


Recombinant Laminin-511 Fragment (iMatrix-511) Coating Supports Maintenance of Human Nucleus Pulposus Progenitor Cells In Vitro


 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
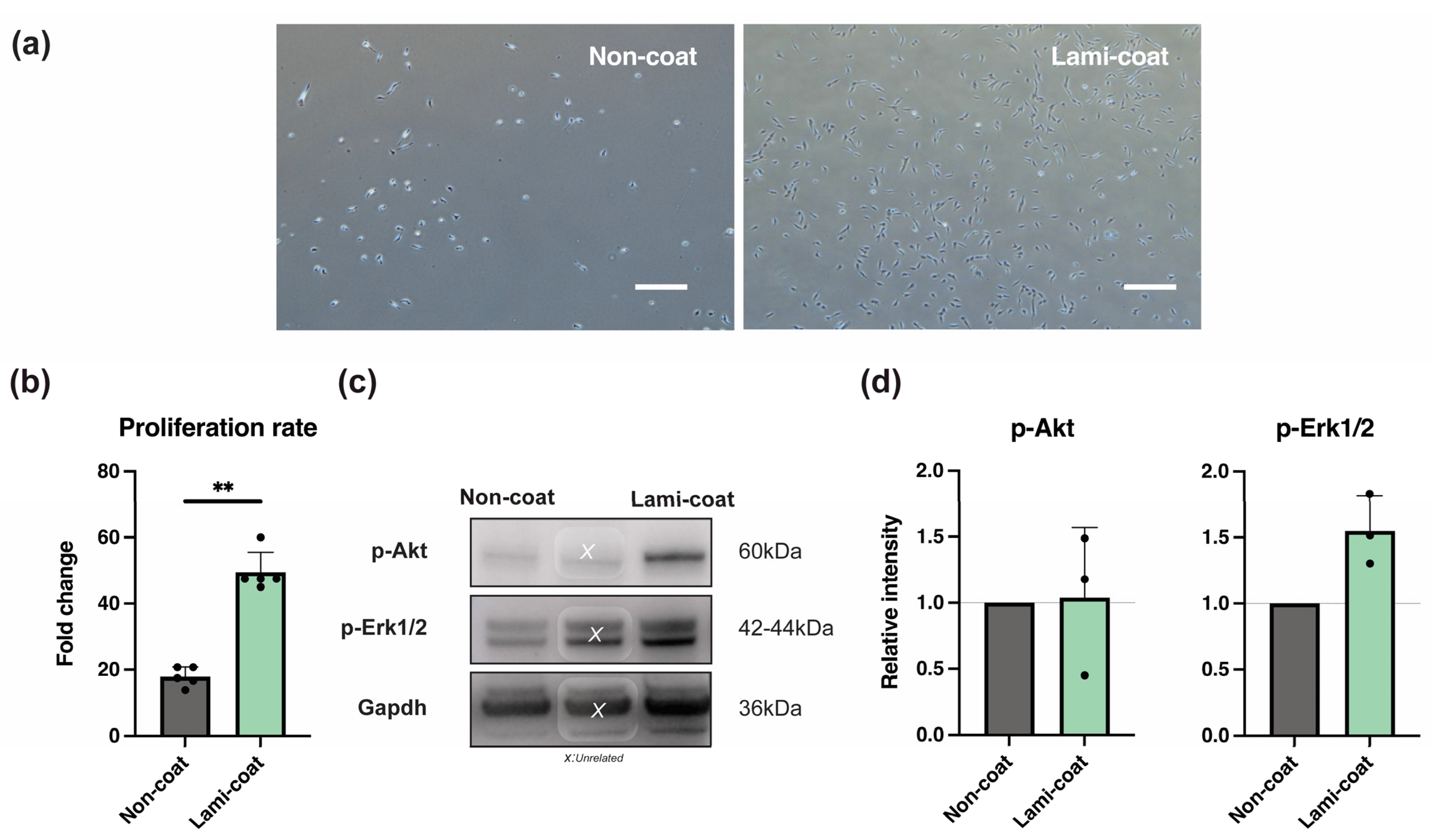
2.1. Laminin-511 Coating Enhances NP Cell Proliferation Potency
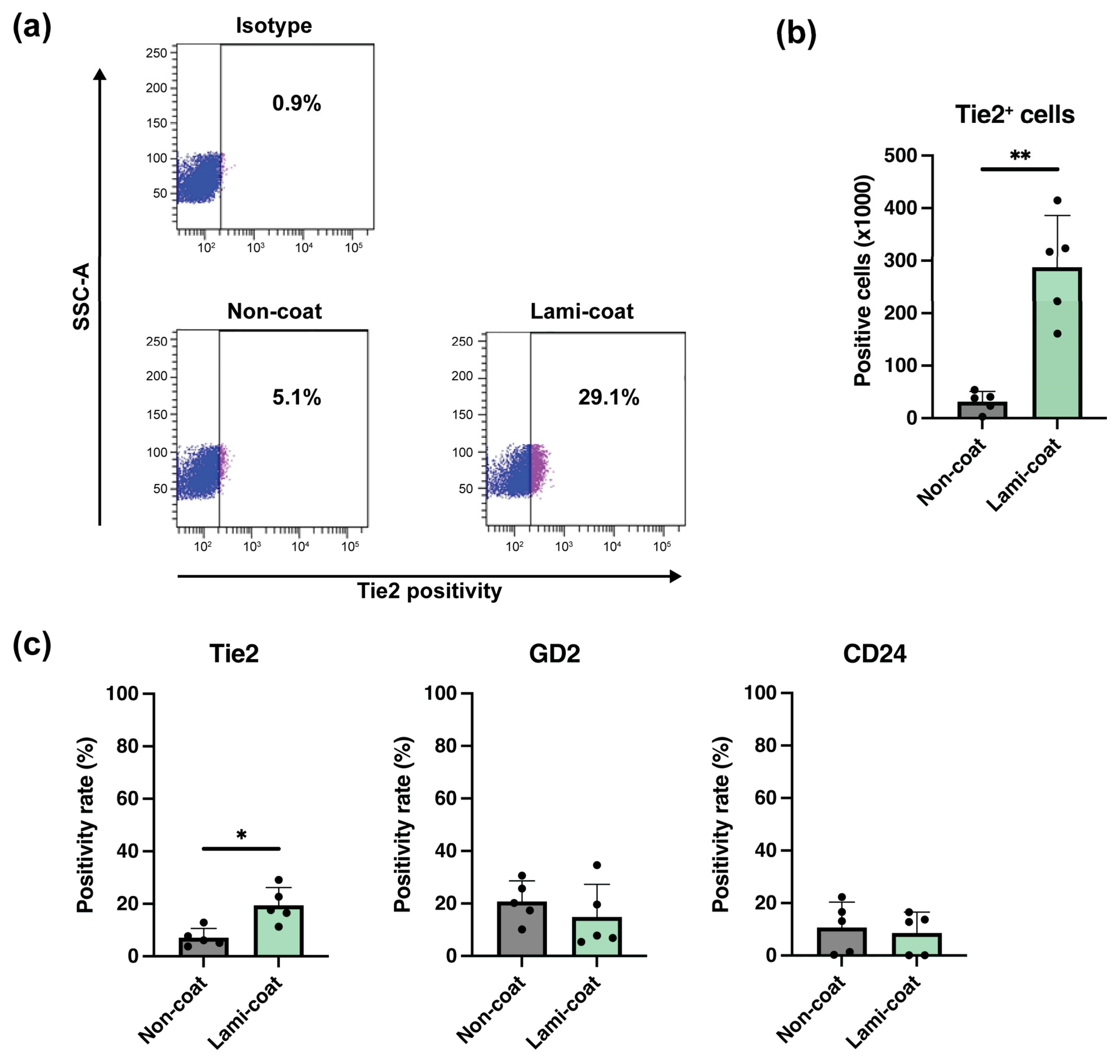
2.2. Laminin-511 Coating Strongly Increases Tie2-Expressing NP Cell Yields
2.3. Laminin-511 Supports Enhanced Type II Collagen Production
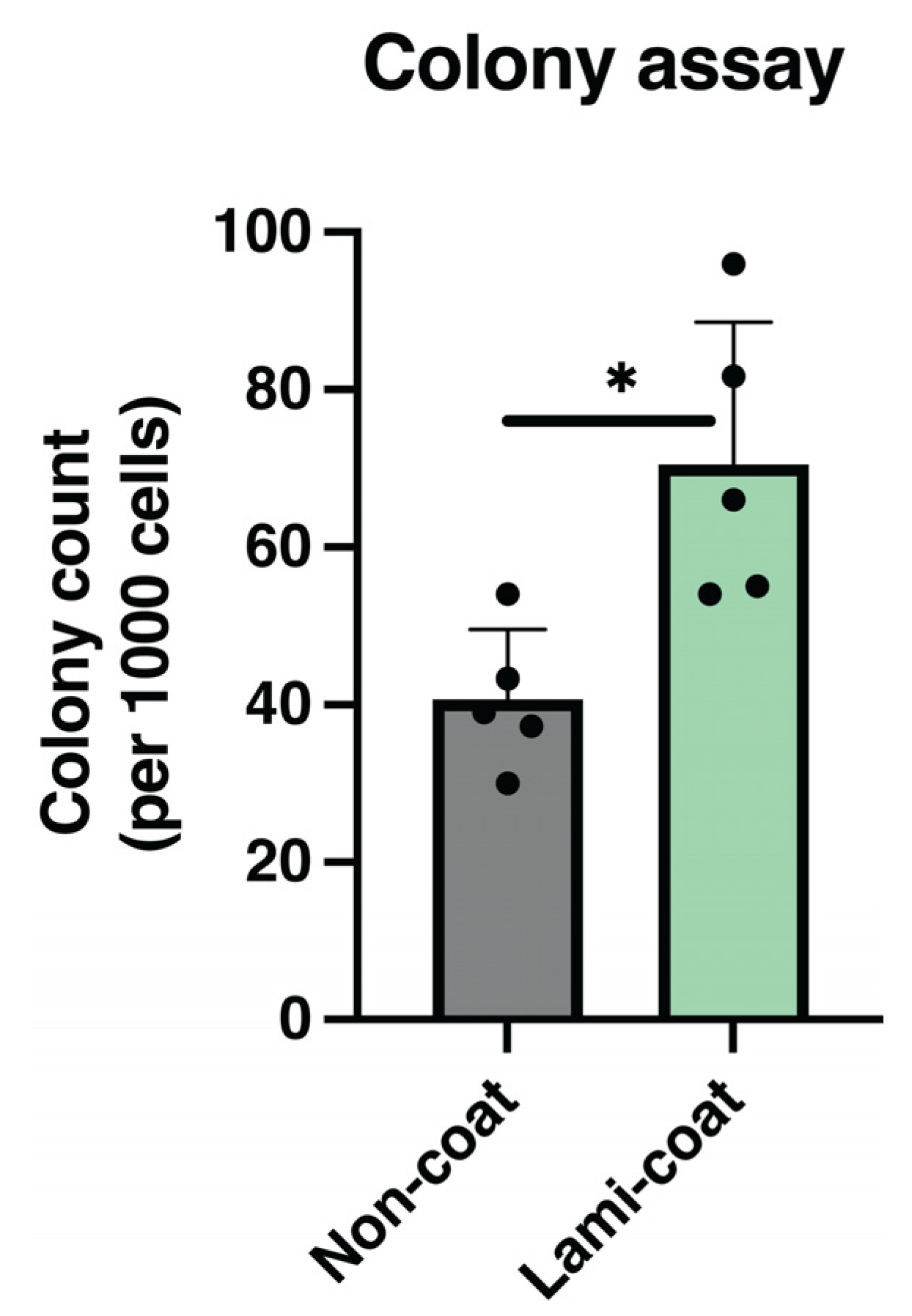
2.4. Colony Formation Rate Increased by Laminin-511
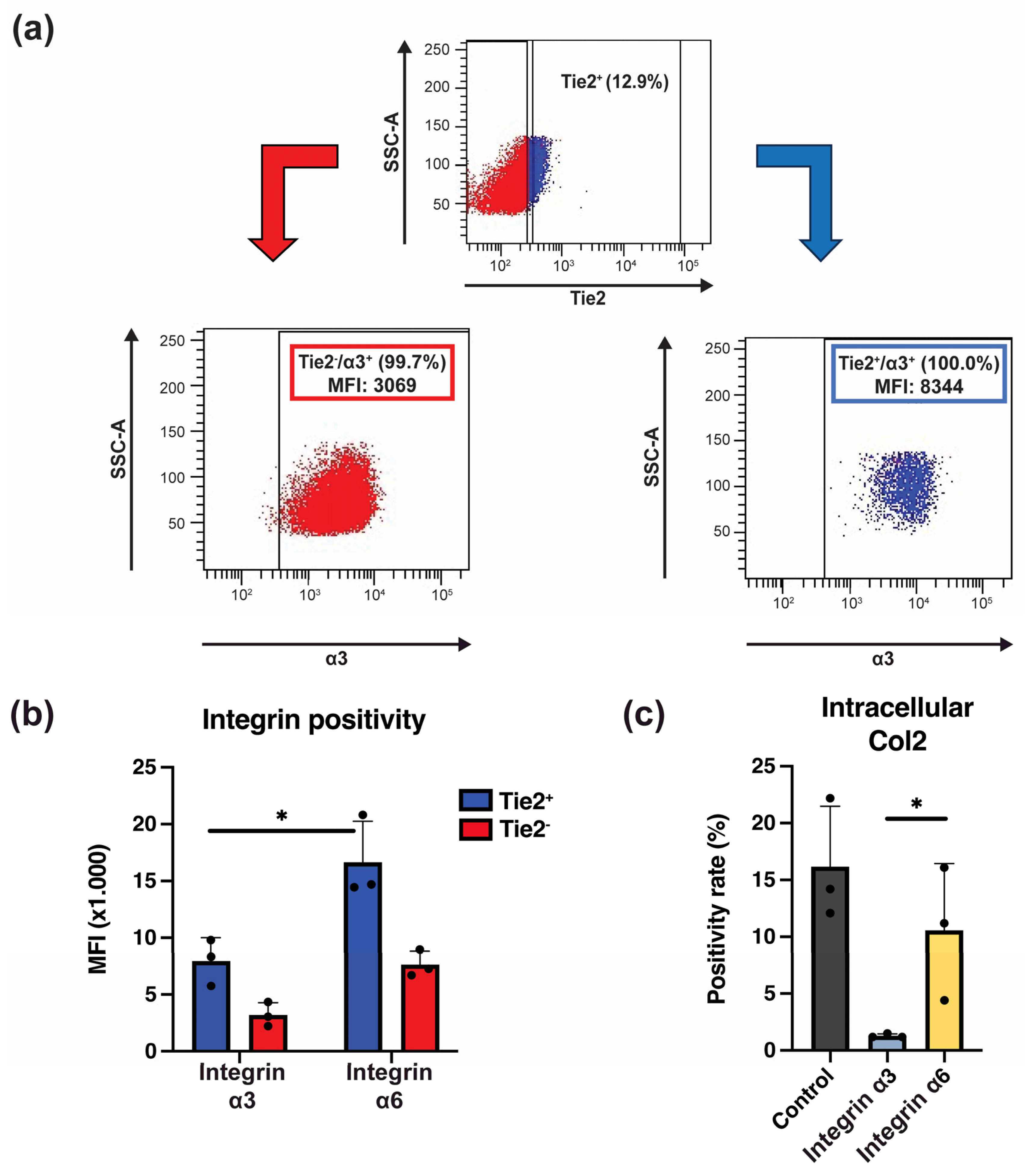
2.5. Integrins Are Crucial in Laminin-511-Mediated NP Progenitor Cell Retention
3. Discussion
4. Materials and Methods
4.1. Methods of NP Cell Isolation
4.2. Laminin-Coating
4.3. Cell Viability and Counting
4.4. Western Blot Analysis
4.5. Flow Cytometry Analysis
4.6. Colony Formation Assay Method
4.7. Integrin Blocking Assay
4.8. Statistics and Visualization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Glossary
References
- Vos, T.; Flaxman, A.D.; Naghavi, M.; Lozano, R.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S.; Aboyans, V.; et al. Years lived with disability (YLDs) for 1160 sequelae of 289 diseases and injuries 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2163–2196. [Google Scholar] [CrossRef]
- Huang, Z.; Guo, W.; Martin, J.T. Socioeconomic status, mental health, and nutrition are the principal traits for low back pain phenotyping: Data from the osteoarthritis initiative. JOR Spine 2023, 6, e1248. [Google Scholar] [CrossRef]
- Oichi, T.; Taniguchi, Y.; Oshima, Y.; Tanaka, S.; Saito, T. Pathomechanism of intervertebral disc degeneration. JOR Spine 2020, 3, e1076. [Google Scholar] [CrossRef] [PubMed]
- Fainor, M.; Orozco, B.S.; Muir, V.G.; Mahindroo, S.; Gupta, S.; Mauck, R.L.; Burdick, J.A.; Smith, H.E.; Gullbrand, S.E. Mechanical crosstalk between the intervertebral disc, facet joints, and vertebral endplate following acute disc injury in a rabbit model. JOR Spine 2023, e1287. [Google Scholar] [CrossRef]
- Roughley, P.J. Biology of intervertebral disc aging and degeneration: Involvement of the extracellular matrix. Spine 2004, 29, 2691–2699. [Google Scholar] [CrossRef] [PubMed]
- Costi, J.J.; Ledet, E.H.; O’Connell, G.D. Spine biomechanical testing methodologies: The controversy of consensus vs scientific evidence. JOR Spine 2021, 4, e1138. [Google Scholar] [CrossRef]
- Newell, N.; Little, J.P.; Christou, A.; Adams, M.A.; Adam, C.J.; Masouros, S.D. Biomechanics of the human intervertebral disc: A review of testing techniques and results. J. Mech. Behav. Biomed. Mater. 2017, 69, 420–434. [Google Scholar] [CrossRef] [PubMed]
- Fearing, B.V.; Hernandez, P.A.; Setton, L.A.; Chahine, N.O. Mechanotransduction and cell biomechanics of the intervertebral disc. JOR Spine 2018, 1, e1026. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.H.; Li, C.; Mudd, J.L.; Go, G.; Sutherland, A.E. Compositional and structural requirements for laminin and basement membranes during mouse embryo implantation and gastrulation. Development 2004, 131, 2247–2256. [Google Scholar] [CrossRef]
- Humphreys, M.D.; Ward, L.; Richardson, S.M.; Hoyland, J.A. An optimized culture system for notochordal cell expansion with retention of phenotype. JOR Spine 2018, 1, e1028. [Google Scholar] [CrossRef]
- Wang, F.; Cheung, C.W.; Wong, S.S.C. Regenerative medicine for the treatment of chronic low back pain: A narrative review. J. Int. Med. Res. 2023, 51, 3000605231155777. [Google Scholar] [CrossRef]
- Härtl, R.; Bonassar, L.; Bonassar, L.J. Biological Approaches to Spinal Disc Repair and Regeneration for Clinicians; Thieme Medical Publishers, Incorporated: Stuttgart, Germany, 2017. [Google Scholar]
- Schol, J.; Sakai, D. Comprehensive narrative review on the analysis of outcomes from cell transplantation clinical trials for discogenic low back pain. N. Am. Spine Soc. J. 2023, 13, 100195. [Google Scholar] [CrossRef] [PubMed]
- Vadala, G.; Ambrosio, L.; Russo, F.; Papalia, R.; Denaro, V. Stem Cells and Intervertebral Disc Regeneration Overview-What They Can and Can’t Do. Int. J. Spine Surg. 2021, 15, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, J.; Diwan, A.D.; Tipper, J.L. Advanced Strategies for the Regeneration of Lumbar Disc Annulus Fibrosus. Int. J. Mol. Sci. 2020, 21, 4889. [Google Scholar] [CrossRef]
- Williams, R.J.; Tryfonidou, M.A.; Snuggs, J.W.; Le Maitre, C.L. Cell sources proposed for nucleus pulposus regeneration. JOR Spine 2021, 4, e1175. [Google Scholar] [CrossRef]
- McDonnell, E.E.; Wilson, N.; Barcellona, M.N.; Ni Neill, T.; Bagnall, J.; Brama, P.A.J.; Cunniffe, G.M.; Darwish, S.L.; Butler, J.S.; Buckley, C.T. Preclinical to clinical translation for intervertebral disc repair: Effects of species-specific scale, metabolism, and matrix synthesis rates on cell-based regeneration. JOR Spine 2023, 6, e1279. [Google Scholar] [CrossRef] [PubMed]
- Sakai, D.; Nakamura, Y.; Nakai, T.; Mishima, T.; Kato, S.; Grad, S.; Alini, M.; Risbud, M.V.; Chan, D.; Cheah, K.S.; et al. Exhaustion of nucleus pulposus progenitor cells with ageing and degeneration of the intervertebral disc. Nat. Commun. 2012, 3, 1264. [Google Scholar] [CrossRef]
- Sakai, D.; Schol, J.; Bach, F.C.; Tekari, A.; Sagawa, N.; Nakamura, Y.; Chan, S.C.W.; Nakai, T.; Creemers, L.B.; Frauchiger, D.A.; et al. Successful fishing for nucleus pulposus progenitor cells of the intervertebral disc across species. JOR Spine 2018, 1, e1018. [Google Scholar] [CrossRef]
- Zhang, X.; Guerrero, J.; Croft, A.S.; Albers, C.E.; Hackel, S.; Gantenbein, B. Spheroid-Like Cultures for Expanding Angiopoietin Receptor-1 (aka. Tie2) Positive Cells from the Human Intervertebral Disc. Int. J. Mol. Sci. 2020, 21, 9423. [Google Scholar] [CrossRef]
- Sako, K.; Sakai, D.; Nakamura, Y.; Matsushita, E.; Schol, J.; Warita, T.; Horikita, N.; Sato, M.; Watanabe, M. Optimization of Spheroid Colony Culture and Cryopreservation of Nucleus Pulposus Cells for the Development of Intervertebral Disc Regenerative Therapeutics. Appl. Sci. 2021, 11, 3309. [Google Scholar] [CrossRef]
- Sako, K.; Sakai, D.; Nakamura, Y.; Schol, J.; Matsushita, E.; Warita, T.; Horikita, N.; Sato, M.; Watanabe, M. Effect of Whole Tissue Culture and Basic Fibroblast Growth Factor on Maintenance of Tie2 Molecule Expression in Human Nucleus Pulposus Cells. Int. J. Mol. Sci. 2021, 22, 4723. [Google Scholar] [CrossRef]
- Ishii, T.; Sakai, D.; Schol, J.; Nakai, T.; Suyama, K.; Watanabe, M. Sciatic nerve regeneration by transplantation of in vitro differentiated nucleus pulposus progenitor cells. Regen. Med. 2017, 12, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, J.; Hackel, S.; Croft, A.S.; Albers, C.E.; Gantenbein, B. The effects of 3D culture on the expansion and maintenance of nucleus pulposus progenitor cell multipotency. JOR Spine 2021, 4, e1131. [Google Scholar] [CrossRef]
- Tekari, A.; Chan, S.C.W.; Sakai, D.; Grad, S.; Gantenbein, B. Angiopoietin-1 receptor Tie2 distinguishes multipotent differentiation capability in bovine coccygeal nucleus pulposus cells. Stem Cell Res. Ther. 2016, 7, 75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Z.; Chen, P.; Ma, C.Y.; Li, C.; Au, T.Y.K.; Tam, V.; Peng, Y.; Wu, R.; Cheung, K.M.C.; et al. Directed Differentiation of Notochord-like and Nucleus Pulposus-like Cells Using Human Pluripotent Stem Cells. Cell Rep. 2020, 30, 2791–2806.e2795. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Shang, Y.; Yu, J.; Zeng, X.; Lin, J.; Tu, M.; Cheang, L.H.; Zhang, J. Regenerative potential of human nucleus pulposus resident stem/progenitor cells declines with ageing and intervertebral disc degeneration. Int. J. Mol. Med. 2018, 42, 2193–2202. [Google Scholar] [CrossRef] [PubMed]
- Sako, K.; Schol, J.; Sakai, D.; Nakamura, Y.; Warita, T.; Matsushita, E.; Watanabe, M. Tie2 rejuvenation of isolated nucleus pulposus progenitor cells from young and aged donor tissue sources shows that age still matters. In Proceedings of the ISSLS, Melbourne, Australia, 3 May 2023. [Google Scholar]
- Nukaga, T.; Sakai, D.; Schol, J.; Suyama, K.; Nakai, T.; Hiyama, A.; Watanabe, M. Minimal Sustainability of Dedifferentiation by ROCK Inhibitor on Rat Nucleus Pulposus Cells In Vitro. Spine Surg. Relat. Res. 2019, 3, 385–391. [Google Scholar] [CrossRef]
- Zhang, X.; Guerrero, J.; Croft, A.S.; Oswald, K.A.C.; Albers, C.E.; Häckel, S.; Gantenbein, B. Towards Tissue-Specific Stem Cell Therapy for the Intervertebral Disc: PPARδ Agonist Increases the Yield of Human Nucleus Pulposus Progenitor Cells in Expansion. Surgeries 2021, 2, 92–104. [Google Scholar] [CrossRef]
- Bischof, M.C.; Häckel, S.; Oberli, A.; Croft, A.S.; Oswald, K.A.C.; Albers, C.E.; Gantenbein, B.; Guerrero, J. Influence of Angiopoietin Treatment with Hypoxia and Normoxia on Human Intervertebral Disc Progenitor Cell’s Proliferation, Metabolic Activity, and Phenotype. Appl. Sci. 2021, 11, 7144. [Google Scholar] [CrossRef]
- Croft, A.S.; Guerrero, J.; Oswald, K.A.C.; Hackel, S.; Albers, C.E.; Gantenbein, B. Effect of different cryopreservation media on human nucleus pulposus cells’ viability and trilineage potential. JOR Spine 2021, 4, e1140. [Google Scholar] [CrossRef]
- Nakahata, T.; Ogawa, M. Clonal origin of murine hemopoietic colonies with apparent restriction to granuclocyte-macrophage-megakaryocyte (GMM) differentiation. J. Cell. Physiol. 1982, 111, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.H.; Yurchenco, P.D. Laminin functions in tissue morphogenesis. Annu. Rev. Cell Dev. Biol. 2004, 20, 255–284. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Futaki, S.; Suemori, H.; Taniguchi, Y.; Yamada, M.; Kawasaki, M.; Hayashi, M.; Kumagai, H.; Nakatsuji, N.; Sekiguchi, K.; et al. Laminin E8 fragments support efficient adhesion and expansion of dissociated human pluripotent stem cells. Nat. Commun. 2012, 3, 1236. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jing, L.; Gilchrist, C.L.; Richardson, W.J.; Fitch, R.D.; Setton, L.A. Expression of laminin isoforms, receptors, and binding proteins unique to nucleus pulposus cells of immature intervertebral disc. Connect. Tissue Res. 2009, 50, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Speer, J.; Barcellona, M.; Jing, L.; Liu, B.; Lu, M.; Kelly, M.; Buchowski, J.; Zebala, L.; Luhmann, S.; Gupta, M.; et al. Integrin-mediated interactions with a laminin-presenting substrate modulate biosynthesis and phenotypic expression for cells of the human nucleus pulposus. Eur. Cells Mater. 2021, 41, 793–810. [Google Scholar] [CrossRef]
- Nettles, D.L.; Richardson, W.J.; Setton, L.A. Integrin expression in cells of the intervertebral disc. J. Anat. 2004, 204, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, C.L.; Chen, J.; Richardson, W.J.; Loeser, R.F.; Setton, L.A. Functional integrin subunits regulating cell-matrix interactions in the intervertebral disc. J. Orthop. Res. 2007, 25, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Takara-Bio. iMatrix-511/iMatrix-511 Silk. Available online: https://catalog.takara-bio.co.jp/product/basic_info.php?unitid=U100007716 (accessed on 15 September 2023).
- Nakagawa, M.; Taniguchi, Y.; Senda, S.; Takizawa, N.; Ichisaka, T.; Asano, K.; Morizane, A.; Doi, D.; Takahashi, J.; Nishizawa, M.; et al. A novel efficient feeder-free culture system for the derivation of human induced pluripotent stem cells. Sci. Rep. 2014, 4, 3594. [Google Scholar] [CrossRef]
- Sugawara, Y.; Hamada, K.; Yamada, Y.; Kumai, J.; Kanagawa, M.; Kobayashi, K.; Toda, T.; Negishi, Y.; Katagiri, F.; Hozumi, K.; et al. Characterization of dystroglycan binding in adhesion of human induced pluripotent stem cells to laminin-511 E8 fragment. Sci. Rep. 2019, 9, 13037. [Google Scholar] [CrossRef]
- Tanigawa, S.; Naganuma, H.; Kaku, Y.; Era, T.; Sakuma, T.; Yamamoto, T.; Taguchi, A.; Nishinakamura, R. Activin Is Superior to BMP7 for Efficient Maintenance of Human iPSC-Derived Nephron Progenitors. Stem Cell Rep. 2019, 13, 322–337. [Google Scholar] [CrossRef]
- Kokubu, Y.; Nagino, T.; Sasa, K.; Oikawa, T.; Miyake, K.; Kume, A.; Fukuda, M.; Fuse, H.; Tozawa, R.; Sakurai, H. Phenotypic Drug Screening for Dysferlinopathy Using Patient-Derived Induced Pluripotent Stem Cells. Stem Cells Transl. Med. 2019, 8, 1017–1029. [Google Scholar] [CrossRef]
- Miyake, T.; Shimada, M.; Matsumoto, Y.; Okino, A. DNA Damage Response After Ionizing Radiation Exposure in Skin Keratinocytes Derived from Human-Induced Pluripotent Stem Cells. Int. J. Radiat. Oncol. Biol. Phys. 2019, 105, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Basatvat, S.; Bach, F.C.; Barcellona, M.N.; Binch, A.L.; Buckley, C.T.; Bueno, B.; Chahine, N.O.; Chee, A.; Creemers, L.B.; Dudli, S.; et al. Harmonization and standardization of nucleus pulposus cell extraction and culture methods. JOR Spine 2023, 6, e1238. [Google Scholar] [CrossRef] [PubMed]
- Fearing, B.V.; Speer, J.E.; Jing, L.; Kalathil, A.; Kelly, M.P.; Buchowski, J.M.; Zebala, L.P.; Luhmann, S.; Gupta, M.C.; Setton, L.A. Verteporfin treatment controls morphology, phenotype, and global gene expression for cells of the human nucleus pulposus. JOR Spine 2020, 3, e1111. [Google Scholar] [CrossRef] [PubMed]
- Risbud, M.V.; Guttapalli, A.; Albert, T.J.; Shapiro, I.M. Hypoxia activates MAPK activity in rat nucleus pulposus cells: Regulation of integrin expression and cell survival. Spine 2005, 30, 2503–2509. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, K.; Hua, W.; Li, S.; Liu, X.; Song, Y.; Zhang, Y.; Shao, Z.; Li, S.; Yang, C. Fibronectin induced ITGbeta1/FAK-dependent apoptotic pathways determines the fate of degenerative NP cells. J. Orthop. Res. 2019, 37, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Bellacosa, A.; Kumar, C.C.; Di Cristofano, A.; Testa, J.R. Activation of AKT kinases in cancer: Implications for therapeutic targeting. Adv. Cancer Res. 2005, 94, 29–86. [Google Scholar] [CrossRef]
- Colognato, H.; Yurchenco, P.D. Form and function: The laminin family of heterotrimers. Dev. Dyn. 2000, 218, 213–234. [Google Scholar] [CrossRef]
- Speer, J.E.; Barcellona, M.N.; Lu, M.Y.; Zha, Z.; Jing, L.; Gupta, M.C.; Buchowski, J.M.; Kelly, M.P.; Setton, L.A. Development of a library of laminin-mimetic peptide hydrogels for control of nucleus pulposus cell behaviors. J. Tissue Eng. 2021, 12, 20417314211021220. [Google Scholar] [CrossRef]
- Molinos, M.; Fiordalisi, M.F.; Caldeira, J.; Almeida, C.R.; Barbosa, M.A.; Goncalves, R.M. Alterations of bovine nucleus pulposus cells with aging. Aging Cell 2023, 22, e13873. [Google Scholar] [CrossRef]
- Sun, Y.; Lv, M.; Zhou, L.; Tam, V.; Lv, F.; Chan, D.; Wang, H.; Zheng, Z.; Cheung, K.M.; Leung, V.Y. Enrichment of committed human nucleus pulposus cells expressing chondroitin sulfate proteoglycans under alginate encapsulation. Osteoarthr. Cartil. 2015, 23, 1194–1203. [Google Scholar] [CrossRef] [PubMed]
- Ido, H.; Nakamura, A.; Kobayashi, R.; Ito, S.; Li, S.; Futaki, S.; Sekiguchi, K. The requirement of the glutamic acid residue at the third position from the carboxyl termini of the laminin gamma chains in integrin binding by laminins. J. Biol. Chem. 2007, 282, 11144–11154. [Google Scholar] [CrossRef] [PubMed]
- Nishiuchi, R.; Takagi, J.; Hayashi, M.; Ido, H.; Yagi, Y.; Sanzen, N.; Tsuji, T.; Yamada, M.; Sekiguchi, K. Ligand-binding specificities of laminin-binding integrins: A comprehensive survey of laminin-integrin interactions using recombinant alpha3beta1, alpha6beta1, alpha7beta1 and alpha6beta4 integrins. Matrix Biol. J. Int. Soc. Matrix Biol. 2006, 25, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Silverman, L.I.; Dulatova, G.; Tandeski, T.; Erickson, I.E.; Lundell, B.; Toplon, D.; Wolff, T.; Howard, A.; Chintalacharuvu, S.; Foley, K.T. In vitro and in vivo evaluation of discogenic cells, an investigational cell therapy for disc degeneration. Spine J. Off. J. N. Am. Spine Soc. 2020, 20, 138–149. [Google Scholar] [CrossRef]
- US National Library of Medicine-ClinicalTrials.gov. 2019. Available online: https://clinicaltrials.gov/ct2/show/NCT03955315 (accessed on 28 January 2020).
- US National Library of Medicine-ClinicalTrials.gov. 2017. Available online: https://clinicaltrials.gov/ct2/show/NCT03347708 (accessed on 28 January 2020).
- DiscGenics. DiscGenics Announces Presentation of Positive Interim Clinical Data from Phase 1/2 Study of Cell Therapy for Degenerative Disc Disease. Available online: https://www.discgenics.com/news-posts/2022/2/25/discgenics-announces-presentation-of-positive-interim-clinical-data-from-phase-12-study-of-cell-therapy-for-degenerative-disc-disease (accessed on 12 September 2023).
- Takahashi, T.; Donahue, R.P.; Nordberg, R.C.; Hu, J.C.; Currall, S.C.; Athanasiou, K.A. Commercialization of regenerative-medicine therapies. Nat. Rev. Bioeng. 2023. [Google Scholar] [CrossRef]
- Benndorf, R.A.; Gehling, U.M.; Appel, D.; Maas, R.; Schwedhelm, E.; Schlagner, K.; Silberhorn, E.; Hossfeld, D.K.; Rogiers, X.; Boger, R. Mobilization of putative high-proliferative-potential endothelial colony-forming cells during antihypertensive treatment in patients with essential hypertension. Stem Cells Dev. 2007, 16, 329–338. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nakano, T.; Nakahata, T.; Asai, H.; Yagi, Y.; Tsuji, K.; Komiyama, A.; Akabane, T.; Kojima, S.; Kitamura, Y. Formation of mast cell colonies in methylcellulose by mouse peritoneal cells and differentiation of these cloned cells in both the skin and the gastric mucosa of W/Wv mice: Evidence that a common precursor can give rise to both “connective tissue-type” and “mucosal” mast cells. J. Immunol. 1986, 136, 1378–1384. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Sex | Age (y) | Pathology | Experimental Use |
|---|---|---|---|---|
| A22 | M | 22 | LDH | FCM, CFA, WB |
| T20 | M | 20 | LDH | FCM, CFA |
| A18 | M | 18 | LDH | FCM, CFA |
| T18 | M | 18 | LDH | FCM |
| T17 | F | 17 | LDH | FCM |
| T16 | M | 16 | LDH | FCM, CFA |
| A23 | M | 23 | LDH | WB, CFA |
| T21 | M | 21 | LDH | WB |
| A20 | M | 20 | LDH | WB |
| A16 | M | 16 | LDH | WB |
| K18 | M | 18 | LDH | WB |
| A16 | M | 16 | LDH | WB |
| T19 | M | 19 | LDH | FCM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soma, H.; Sakai, D.; Nakamura, Y.; Tamagawa, S.; Warita, T.; Schol, J.; Matsushita, E.; Naiki, M.; Sato, M.; Watanabe, M. Recombinant Laminin-511 Fragment (iMatrix-511) Coating Supports Maintenance of Human Nucleus Pulposus Progenitor Cells In Vitro. Int. J. Mol. Sci. 2023, 24, 16713. https://doi.org/10.3390/ijms242316713
Soma H, Sakai D, Nakamura Y, Tamagawa S, Warita T, Schol J, Matsushita E, Naiki M, Sato M, Watanabe M. Recombinant Laminin-511 Fragment (iMatrix-511) Coating Supports Maintenance of Human Nucleus Pulposus Progenitor Cells In Vitro. International Journal of Molecular Sciences. 2023; 24(23):16713. https://doi.org/10.3390/ijms242316713
Chicago/Turabian StyleSoma, Hazuki, Daisuke Sakai, Yoshihiko Nakamura, Shota Tamagawa, Takayuki Warita, Jordy Schol, Erika Matsushita, Mitsuru Naiki, Masato Sato, and Masahiko Watanabe. 2023. "Recombinant Laminin-511 Fragment (iMatrix-511) Coating Supports Maintenance of Human Nucleus Pulposus Progenitor Cells In Vitro" International Journal of Molecular Sciences 24, no. 23: 16713. https://doi.org/10.3390/ijms242316713
APA StyleSoma, H., Sakai, D., Nakamura, Y., Tamagawa, S., Warita, T., Schol, J., Matsushita, E., Naiki, M., Sato, M., & Watanabe, M. (2023). Recombinant Laminin-511 Fragment (iMatrix-511) Coating Supports Maintenance of Human Nucleus Pulposus Progenitor Cells In Vitro. International Journal of Molecular Sciences, 24(23), 16713. https://doi.org/10.3390/ijms242316713