

Potential Roles of microRNAs for Assessing Cardiovascular Risk in Pre-Eclampsia-Exposed Postpartum Women and Offspring

 , , , and
, , , and
Abstract
:1. Introduction
2. Pathogenesis of Pre-Eclampsia
3. Risk of Cardiovascular Disease in Postpartum Women Exposed to Pre-Eclampsia
4. Risk of Cardiovascular Disease in Offspring Exposed to Pre-Eclampsia
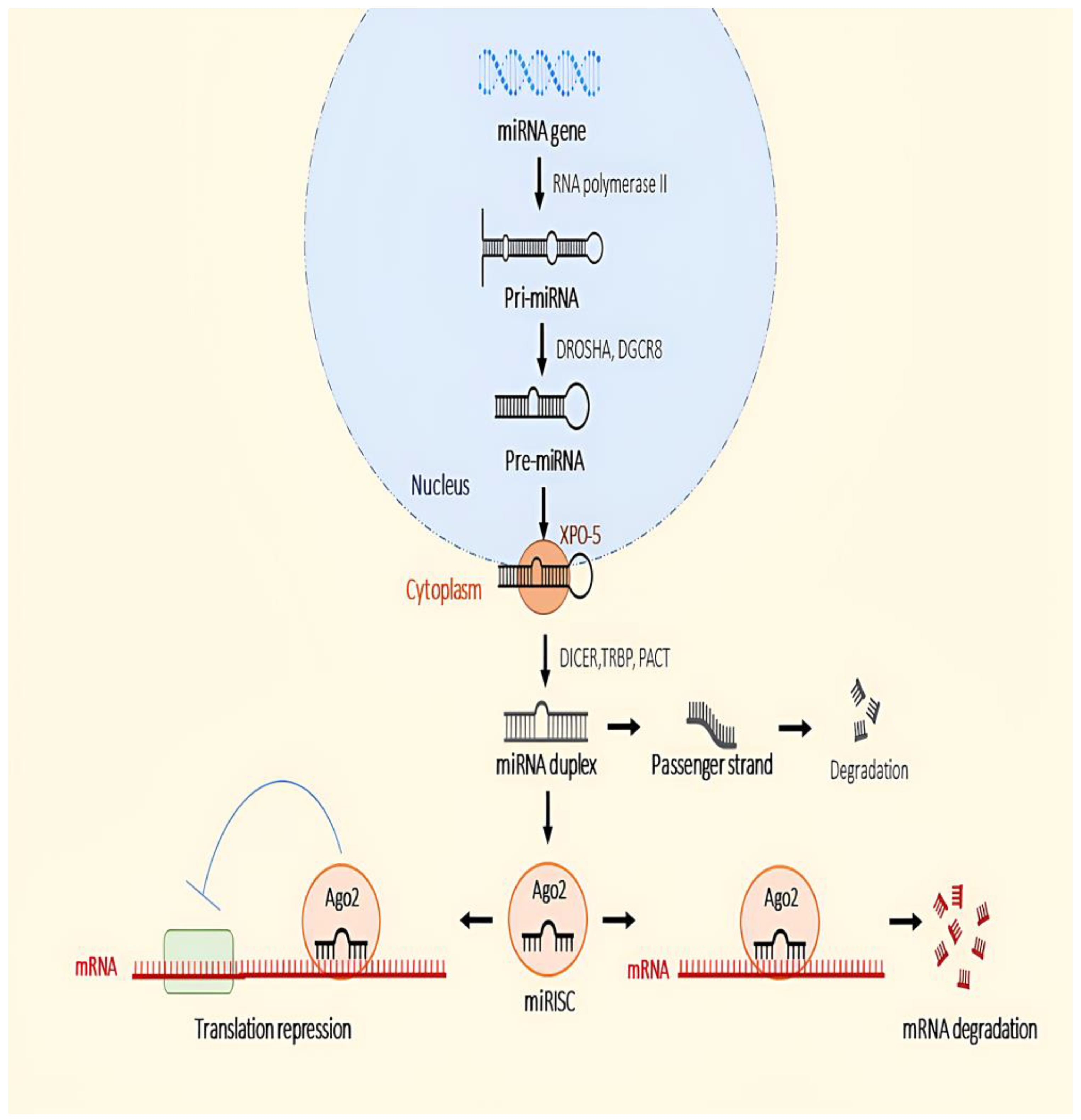
5. microRNA Biogenesis
6. microRNA Dysregulation in Postpartum Women Exposed to Pre-Eclampsia

{kind=link}
| miRNA | Regulation | Role in CVD | Sample | Reference |
|---|---|---|---|---|
| miR-221-3p | Downregulated | Cell proliferation | Plasma | [107] |
| miR-126-3p | Downregulated | Angiogenesis | [110] | |
| miR-146a-5p | Downregulated | Angiogenesis | ||
| miR-122-5p | Downregulated | Fat metabolism | ||
| miR-103a-3p | Downregulated | Anti-apoptosis and autophagy | [117] | |
| miR-1-3p | Upregulated | Action potential regulation | Whole Peripheral Blood | [120,125] |
| miR-23a-3p | Upregulated | Apoptosis, cell regeneration | [120] | |
| miR-499a-5p | Upregulated | Cell proliferation | ||
| miR-17-5p | Upregulated | Apoptosis | [125] | |
| miR-20b-5p | Upregulated | Cell proliferation, migration | ||
| miR-29a-3p | Upregulated | Cell migration | ||
| miR-126-3p | Upregulated | Angiogenesis | ||
| miR-133a-3p | Upregulated | Anti-angiogenesis | ||
| miR-130b-3p | Downregulated | Angiogenesis |
7. microRNA Dysregulation in Offspring Exposed to Pre-Eclampsia
| miRNA | Regulation | Role in CVD | Sample | Reference |
|---|---|---|---|---|
| miR-26a-5p | Downregulated | Apoptosis inhibitor, autophagy regulation | Cord Blood | [133] |
| miR-145-5p | Downregulated | Apoptosis inhibitor | ||
| miR-574-3p | Downregulated | Regulation of mitochondrial function | ||
| miR-195-5p | Downregulated | Apoptosis inhibitor | ||
| miR-199a-5p | Downregulated | Induced cell proliferation, anti-inflammation | ||
| miR-221-3p | Downregulated | Anti-angiogenesis | ||
| miR-92a-3p | Upregulated | Inflammation | ||
| miR-499a-5p | Upregulated | Cell proliferation | Placenta | [138] |
| miR-29a/c-3p | Downregulated | Cell migration | HUVEC | [142] |
| miR-146a | Upregulated | Inflammation | [143] | |
| miR-1270 | Downregulated | Cell migration | EPC | [146] |
| miR-133a-3p | Upregulated | Anti-angiogenesis | Whole Peripheral Blood | [147] |
| miR-1-3p | Upregulated | Action potential regulation | ||
| miR-103a-3p | Upregulated | Apoptosis, autophagy | ||
| miR-20a-5p | Upregulated | Inflammation | ||
| miR-342-3p | Downregulated | Anti-inflammation |
8. Limitations and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Sidek, S.N.; Abdul Ghani, R.B. Knowledge, Attitude and Practices on Maternal Hypertensive Disorder Among Mothers in Kuantan, Pahang, Malaysia. Asian J. Med. Biomed. 2022, 5, 24–35. [Google Scholar] [CrossRef]
- Brown, M.A.; Magee, L.A.; Kenny, L.C.; Karumanchi, S.A.; McCarthy, F.P.; Saito, S.; Hall, D.R.; Warren, C.E.; Adoyi, G.; Ishaku, S. Hypertensive Disorders of Pregnancy. Hypertension 2018, 72, 24–43. [Google Scholar] [CrossRef] [PubMed]
- Lisonkova, S.; Joseph, K.S. Incidence of Preeclampsia: Risk Factors and Outcomes Associated with Early- versus Late-Onset Disease. Am. J. Obstet. Gynecol. 2013, 209, 544.e1–544.e12. [Google Scholar] [CrossRef] [PubMed]
- Abalos, E.; Cuesta, C.; Grosso, A.L.; Chou, D.; Say, L. Global and regional estimates of preeclampsia and eclampsia: A systematic review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Staff, A.C.; Figueiro-Filho, E.; Davidge, S.T. Chapter 19—Long-Term Effects of Preeclampsia on Mothers and Offspring. In Chesley’s Hypertensive Disorders in Pregnancy, 5th ed.; Taylor, R.N., Conrad, K.P., Davidge, S.T., Staff, A.C., Roberts, J.M., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 419–434. [Google Scholar] [CrossRef]
- Tuberville, H.R.; Sasser, J.M. Preeclampsia beyond pregnancy: Long-term consequences for mother and child. Am. J. Physiol.-Ren. Physiol. 2020, 318, 1315–1326. [Google Scholar] [CrossRef]
- Karatza, A.A.; Dimitriou, G. Preeclampsia Emerging as a Novel Risk Factor for Cardiovascular Disease in the Offspring. Curr. Pediatr. Rev. 2020, 16, 194–199. [Google Scholar] [CrossRef]
- Mongraw-Chaffin, M.L.; Cirillo, P.M.; Cohn, B.A. Preeclampsia and Cardiovascular Disease Death. Hypertension 2010, 56, 166–171. [Google Scholar] [CrossRef]
- Cederlöf, E.T.; Lundgren, M.; Lindahl, B.; Christersson, C. Pregnancy Complications and Risk of Cardiovascular Disease Later in Life: A Nationwide Cohort Study. J. Am. Heart Assoc. 2022, 11, e023079. [Google Scholar] [CrossRef]
- Fraser, A.; Nelson, S.M.; Macdonald-Wallis, C.; Sattar, N.; Lawlor, D.A. Hypertensive Disorders of Pregnancy and Cardiometabolic Health in Adolescent Offspring. Hypertension 2013, 62, 614–620. [Google Scholar] [CrossRef]
- Staley, J.R.; Bradley, J.; Silverwood, R.J.; Howe, L.D.; Tilling, K.; Lawlor, D.A.; Macdonald-Wallis, C. Associations of Blood Pressure in Pregnancy With Offspring Blood Pressure Trajectories During Childhood and Adolescence: Findings From a Prospective Study. J. Am. Heart Assoc. 2015, 4, e001422. [Google Scholar] [CrossRef]
- Pogribny, I.P. MicroRNAs as biomarkers for clinical studies. Exp. Biol. Med. 2018, 243, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Saliminejad, K.; Khorshid, H.R.K.; Fard, S.S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol. 2018, 234, 5451–5465. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Y.; Zheng, Y.S.; Li, Z.G.; Cui, Y.M.; Jiang, J.C. MiR-92a contributes to the cardiovascular disease development in diabetes mellitus through NF-κB and downstream inflammatory pathways. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3070–3079. [Google Scholar] [CrossRef] [PubMed]
- Baulina, N.; Pisklova, M.; Kiselev, I.; Chumakova, O.; Zateyshchikov, D.; Favorova, O. Circulating miR-499a-5p Is a Potential Biomarker of MYH7—Associated Hypertrophic Cardiomyopathy. Int. J. Mol. Sci. 2022, 23, 3791. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Y.; Zhao, X.; Liu, Y.-Z.; Meng, Z.; Wang, D.; Yang, F.; Shi, Q.-W. Plasma MicroRNA-126-5p is Associated with the Complexity and Severity of Coronary Artery Disease in Patients with Stable Angina Pectoris. Cell. Physiol. Biochem. 2016, 39, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Saini, V.M.; Liu, K.R.; Surve, A.S.; Gupta, S.; Gupta, A. MicroRNAs as biomarkers for monitoring cardiovascular changes in Type II Diabetes Mellitus (T2DM) and exercise. J. Diabetes Metab. Disord. 2022, 21, 1819–1832. [Google Scholar] [CrossRef] [PubMed]
- Armaly, Z.; Jadaon, J.E.; Jabbour, A.; Abassi, Z.A. Preeclampsia: Novel Mechanisms and Potential Therapeutic Approaches. Front. Physiol. 2018, 9, 973. [Google Scholar] [CrossRef]
- Sircar, M.; Thadhani, R.; Karumanchi, S.A. Pathogenesis of preeclampsia. Curr. Opin. Nephrol. Hypertens. 2015, 24, 131–138. [Google Scholar] [CrossRef]
- Goulopoulou, S. Maternal Vascular Physiology in Preeclampsia. Hypertension 2017, 70, 1066–1073. [Google Scholar] [CrossRef]
- Lyall, F.; Robson, S.C.; Bulmer, J.N. Spiral Artery Remodeling and Trophoblast Invasion in Preeclampsia and Fetal Growth Restriction. Hypertension 2013, 62, 1046–1054. [Google Scholar] [CrossRef]
- Zhou, Y.; Gormley, M.J.; Hunkapiller, N.M.; Kapidzic, M.; Stolyarov, Y.; Feng, V.; Nishida, M.; Drake, P.M.; Bianco, K.; Wang, F.; et al. Reversal of gene dysregulation in cultured cytotrophoblasts reveals possible causes of preeclampsia. J. Clin. Investig. 2013, 123, 2862–2872. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Chaiworapongsa, T. Preeclampsia: A link between trophoblast dysregulation and an antiangiogenic state. J. Clin. Investig. 2013, 123, 2775–2777. [Google Scholar] [CrossRef] [PubMed]
- Pijnenborg, R.; Vercruysse, L.; Hanssens, M. The Uterine Spiral Arteries In Human Pregnancy: Facts and Controversies. Placenta 2006, 27, 939–958. [Google Scholar] [CrossRef]
- Staff, A.C.; Fjeldstad, H.E.; Fosheim, I.K.; Moe, K.; Turowski, G.; Johnsen, G.M.; Alnaes-Katjavivi, P.; Sugulle, M. Failure of physiological transformation and spiral artery atherosis: Their roles in preeclampsia. Am. J. Obstet. Gynecol. 2022, 226, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Hecht, J.L.; Zsengeller, Z.K.; Spiel, M.; Karumanchi, S.A.; Rosen, S. Revisiting decidual vasculopathy. Placenta 2016, 42, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Rana, S.; Karumanchi, S.A. Preeclampsia: The Role of Angiogenic Factors in Its Pathogenesis. Physiology 2009, 24, 147–158. [Google Scholar] [CrossRef]
- Chau, K.; Hennessy, A.; Makris, A. Placental growth factor and pre-eclampsia. J. Hum. Hypertens. 2017, 31, 782–786. [Google Scholar] [CrossRef]
- Zhou, Y.; McMaster, M.; Woo, K.; Janatpour, M.; Perry, J.; Karpanen, T.; Alitalo, K.; Damsky, C.; Fisher, S.J. Vascular Endothelial Growth Factor Ligands and Receptors That Regulate Human Cytotrophoblast Survival Are Dysregulated in Severe Preeclampsia and Hemolysis, Elevated Liver Enzymes, and Low Platelets Syndrome. Am. J. Pathol. 2002, 160, 1405–1423. [Google Scholar] [CrossRef]
- Kendall, R.L.; Thomas, K.A. Inhibition of vascular endothelial cell growth factor activity by an endogenously encoded soluble receptor. Proc. Natl. Acad. Sci. USA 1993, 90, 10705–10709. [Google Scholar] [CrossRef]
- Shibuya, M.; Matsui, H.; Sasagawa, T.; Nagamatsu, T. A simple detection method for the serum sFLT1 protein in preeclampsia. Sci. Rep. 2021, 11, 20613. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, L.; Li, R. Values of serum sFlt-1, PLGF levels, and sFlt-1/PLGF ratio in diagnosis and prognosis evaluation of preeclamptic patients. Clin. Exp. Hypertens. 2020, 42, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Pant, V.; Yadav, B.K.; Sharma, J. A cross sectional study to assess the sFlt-1:PlGF ratio in pregnant women with and without preeclampsia. BMC Pregnancy Childbirth 2019, 19, 266. [Google Scholar] [CrossRef] [PubMed]
- Tannetta, D.S.; Dragovic, R.A.; Gardiner, C.; Redman, C.W.; Sargent, I.L. Characterisation of Syncytiotrophoblast Vesicles in Normal Pregnancy and Pre-Eclampsia: Expression of Flt-1 and Endoglin. PLoS ONE 2013, 8, e56754. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Huang, Y.; Jiang, R.; Teng, Y. Syncytiotrophoblast-derived microparticle shedding in early-onset and late-onset severe pre-eclampsia. Int. J. Gynaecol. Obstet. 2012, 119, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, C.; Tannetta, D.S.; Simms, C.A.; Harrison, P.; Redman, C.W.G.; Sargent, I.L. Syncytiotrophoblast Microvesicles Released from Pre-Eclampsia Placentae Exhibit Increased Tissue Factor Activity. PLoS ONE 2011, 6, e26313. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Han, L.; Huang, P.; Chen, Y.; Wang, Y.; Xue, F. Syncytiotrophoblast-Derived Extracellular Vesicles in Pathophysiology of Preeclampsia. Front. Physiol. 2019, 10, 1236. [Google Scholar] [CrossRef]
- Huppertz, B. The Critical Role of Abnormal Trophoblast Development in the Etiology of Preeclampsia. Curr. Pharm. Biotechnol. 2018, 19, 771–780. [Google Scholar] [CrossRef]
- Staff, A.R.; Redman, C.W.G. The Differences between Early- and Late-Onset Pre-Eclampsia. In Comprehensive Gynecology and Obstetrics; Saito, S., Ed.; Springer: Singapore, 2018; pp. 157–172. [Google Scholar] [CrossRef]
- Kamrani, A.; Alipourfard, I.; Ahmadi-Khiavi, H.; Yousefi, M.; Rostamzadeh, D.; Izadi, M.; Ahmadi, M. The role of epigenetic changes in preeclampsia. BioFactors 2019, 45, 712–724. [Google Scholar] [CrossRef]
- Melchiorre, K.; Sutherland, G.R.; Liberati, M.; Thilaganathan, B. Preeclampsia Is Associated With Persistent Postpartum Cardiovascular Impairment. Hypertension 2011, 58, 709–715. [Google Scholar] [CrossRef]
- Honigberg, M.C.; Zekavat, S.M.; Aragam, K.; Klarin, D.; Bhatt, D.L.; Scott, N.S.; Peloso, G.M.; Natarajan, P. Long-Term Cardiovascular Risk in Women With Hypertension During Pregnancy. J. Am. Coll. Cardiol. 2019, 74, 2743–2754. [Google Scholar] [CrossRef]
- Leon, L.J.; McCarthy, F.P.; Direk, K.; Gonzalez-Izquierdo, A.; Prieto-Merino, D.; Casas, J.P.; Chappell, L. Preeclampsia and Cardiovascular Disease in a Large UK Pregnancy Cohort of Linked Electronic Health Records. Circulation 2019, 140, 1050–1060. [Google Scholar] [CrossRef] [PubMed]
- Bellamy, L.; Casas, J.-P.; Hingorani, A.D.; Williams, D.J. Pre-eclampsia and risk of cardiovascular disease and cancer in later life: Systematic review and meta-analysis. BMJ 2007, 335, 974. [Google Scholar] [CrossRef] [PubMed]
- Powe, C.E.; Levine, R.J.; Karumanchi, S.A. Preeclampsia, a Disease of the Maternal Endothelium. Circulation 2011, 123, 2856–2869. [Google Scholar] [CrossRef] [PubMed]
- Sattar, N.; Ramsay, J.; Crawford, L.; Cheyne, H.; Greer, I.A. Classic and Novel Risk Factor Parameters in Women with a History of Preeclampsia. Hypertension 2003, 42, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.R.; Karumanchi, S.A.; Brown, N.J.; Royle, C.M.; Mcelrath, T.F.; Seely, E.W. Increased Sensitivity to Angiotensin II Is Present Postpartum in Women With a History of Hypertensive Pregnancy. Hypertension 2010, 55, 1239–1245. [Google Scholar] [CrossRef]
- Liao, L.; Zhao, X.; Zhou, M.; Deng, Y.; Li, Y.; Peng, C. sFlt-1: A Double Regulator in Angiogenesis-related Diseases. Curr. Pharm. Des. 2021, 27, 4160–4170. [Google Scholar] [CrossRef]
- Burke, S.D.; Zsengellér, Z.K.; Khankin, E.V.; Lo, A.S.; Rajakumar, A.; DuPont, J.J.; McCurley, A.; Moss, M.E.; Zhang, D.; Clark, C.D.; et al. Soluble fms-like tyrosine kinase 1 promotes angiotensin II sensitivity in preeclampsia. J. Clin. Investig. 2016, 126, 2561–2574. [Google Scholar] [CrossRef]
- Garrido-Gimenez, C.; Mendoza, M.; Cruz-Lemini, M.; Galian-Gay, L.; Sanchez-Garcia, O.; Granato, C.; Rodriguez-Sureda, V.; Rodriguez-Palomares, J.; Carreras-Moratonas, E.; Cabero-Roura, L.; et al. Angiogenic Factors and Long-Term Cardiovascular Risk in Women That Developed Preeclampsia During Pregnancy. Hypertension 2020, 76, 1808–1816. [Google Scholar] [CrossRef]
- Hammadah, M.; Georgiopoulou, V.V.; Kalogeropoulos, A.P.; Weber, M.; Wang, X.; Samara, M.A.; Wu, Y.; Butler, J.; Tang, W.W. Elevated Soluble Fms-Like Tyrosine Kinase-1 and Placental-Like Growth Factor Levels Are Associated with Development and Mortality Risk in Heart Failure. Circ. Heart Fail. 2016, 9, e002115. [Google Scholar] [CrossRef]
- Mauricio, R.; Singh, K.; Sanghavi, M.; Ayers, C.R.; Rohatgi, A.; Vongpatanasin, W.; de Lemos, J.A.; Khera, A. Soluble Fms-like tyrosine kinase-1 (sFlt-1) is associated with subclinical and clinical atherosclerotic cardiovascular disease: The Dallas Heart Study. Atherosclerosis 2022, 346, 46–52. [Google Scholar] [CrossRef]
- Gruson, D.; Hermans, M.P.; Ferracin, B.; Ahn, S.A.; Rousseau, M.F. Sflt-1 in heart failure: Relation with disease severity and biomarkers. Scandinavian J. Clin. Lab. Investig. 2016, 76, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, A.B.; Rolo, L.C.; Nardozza, L.M.M.; Araujo Júnior, E. Chapter 6—Epigenetics and Preeclampsia: Programming of Future Outcomes. Preeclampsia 2017, 1710, 73–83. [Google Scholar] [CrossRef]
- Davies, E.L.; Bell, J.S.; Bhattacharya, S. Preeclampsia and preterm delivery: A population-based case–control study. Hypertens. Pregnancy 2016, 35, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Kerkhof, G.F.; Breukhoven, P.E.; Leunissen, R.W.J.; Willemsen, R.H.; Hokken-Koelega, A.C.S. Does Preterm Birth Influence Cardiovascular Risk in Early Adulthood? J. Pediatr. 2012, 161, 390–396.e1. [Google Scholar] [CrossRef] [PubMed]
- Alsnes, I.V.; Vatten, L.J.; Fraser, A.; Bjørngaard, J.H.; Rich-Edwards, J.; Romundstad, P.R.; Åsvold, B.O. Hypertension in Pregnancy and Offspring Cardiovascular Risk in Young Adulthood. Hypertension 2017, 69, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Espliguero, R.; Viana-Llamas, M.C.; Silva-Obregón, J.A.; Avanzas, P. The Role of C-reactive Protein in Patient Risk Stratification and Treatment. Eur. Cardiol. 2021, 16, e28. [Google Scholar] [CrossRef] [PubMed]
- Tajfard, M.; Sany, S.B.T.; Avan, A.; Latiff, L.A.; Rahimi, H.R.; Moohebati, M.; Hasanzadeh, M.; Ghazizadeh, H.; Esmaeily, H.; Doosti, H.; et al. Relationship between serum high sensitivity C-reactive protein with angiographic severity of coronary artery disease and traditional cardiovascular risk factors. J. Cell. Physiol. 2018, 234, 10289–10299. [Google Scholar] [CrossRef]
- Lewandowski, A.J.; Davis, E.F.; Yu, G.; Digby, J.E.; Boardman, H.; Whitworth, P.; Singhal, A.; Lucas, A.; McCormick, K.; Shore, A.C.; et al. Elevated Blood Pressure in Preterm-Born Offspring Associates With a Distinct Antiangiogenic State and Microvascular Abnormalities in Adult Life. Hypertension 2015, 65, 607–614. [Google Scholar] [CrossRef]
- Akçakuş, M.; Altunay, L.; Yıkılmaz, A.; Yazıcı, C.; Köklü, E. The Relationship between Abdominal Aortic Intima-Media Thickness and Lipid Profile in Neonates Born to Mothers with Preeclampsia. J. Pediatr. Endocrinol. Metab. 2010, 23, 1143–1149. [Google Scholar] [CrossRef]
- Alsnes, I.V.; Janszky, I.; Forman, M.R.; Vatten, L.J.; Økland, I. A population-based study of associations between preeclampsia and later cardiovascular risk factors. Am. J. Obstet. Gynecol. 2014, 211, 657.e1–657.e7. [Google Scholar] [CrossRef]
- Kosmas, C.E.; Martinez, I.; Sourlas, A.; Bouza, K.V.; Campos, F.N.; Torres, V.; Montan, P.D.; Guzman, E. High-density lipoprotein (HDL) functionality and its relevance to atherosclerotic cardiovascular disease. Drugs Context 2018, 7, 212525. [Google Scholar] [CrossRef] [PubMed]
- Lazdam, M.; de la Horra, A.; Pitcher, A.; Mannie, Z.; Diesch, J.; Trevitt, C.; Kylintireas, I.; Contractor, H.; Singhal, A.; Lucas, A.; et al. Elevated Blood Pressure in Offspring Born Premature to Hypertensive Pregnancy. Hypertension 2010, 56, 159–165. [Google Scholar] [CrossRef]
- Timpka, S.; Macdonald-Wallis, C.; Hughes, A.D.; Chaturvedi, N.; Franks, P.W.; Lawlor, D.A.; Fraser, A. Hypertensive Disorders of Pregnancy and Offspring Cardiac Structure and Function in Adolescence. J. Am. Heart Assoc. 2016, 5, e003906. [Google Scholar] [CrossRef] [PubMed]
- Hashem, M.; Kalashyan, H.; Choy, J.; Chiew, S.K.; Shawki, A.H.; Dawood, A.H.; Becher, H. Left Ventricular Relative Wall Thickness Versus Left Ventricular Mass Index in Non-Cardioembolic Stroke Patients. Medicine 2015, 94, e872. [Google Scholar] [CrossRef] [PubMed]
- Yun, M.; Li, S.; Yan, Y.; Sun, D.; Guo, Y.; Fernandez, C.; Bazzano, L.; He, J.; Zhang, T.; Chen, W. Blood Pressure and Left Ventricular Geometric Changes: A Directionality Analysis. Hypertension 2021, 78, 1259–1266. [Google Scholar] [CrossRef]
- Vrablik, M.; Dlouha, D.; Todorovova, V.; Stefler, D.; Hubacek, J.A. Genetics of Cardiovascular Disease: How Far Are We from Personalized CVD Risk Prediction and Management? Int. J. Mol. Sci. 2021, 22, 4182. [Google Scholar] [CrossRef]
- Ward-Caviness, C.K.; Neas, L.M.; Blach, C.; Haynes, C.S.; LaRocque-Abramson, K.; Grass, E.; Dowdy, Z.E.; Devlin, R.B.; Diaz-Sanchez, D.; Cascio, W.E.; et al. A genome-wide trans-ethnic interaction study links the PIGR-FCAMR locus to coronary atherosclerosis via interactions between genetic variants and residential exposure to traffic. PLoS ONE 2017, 12, e0173880. [Google Scholar] [CrossRef]
- Khera, A.V.; Emdin, C.A.; Drake, I.; Natarajan, P.; Bick, A.G.; Cook, N.R.; Chasman, D.I.; Baber, U.; Mehran, R.; Rader, D.J.; et al. Genetic Risk, Adherence to a Healthy Lifestyle, and Coronary Disease. N. Engl. J. Med. 2016, 375, 2349–2358. [Google Scholar] [CrossRef]
- Cruz-Lemini, M.; Crispi, F.; Valenzuela-Alcaraz, B.; Figueras, F.; Sitges, M.; Bijnens, B.; Gratacós, E. Fetal cardiovascular remodeling persists at 6 months in infants with intrauterine growth restriction. Ultrasound Obstet. Gynecol. 2019, 48, 349–356. [Google Scholar] [CrossRef]
- Tzafrir, T.; Wainstock, T.; Sheiner, E.; Miodownik, S.; Pariente, G. Fetal Growth Restriction and Long-Term Cardiovascular Morbidity of Offspring in Dichorionic–Diamniotic Twin Pregnancies. J. Clin. Med. 2023, 12, 1628. [Google Scholar] [CrossRef]
- Tüfekci, K.U.; Meuwissen, R.L.J.; Genç, Ş. The Role of MicroRNAs in Biological Processes. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2014; pp. 15–31. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-K.; Kim, B.; Kim, V.N. Re-evaluation of the roles of DROSHA, Exportin 5, and DICER in microRNA biogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, E1881–E1889. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xiang, Y.; Chen, S.; Shi, M.; Jiang, X.; He, Z.; Gao, S. Mechanisms of MicroRNA Biogenesis and Stability Control in Plants. Front. Plant Sci. 2022, 13, 844149. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Ayub, H.; Khan, T.; Wahid, F. MicroRNA biogenesis, gene silencing mechanisms and role in breast, ovarian and prostate cancer. Biochimie 2019, 167, 12–24. [Google Scholar] [CrossRef]
- Annese, T.; Tamma, R.; De Giorgis, M.; Ribatti, D. microRNAs Biogenesis, Functions and Role in Tumor Angiogenesis. Front. Oncol. 2020, 10, 581007. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Okada, C.; Yamashita, E.; Lee, S.J.; Shibata, S.; Katahira, J.; Nakagawa, A.; Yoneda, Y.; Tsukihara, T. A High-Resolution Structure of the Pre-microRNA Nuclear Export Machinery. Science 2009, 326, 1275–1279. [Google Scholar] [CrossRef]
- Zeng, Y.; Cullen, B.R. Structural Requirements for Pre-microrna Binding and Nuclear Export by Exportin 5. Nucleic Acids Res. 2004, 32, 4776–4785. [Google Scholar] [CrossRef]
- Cullen, B.R. Transcription and Processing of Human microRNA Precursors. Mol. Cell 2004, 16, 861–865. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, X.; Graves, P.; Zeng, Y. A comprehensive analysis of precursor microRNA cleavage by human Dicer. RNA 2012, 18, 2083–2092. [Google Scholar] [CrossRef]
- Lee, Y.; Hur, I.; Park, S.-Y.; Kim, Y.-K.; Suh, M.R.; Kim, V.N. The role of PACT in the RNA silencing pathway. EMBO J. 2006, 25, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Fareh, M.; Yeom, K.-H.; Haagsma, A.C.; Chauhan, S.; Heo, I.; Joo, C. TRBP ensures efficient Dicer processing of precursor microRNA in RNA-crowded environments. Nat. Commun. 2016, 7, 13694. [Google Scholar] [CrossRef]
- Medley, J.C.; Panzade, G.; Zinovyeva, A.Y. microRNA strand selection: Unwinding the rules. WIREs RNA 2021, 12, e1627. [Google Scholar] [CrossRef] [PubMed]
- Suster, I.; Feng, Y. Multifaceted Regulation of MicroRNA Biogenesis: Essential Roles and Functional Integration in Neuronal and Glial Development. Int. J. Mol. Sci. 2021, 22, 6765. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Asano, Y.; Ui-Tei, K. Modulation of MicroRNA Processing by Dicer via Its Associated dsRNA Binding Proteins. Non-Coding RNA 2021, 7, 57. [Google Scholar] [CrossRef] [PubMed]
- Daugaard, I.; Hansen, T.B. Biogenesis and Function of Ago-Associated RNAs. Trends Genet. 2017, 33, 208–219. [Google Scholar] [CrossRef]
- Abdelfattah, A.M.; Park, C.; Choi, M.Y. Update on non-canonical microRNAs. Biomol. Concepts 2014, 5, 275–287. [Google Scholar] [CrossRef]
- Miyoshi, K.; Miyoshi, T.; Siomi, H. Many ways to generate microRNA-like small RNAs: Non-canonical pathways for microRNA production. Mol. Genet. Genom. 2010, 284, 95–103. [Google Scholar] [CrossRef]
- Westholm, J.O.; Lai, E.C. Mirtrons: microRNA biogenesis via splicing. Biochimie 2011, 93, 1897–1904. [Google Scholar] [CrossRef]
- Okamura, K.; Hagen, J.W.; Duan, H.; Tyler, D.M.; Lai, E.C. The Mirtron Pathway Generates microRNA-Class Regulatory RNAs in Drosophila. Cell 2007, 130, 89–100. [Google Scholar] [CrossRef]
- Wajahat, M.; Bracken, C.P.; Orang, A. Emerging Functions for snoRNAs and snoRNA-Derived Fragments. Int. J. Mol. Sci. 2021, 22, 10193. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.; Patkaniowska, A.; Urlaub, H.; Lührmann, R.; Tuschl, T. Single-Stranded Antisense siRNAs Guide Target RNA Cleavage in RNAi. Cell 2002, 110, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Babiarz, J.E.; Ruby, J.G.; Wang, Y.; Bartel, D.P.; Blelloch, R. Mouse ES cells express endogenous shRNAs, siRNAs, and other Microprocessor-independent, Dicer-dependent small RNAs. Genes Dev. 2008, 22, 2773–2785. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xu, Z.; Sheng, J. tRNA-Derived Small RNA: A Novel Regulatory Small Non-Coding RNA. Genes 2018, 9, 246. [Google Scholar] [CrossRef]
- Stavast, C.; Erkeland, S. The Non-Canonical Aspects of MicroRNAs: Many Roads to Gene Regulation. Cells 2019, 8, 1465. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, Y.; Li, S.; Zhang, J.; Jiang, D.; Li, X.; Li, Y.; Du, J. A Machine Learning-Based Prediction Model for Cardiovascular Risk in Women With Preeclampsia. Front. Cardiovasc. Med. 2021, 8, 736491. [Google Scholar] [CrossRef]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef]
- Riolo, G.; Cantara, S.; Marzocchi, C.; Ricci, C. miRNA Targets: From Prediction Tools to Experimental Validation. Methods Protoc. 2020, 4, 1. [Google Scholar] [CrossRef]
- Khan, I.A.; Rashid, S.; Singh, N.; Rashid, S.; Singh, V.; Gunjan, D.; Das, P.; Dash, N.R.; Pandey, R.M.; Chauhan, S.S.; et al. Panel of serum miRNAs as potential non-invasive biomarkers for pancreatic ductal adenocarcinoma. Sci. Rep. 2021, 11, 2824. [Google Scholar] [CrossRef]
- Zhai, T.-Y.; Dou, M.; Ma, Y.-B.; Wang, H.; Liu, F.; Zhang, L.-D.; Chong, T.; Wang, Z.-M.; Xue, L. miR-20b-5p is a novel biomarker for detecting prostate cancer. Oncol. Lett. 2022, 24, 426. [Google Scholar] [CrossRef]
- Yang, Q.; Zhao, Q.; Yin, Y. miR 133b is a potential diagnostic biomarker for Alzheimer’s disease and has a neuroprotective role. Exp. Ther. Med. 2019, 18, 2711–2718. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Meng, T.; Chen, J.; Kang, J.; Li, X.; Yu, G.; Tian, L.; Jin, Z.; Dong, H.; Zhang, X.; et al. miR-21a-5p Promotes Inflammation following Traumatic Spinal Cord Injury through Upregulation of Neurotoxic Reactive Astrocyte (A1) Polarization by Inhibiting the CNTF/STAT3/Nkrf Pathway. Int. J. Biol. Sci. 2021, 17, 2795–2810. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhang, Z.; Chen, L.; Zhang, J. miRNA 101 3p.1 as an independent diagnostic biomarker aggravates chronic obstructive pulmonary disease via activation of the EGFR/PI3K/AKT signaling pathway. Mol. Med. Rep. 2019, 20, 4293–4302. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.S.Q.; Casselman, R.; Tayade, C.; Smith, G.N. Differential expression of plasma microRNA in preeclamptic patients at delivery and 1 year postpartum. Am. J. Obstet. Gyneology 2015, 213, 367. [Google Scholar] [CrossRef] [PubMed]
- Verjans, R.; Peters, T.; Beaumont, F.J.; van Leeuwen, R.; van Herwaarden, T.; Verhesen, W.; Munts, C.; Bijnen, M.; Henkens, M.; Diez, J.; et al. MicroRNA-221/222 Family Counteracts Myocardial Fibrosis in Pressure Overload–Induced Heart Failure. Hypertension 2018, 71, 280–288. [Google Scholar] [CrossRef]
- Dentelli, P.; Rosso, A.; Orso, F.; Olgasi, C.; Taverna, D.; Brizzi, M.F. microRNA-222 Controls Neovascularization by Regulating Signal Transducer and Activator of Transcription 5A Expression. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1562–1568. [Google Scholar] [CrossRef]
- Dayan, N.; Schlosser, K.; Stewart, D.J.; Delles, C.; Kaur, A.; Pilote, L. Circulating MicroRNAs Implicate Multiple Atherogenic Abnormalities in the Long-Term Cardiovascular Sequelae of Preeclampsia. Am. J. Hypertens. 2018, 31, 1093–1097. [Google Scholar] [CrossRef]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The Endothelial-Specific MicroRNA miR-126 Governs Vascular Integrity and Angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef]
- Potus, F.; Ruffenach, G.; Dahou, A.; Thebault, C.; Breuils-Bonnet, S.; Tremblay, È.; Nadeau, V.; Paradis, R.; Graydon, C.; Wong, R.; et al. Downregulation of MicroRNA-126 Contributes to the Failing Right Ventricle in Pulmonary Arterial Hypertension. Circulation 2015, 132, 932–943. [Google Scholar] [CrossRef]
- Rawal, S.; Munasinghe, P.E.; Shindikar, A.; Paulin, J.M.; Cameron, V.A.; Manning, P.J.; Williams, M.J.A.; Jones, G.T.; Bunton, R.; Galvin, I.F.; et al. Down-regulation of proangiogenic microRNA-126 and microRNA-132 are early modulators of diabetic cardiac microangiopathy. Cardiovasc. Res. 2017, 113, 90–101. [Google Scholar] [CrossRef]
- Shen, L.; Li, C.; Zhang, H.; Qiu, S.; Fu, T.; Xu, Y. Downregulation of miR-146a Contributes to Cardiac Dysfunction Induced by the Tyrosine Kinase Inhibitor Sunitinib. Front. Pharmacol. 2019, 10, 914. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Rizvi, A.; Pradhan, A.; Perrone, M.A.; Ali, W. Circulating microRNA-126 &122 in patients with coronary artery disease: Correlation with small dense LDL. Prostaglandins Other Lipid Mediat. 2021, 153, 106536. [Google Scholar] [CrossRef] [PubMed]
- Laffont, B.; Rayner, K.J. MicroRNAs in the Pathobiology and Therapy of Atherosclerosis. Can. J. Cardiol. 2017, 33, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Abad, F. Changes of Blood Biomarkers Following Pre-eclampsia and Predictors of Future Cardiovascular Risk. Ph.D. Thesis, University of Southampton, Southampton, UK, 2020. [Google Scholar]
- Zhang, C.; Zhang, C.; Wang, H.; Qi, Y.; Kan, Y.; Ge, Z. Effects of miR 103a 3p on the autophagy and apoptosis of cardiomyocytes by regulating Atg5. Int. J. Mol. Med. 2019, 43, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Xu, J.; Bai, Y.; Zhang, H.; Zhou, W.; Cheng, M.; Zhang, D.; Zhang, L.; Zhang, S. microRNA 103a regulates the calcification of vascular smooth muscle cells by targeting runt related transcription factor 2 in high phosphorus conditions. Exp. Ther. Med. 2021, 22, 1036. [Google Scholar] [CrossRef]
- Hromadnikova, I.; Kotlabova, K.; Dvorakova, L.; Krofta, L. Evaluation of Vascular Endothelial Function in Young and Middle-Aged Women with Respect to a History of Pregnancy, Pregnancy-Related Complications, Classical Cardiovascular Risk Factors, and Epigenetics. Int. J. Mol. Sci. 2020, 21, 430. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, M.; Wen, H.; Hu, C.; Liu, R.; Wei, H.; Ai, C.; Wang, G.; Liao, X.; Li, X. Comparing the diagnostic values of circulating microRNAs and cardiac troponin T in patients with acute myocardial infarction. Clinics 2013, 68, 75–80. [Google Scholar] [CrossRef]
- Terentyev, D.; Belevych, A.E.; Terentyeva, R.; Martin, M.M.; Malana, G.E.; Kuhn, D.E.; Abdellatif, M.; Feldman, D.S.; Elton, T.S.; Gyorke, S.; et al. miR-1 Overexpression Enhances Ca 2+ Release and Promotes Cardiac Arrhythmogenesis by Targeting PP2A Regulatory Subunit B56α and Causing CaMKII-Dependent Hyperphosphorylation of RyR2. Circ. Res. 2009, 104, 514–521. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. Cardiac-specific miRNA in Cardiogenesis, Heart Function, and Cardiac Pathology (with Focus on Myocardial Infarction). J. Mol. Cell. Cardiol. 2016, 94, 107–121. [Google Scholar] [CrossRef]
- Wang, S.; He, W.; Wang, C. MiR-23a Regulates the Vasculogenesis of Coronary Artery Disease by Targeting Epidermal Growth Factor Receptor. Cardiovasc. Ther. 2016, 34, 199–208. [Google Scholar] [CrossRef]
- Hromadníková, I.; Dvořáková, L.; Krofta, L. Postpartum Profiling of microRNAs Involved in Pathogenesis of Cardiovascular/Cerebrovascular Diseases in Women Exposed to Pregnancy-related Complications. Int. J. Cardiol. 2019, 291, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Fan, T.; Hu, Q.; Xu, W.; Yang, J.; Xu, C.; Zhang, B.; Chen, J.; Jiang, H. Downregulation of microRNA-17-5p improves cardiac function after myocardial infarction via attenuation of apoptosis in endothelial cells. Mol. Genet. Genom. 2018, 293, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Jiang, S.; Wu, N.; Shi, E.; Yang, L.; Li, Q. MiR-17-5p-mediated Endoplasmic Reticulum Stress Promotes Acute Myocardial Ischemia Injury Through Targeting Tsg101. Cell Stress Chaperones 2020, 26, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Cheng, R.; Liang, C.; Yao, Y.; Zhang, W.H.; Zhang, J.; Zhang, M.; Li, B.Y.; Xu, C.; Zhang, R. MicroRNA-20b Promotes Cardiac Hypertrophy by the Inhibition of Mitofusin 2-Mediated Inter-organelle Ca2+ Cross-Talk. Mol. Ther. Nucleic Acids 2020, 19, 1343–1356. [Google Scholar] [CrossRef]
- Martinez-Arroyo, O.; Ortega, A.; Flores-Chova, A.; Sanchez-Garcia, B.; Garcia-Garcia, A.B.; Chaves, F.J.; Martin-Escudero, J.C.; Forner, M.J.; Redon, J.; Cortes, R. High miR-126-3p levels associated with cardiovascular events in a general population. Eur. J. Intern. Med. 2023, 113, 49–56. [Google Scholar] [CrossRef]
- Yan, W.; Wang, Y.; Chen, Y.; Guo, Y.; Li, Q.; Wei, X. Exosomal miR-130b-3p Promotes Progression and Tubular Formation Through Targeting PTEN in Oral Squamous Cell Carcinoma. Front. Cell Dev. Biol. 2021, 9, 616306. [Google Scholar] [CrossRef]
- Oudit, G.Y.; Kassiri, Z.; Zhou, J.; Liu, Q.C.; Liu, P.P.; Backx, P.H.; Dawood, F.; Crackower, M.A.; Scholey, J.W.; Penninger, J.M. Loss of PTEN attenuates the development of pathological hypertrophy and heart failure in response to biomechanical stress. Cardiovasc. Res. 2008, 78, 505–514. [Google Scholar] [CrossRef]
- Liang, T.; Gao, F.; Chen, J. Role of PTEN-less in cardiac injury, hypertrophy and regeneration. Cell Regen. 2021, 10, 25. [Google Scholar] [CrossRef]
- Hromadníková, I.; Ivankova, K.; Vedmetskaya, Y.; Krofta, L. Profiling of cardiovascular and cerebrovascular disease associated microRNA expression in umbilical cord blood in gestational hypertension, preeclampsia and fetal growth restriction. Int. J. Cardiol. 2017, 249, 402–409. [Google Scholar] [CrossRef]
- Hromadnikova, I.; Kotlabova, K.; Hympanova, L.; Krofta, L. Cardiovascular and Cerebrovascular Disease Associated microRNAs Are Dysregulated in Placental Tissues Affected with Gestational Hypertension, Preeclampsia and Intrauterine Growth Restriction. PLoS ONE 2015, 10, e0138383. [Google Scholar] [CrossRef]
- Xing, X.; Guo, S.; Zhang, G.; Liu, Y.; Bi, S.; Wang, X.; Lu, Q. miR-26a-5p protects against myocardial ischemia/reperfusion injury by regulating the PTEN/PI3K/AKT signaling pathway. Braz. J. Med. Biol. Res. 2020, 53, e9106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, Q.; Feng, Y.; Chen, X.; Yang, L.; Xu, M.; Wang, X.; Li, W.; Niu, X.; Gao, D. MicroRNA-26a Protects the Heart Against Hypertension-Induced Myocardial Fibrosis. J. Am. Heart Assoc. 2020, 9, e017970. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Xu, D.-L.; Liu, X.-B.; Bi, S.-J.; Zhang, J. MicroRNA 145 5p inhibits hypoxia/reoxygenation induced apoptosis in H9c2 cardiomyocytes by targeting ROCK1. Exp. Ther. Med. 2021, 22, 796. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Qi, H.; Huan, L.; Yang, Y. Effect of miR-195-5p on cardiomyocyte apoptosis in rats with heart failure by regulating TGF-β1/Smad3 signaling pathway. Biosci. Rep. 2020, 40, BSR20200566. [Google Scholar] [CrossRef]
- Wu, J.; Subbaiah, K.C.V.; Jiang, F.; Hedaya, O.; Mohan, A.; Yang, T.; Welle, K.; Ghaemmaghami, S.; Tang, W.H.W.; Small, E.; et al. MicroRNA-574 regulates FAM210A expression and influences pathological cardiac remodeling. EMBO Mol. Med. 2021, 13, e12710. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Pang, B.; Xiao, Y.; Zhou, S.; He, B.; Zhang, F.; Liu, W.; Peng, H.; Li, P. The protective microRNA-199a-5p-mediated unfolded protein response in hypoxic cardiomyocytes is regulated by STAT3 pathway. J. Physiol. Biochem. 2019, 75, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yan, C.; Fan, J.; Hou, Z.; Han, Y. MiR-221-3p targets Hif-1α to inhibit angiogenesis in heart failure. Lab. Investig. 2021, 101, 104–115. [Google Scholar] [CrossRef]
- Zhou, C.; Zou, Q.; Li, H.; Wang, R.; Liu, A.; Magness, R.R.; Zheng, J. Preeclampsia Downregulates MicroRNAs in Fetal Endothelial Cells: Roles of miR-29a/c-3p in Endothelial Function. J. Clin. Endocrinol. Metab. 2017, 102, 3470–3479. [Google Scholar] [CrossRef]
- Yu, G.Z.; Reilly, S.; Lewandowski, A.J.; Aye, C.Y.; Simpson, L.J.; Newton, L.D.; Davis, E.F.; Zhu, S.J.; Fox, W.R.; Goel, A.; et al. Neonatal MicroRNA Profile Determines Endothelial Function in Offspring of Hypertensive Pregnancies. Hypertension 2018, 72, 937–945. [Google Scholar] [CrossRef]
- Petrkova, J.; Borucka, J.; Kalab, M.; Klevcova, P.; Michalek, J.; Taborsky, M.; Petrek, M. Increased Expression of miR-146a in Valvular Tissue From Patients With Aortic Valve Stenosis. Front. Cardiovasc. Med. 2019, 6, 86. [Google Scholar] [CrossRef]
- Shimada, B.K.; Yang, Y.; Zhu, J.; Wang, S.; Suen, A.; Kronstadt, S.M.; Jeyaram, A.; Jay, S.M.; Zou, L.; Chao, W. Extracellular miR-146a-5p Induces Cardiac Innate Immune Response and Cardiomyocyte Dysfunction. ImmunoHorizons 2020, 4, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Brodowski, L.; Schröder-Heurich, B.; von Hardenberg, S.; Richter, K.; von Kaisenberg, C.S.; Dittrich-Breiholz, O.; Meyer, N.; Dörk, T.; von Versen-Höynck, F. MicroRNA Profiles of Maternal and Neonatal Endothelial Progenitor Cells in Preeclampsia. Int. J. Mol. Sci. 2021, 22, 5320. [Google Scholar] [CrossRef] [PubMed]
- Hromadnikova, I.; Kotlabova, K.; Dvorakova, L.; Krofta, L.; Sirc, J. Postnatal Expression Profile of microRNAs Associated with Cardiovascular and Cerebrovascular Diseases in Children at the Age of 3 to 11 Years in Relation to Previous Occurrence of Pregnancy-Related Complications. Int. J. Mol. Sci. 2019, 20, 654. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Ma, Z.; Ding, Y.; Bedarida, T.; Chen, L.; Xie, Z.; Song, P.; Zou, M.-H. Circulating miR-103a-3p contributes to angiotensin II-induced renal inflammation and fibrosis via a SNRK/NF-κB/p65 regulatory axis. Nat. Commun. 2019, 10, 2145. [Google Scholar] [CrossRef]
- Fu, X.; Qie, J.; Fu, Q.; Chen, J.; Jin, Y.; Ding, Z. miR-20a-5p/TGFBR2 Axis Affects Pro-inflammatory Macrophages and Aggravates Liver Fibrosis. Front. Oncol. 2020, 10, 107. [Google Scholar] [CrossRef]
- Ray, S.L.; Coulson, D.J.; Yeoh, M.L.Y.; Tamara, A.; Latief, J.S.; Bakhashab, S.; Weaver, J.U. The Role of miR-342 in Vascular Health. Study in Subclinical Cardiovascular Disease in Mononuclear Cells, Plasma, Inflammatory Cytokines and PANX2. Int. J. Mol. Sci. 2020, 21, 7217. [Google Scholar] [CrossRef]
- Catuogno, S.; Esposito, C.L.; Quintavalle, C.; Condorelli, G.; De Franciscis, V.; Cerchia, L. Nucleic Acids in Human Glioma Treatment: Innovative Approaches and Recent Results. J. Signal Transduct. 2012, 2012, 735135. [Google Scholar] [CrossRef]
- Chen, S.; Puthanveetil, P.; Feng, B.; Matkovich, S.J.; Dorn, G.W.; Chakrabarti, S. Cardiac miR-133a overexpression prevents early cardiac fibrosis in diabetes. J. Cell. Mol. Med. 2014, 18, 415–421. [Google Scholar] [CrossRef]
- Kurusamy, S.; Ihugba, J.C.; Armesilla, A.L.; Cotton, J.M. P10 MIR-133a overexpression impairs endothelial cell migration and tube formation in vitro. Heart 2018, 104, A5. [Google Scholar] [CrossRef]
- Apicella, C.; Ruano, C.S.M.; Méhats, C.; Miralles, F.; Vaiman, D. The Role of Epigenetics in Placental Development and the Etiology of Preeclampsia. Int. J. Mol. Sci. 2019, 20, 2837. [Google Scholar] [CrossRef]
- Ching, T.; Ha, J.; Song, M.-A.; Tiirikainen, M.; Molnar, J.; Berry, M.J.; Towner, D.; Garmire, L.X. Genome-scale hypomethylation in the cord blood DNAs associated with early onset preeclampsia. Clin. Epigenetics 2015, 7, 21. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Isa, N.I.; Syafruddin, S.E.; Mokhtar, M.H.; Zainal Abidin, S.; Jaffar, F.H.F.; Ugusman, A.; Hamid, A.A. Potential Roles of microRNAs for Assessing Cardiovascular Risk in Pre-Eclampsia-Exposed Postpartum Women and Offspring. Int. J. Mol. Sci. 2023, 24, 16842. https://doi.org/10.3390/ijms242316842
Mohd Isa NI, Syafruddin SE, Mokhtar MH, Zainal Abidin S, Jaffar FHF, Ugusman A, Hamid AA. Potential Roles of microRNAs for Assessing Cardiovascular Risk in Pre-Eclampsia-Exposed Postpartum Women and Offspring. International Journal of Molecular Sciences. 2023; 24(23):16842. https://doi.org/10.3390/ijms242316842
Chicago/Turabian StyleMohd Isa, Nurul Iffah, Saiful Effendi Syafruddin, Mohd Helmy Mokhtar, Shahidee Zainal Abidin, Farah Hanan Fathihah Jaffar, Azizah Ugusman, and Adila A. Hamid. 2023. "Potential Roles of microRNAs for Assessing Cardiovascular Risk in Pre-Eclampsia-Exposed Postpartum Women and Offspring" International Journal of Molecular Sciences 24, no. 23: 16842. https://doi.org/10.3390/ijms242316842
APA StyleMohd Isa, N. I., Syafruddin, S. E., Mokhtar, M. H., Zainal Abidin, S., Jaffar, F. H. F., Ugusman, A., & Hamid, A. A. (2023). Potential Roles of microRNAs for Assessing Cardiovascular Risk in Pre-Eclampsia-Exposed Postpartum Women and Offspring. International Journal of Molecular Sciences, 24(23), 16842. https://doi.org/10.3390/ijms242316842