
The Novel Atypical Antipsychotic Lurasidone Affects Cytochrome P450 Expression in the Liver and Peripheral Blood Lymphocytes


Abstract
1. Introduction
2. Results
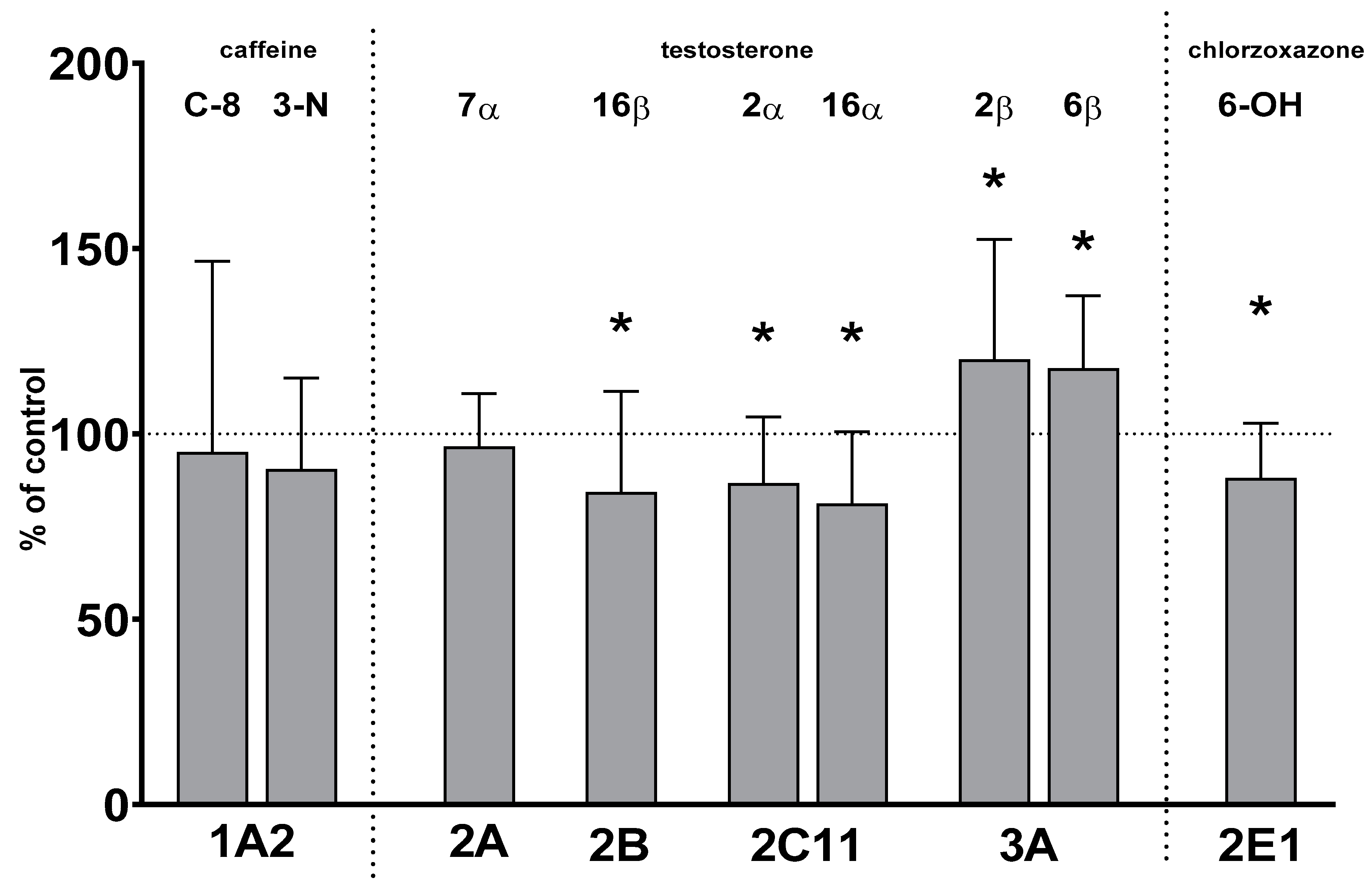
2.1. CYP Enzyme Activity in Rat Liver Microsomes after Prolonged Lurasidone Administration
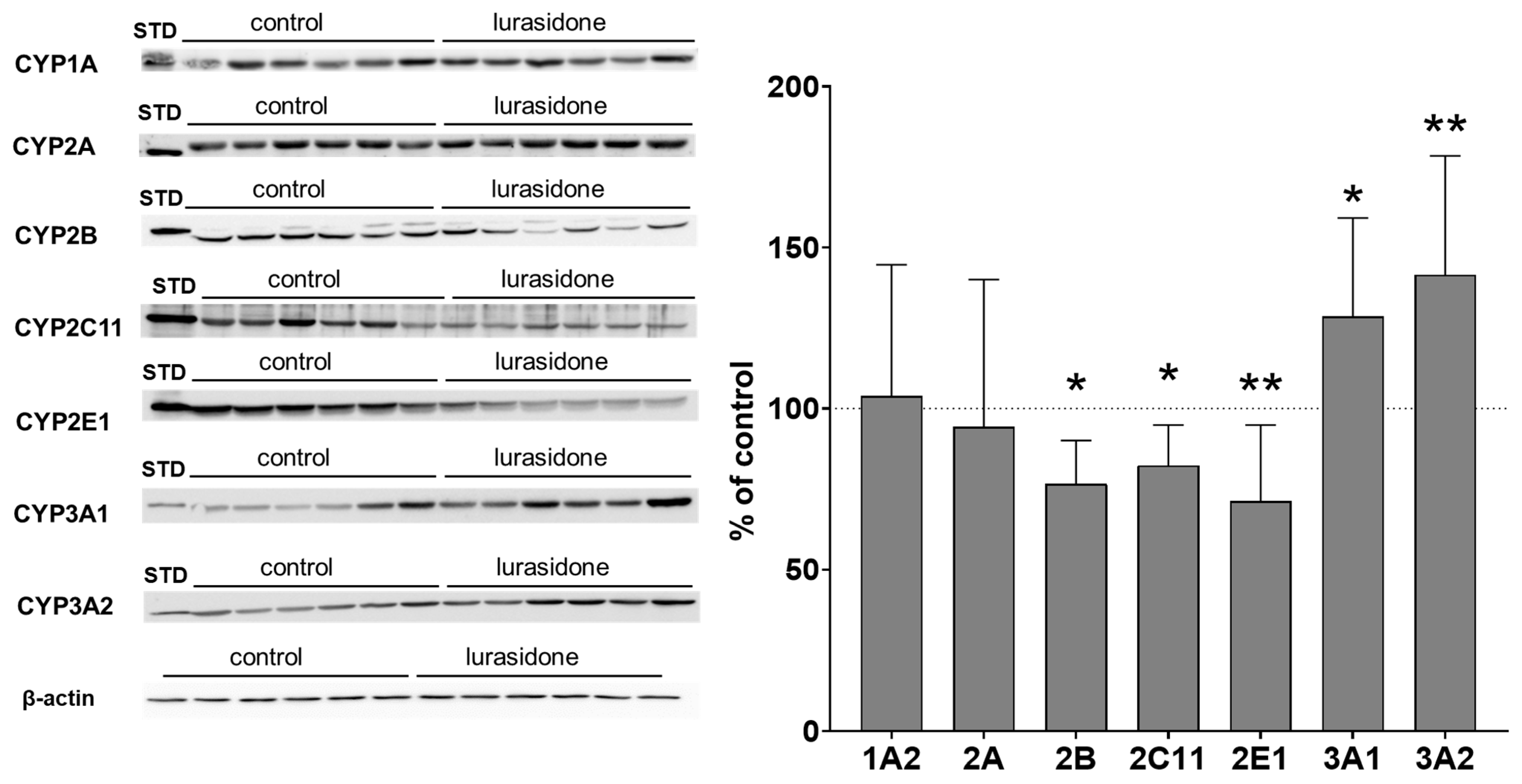
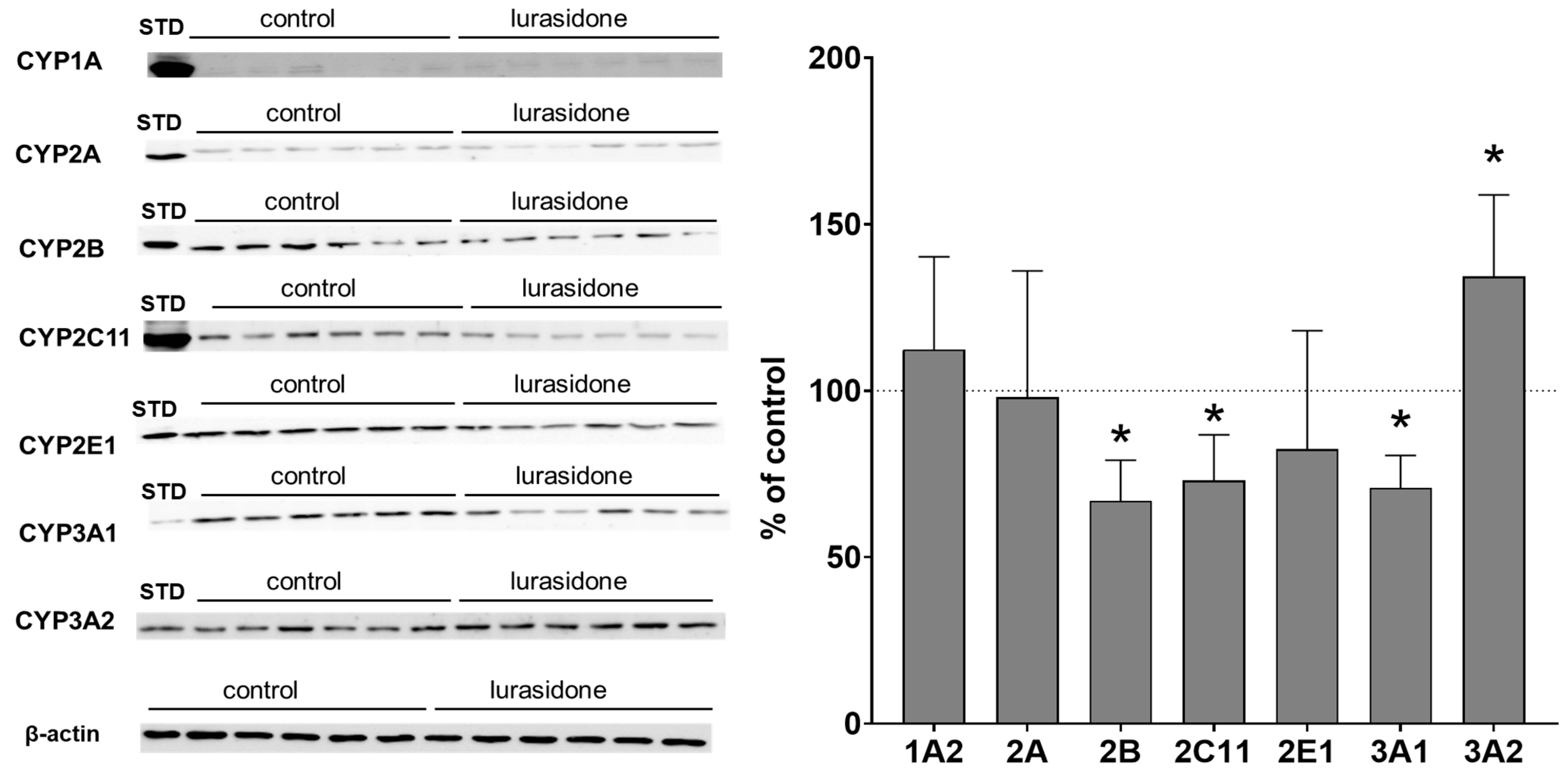
2.2. CYP Protein Level in Liver Microsomes and Lymphocytes after Chronic Lurasidone Treatment
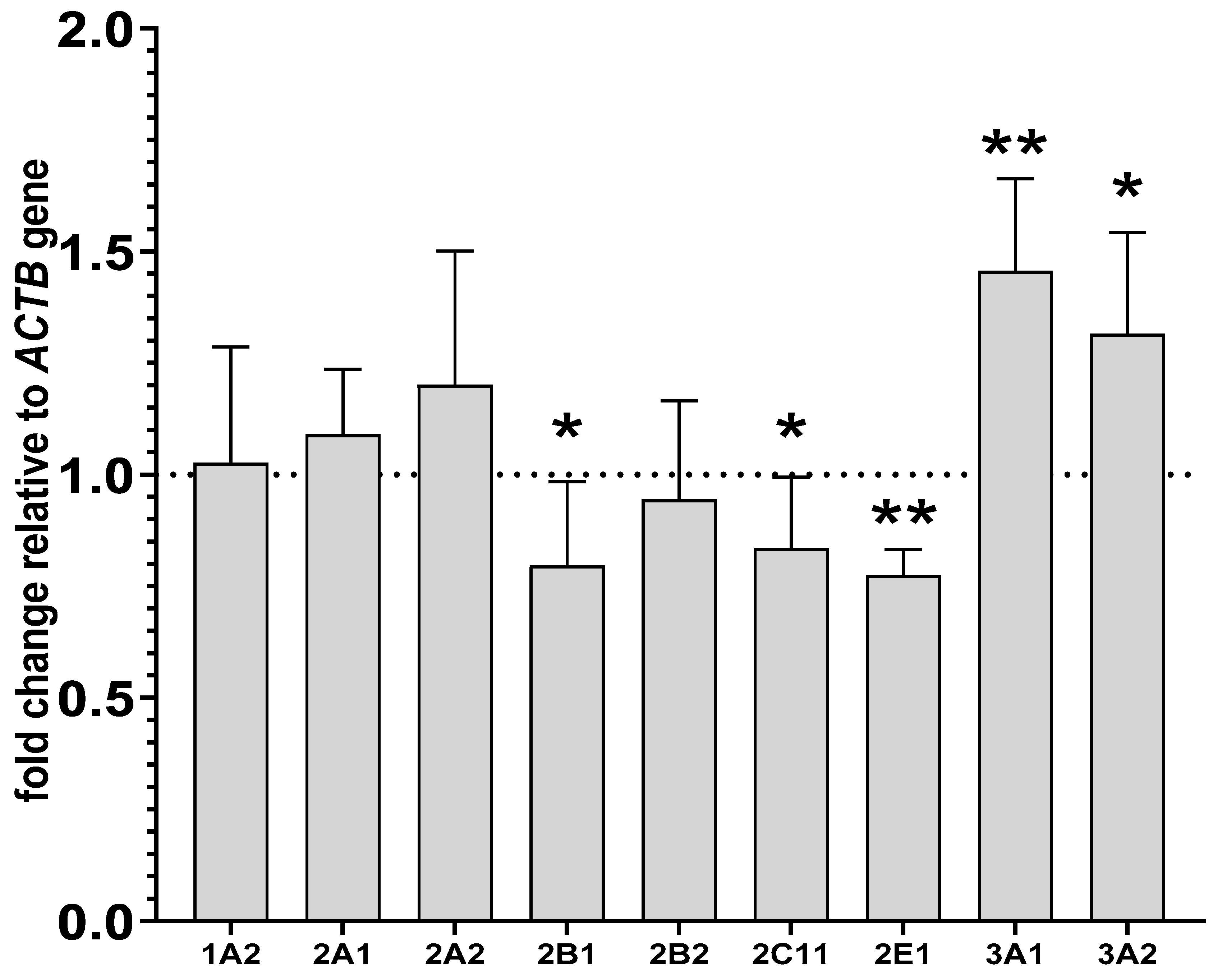
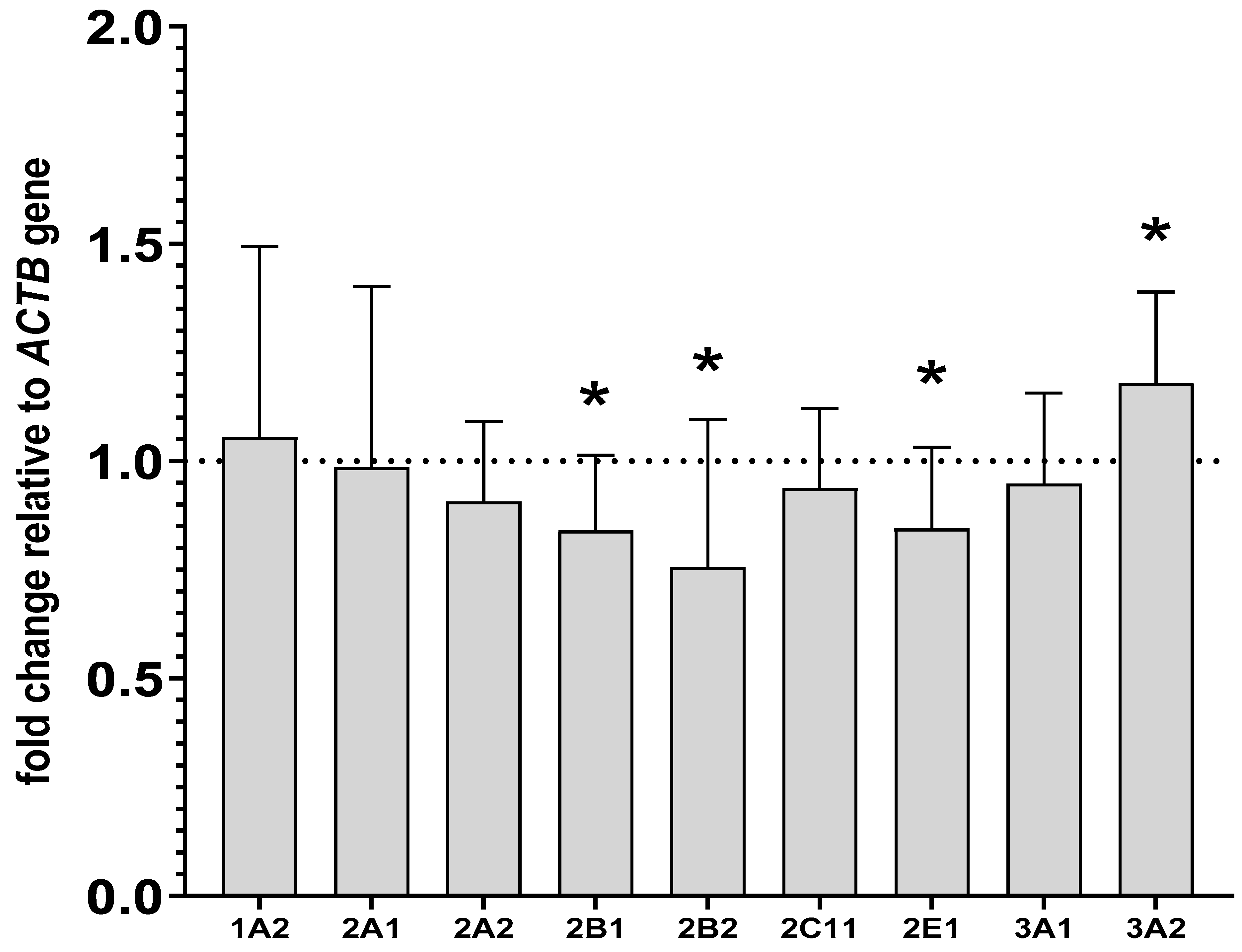
2.3. Alterations in the CYP mRNA Levels in the Liver and Lymphocyte Cells after Lurasidone Treatment
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Animal Treatment
4.3. Estimation of CYP Enzyme Activities in Liver Microsomes
4.4. Lymphocyte Isolation
4.5. Western Blotting
4.6. Assay for CYP mRNA Expression
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zanger, U.M.; Schwab, M. Cytochrome P450 Enzymes in Drug Metabolism: Regulation of Gene Expression, Enzyme Activities, and Impact of Genetic Variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef] [PubMed]
- Miksys, S.; Rao, Y.; Sellers, E.M.; Kwan, M.; Mendis, D.; Tyndale, R.F. Regional and Cellular Distribution of CYP2D Subfamily Members in Rat Brain. Xenobiotica 2000, 30, 547–564. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, S.; Wang, J.; Renukuntla, J.; Sirimulla, S.; Chen, J. A Comprehensive Review of Cytochrome P450 2E1 for Xenobiotic Metabolism. Drug Metab. Rev. 2019, 51, 178–195. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, C.D.; Lu, H.; Cui, J.Y. Epigenetic Regulation of Drug Processing Genes. Toxicol. Mech. Methods 2011, 21, 312–324. [Google Scholar] [CrossRef]
- Furukawa, M.; Nishimura, M.; Ogino, D.; Chiba, R.; Ikai, I.; Ueda, N.; Naito, S.; Kuribayashi, S.; Moustafa, M.A.; Uchida, T.; et al. Cytochrome P450 Gene Expression Levels in Peripheral Blood Mononuclear Cells in Comparison with the Liver. Cancer Sci. 2004, 95, 520–529. [Google Scholar] [CrossRef]
- Saurabh, K.; Parmar, D. Evidence for Cytochrome P450 2B1/2B2 Isoenzymes in Freshly Prepared Peripheral Blood Lymphocytes. Biomarkers 2011, 16, 649–656. [Google Scholar] [CrossRef]
- Sharma, A.; Saurabh, K.; Yadav, S.; Jain, S.K.; Parmar, D. Expression Profiling of Selected Genes of Toxication and Detoxication Pathways in Peripheral Blood Lymphocytes as a Biomarker for Predicting Toxicity of Environmental Chemicals. Int. J. Hyg. Environ. Health 2013, 216, 645–651. [Google Scholar] [CrossRef]
- Dey, A.; Yadav, S.; Dhawan, A.; Seth, P.K.; Parmar, D. Evidence for Cytochrome P450 3A Expression and Catalytic Activity in Rat Blood Lymphocytes. Life Sci. 2006, 79, 1729–1735. [Google Scholar] [CrossRef]
- Dey, A.; Parmar, D.; Dhawan, A.; Dash, D.; Seth, P.K. Cytochrome P450 2E1 Dependent Catalytic Activity and Lipid Peroxidation in Rat Blood Lymphocytes. Life Sci. 2002, 71, 2509–2519. [Google Scholar] [CrossRef]
- Dey, A.; Parmar, D.; Dayal, M.; Dhawan, A.; Seth, P.K. Cytochrome P450 1A1 (CYP1A1) in Blood Lymphocytes Evidence for Catalytic Activity and mRNA Expression. Life Sci. 2001, 69, 383–393. [Google Scholar] [CrossRef]
- Hannon-Fletcher, M.P.; Barnett, Y.A. Lymphocyte Cytochrome P450 Expression: Inducibility Studies in Male Wistar Rats. Br. J. Biomed. Sci. 2008, 65, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Saurabh, K.; Sharma, A.; Yadav, S.; Parmar, D. Polycyclic Aromatic Hydrocarbon Metabolizing Cytochrome P450s in Freshly Prepared Uncultured Rat Blood Lymphocytes. Biochem. Pharmacol. 2010, 79, 1182–1188. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Dinesh, K.; Yadav, S.; Jain, S.K.; Pant, M.C.; Parmar, D. Cytochrome P450 2A Isoenzymes in Freshly Prepared Blood Lymphocytes Isolated from Rats and Validation as a Biomarker for Clinical Studies in Humans. Xenobiotica 2013, 43, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, T.; Hase, I.; Imaoka, S.; Hiroi, T.; Oda, Y.; Asada, A.; Funae, Y. Quantitative RT-PCR for CYP3A4 mRNA in Human Peripheral Lymphocytes: Induction of CYP3A4 in Lymphocytes and in Liver by Rifampicin. Pharmacogenetics 2000, 10, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Raucy, J.L.; Schultz, E.D.; Wester, M.R.; Arora, S.; Johnston, D.E.; Omdahl, J.L.; Carpenter, S.P. Human Lymphocyte Cytochrome P450 2E1, a Putative Marker for Alcohol-Mediated Changes in Hepatic Chlorzoxazone Activity. Drug Metab. Dispos. 1997, 25, 1429–1435. [Google Scholar] [PubMed]
- Shah, P.P.; Saurabh, K.; Pant, M.C.; Mathur, N.; Parmar, D. Evidence for Increased Cytochrome P450 1A1 Expression in Blood Lymphocytes of Lung Cancer Patients. Mutat. Res. 2009, 670, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Temesvári, M.; Kóbori, L.; Paulik, J.; Sárváry, E.; Belic, A.; Monostory, K. Estimation of Drug-Metabolizing Capacity by Cytochrome P450 Genotyping and Expression. J. Pharmacol. Exp. Ther. 2012, 341, 294–305. [Google Scholar] [CrossRef]
- Bahari, A.; Mehrzad, J.; Mahmoudi, M.; Bassami, M.R.; Dehghani, H. Cytochrome P450 Isoforms Are Differently Up-Regulated in Aflatoxin B1-Exposed Human Lymphocytes and Monocytes. Immunopharmacol. Immunotoxicol. 2014, 36, 1–10. [Google Scholar] [CrossRef]
- van Duursen, M.B.M.; Sanderson, J.T.; van den Berg, M. Cytochrome P450 1A1 and 1B1 in Human Blood Lymphocytes Are Not Suitable as Biomarkers of Exposure to Dioxin-like Compounds: Polymorphisms and Interindividual Variation in Expression and Inducibility. Toxicol. Sci. 2005, 85, 703–712. [Google Scholar] [CrossRef]
- Haas, C.E.; Brazeau, D.; Cloen, D.; Booker, B.M.; Frerichs, V.; Zaranek, C.; Frye, R.F.; Kufel, T. Cytochrome P450 mRNA Expression in Peripheral Blood Lymphocytes as a Predictor of Enzyme Induction. Eur. J. Clin. Pharmacol. 2005, 61, 583–593. [Google Scholar] [CrossRef]
- Daniel, W.A.; Bromek, E.; Danek, P.J.; Haduch, A. The Mechanisms of Interactions of Psychotropic Drugs with Liver and Brain Cytochrome P450 and Their Significance for Drug Effect and Drug-Drug Interactions. Biochem. Pharmacol. 2022, 199, 115006. [Google Scholar] [CrossRef] [PubMed]
- Danek, P.J.; Wójcikowski, J.; Daniel, W.A. The atypical neuroleptics iloperidone and lurasidone inhibit human cytochrome P450 enzymes in vitro. Evaluation of potential metabolic interactions. Pharmacol. Rep. 2020, 72, 1685–1694. [Google Scholar] [CrossRef] [PubMed]
- Danek, P.J.; Wójcikowski, J.; Daniel, W.A. Asenapine and Iloperidone Decrease the Expression of Major Cytochrome P450 Enzymes CYP1A2 and CYP3A4 in Human Hepatocytes. A Significance for Drug-Drug Interactions during Combined Therapy. Toxicol. Appl. Pharmacol. 2020, 406, 115239. [Google Scholar] [CrossRef] [PubMed]
- Kot, M.; Haduch, A.; Papp, M.; Daniel, W.A. The Effect of Chronic Treatment with Lurasidone on Rat Liver Cytochrome P450 Expression and Activity in the Chronic Mild Stress Model of Depression. Drug Metab. Dispos. 2017, 45, 1336–1344. [Google Scholar] [CrossRef] [PubMed]
- Jaeschke, R.R.; Sowa-Kućma, M.; Pańczyszyn-Trzewik, P.; Misztak, P.; Styczeń, K.; Datka, W. Lurasidone: The 2016 Update on the Pharmacology, Efficacy and Safety Profile. Pharmacol. Rep. 2016, 68, 748–755. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, T.; Horisawa, T.; Tokuda, K.; Ishiyama, T.; Ogasa, M.; Tagashira, R.; Matsumoto, K.; Nishikawa, H.; Ueda, Y.; Toma, S.; et al. Pharmacological Profile of Lurasidone, a Novel Antipsychotic Agent with Potent 5-Hydroxytryptamine 7 (5-HT7) and 5-HT1A Receptor Activity. J. Pharmacol. Exp. Ther. 2010, 334, 171–181. [Google Scholar] [CrossRef]
- Findling, R.L.; Goldman, R.; Chiu, Y.-Y.; Silva, R.; Jin, F.; Pikalov, A.; Loebel, A. Pharmacokinetics and Tolerability of Lurasidone in Children and Adolescents with Psychiatric Disorders. Clin. Ther. 2015, 37, 2788–2797. [Google Scholar] [CrossRef]
- Meyer, J.M.; Loebel, A.D.; Schweizer, E. Lurasidone: A New Drug in Development for Schizophrenia. Expert. Opin. Investig. Drugs 2009, 18, 1715–1726. [Google Scholar] [CrossRef]
- Wójcikowski, J.; Daniel, W.A. The Role of the Nervous System in the Regulation of Liver Cytochrome P450. Curr. Drug Metab. 2011, 12, 124–138. [Google Scholar] [CrossRef]
- Rane, A.; Liu, Z.; Levol, R.; Bjelfman, C.; Thyr, C.; Ericson, H.; Hansson, T.; Henderson, C.; Wolf, C.R. Differential Effects of Neuroleptic Agents on Hepatic Cytochrome P-450 Isozymes in the Male Rat. Biochim. Biophys. Acta 1996, 1291, 60–66. [Google Scholar] [CrossRef]
- Wójcikowski, J.; Haduch, A.; Daniel, W.A. Effect of Classic and Atypical Neuroleptics on Cytochrome P450 3A (CYP3A) in Rat Liver. Pharmacol. Rep. 2012, 64, 1411–1418. [Google Scholar] [CrossRef] [PubMed]
- Danek, P.J.; Kuban, W.; Daniel, W.A. The Effect of Chronic Iloperidone Treatment on Cytochrome P450 Expression and Activity in the Rat Liver: Involvement of Neuroendocrine Mechanisms. Int. J. Mol. Sci. 2021, 22, 8447. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Nakasa, H.; Ishii, I.; Ariyoshi, N.; Igarashi, T.; Ohmori, S.; Kitada, M. Effects of Endogenous Steroids on CYP3A4-Mediated Drug Metabolism by Human Liver Microsomes. Drug Metab. Dispos. 2002, 30, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Danek, P.J.; Bromek, E.; Daniel, W.A. The Influence of Long-Term Treatment with Asenapine on Liver Cytochrome P450 Expression and Activity in the Rat. The Involvement of Different Mechanisms. Pharmaceuticals 2021, 14, 629. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.J.; Sharma, A.; Choudhuri, G.; Parmar, D. Induction of Blood Lymphocyte Cytochrome P450 2E1 in Early Stage Alcoholic Liver Cirrhosis. Alcohol 2011, 45, 81–87. [Google Scholar] [CrossRef]
- Lin, P.; Hu, S.-W.; Chang, T.-H. Correlation between Gene Expression of Aryl Hydrocarbon Receptor (AhR), Hydrocarbon Receptor Nuclear Translocator (Arnt), Cytochromes P4501A1 (CYP1A1) and 1B1 (CYP1B1), and Inducibility of CYP1A1 and CYP1B1 in Human Lymphocytes. Toxicol. Sci. 2003, 71, 20–26. [Google Scholar] [CrossRef]
- Lampe, J.W.; Stepaniants, S.B.; Mao, M.; Radich, J.P.; Dai, H.; Linsley, P.S.; Friend, S.H.; Potter, J.D. Signatures of Environmental Exposures Using Peripheral Leukocyte Gene Expression: Tobacco Smoke. Cancer Epidemiol. Biomark. Prev. 2004, 13, 445–453. [Google Scholar] [CrossRef]
- Sharma, A.; Saurabh, K.; Yadav, S.; Jain, S.K.; Parmar, D. Ethanol Induced Induction of Cytochrome P450 2E1 and Activation of Mitogen Activated Protein Kinases in Peripheral Blood Lymphocytes. Xenobiotica 2012, 42, 317–326. [Google Scholar] [CrossRef]
- Siest, G.; Jeannesson, E.; Marteau, J.-B.; Samara, A.; Marie, B.; Pfister, M.; Visvikis-Siest, S. Transcription Factor and Drug-Metabolizing Enzyme Gene Expression in Lymphocytes from Healthy Human Subjects. Drug Metab. Dispos. 2008, 36, 182–189. [Google Scholar] [CrossRef]
- Martignoni, M.; Groothuis, G.M.M.; de Kanter, R. Species Differences between Mouse, Rat, Dog, Monkey and Human CYP-Mediated Drug Metabolism, Inhibition and Induction. Expert Opin. Drug Metab. Toxicol. 2006, 2, 875–894. [Google Scholar] [CrossRef]
- Yu, L.; Lu, S.; Zhao, N.; Ni, S.; Yao, T.; Zeng, S. Male-Specific Induction of CYP3A2 in Rats by Zolmitriptan. J. Pharm. Pharmacol. 2008, 60, 1601–1607. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, M.; Hannaway, K.E.; Adelekun, A.E.; Jayathilake, K.; Meltzer, H.Y. Prevention of the Phencyclidine-Induced Impairment in Novel Object Recognition in Female Rats by Co-Administration of Lurasidone or Tandospirone, a 5-HT(1A) Partial Agonist. Neuropsychopharmacology 2012, 37, 2175–2183. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, M.; Panos, J.J.; Kwon, S.; Oyamada, Y.; Rajagopal, L.; Meltzer, H.Y. Comparative Effect of Lurasidone and Blonanserin on Cortical Glutamate, Dopamine, and Acetylcholine Efflux: Role of Relative Serotonin (5-HT)2A and DA D2 Antagonism and 5-HT1A Partial Agonism. J. Neurochem. 2014, 128, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, M.; Neugebauer, N.M.; Oyamada, Y.; Meltzer, H.Y. Nicotinic Receptors and Lurasidone-Mediated Reversal of Phencyclidine-Induced Deficit in Novel Object Recognition. Behav. Brain Res. 2016, 301, 204–212. [Google Scholar] [CrossRef]
- Waxman, D.J. Interactions of hepatic cytochromes P-450 with steroid hormones. Regioselectivity and stereospecificity of steroid metabolism and hormonal regulation of rat P-450 enzyme expression. Biochem. Pharmacol. 1988, 37, 71–84. [Google Scholar] [CrossRef]
- Böyum, A. Isolation of Mononuclear Cells and Granulocytes from Human Blood. Isolation of Monuclear Cells by One Centrifugation, and of Granulocytes by Combining Centrifugation and Sedimentation at 1 g. Scand. J. Clin. Lab. Investig. 1968, 97, 77–89. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytochrome P450 | CYP1A2 | CYP2A1 | CYP2A2 | CYP2B1 | CYP2B2 | CYP2C11 | CYP2E1 | CYP3A1 | CYP3A2 | |
|---|---|---|---|---|---|---|---|---|---|---|
| LIVER | Activity | No change | No change | ↓ | ↓ | ↓ | ↑ | |||
| Protein | No change | No change | ↓ | ↓ | ↓ | ↑ | ↑ | |||
| mRNA | No change | No change | No change | ↓ | No change | ↓ | ↓ | ↑ | ↑ | |
| LYMPHOCYTES | Protein | No change | No change | ↓ | ↓ | (↓) | ↓ | ↑ | ||
| mRNA | No change | No change | No change | ↓ | ↓ | No change | ↓ | No change | ↑ | |
| CYP Enzymes | Microsomes | Lymphocytes | ||||
|---|---|---|---|---|---|---|
| Primary Antibody | Secondary Antibody | Positive Control [μg] | Primary Antibody | Secondary Antibody | Positive Control [μg] | |
| CYP1A | 1:1000 | 1:4000 | 5 | 1:1000 | 1:1000 | 1 |
| CYP2A | 1:2000 | 1:4000 | 5 | 1:2000 | 1:1000 | 1 |
| CYP2B | 1:1000 | 1:4000 | 5 | 1:1000 | 1:1000 | 1 |
| CYP2C11 | 1:1000 | 1:4000 | 5 | 1:1000 | 1:1000 | 1 |
| CYP2E1 | 1:2000 | 1:4000 | 2 | 1:2000 | 1:1000 | 0.4 |
| CYP3A1 | 1:4000 | 1:4000 | 1 | 1:4000 | 1:1000 | 0.2 |
| CYP3A2 | 1:4000 | 1:4000 | 1 | 1:4000 | 1:1000 | 0.2 |
| β-actin | 1:10,000 | 1:2000 | - | 1:20,000 | 1:2000 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danek, P.J.; Daniel, W.A. The Novel Atypical Antipsychotic Lurasidone Affects Cytochrome P450 Expression in the Liver and Peripheral Blood Lymphocytes. Int. J. Mol. Sci. 2023, 24, 16796. https://doi.org/10.3390/ijms242316796
Danek PJ, Daniel WA. The Novel Atypical Antipsychotic Lurasidone Affects Cytochrome P450 Expression in the Liver and Peripheral Blood Lymphocytes. International Journal of Molecular Sciences. 2023; 24(23):16796. https://doi.org/10.3390/ijms242316796
Chicago/Turabian StyleDanek, Przemysław J., and Władysława A. Daniel. 2023. "The Novel Atypical Antipsychotic Lurasidone Affects Cytochrome P450 Expression in the Liver and Peripheral Blood Lymphocytes" International Journal of Molecular Sciences 24, no. 23: 16796. https://doi.org/10.3390/ijms242316796
APA StyleDanek, P. J., & Daniel, W. A. (2023). The Novel Atypical Antipsychotic Lurasidone Affects Cytochrome P450 Expression in the Liver and Peripheral Blood Lymphocytes. International Journal of Molecular Sciences, 24(23), 16796. https://doi.org/10.3390/ijms242316796