Mitochondrial DNA copy number and incident heart failure: The atherosclerosis risk in communities (ARIC) study. Circulation 2020, 141, 1823–1825. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Koyama, H.; Kurajoh, M.; Shoji, T.; Tsutsumi, Z.; Moriwaki, Y. Biochemistry of uridine in plasma. Clin. Chim. Acta 2011, 412, 1712–1724. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, S.; Xie, C.; Fang, J. Uridine metabolism and its role in glucose, lipid, and amino acid homeostasis. BioMed Res. Int. 2020, 2020, 7091718. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.; Cheng, S.; Li, Q.; Chen, C.; Ouyang, M. Protective effect of uridine on atrial fibrillation: A Mendelian randomisation study. Sci. Rep. 2023, 13, 19639. [Google Scholar] [CrossRef]
- Cheng, T.; Wang, H.; Hu, Y. The causal effects of genetically determined human blood metabolites on the risk of atrial fibrillation. Front. Cardiovasc. Med. 2023, 10, 1211458. [Google Scholar] [CrossRef]
- Ma, S.; Ma, J.; Tu, Q.; Zheng, C.; Chen, Q.; Lv, W. Isoproterenol increases left atrial fibrosis and susceptibility to atrial fibrillation by inducing atrial ischemic infarction in rats. Front. Pharmacol. 2020, 11, 493. [Google Scholar] [CrossRef]
- Jiang, N.; Zhao, Z. Intestinal aging is alleviated by uridine via regulating inflammation and oxidative stress in vivo and in vitro. Cell Cycle 2022, 21, 1519–1531. [Google Scholar] [CrossRef]
- Zhang, G.X.; Kimura, S.; Nishiyama, A.; Shokoji, T.; Rahman, M.; Yao, L.; Nagai, Y.; Fujisawa, Y.; Miyatake, A.; Abe, Y. Cardiac oxidative stress in acute and chronic isoproterenol-infused rats. Cardiovasc. Res. 2005, 65, 230–238. [Google Scholar] [CrossRef]
- Ahmad, T.; Khan, T.; Kirabo, A.; Shah, A.J. Antioxidant flavonoid diosmetin is cardioprotective in a rat model of myocardial infarction induced by beta 1-adrenergic receptors activation. Curr. Issues Mol. Biol. 2023, 45, 4675–4686. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, L.A.; Hassan, O.F.; Galal, O.; Mansour, D.F.; El-Khatib, A. Beneficial effects of benfotiamine, a NADPH oxidase inhibitor, in isoproterenol-induced myocardial infarction in rats. PLoS ONE 2020, 15, e0232413. [Google Scholar] [CrossRef] [PubMed]
- Abramicheva, P.A.; Andrianova, N.V.; Babenko, V.A.; Zorova, L.D.; Zorov, S.D.; Pevzner, I.B.; Popkov, V.A.; Semenovich, D.S.; Yakupova, E.I.; Silachev, D.N.; et al. Mitochondrial network: Electric cable and more. Biochemistry (Moscow) 2023, 88, 1596–1607. [Google Scholar] [CrossRef]
- Dubinin, M.V.; Starinets, V.S.; Belosludtseva, N.V.; Mikheeva, I.B.; Chelyadnikova, Y.A.; Penkina, D.K.; Vedernikov, A.A.; Belosludtsev, K.N. The effect of uridine on the state of skeletal muscles and the functioning of mitochondria in Duchenne dystrophy. Int. J. Mol. Sci. 2022, 23, 10660. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hady, E.; Mohamed, F.; Ahmed, M.; Abdel-Salam, M.; Ayobe, M. Supplementation of lipoic acid, zinc and clopidogrel reduces mortality rate and incidence of ventricular arrhythmia in experimental myocardial infarction. Front. Physiol. 2021, 12, 582223. [Google Scholar] [CrossRef]
- Ahmad, T.; Fiuzat, M.; Neely, B.; Neely, M.L.; Pencina, M.J.; Kraus, W.E.; Zannad, F.; Whellan, D.J.; Donahue, M.P.; Piña, I.L.; et al. Biomarkers of myocardial stress and fibrosis as predictors of mode of death in patients with chronic heart failure. JACC Heart Fail. 2014, 2, 260–268. [Google Scholar] [CrossRef]
- Botros, M.; Sikaris, K. The De Ritis Ratio: The Test of Time. Clin. Biochem. Rev. 2013, 34, 117–130. [Google Scholar]
- Yogeeta, S.K.; Raghavendran, H.R.; Gnanapragasam, A.; Subhashini, R.; Devaki, T. Ferulic acid with ascorbic acid synergistically extenuates the mitochondrial dysfunction during beta-adrenergic catecholamine induced cardiotoxicity in rats. Chem. Biol. Interact. 2006, 163, 160–169. [Google Scholar] [CrossRef]
- Mironova, G.D.; Negoda, A.E.; Marinov, B.S.; Paucek, P.; Costa, A.D.; Grigoriev, S.M.; Skarga, Y.Y.; Garlid, K.D. Functional distinctions between the mitochondrial ATP-dependent K+ channel (mitoKATP) and its inward rectifier subunit (mitoKIR). J. Biol. Chem. 2004, 279, 32562–32568. [Google Scholar] [CrossRef]
- Belosludtseva, N.V.; Pavlik, L.L.; Belosludtsev, K.N.; Saris, N.L.; Shigaeva, M.I.; Mironova, G.D. The short-term opening of cyclosporin A-independent palmitate/Sr2+-induced pore can underlie ion efflux in the oscillatory mode of functioning of rat liver mitochondria. Membranes 2022, 12, 667. [Google Scholar] [CrossRef] [PubMed]
- Skulachev, V.P.; Vyssokikh, M.Y.; Chernyak, B.V.; Mulkidjanian, A.Y.; Skulachev, M.V.; Shilovsky, G.A.; Lyamzaev, K.G.; Borisov, V.B.; Severin, F.F.; Sadovnichii, V.A. Six functions of respiration: Isn’t it time to take control over ROS production in mitochondria, and aging along with it? Int. J. Mol. Sci. 2023, 24, 12540. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | V Respiration, nmol O2·min−1·mg−1 pr | RCR | ADP/O | Tph, s | |||
|---|---|---|---|---|---|---|---|
| State 2 | State 3 | State 4 | State 3UDNP | ||||
| CTR | 4.6 ± 0.5 | 67.4 ± 3.7 | 13.9 ± 0.8 | 63.8 ± 3.4 | 4.90 ± 0.22 | 2.96 ± 0.12 | 43.6 ± 2.8 |
| CTR + U | 4.2 ± 1.3 | 57.9 ± 1.2 | 12.5 ± 1.3 | 61.5 ± 3.3 | 4.67 ± 0.20 | 2.74 ± 0.07 | 42.0 ± 4.3 |
| ISO | 4.0 ± 0.8 | 45.4 ± 1.8 * | 13.5 ± 1.0 | 42.9 ± 1.8 * | 3.38 ± 0.18 * | 2.18 ± 0.08 * | 69.6 ± 3.9 * |
| ISO + U | 3.9 ± 0.9 | 54.8 ± 2.1 *,# | 13.1 ± 0.7 | 47.9 ± 2.6 * | 4.20 ± 0.19 *,# | 2.53 ± 0.10 | 49.7 ± 4.6 *,# |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belosludtseva, N.V.; Pavlik, L.L.; Mikheeva, I.B.; Talanov, E.Y.; Serov, D.A.; Khurtin, D.A.; Belosludtsev, K.N.; Mironova, G.D. Protective Effect of Uridine on Structural and Functional Rearrangements in Heart Mitochondria after a High-Dose Isoprenaline Exposure Modelling Stress-Induced Cardiomyopathy in Rats. Int. J. Mol. Sci. 2023, 24, 17300. https://doi.org/10.3390/ijms242417300
Belosludtseva NV, Pavlik LL, Mikheeva IB, Talanov EY, Serov DA, Khurtin DA, Belosludtsev KN, Mironova GD. Protective Effect of Uridine on Structural and Functional Rearrangements in Heart Mitochondria after a High-Dose Isoprenaline Exposure Modelling Stress-Induced Cardiomyopathy in Rats. International Journal of Molecular Sciences. 2023; 24(24):17300. https://doi.org/10.3390/ijms242417300
Chicago/Turabian StyleBelosludtseva, Natalia V., Lubov L. Pavlik, Irina B. Mikheeva, Eugeny Yu. Talanov, Dmitriy A. Serov, Dmitriy A. Khurtin, Konstantin N. Belosludtsev, and Galina D. Mironova. 2023. "Protective Effect of Uridine on Structural and Functional Rearrangements in Heart Mitochondria after a High-Dose Isoprenaline Exposure Modelling Stress-Induced Cardiomyopathy in Rats" International Journal of Molecular Sciences 24, no. 24: 17300. https://doi.org/10.3390/ijms242417300
APA StyleBelosludtseva, N. V., Pavlik, L. L., Mikheeva, I. B., Talanov, E. Y., Serov, D. A., Khurtin, D. A., Belosludtsev, K. N., & Mironova, G. D. (2023). Protective Effect of Uridine on Structural and Functional Rearrangements in Heart Mitochondria after a High-Dose Isoprenaline Exposure Modelling Stress-Induced Cardiomyopathy in Rats. International Journal of Molecular Sciences, 24(24), 17300. https://doi.org/10.3390/ijms242417300