
Biological Impact of Organic Extracts from Urban-Air Particulate Matter: An In Vitro Study of Cytotoxic and Metabolic Effects in Lung Cells




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
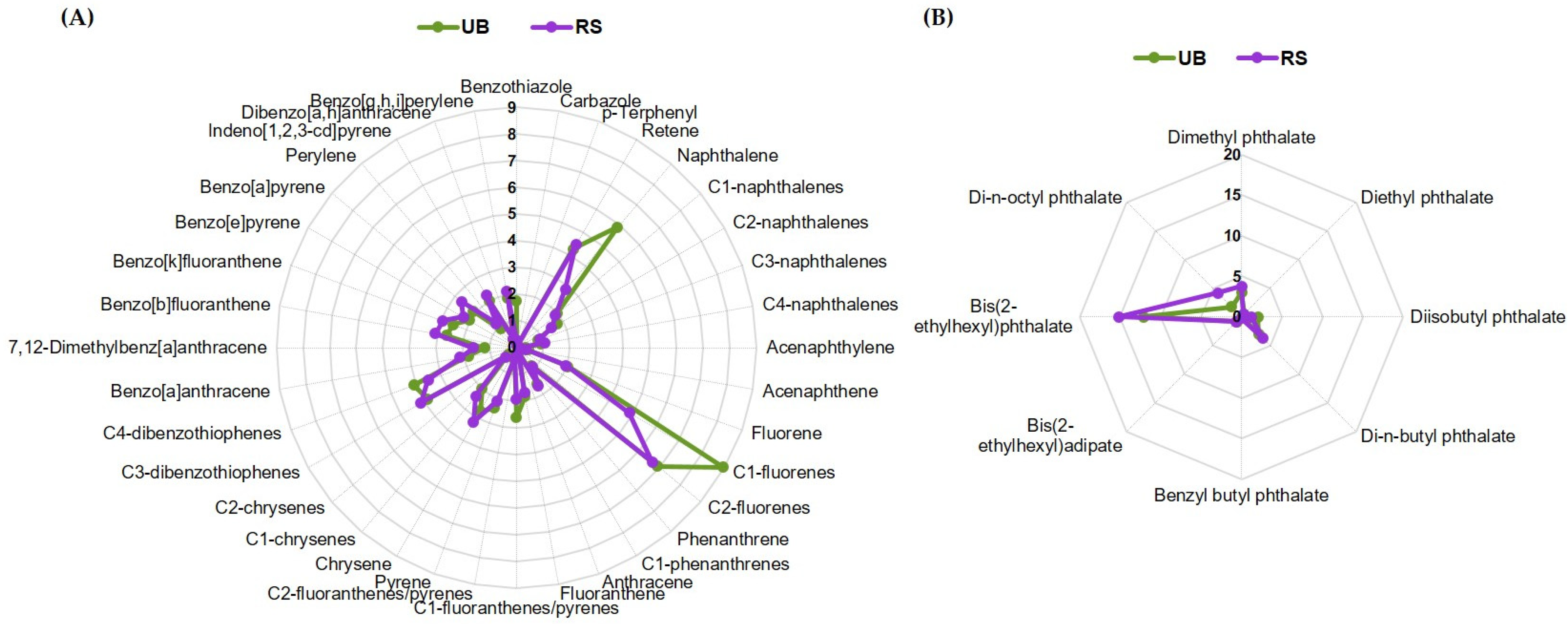
2.1. Composition of PM10 in PAHs and Plasticizers
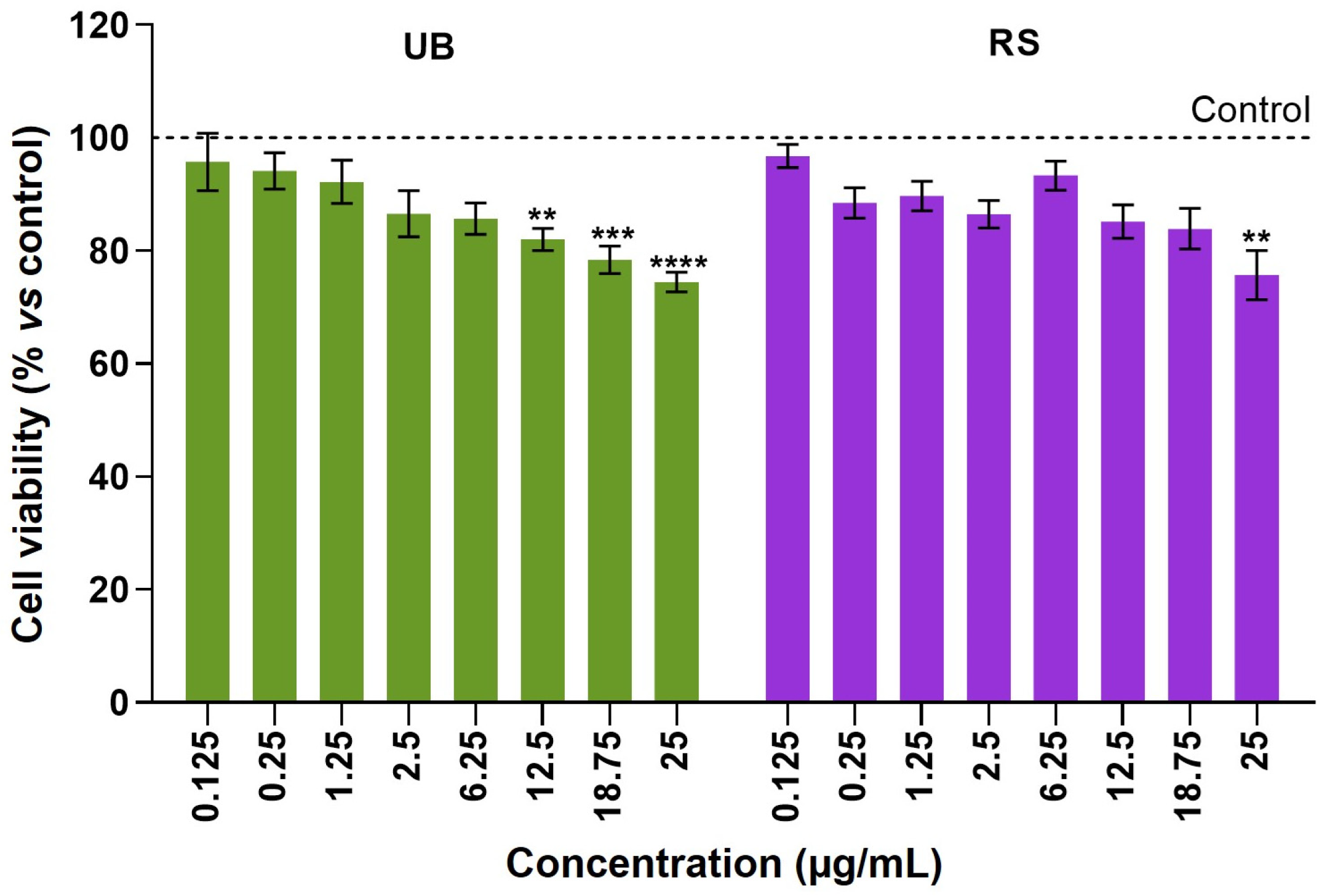
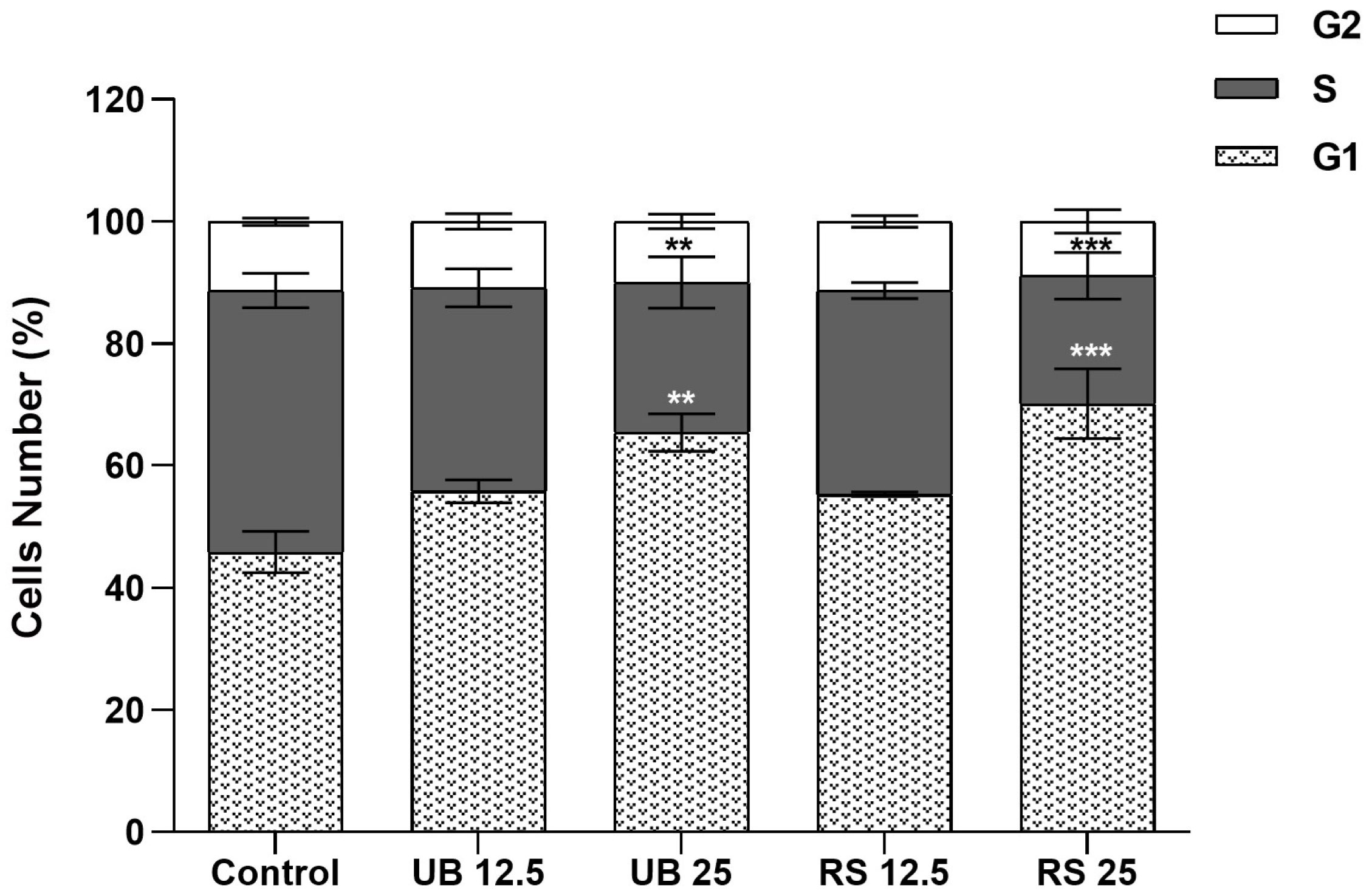
2.2. Cytotoxicity of PM10 Organic Extracts
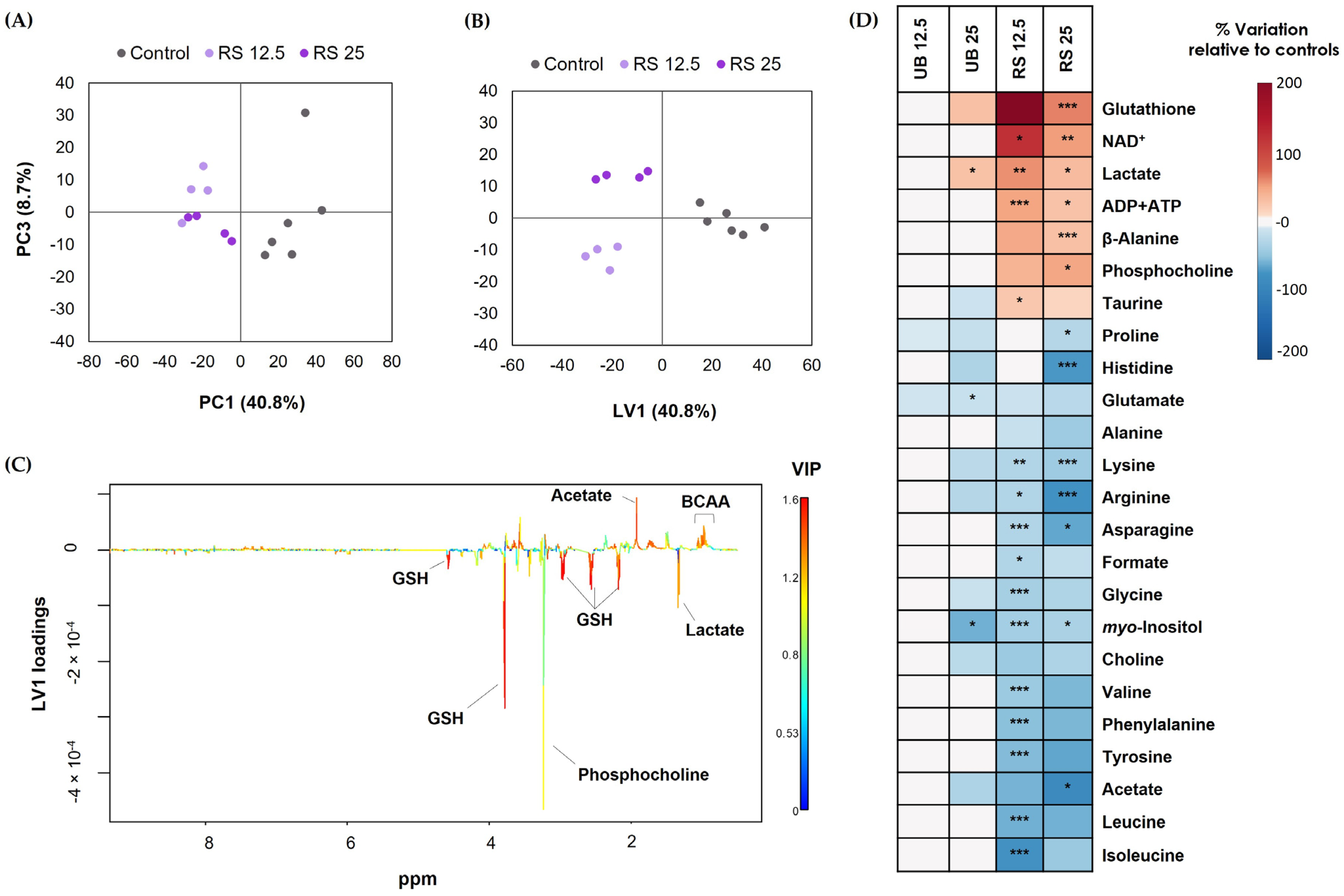
2.3. Impact of PM10 Organic Extracts on the Intracellular Metabolic Profile
3. Materials and Methods
3.1. PM10 Sampling
3.2. Chemical Characterization of PM10 Samples
3.3. PM10 Extraction and Sample Preparation for Biological Assays
3.4. Cell Culture
3.5. Cell Viability Assay
3.6. Quantification of Intracellular ROS
3.7. Mitochondrial Membrane Potential (MMP)
3.8. Cell Cycle Analysis
3.9. NMR Metabolomics
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nazarenko, Y.; Pal, D.; Ariya, P.A. Air quality standards for the concentration of particulate matter 2.5, global descriptive analysis. Bull. World Health Organ. 2021, 99, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Thangavel, P.; Park, D.; Lee, Y.-C. Recent Insights into Particulate Matter (PM2.5)-Mediated Toxicity in Humans: An Overview. Int. J. Environ. Res. Public Health 2022, 19, 7511. [Google Scholar] [CrossRef] [PubMed]
- Visani, G.; Manti, A.; Valentini, L.; Canonico, B.; Loscocco, F.; Isidori, A.; Gabucci, E.; Gobbi, P.; Montanari, S.; Rocchi, M.; et al. Environmental nanoparticles are significantly over-expressed in acute myeloid leukemia. Leuk. Res. 2022, 50, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.; Santa-Helena, E.; De Falco, A.; de Paula Ribeiro, J.; Gioda, A.; Gioda, C.R. Toxicological Effects of Fine Particulate Matter (PM2.5): Health Risks and Associated Systemic Injuries—Systematic Review. Water Air Soil Pollut. 2023, 234, 346. [Google Scholar] [CrossRef] [PubMed]
- Adar, S.D.; Filigrana, P.A.; Clements, N.; Peel, J.L. Ambient Coarse Particulate Matter and Human Health: A Systematic Review and Meta-Analysis. Curr. Environ. Health Rep. 2014, 1, 258–274. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Soukup, J.M.; Sioutas, C.; Cassee, F.R. Response of human alveolar macrophages to ultrafine, fine, and coarse urban air pollution particles. Exp. Lung Res. 2003, 29, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Schins, R.P.F.; Lightbody, J.H.; Borm, P.J.A.; Shi, T.; Donaldson, K.; Stone, V. Inflammatory effects of coarse and fine particulate matter in relation to chemical and biological constituents. Toxicol. Appl. Pharmacol. 2004, 195, 1–11. [Google Scholar] [CrossRef]
- Kyung, S.Y.; Jeong, S.H. Particulate-Matter Related Respiratory Diseases. Tuberc. Respir. Dis. 2020, 83, 116. [Google Scholar] [CrossRef]
- Huang, F.; Pan, B.; Wu, J.; Chen, E.; Chen, L. Relationship between exposure to PM2.5 and lung cancer incidence and mortality: A meta-analysis. Oncotarget 2017, 8, 43322–43331. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Y.; Lin, Z.; Zhou, X.; Chen, T.; He, H.; Huang, H.; Yang, T.; Jiang, Y.; Xu, W.; et al. Mitochondrial OGG1 protects against PM2.5 induced oxidative DNA damage in BEAS-2B cells. Exp. Mol. Pathol. 2015, 99, 365–373. [Google Scholar] [CrossRef]
- Loaiza-Ceballos, M.C.; Marin-Palma, D.; Zapata, W.; Hernandez, J.C. Viral respiratory infections and air pollutants. Air Qual. Atmos. Health 2022, 15, 105–114. [Google Scholar] [CrossRef]
- Ji, J.S.; Liu, L.; Zhang, J.; Kan, H.; Zhao, B.; Burkart, K.G.; Zeng, Y. NO2 and PM2.5 air pollution co-exposure and temperature effect modification on pre-mature mortality in advanced age: A longitudinal cohort study in China. Environ. Health Glob. Access Sci. Source 2022, 21, 97. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, R.; Sera, F.; Vicedo-Cabrera, A.M.; Guo, Y.; Tong, S.; Lavigne, E.; Correa, P.M.; Ortega, N.V.; Achilleos, S.; et al. Interactive effects of ambient fine particulate matter and ozone on daily mortality in 372 cities: Two stage time series analysis. BMJ 2023, 383, e075203. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, H.; Zhang, X.; Xing, W.; Wang, Y.; Bai, P.; Zhang, L.; Hayakawa, K.; Toriba, A.; Tang, N. Exposure to Atmospheric Particulate Matter-Bound Polycyclic Aromatic Hydrocarbons and Their Health Effects: A Review. Int. J. Environ. Res. Public Health 2021, 18, 2177. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Shafy, H.I.; Mansour, M.S.M. A review on polycyclic aromatic hydrocarbons: Source, environmental impact, effect on human health and remediation. Egypt. J. Pet. 2016, 25, 107–123. [Google Scholar] [CrossRef]
- Zhou, Q.; Chen, J.; Zhang, J.; Zhou, F.; Zhao, J.; Wei, X.; Zheng, K.; Wu, J.; Li, B.; Pan, B. Toxicity and endocrine-disrupting potential of PM2.5: Association with particulate polycyclic aromatic hydrocarbons, phthalate esters, and heavy metals. Environ. Pollut. 2022, 292, 118349. [Google Scholar] [CrossRef] [PubMed]
- Hussain, K.; Hoque, R.R.; Balachandran, S.; Medhi, S.; Idris, M.G.; Rahman, M.; Hussain, F.L. Monitoring and Risk Analysis of PAHs in the Environment. In Handbook of Environmental Materials Management; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Jameson, C.W. Polycyclic aromatic hydrocarbons and associated occupational exposures. In Tumour Site Concordance and Mechanisms of Carcinogenesis; Part 1-Concordance between cancer in humans and in experimental animals; Baan, R.A., Stewart, B.W., Straif, K., Eds.; IARC Scientific Publications: Lion, France, 2019; Chapter 7; pp. 59–63. [Google Scholar]
- Luo, K.; Zeng, Y.; Li, M.; Man, Y.; Zeng, L.; Zhang, Q.; Luo, J.; Kang, Y. Inhalation bioacessibility and absorption of polycyclic aromatic hydrocarbons (PAHs) in indoor PM2.5 and its implication in risk assessment. Sci. Total Environ. 2021, 774, 145770. [Google Scholar] [CrossRef] [PubMed]
- Vicente, E.D.; Vicente, A.; Nunes, T.; Calvo, A.; del Blanco-Alegre, C.; Oduber, F.; Castro, A.; Fraile, R.; Amato, F.; Alves, C. Household Dust: Loadings and PM10-Bound Plasticizers and Polycyclic Aromatic Hydrocarbons. Atmosphere 2019, 10, 785. [Google Scholar] [CrossRef]
- Fadel, M.; Ledoux, F.; Afif, C.; Courcot, D. Human health risk assessment for PAHs, phthalates, elements, PCDD/Fs, and DL-PCBs in PM2.5 and for NMVOCs in two East-Mediterranean urban sites under industrial influence. Atmos. Pollut. Res. 2022, 13, 101261. [Google Scholar] [CrossRef]
- Wang, J.; Guinot, B.; Dong, Z.; Li, X.; Xu, H.; Xiao, S.; Ho, S.S.H.; Liu, S.; Cao, J. PM2.5-Bound Polycyclic Aromatic Hydrocarbons (PAHs), Oxygenated-PAHs and Phthalate Esters (PAEs) inside and outside Middle School Classrooms in Xi’an, China: Concentration, Characteristics and Health Risk Assessment. Aerosol Air Qual. Res. 2017, 17, 1811–1824. [Google Scholar] [CrossRef]
- Mathieu-Denoncourt, J.; Wallace, S.J.; de Solla, S.R.; Langlois, V.S. Plasticizer endocrine disruption: Highlighting developmental and reproductive effects in mammals and non-mammalian aquatic species. Gen. Comp. Endocrinol. 2015, 219, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Kay, V.R.; Chambers, C.; Foster, W.G. Reproductive and developmental effects of phthalate diesters in females. Crit. Rev. Toxicol. 2013, 43, 200–219. [Google Scholar] [CrossRef] [PubMed]
- Fandiño-Del-Rio, M.; Matsui, E.C.; Peng, R.D.; Meeker, J.D.; Quirós-Alcalá, L. Phthalate biomarkers and associations with respiratory symptoms and healthcare utilization among low-income urban children with asthma. Environ. Res. 2022, 212, 113239. [Google Scholar] [CrossRef]
- Peixoto, M.S.; de Oliveira Galvão, M.F.; Batistuzzo de Medeiros, S.R. Cell death pathways of particulate matter toxicity. Chemosphere 2017, 188, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Quezada-Maldonado, E.M.; Sánchez-Pérez, Y.; Chirino, Y.I.; García-Cuellar, C.M. Airborne particulate matter induces oxidative damage, DNA adduct formation and alterations in DNA repair pathways. Environ. Pollut. 2021, 287, 117313. [Google Scholar] [CrossRef] [PubMed]
- Leni, Z.; Künzi, L.; Geiser, M. Air pollution causing oxidative stress. Curr. Opin. Toxicol. 2020, 20–21, 1–8. [Google Scholar] [CrossRef]
- Wei, T.; Tang, M. Biological effects of airborne fine particulate matter (PM2.5) exposure on pulmonary immune system. Environ. Toxicol. Pharmacol. 2018, 60, 195–201. [Google Scholar] [CrossRef]
- Vattanasit, U.; Navasumrit, P.; Khadka, M.B.; Kanitwithayanun, J.; Promvijit, J.; Autrup, H.; Ruchirawat, M. Oxidative DNA damage and inflammatory responses in cultured human cells and in humans exposed to traffic-related particles. Int. J. Hyg. Environ. Health 2014, 217, 23–33. [Google Scholar] [CrossRef]
- Silva, T.D.; Alves, C.; Oliveira, H.; Duarte, I.F. Metabolic dysregulations underlying the pulmonary toxicity of atmospheric fine particulate matter: Focus on energy-producing pathways and lipid metabolism. Air Qual. Atmos. Health 2022, 15, 2051–2065. [Google Scholar] [CrossRef]
- Araújo, A.M.; Carvalho, F.; de Pinho, P.; Carvalho, M. Toxicometabolomics: Small Molecules to Answer Big Toxicological Questions. Metabolites 2021, 11, 692. [Google Scholar] [CrossRef]
- Halama, A. Metabolomics in cell culture-A strategy to study crucial metabolic pathways in cancer development and the response to treatment. Arch. Biochem. Biophys. 2014, 564, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Shon, J.C.; Lee, S.M.; Jung, J.H.; Wu, Z.; Kwon, Y.S.; Sim, H.J.; Seo, J.S. Integrated metabolomics and lipidomics reveals high accumulation of polyunsaturated lysoglycerophospholipids in human lung fibroblasts exposed to fine particulate matter. Ecotoxicol. Environ. Saf. 2020, 202, 110896. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Lu, R.; Cui, J.; Sun, H.; Yang, H.; Meng, Q.; Wu, S.; Aschner, M.; Li, X.; Chen, R. Inhibition of ATP citrate lyase (ACLY) protects airway epithelia from PM2.5-induced epithelial-mesenchymal transition. Ecotoxicol. Environ. Saf. 2019, 167, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Zou, Y.; Abbas, A.; Dai, B. Nuclear magnetic resonance-based metabolomic investigation reveals metabolic perturbations in PM2.5-treated A549 cells. Environ. Sci. Pollut. Res. 2018, 25, 31656–31665. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhu, L.; Li, R.; Wang, H.; Cai, Z. Omics approach reveals metabolic disorders associated with the cytotoxicity of airborne particulate matter in human lung carcinoma cells. Environ. Pollut. 2019, 246, 45–52. [Google Scholar] [CrossRef]
- Song, Y.; Li, R.; Zhang, Y.; Wei, J.; Chen, W.; Chung, C.K.A.; Cai, Z. Mass spectrometry-based metabolomics reveals the mechanism of ambient fine particulate matter and its components on energy metabolic reprogramming in BEAS-2B cells. Sci. Total Environ. 2019, 651, 3139–3150. [Google Scholar] [CrossRef]
- Song, X.; Liu, J.; Geng, N.; Shan, Y.; Zhang, B.; Zhao, B.; Ni, Y.; Liang, Z.; Chen, J.; Zhang, L.; et al. Multi-omics analysis to reveal disorders of cell metabolism and integrin signaling pathways induced by PM2.5. J. Hazard. Mater. 2022, 424, 127573. [Google Scholar] [CrossRef]
- Liu, D.; Liu, Y.; Wang, R.; Feng, L.; Xu, L.; Jin, C. Metabolic profiling disturbance of PM2.5 revealed by Raman spectroscopy and mass spectrometry-based nontargeted metabolomics. Environ. Sci. Pollut. Res. Int. 2022, 29, 74500–74511. [Google Scholar] [CrossRef]
- Sánchez-Soberón, F.; Cuykx, M.; Serra, N.; Linares, V.; Bellés, M.; Covaci, A.; Schuhmacher, M. In-vitro metabolomics to evaluate toxicity of particulate matter under environmentally realistic conditions. Chemosphere 2018, 209, 137–146. [Google Scholar] [CrossRef]
- Villasclaras, P.; Jaén, C.; van Drooge, B.L.; Grimalt, J.O.; Tauler, R.; Bedia, C. Phenotypic and Metabolomic Characterization of 3D Lung Cell Cultures Exposed to Airborne Particulate Matter from Three Air Quality Network Stations in Catalonia. Toxics 2022, 10, 632. [Google Scholar] [CrossRef]
- Kang, H.-J.; Lee, S.-Y.; Kwon, J.-H. Physico-chemical properties and toxicity of alkylated polycyclic aromatic hydrocarbons. J. Hazard. Mater. 2016, 312, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Ismail, I.M.I.; Khoder, M.; Shamy, M.; Alghamdi, M.; Costa, M.; Ali, L.N.; Wang, W.; Eqani, S.A.M.A.S. Polycyclic aromatic hydrocarbons (PAHs) in indoor dust samples from Cities of Jeddah and Kuwait: Levels, sources and non-dietary human exposure. Sci. Total Environ. 2016, 573, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.B.; Shaikh, S.; Jain, K.R.; Desai, C.; Madamwar, D. Polycyclic Aromatic Hydrocarbons: Sources, Toxicity, and Remediation Approaches. Front. Microbiol. 2020, 11, 562813. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, D.; Vicente, E.D.; Vicente, A.; Gonçalves, C.; Lopes, I.; Alves, C.A.; Oliveira, H. Toxicological and Mutagenic Effects of Particulate Matter from Domestic Activities. Toxics 2023, 11, 505. [Google Scholar] [CrossRef] [PubMed]
- Vicente, E.D.; Figueiredo, D.; Gonçalves, C.; Vicente, A.; Lopes, I.; Oliveira, H.; Alves, C.A. Cytotoxicity and mutagenicity of particulate matter from the open burning of pruning wastes. Air Qual. Atmos. Health 2022, 15, 299–310. [Google Scholar] [CrossRef]
- Vicente, E.D.; Figueiredo, D.; Gonçalves, C.; Lopes, I.; Oliveira, H.; Kováts, N.; Pinheiro, T.; Alves, C.A. In vitro toxicity of indoor and outdoor PM10 from residential wood combustion. Sci. Total Environ. 2021, 782, 146820. [Google Scholar] [CrossRef] [PubMed]
- Vicente, E.D.; Figueiredo, D.; Gonçalves, C.; Lopes, I.; Oliveira, H.; Kováts, N.; Pinheiro, T.; Alves, C.A. In vitro toxicity of particulate matter emissions from residential pellet combustion. J. Environ. Sci. 2022, 115, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xia, T.; Nel, A.E. The role of oxidative stress in ambient particulate matter-induced lung diseases and its implications in the toxicity of engineered nanoparticles. Free Radic. Biol. Med. 2008, 44, 1689–1699. [Google Scholar] [CrossRef]
- Deng, X.; Rui, W.; Zhang, F.; Ding, W. PM2.5 induces Nrf2-mediated defense mechanisms against oxidative stress by activating PIK3/AKT signaling pathway in human lung alveolar epithelial A549 cells. Cell Biol. Toxicol. 2013, 29, 143–157. [Google Scholar] [CrossRef]
- Zou, Y.; Jin, C.; Su, Y.; Li, J.; Zhu, B. Water soluble and insoluble components of urban PM2.5 and their cytotoxic effects on epithelial cells (A549) in vitro. Environ. Pollut. 2016, 212, 627–635. [Google Scholar] [CrossRef]
- Libalova, H.; Milcova, A.; Cervena, T.; Vrbova, K.; Rossnerova, A.; Novakova, Z.; Topinka, J.; Rossner, P. Kinetics of ROS generation induced by polycyclic aromatic hydrocarbons and organic extracts from ambient air particulate matter in model human lung cell lines. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2018, 827, 50–58. [Google Scholar] [CrossRef]
- De Oliveira Alves, N.; Vessoni, A.T.; Quinet, A.; Fortunato, R.S.; Kajitani, G.S.; Peixoto, M.S.; Hacon, S.D.S.; Artaxo, P.; Saldiva, P.; Menck, C.F.M.; et al. Biomass burning in the Amazon region causes DNA damage and cell death in human lung cells. Sci. Rep. 2017, 7, 10937. [Google Scholar] [CrossRef] [PubMed]
- Hickey, C.; Gordon, C.; Galdanes, K.; Blaustein, M.; Horton, L.; Chillrud, S.; Ross, J.; Yinon, L.; Chen, L.C.; Gordon, T. Toxicity of particles emitted by fireworks. Part. Fibre Toxicol. 2020, 17, 28. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.J.; Cao, Z.M.; Guo, X.W.; Chen, H.J.; Lian, Y.; Zheng, W.J.; Chen, Y.J.; Lian, H.Z.; Hu, X. Cytotoxicity and toxicoproteomic analyses of human lung epithelial cells exposed to extracts of atmospheric particulate matters on PTFE filters using acetone and water. Ecotoxicol. Environ. Saf. 2020, 191, 110223. [Google Scholar] [CrossRef] [PubMed]
- Penning, T.M.; Burczynski, M.E.; Hung, C.F.; McCoull, K.D.; Palackal, N.T.; Tsuruda, L.S. Dihydrodiol dehydrogenases and polycyclic aromatic hydrocarbon activation: Generation of reactive and redox active o-quinones. Chem. Res. Toxicol. 1999, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Reddam, A.; Mclarnan, S.; Kupsco, A. Environmental Chemical Exposures and Mitochondrial Dysfunction: A Review of Recent Literature. Curr. Environ. Health Rep. 2022, 9, 631–649. [Google Scholar] [CrossRef] [PubMed]
- Schins, R.P.F.; Knaapen, A.M. Genotoxicity of poorly soluble particles. Inhal. Toxicol. 2007, 19, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Zárate, E.; Sánchez-Pérez, Y.; Gutiérrez-Ruiz, M.C.; Chirino, Y.I.; Osornio-Vargas, Á.R.; Morales-Bárcenas, R.; Souza-Arroyo, V.; García-Cuellar, C.M. Atmospheric particulate matter (PM10) exposure-induced cell cycle arrest and apoptosis evasion through STAT3 activation via PKCζ and Src kinases in lung cells. Environ. Pollut. 2016, 214, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Zhang, M.; Sun, Y.; Fang, Z.; Wang, H.; Li, S.; Peng, Y.; Li, J.; Li, J.; Tian, J.; et al. Mechanism of PM2.5-induced human bronchial epithelial cell toxicity in central China. J. Hazard. Mater. 2020, 396, 122747. [Google Scholar] [CrossRef]
- Gao, M.; Liang, C.; Hong, W.; Yu, X.; Zhou, Y.; Sun, R.; Li, H.; Huang, H.; Gan, X.; Yuan, Z.; et al. Biomass-related PM2.5 induces mitochondrial fragmentation and dysfunction in human airway epithelial cells. Environ. Pollut. 2022, 292, 118464. [Google Scholar] [CrossRef]
- Leclercq, B.; Kluza, J.; Antherieu, S.; Sotty, J.; Alleman, L.Y.; Perdrix, E.; Loyens, A.; Coddeville, P.; Lo Guidice, J.M.; Marchetti, P.; et al. Air pollution-derived PM2.5 impairs mitochondrial function in healthy and chronic obstructive pulmonary diseased human bronchial epithelial cells. Environ. Pollut. 2018, 243, 1434–1449. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, X.; Li, X.; Lv, S.; Ma, R.; Qi, Y.; Abulikemu, A.; Duan, H.; Guo, C.; Li, Y.; et al. PM2.5 triggered apoptosis in lung epithelial cells through the mitochondrial apoptotic way mediated by a ROS-DRP1-mitochondrial fission axis. J. Hazard. Mater. 2020, 397, 122608. [Google Scholar] [CrossRef] [PubMed]
- Pardo, M.; Xu, F.; Shemesh, M.; Qiu, X.; Barak, Y.; Zhu, T.; Rudich, Y. Nrf2 protects against diverse PM2.5 components-induced mitochondrial oxidative damage in lung cells. Sci. Total Environ. 2019, 669, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Sotty, J.; Kluza, J.; De Sousa, C.; Tardivel, M.; Anthérieu, S.; Alleman, L.Y.; Canivet, L.; Perdrix, E.; Loyens, A.; Marchetti, P.; et al. Mitochondrial alterations triggered by repeated exposure to fine (PM2.5-0.18) and quasi-ultrafine (PM0.18) fractions of ambient particulate matter. Environ. Int. 2020, 142, 105830. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhong, Y.; Hou, T.; Liao, J.; Zhang, C.; Sun, C.; Wang, G. PM2.5 induces EMT and promotes CSC properties by activating Notch pathway in vivo and vitro. Ecotoxicol. Environ. Saf. 2019, 178, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, Y.; Liu, L.; Wang, Q.; Zeng, J.; Chen, C. PM2.5 exposure perturbs lung microbiome and its metabolic profile in mice. Sci. Total Environ. 2020, 721, 137432. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; MacNee, W. Lung glutathione and oxidative stress: Implications in cigarette smoke-induced airway disease. Am. J. Physiol. 1999, 277, 1067–1088. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; St. Croix, C.M.; Sussman, N.; Zhao, Q.; Pitt, B.R.; Kagan, V.E. Contribution of Glutathione and Metallothioneins to Protection against Copper Toxicity and Redox Cycling: Quantitative Analysis Using MT+/+ and MT−/− Mouse Lung Fibroblast Cells. Chem. Res. Toxicol. 2002, 15, 1080–1087. [Google Scholar] [CrossRef]
- Zhao, Y.; Gu, X.; Ma, H.; He, X.; Liu, M.; Ding, Y. Association of Glutathione Level and Cytotoxicity of Gold Nanoparticles in Lung Cancer Cells. J. Phys. Chem. C 2011, 115, 12797–12802. [Google Scholar] [CrossRef]
- Surai, P.F.; Earle-Payne, K.; Kidd, M.T. Taurine as a Natural Antioxidant: From Direct Antioxidant Effects to Protective Action in Various Toxicological Models. Antioxidants 2021, 10, 1876. [Google Scholar] [CrossRef]
- Pio, C.; Rienda, I.C.; Nunes, T.; Gonçalves, C.; Tchepel, O.; Pina, N.K.; Rodrigues, J.; Lucarelli, F.; Alves, C.A. Impact of biomass burning and non-exhaust vehicle emissions on PM10 levels in a mid-size non-industrial western Iberian city. Atmos. Environ. 2022, 289, 119293. [Google Scholar] [CrossRef]
- Pio, C.; Cerqueira, M.; Harrison, R.M.; Nunes, T.; Mirante, F.; Alves, C.; Oliveira, C.; Sanchez de la Campa, A.; Artíñano, B.; Matos, M. OC/EC ratio observations in Europe: Re-thinking the approach for apportionment between primary and secondary organic carbon. Atmos. Environ. 2011, 45, 6121–6132. [Google Scholar] [CrossRef]
- Alves, C.; Rienda, I.C.; Vicente, A.; Vicente, E.; Gonçalves, C.; Candeias, C.; Rocha, F.; Lucarelli, F.; Pazzi, G.; Kováts, N.; et al. Morphological properties, chemical composition, cancer risks and toxicological potential of airborne particles from traffic and urban background sites. Atmos. Res. 2021, 264, 105837. [Google Scholar] [CrossRef]
- Alves, C.; Soares, M.; Figueiredo, D.; Oliveira, H. Effects of particle-bound polycyclic aromatic hydrocarbons from different traffic sources on human alveolar epithelial cell line A549. Atmos. Environ. 2023, 303, 6414. [Google Scholar] [CrossRef]
- Carrola, J.; Bastos, V.; Jarak, I.; Oliveira-Silva, R.; Malheiro, E.; Daniel-da-Silva, A.L.; Oliveira, H.; Santos, C.; Gil, A.M.; Duarte, I.F. Metabolomics of silver nanoparticles toxicity in HaCaT cells: Structure–activity relationships and role of ionic silver and oxidative stress. Nanotoxicology 2016, 10, 1105–1117. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, T.D.; Alves, C.; Oliveira, H.; Duarte, I.F. Biological Impact of Organic Extracts from Urban-Air Particulate Matter: An In Vitro Study of Cytotoxic and Metabolic Effects in Lung Cells. Int. J. Mol. Sci. 2023, 24, 16896. https://doi.org/10.3390/ijms242316896
Silva TD, Alves C, Oliveira H, Duarte IF. Biological Impact of Organic Extracts from Urban-Air Particulate Matter: An In Vitro Study of Cytotoxic and Metabolic Effects in Lung Cells. International Journal of Molecular Sciences. 2023; 24(23):16896. https://doi.org/10.3390/ijms242316896
Chicago/Turabian StyleSilva, Tatiana D., Célia Alves, Helena Oliveira, and Iola F. Duarte. 2023. "Biological Impact of Organic Extracts from Urban-Air Particulate Matter: An In Vitro Study of Cytotoxic and Metabolic Effects in Lung Cells" International Journal of Molecular Sciences 24, no. 23: 16896. https://doi.org/10.3390/ijms242316896
APA StyleSilva, T. D., Alves, C., Oliveira, H., & Duarte, I. F. (2023). Biological Impact of Organic Extracts from Urban-Air Particulate Matter: An In Vitro Study of Cytotoxic and Metabolic Effects in Lung Cells. International Journal of Molecular Sciences, 24(23), 16896. https://doi.org/10.3390/ijms242316896