
Paracrine Responses of Cardiosphere-Derived Cells to Cytokines and TLR Ligands: A Comparative Analysis
 , , ,
, , ,
Abstract
:
1. Introduction
2. Results
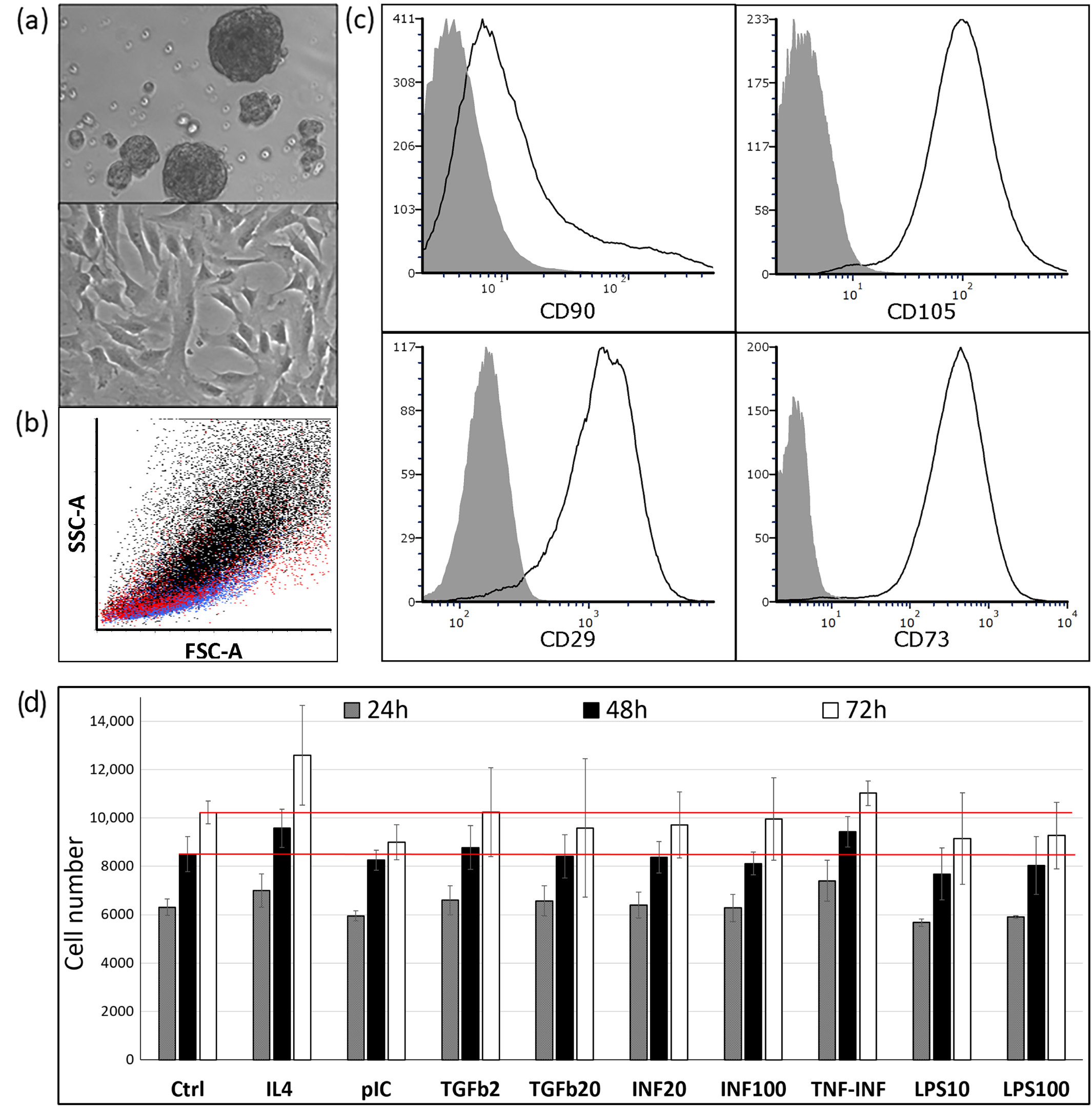
2.1. The Phenotype and Culture Characteristics of CDCs
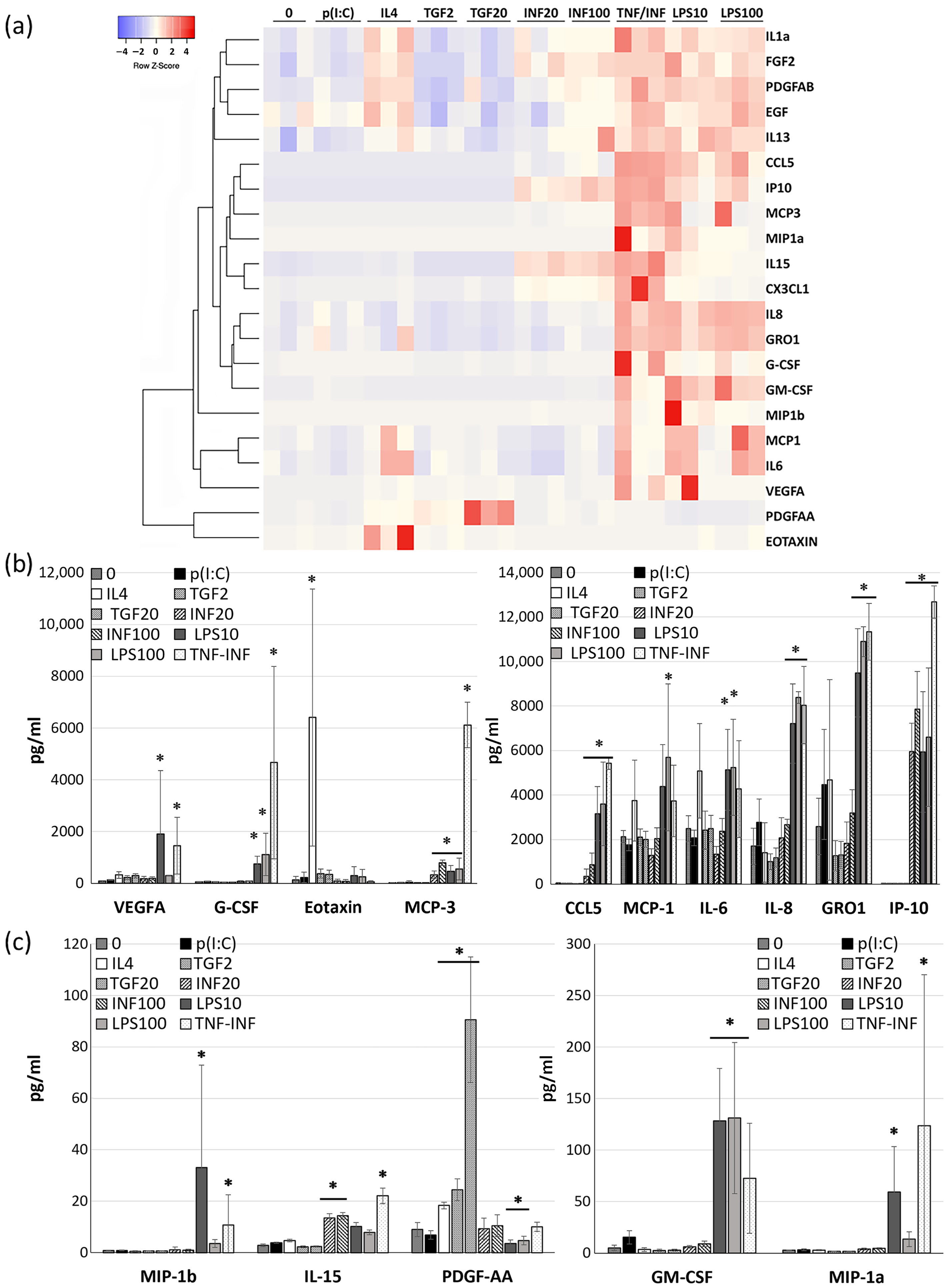
2.2. The Effects of Cytokines and TLR Ligands on Cytokine Expression Pattern of CDCs
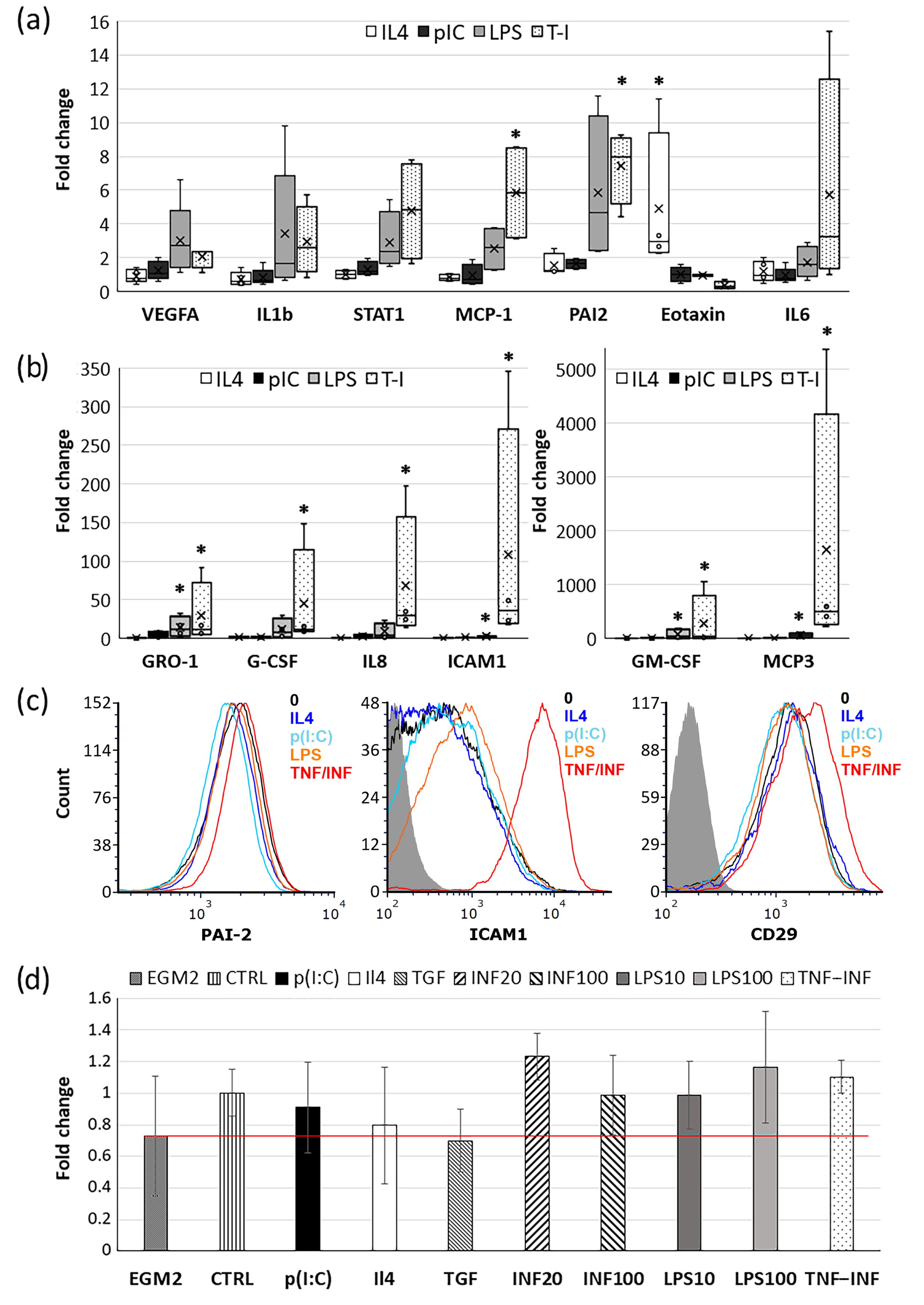
2.3. Effects of Cytokines and TLR Ligands on CDC Gene Expression
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.1.1. Isolation of Peripheral Blood Mononuclear Cells (PBMCs)
4.1.2. Coculture of PBMCs with Cardiosphere-Derived Cells (CDCs)
4.2. Cytokine Treatments
4.3. Cell Viability Assay
4.4. Flow Cytometry
4.5. qRT-PCR Analysis
4.6. Multiplex Immunoassay (Magpix)
4.7. Fibrin Gel Angiogenesis Bead Assay
4.8. Statistical Analysis
5. Conclusions
Study Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, T.S.; Cheng, K. Direct comparison of different stem cell types and subpopulations reveals superior paracrine potency and myocardial repair efficacy with cardiosphere-derived cells. J. Am. Coll. Cardiol. 2012, 59, 942–953. [Google Scholar] [CrossRef] [PubMed]
- Marbán, E.; Liao, K. On the cellular origin of cardiosphere-derived cells (CDCs). Basic. Res. Cardiol. 2022, 117, 12. [Google Scholar] [CrossRef]
- Messina, E.; De Angelis, L. Isolation and expansion of adult cardiac stem cells from human and murine heart. Circ. Res. 2004, 95, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Bearzi, C.; Rota, M. Human cardiac stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 14068–14073. [Google Scholar] [CrossRef] [PubMed]
- Carr, C.A.; Stuckey, D.J. Cardiosphere-derived cells improve function in the infarcted rat heart for at least 16 weeks--an MRI study. PLoS ONE 2011, 6, e25669. [Google Scholar] [CrossRef] [PubMed]
- Barile, L.; Gherghiceanu, M. Human cardiospheres as a source of multipotent stem and progenitor cells. Stem Cells Int. 2013, 2013, 916837. [Google Scholar] [CrossRef] [PubMed]
- Dey, D.; Leng, H. Dissecting the molecular relationship among various cardiogenic progenitor cells. Circ. Res. 2013, 112, 1253–1262. [Google Scholar] [CrossRef]
- Li, T.S.; Cheng, K. Cardiospheres recapitulate a niche-like microenvironment rich in stemness and cell-matrix interactions, rationalizing their enhanced functional potency for myocardial repair. Stem Cells 2010, 28, 2088–2098. [Google Scholar] [CrossRef]
- Tang, X.L.; Li, Q. Long-Term Outcome of Administration of c-kit(POS) Cardiac Progenitor Cells After Acute Myocardial Infarction: Transplanted Cells Do not Become Cardiomyocytes, but Structural and Functional Improvement and Proliferation of Endogenous Cells Persist for at Least One Year. Circ. Res. 2016, 118, 1091–1105. [Google Scholar]
- Ellison-Hughes, G.M.; Madeddu, P. Exploring pericyte and cardiac stem cell secretome unveils new tactics for drug discovery. Pharmacol. Ther. 2017, 171, 1–12. [Google Scholar] [CrossRef]
- Dutton, L.C.; Dudhia, J. Cardiosphere-derived cells suppress allogeneic lymphocytes by production of PGE2 acting via the EP4 receptor. Sci. Rep. 2018, 8, 13351. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, Z. Pericardial application as a new route for implanting stem-cell cardiospheres to treat myocardial infarction. J. Physiol. 2018, 596, 2037–2054. [Google Scholar] [CrossRef] [PubMed]
- Lopez, E.; Marinaro, F. The Immunomodulatory Signature of Extracellular Vesicles from Cardiosphere-Derived Cells: A Proteomic and miRNA Profiling. Front. Cell Dev. Biol. 2020, 8, 321. [Google Scholar] [CrossRef]
- Neri, M.; Fineschi, V. Cardiac oxidative stress and inflammatory cytokines response after myocardial infarction. Curr. Vasc. Phamacol. 2015, 13, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Martins-Marques, T. Connecting different heart diseases through intercellular communication. Biol. Open 2021, 10, bio058777. [Google Scholar] [CrossRef] [PubMed]
- Moeinabadi-Bidgoli, K.; Babajani, A.; Yazdanpanah, G.; Farhadihosseinabadi, B.; Jamshidi, E.; Bahrami, S.; Niknejad, H. Translational insights into stem cell preconditioning: From molecular mechanisms to preclinical applications. Biomed. Pharmacother. 2021, 142, 112026. [Google Scholar] [CrossRef] [PubMed]
- Matta, A.; Nader, V.; Lebrin, M.; Gross, F.; Prats, A.C.; Cussac, D.; Galinier, M.; Roncalli, J. Pre-Conditioning Methods and Novel Approaches with Mesenchymal Stem Cells Therapy in Cardiovascular Disease. Cells. 2022, 11, 1620. [Google Scholar] [CrossRef]
- Yan, F.; Yao, Y.; Chen, L.; Li, Y.; Sheng, Z.; Ma, G. Hypoxic preconditioning improves survival of cardiac progenitor cells: Role of stromal cell derived factor-1α-CXCR4 axis. PLoS ONE. 2012, 7, e37948. [Google Scholar] [CrossRef]
- Tan, S.C.; Gomes, R.S.; Yeoh, K.K.; Perbellini, F.; Malandraki-Miller, S.; Ambrose, L.; Heather, L.C.; Faggian, G.; Schofield, C.J.; Davies, K.E.; et al. Preconditioning of Cardiosphere-Derived Cells with Hypoxia or Prolyl-4-Hydroxylase Inhibitors Increases Stemness and Decreases Reliance on Oxidative Metabolism. Cell Transplant. 2016, 25, 35–53. [Google Scholar] [CrossRef]
- Waterman, R.S.; Tomchuck, S.L. A new mesenchymal stem cell (MSC) paradigm: Polarization into a pro-inflammatory MSC1 or an Immunosuppressive MSC2 phenotype. PLoS ONE 2010, 5, e10088. [Google Scholar] [CrossRef]
- Waterman, R.S.; Morgenweck, J. Anti-inflammatory mesenchymal stem cells (MSC2) attenuate symptoms of painful diabetic peripheral neuropathy. Stem Cells Transl. Med. 2012, 1, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Fonsatti, E.; Maio, M. Highlights on endoglin (CD105): From basic findings towards clinical applications in human cancer. J. Transl. Med. 2004, 2, 18. [Google Scholar] [CrossRef] [PubMed]
- Malliaras, K.; Makkar, R.R. Intracoronary cardiosphere-derived cells after myocardial infarction: Evidence of therapeutic regeneration in the final 1-year results of the CADUCEUS trial (cardiosphere-derived autologous stem cells to reverse ventricular dysfunction). J. Am. Coll. Cardiol. 2014, 63, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Ishigami, S.; Ohtsuki, S. Intracoronary autologous cardiac progenitor cell transfer in patients with hypoplastic left heart syndrome: The TICAP prospective phase 1 controlled trial. Circ. Res. 2015, 116, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.; Barile, L. Regenerative potential of cardiosphere-derived cells expanded from percutaneous endomyocardial biopsy specimens. Circulation 2007, 115, 896–908. [Google Scholar] [CrossRef] [PubMed]
- Kogan, P.S.; Wirth, F. Uncovering the molecular identity of cardio-sphere-derived cells (CDCs) by single-cell RNA sequencing. Basic. Res. Cardiol. 2022, 117, 11. [Google Scholar] [CrossRef] [PubMed]
- Chimenti, I.; Smith, R.R. Relative roles of direct regeneration versus paracrine effects of human cardiosphere-derived cells transplanted into infarcted mice. Circ. Res. 2010, 106, 971–980. [Google Scholar] [CrossRef]
- Tseliou, E.; de Couto, G. Angiogenesis, cardiomyocyte proliferation and anti-fibrotic effects underlie structural preservation post-infarction by intramyocardially-injected cardiospheres. PLoS ONE 2014, 9, e88590. [Google Scholar] [CrossRef]
- Poltorak, A.; He, X. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: Mutations in Tlr4 gene. Science 1998, 282, 2085–2088. [Google Scholar] [CrossRef]
- Zhao, W.; Zhao, T. Platelet-derived growth factor involvement in myocardial remodeling following infarction. J. Mol. Cell Cardiol. 2011, 51, 830–838. [Google Scholar] [CrossRef]
- Ezoe, K.; Horikoshi, T. Tumor necrosis factor-alpha increased the integrin alpha 2 beta 1 expression and cell attachment to type I collagen in human dermal fibroblasts. Biochem. Biophys. Res. Commun. 1993, 192, 281–287. [Google Scholar] [CrossRef]
- van Megen, K.M.; van 't Wout, E.T. Activated Mesenchymal Stromal Cells Process and Present Antigens Regulating Adaptive Immunity. Front. Immunol. 2019, 10, 694. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Zhao, X. In-flammatory cytokine-induced intercellular adhesion molecule-1 and vascular cell adhesion mole-cule-1 in mesenchymal stem cells are critical for immunosuppression. J. Immunol. 2010, 184, 2321–2328. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Huang, K.; Xia, W.; Shi, J.; Liu, Q.; Zhang, X.; Li, G.; Chen, J.; Wang, T.; Chen, X.; et al. Mesenchymal Stromal Cells Rapidly Suppress TCR Signaling-Mediated Cytokine Transcription in Activated T Cells Through the ICAM-1/CD43 Interaction. Front. Immunol. 2021, 12, 609544. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, R.J.; Yates, C.C.; Wells, A. IP-10 blocks vascular endothelial growth factor-induced endothelial cell motility and tube formation via inhibition of calpain. Circ. Res. 2006, 98, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Angiolillo, A.L.; Sgadari, C. Human interfer-on-inducible protein 10 is a potent inhibitor of angiogenesis in vivo. J. Exp. Med. 1995, 182, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Emmert, M.Y.; Burrello, J. Intracoronary delivery of extracellular vesicles from human cardiac progenitor cells reduces infarct size in porcine acute myocardial infarction. Eur. Heart J. 2023, ehad636. [Google Scholar] [CrossRef] [PubMed]
- Barile, L.; Milano, G.; Vassalli, G. Beneficial effects of exosomes secreted by cardiac-derived progenitor cells and other cell types in myocardial ischemia. Stem Cell Investig. 2017, 4, 93. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.L.; Zhu, W.; Cheng, M.; Chen, L.; Zhang, J.; Sun, T.; Kishore, R.; Phillips, M.I.; Losordo, D.W.; Qin, G. Hypoxic preconditioning enhances the benefit of cardiac progenitor cell therapy for treatment of myocardial infarction by inducing CXCR4 expression. Circ. Res. 2009, 104, 1209–1216. [Google Scholar] [CrossRef]
- Zhang, L.; Geng, W.R.; Hu, J.; Chen, X.M.; Shen, Y.L.; Wang, L.L.; Jiang, J.P.; Chen, Y.Y. Lipopolysaccharide pretreatment promotes cardiac stem cell migration through heat shock protein 90-dependent β-catenin activation. Life Sci. 2016, 153, 132–140. [Google Scholar] [CrossRef]
- Yue, R.; Fu, W.; Liao, X.; Lan, C.; Liao, Q.; Li, L.; Yang, D.; Xia, X.; Chen, X.; Zeng, C.; et al. Metformin promotes the survival of transplanted cardiosphere-derived cells thereby enhancing their therapeutic effect against myocardial infarction. Stem Cell Res. Ther. 2017, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Höving, A.L.; Schmidt, K.E.; Merten, M.; Hamidi, J.; Rott, A.K.; Faust, I.; Greiner, J.F.W.; Gummert, J.; Kaltschmidt, B.; Kaltschmidt, C.; et al. Blood Serum Stimulates p38-Mediated Proliferation and Changes in Global Gene Expression of Adult Human Cardiac Stem Cells. Cells 2020, 9, 1472. [Google Scholar] [CrossRef] [PubMed]
- Finger, S.; Knorr, M. A sequential interferon gamma directed chemotactic cellular immune response determines survival and cardiac function post-myocardial infarction. Cardiovasc. Res. 2019, 115, 1907–1917. [Google Scholar] [CrossRef] [PubMed]
- Nian, M.; Lee, P.; Khaper, N.; Liu, P. Inflammatory cytokines and postmyocardial infarction remodeling. Circ. Res. 2004, 94, 1543–1553. [Google Scholar] [CrossRef] [PubMed]
- Domenis, R.; Cifù, A. Pro inflammatory stimuli enhance the immunosuppressive functions of adipose mesenchymal stem cells-derived exosomes. Sci. Rep. 2018, 8, 13325. [Google Scholar] [CrossRef] [PubMed]
- Daseke, M.J., 2nd; Tenkorang-Impraim, M.A.A. Exogenous IL4 shuts off pro-inflammation in neutrophils while stimulating anti-inflammation in macrophages to induce neutrophil phagocytosis following myocardial infarction. J. Mol. Cell Cardiol. 2020, 145, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, S.; Tapping, R.I. Toll-like receptor signaling in cell proliferation and survival. Cytokine. 2010, 49, 1–9. [Google Scholar] [CrossRef] [PubMed]
- van den Borne, P.; Quax, P.H. The multifaceted functions of CXCL10 in cardiovascular disease. Biomed. Res. Int. 2014, 2014, 893106. [Google Scholar] [CrossRef]
- Rosenblatt-Velin, N.; Badoux, S.; Liaudet, L. Pharmacological Therapy in the Heart as an Alternative to Cellular Therapy: A Place for the Brain Natriuretic Peptide? Stem Cells Int. 2016, 2016, 5961342. [Google Scholar] [CrossRef]
- Stastna, M.; Chimenti, I.; Marbán, E.; Van Eyk, J.E. Identification and functionality of proteomes secreted by rat cardiac stem cells and neonatal cardiomyocytes. Proteomics. 2010, 10, 245–253. [Google Scholar] [CrossRef]
- Mestas, J.; Hughes, C.C. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | IL4 | p(I:C) | LPS (*) | TNF/INF (*) |
|---|---|---|---|---|
| CD274 | 0.68 ± 0.27 | 1.14 ± 0.32 | 2.67 ± 0.13 | 38.57 ± 30.51 |
| HLA-DRB1 | 1.12 ± 0.28 | 2.29 ± 1.09 | 7.03 ± 2.73 | 699.55 ± 187.95 |
| IDO1/2 | 1.31 ± 0.71 | 3.23 ± 1.89 | 35.05 ± 41.07 | 9741.64 ± 5122.44 |
| IP10 | 2.48 ± 3.10 | 1.81 ± 2.44 | 315.23 ± 242.77 | 70,727.20 ± 18,141.92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zubkova, E.; Dergilev, K.; Beloglazova, I.; Kalinin, A.; Guseva, A.; Andreev, A.; Partigulov, S.; Lepilin, M.; Menshikov, M.; Parfyonova, Y. Paracrine Responses of Cardiosphere-Derived Cells to Cytokines and TLR Ligands: A Comparative Analysis. Int. J. Mol. Sci. 2023, 24, 17278. https://doi.org/10.3390/ijms242417278
Zubkova E, Dergilev K, Beloglazova I, Kalinin A, Guseva A, Andreev A, Partigulov S, Lepilin M, Menshikov M, Parfyonova Y. Paracrine Responses of Cardiosphere-Derived Cells to Cytokines and TLR Ligands: A Comparative Analysis. International Journal of Molecular Sciences. 2023; 24(24):17278. https://doi.org/10.3390/ijms242417278
Chicago/Turabian StyleZubkova, Ekaterina, Konstantin Dergilev, Irina Beloglazova, Alexander Kalinin, Alika Guseva, Alexander Andreev, Stanislav Partigulov, Mikhail Lepilin, Mikhail Menshikov, and Yelena Parfyonova. 2023. "Paracrine Responses of Cardiosphere-Derived Cells to Cytokines and TLR Ligands: A Comparative Analysis" International Journal of Molecular Sciences 24, no. 24: 17278. https://doi.org/10.3390/ijms242417278
APA StyleZubkova, E., Dergilev, K., Beloglazova, I., Kalinin, A., Guseva, A., Andreev, A., Partigulov, S., Lepilin, M., Menshikov, M., & Parfyonova, Y. (2023). Paracrine Responses of Cardiosphere-Derived Cells to Cytokines and TLR Ligands: A Comparative Analysis. International Journal of Molecular Sciences, 24(24), 17278. https://doi.org/10.3390/ijms242417278