
The Molecular Mechanism of Potassium Absorption, Transport, and Utilization in Rice



Abstract
:1. Introduction
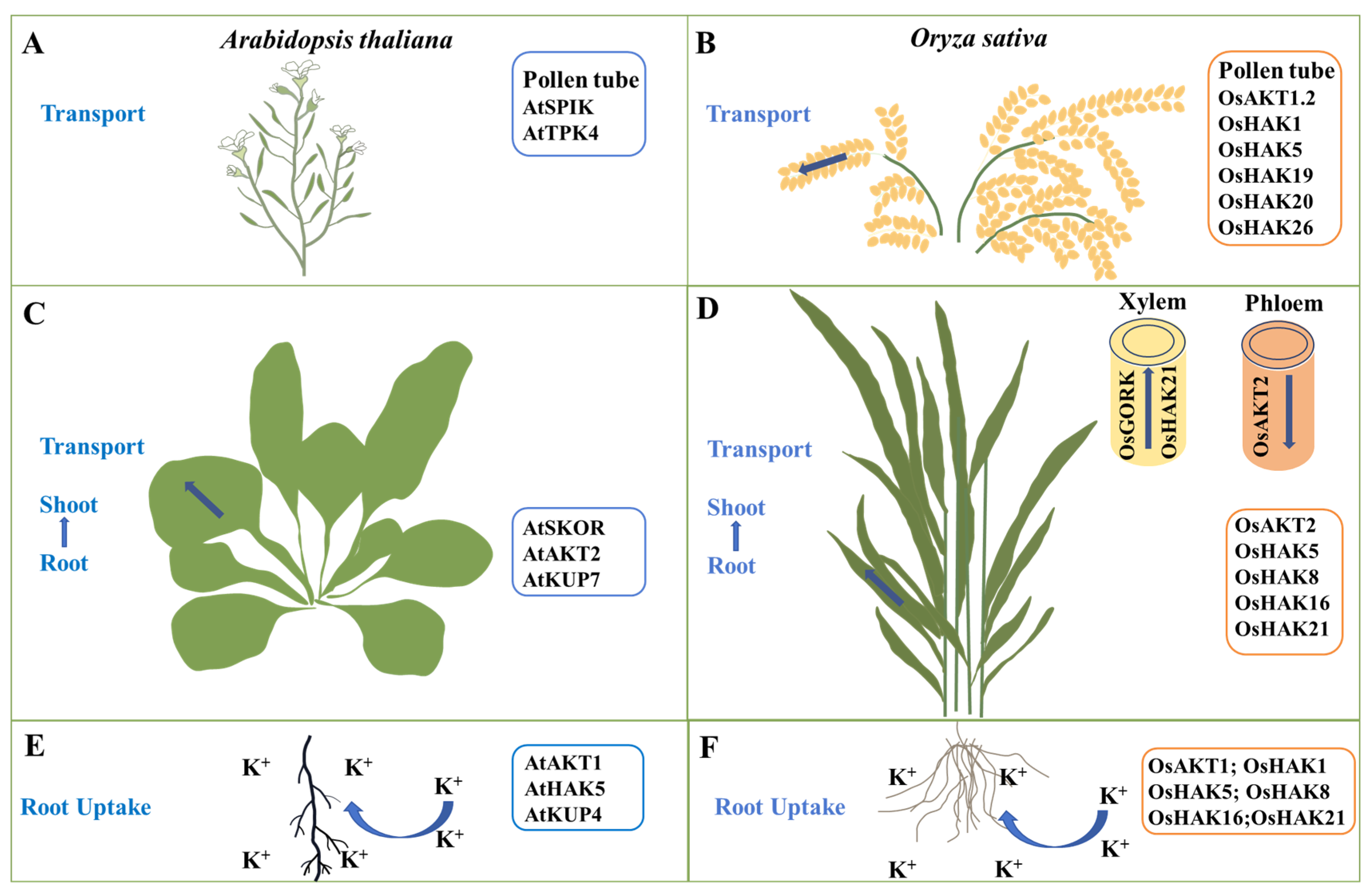
2. Expression Patterns of K+ Channels and Transporters in Rice
3. K+ Absorption and Transport in Rice Roots
3.1. K+ Absorption in Rice Roots
3.2. K+ Transport from Rice Roots to Shoots
4. K+ Transport in Rice Shoots
4.1. K+ Transport in Rice Leaves
4.2. K+ Transport in Rice Guard Cells
4.3. K+ Transport during Rice Pollen Development
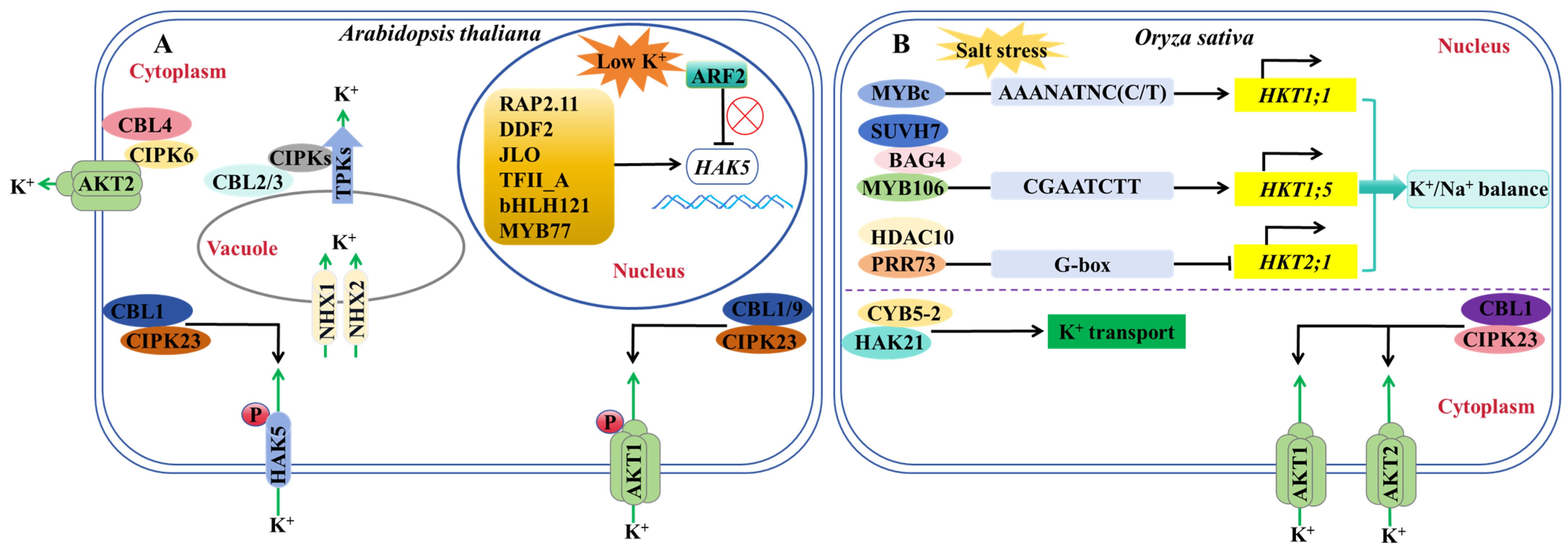
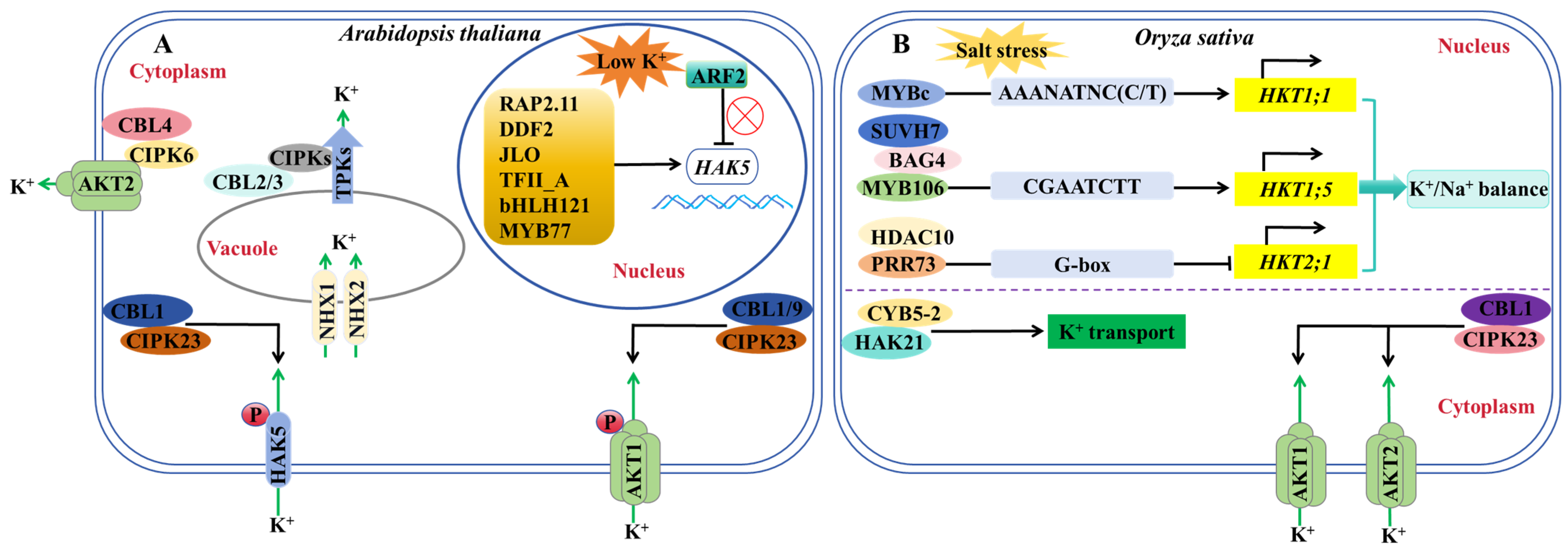
5. Regulation of K+ Channels and Transporters
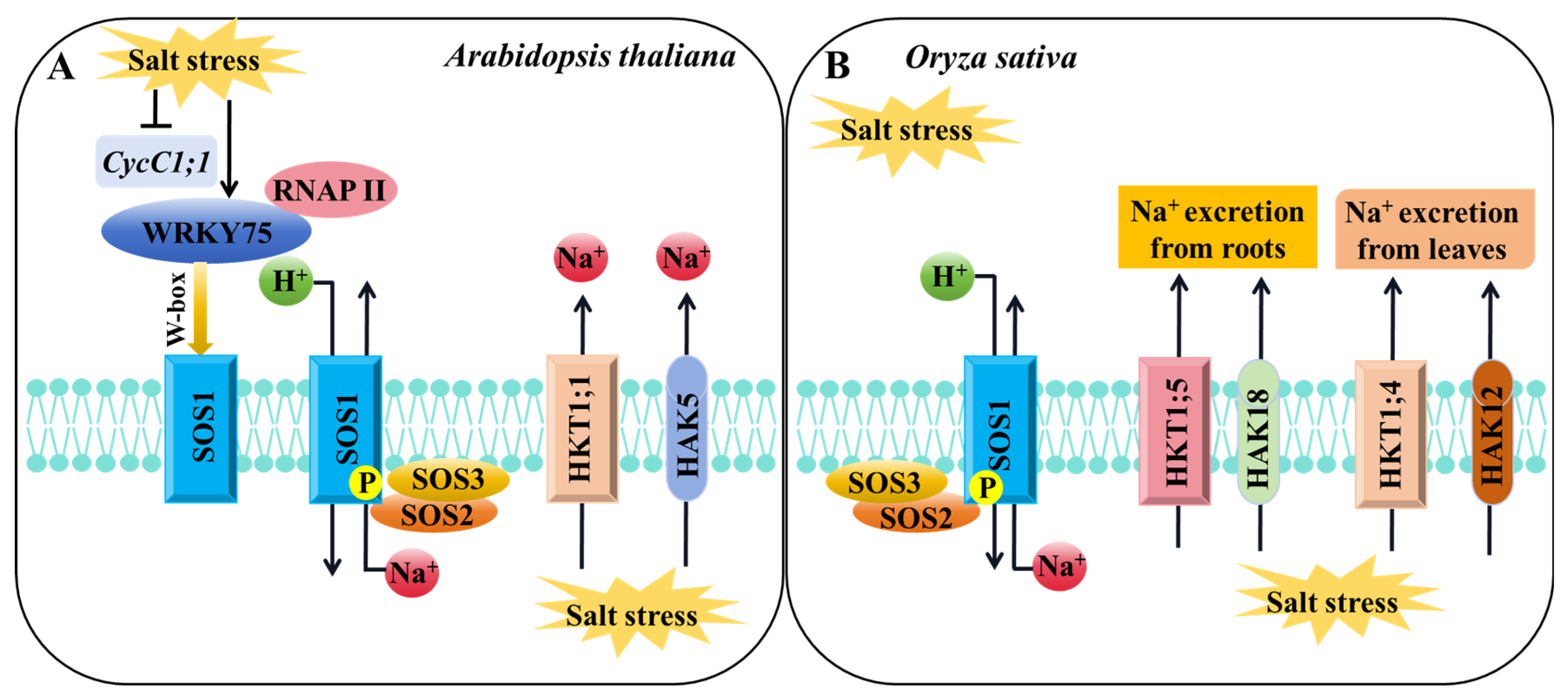
6. Relationship between Potassium and Salt Stress Response in Rice
7. Synergistic Regulation of Potassium with Nitrogen and Phosphorus
7.1. Synergistic Regulation of Potassium and Nitrogen
7.2. Synergistic Regulation of Potassium and Phosphorus
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Very, A.A.; Sentenac, H. Molecular mechanisms and regulation of K+ transport in higher plants. Annu. Rev. Plant Biol. 2003, 54, 575–603. [Google Scholar] [CrossRef]
- Shin, R. Strategies for improveing potassium use efficiency in plants. Mol. Cells 2014, 37, 435–502. [Google Scholar] [CrossRef]
- Clarkson, D.T.; Hanson, J.B. The mineral nutrition of higher plants. Annu. Rev. Plant Physiol. 1980, 31, 239–298. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.S.; Shen, Q.R.; Guo, S.W. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef]
- Pan, Y.H.; Lu, Z.F.; Lu, J.W.; Li, X.K.; Cong, R.H.; Ren, T. Effects of low sink demand on leaf photosynthesis under potassium deficiency. Plant Physiol. Biochem. 2017, 113, 110–121. [Google Scholar] [CrossRef] [PubMed]
- de Bang, T.C.; Husted, S.; Laursen, K.H.; Persson, D.P.; Schjoerring, J.K. The molecular-physiological functions of mineral macronutrients and their consequences for deficiency symptoms in plants. New Phytol. 2021, 229, 2446–2469. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.T.; Long, Y.; He, F.; Zhang, C.Y.; Wang, R.Y.; Zhang, T.; Wu, W.; Hao, Z.Y.; Wang, Y.; Wang, G.L.; et al. The fungal pathogen Magnaporthe oryzae suppresses innate immunity by modulating a host potassium channel. PLoS Pathog. 2018, 14, e1006878. [Google Scholar] [CrossRef]
- Gajdanowicz, P.; Michael, E.; Sandmann, M.; Rocha, M.; Corrêa, L.G.; Ramírez-Aguilar, S.J.; Gomez-Porras, J.L.; González, W.; Thibaud, J.B.; Dongen, J.T.; et al. Potassium (K+) gradients serve as a mobile energy source in plant vascular tissues. Proc. Natl. Acad. Sci. USA 2011, 108, 864–869. [Google Scholar] [CrossRef]
- Wigoda, N.; Moshelion, M.; Moran, N. Is the leaf bundle sheath a smart flux value for K+ nutrition? J. Plant Physiol. 2014, 171, 715–722. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, W.H. Genetic approaches for improvement of the crop potassium acquisition and utilization efficiency. Curr. Opin. Plant Biol. 2015, 25, 46–52. [Google Scholar] [CrossRef]
- Grabov, A. Plant KT/KUP/HAK potassium transporters: Single family-Multiple functions. Ann. Bot. 2007, 99, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Bohm, J.; Messerer, M.; Muller, H.; Scholz-Starke, J.; Gradogna, A.; Scherzer, S.; Maierhofer, T.; Bazihizina, N.; Zhang, H.M.; Stigloher, C.; et al. Understanding the molecular basis of salt sequestration in epidermal bladder cells of Chenopodium quinoa. Curr. Biol. 2018, 28, 3075–3085. [Google Scholar] [CrossRef] [PubMed]
- Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ nutrition in plants. Front. Plant Sci. 2019, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Papazian, D.M.; Schwarz, T.L.; Tempel, B.L.; Jan, L.Y. Cloning of genomic and complementary DNA from shaker, a putative channel gene from Drosphila. Science 1987, 237, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, W.H. Potassium transport and signaling in higher plants. Annu. Rev. Plant Biol. 2013, 64, 451–476. [Google Scholar] [CrossRef]
- Pilot, G.; Gaymard, F.; Mouline, K.; Chérel, I.; Sentenac, H. Regulated expression of Arabidopsis shaker K+ channel genes involved in K+ uptake and distribution in the plant. Plant Mol. Biol. 2003, 51, 773–787. [Google Scholar] [CrossRef] [PubMed]
- Pilot, G.; Pratelli, R.; Gaymard, F.; Meyer, Y.; Sentenac, H. Five-group distribution of the Shaker-like K+ channel family in higher plants. J. Mol. Evol. 2003, 56, 418–434. [Google Scholar] [CrossRef] [PubMed]
- Deeken, R.; Ivashikina, N.; Czirjak, T.; Philippar, K.; Becker, D.; Ache, P.; Hedrich, R. Tumour development in Arabidopsis thaliana involves the Shaker-like K+ channels AKT1 and AKT2/3. Plant J. 2003, 34, 778–787. [Google Scholar] [CrossRef]
- Gaymard, F.; Pilot, G.; Lacombe, B.; Bouchez, D.; Bruneau, D.; Boucherez, J.; Michaux-Ferrière, N.; Thibaud, J.B.; Sentenac, H. Identification and disruption of a plant shaker-like outward channel involved in K+ release into the xylem sap. Cell 1998, 94, 647–655. [Google Scholar] [CrossRef]
- Ronzier, E.; Corratgé-Faillie, C.; Sanchez, F.; Prado, K.; Brière, C.; Leonhardt, N.; Thibaud, J.B.; Xiong, T.C. CPK13, a noncanonical Ca2+-dependent protein kinase, specifically inhibits KAT2 and KAT1 shaker K+ channels and reduces stomatal opening. Plant Physiol. 2014, 166, 314–326. [Google Scholar] [CrossRef]
- Li, J.; Long, Y.; Qi, G.N.; Li, J.; Xu, Z.J.; Wu, W.H.; Wang, Y. The OsAKT1 channel is critical for K+ uptake in rice roots and is modulated by the rice CBL1-CIPK23 complex. Plant Cell 2014, 26, 3387–3402. [Google Scholar] [CrossRef]
- Ahmad, I.; Mian, A.; Maathuis, F.J.M. Overexpression of the rice AKT1 potassium channel affects potassium nutrition and rice drought tolerance. J. Exp. Bot. 2016, 67, 2689–2698. [Google Scholar] [CrossRef]
- Tian, Q.X.; Shen, L.K.; Luan, J.X.; Zhou, Z.Z.; Guo, D.S.; Shen, Y.; Jing, W.; Zhang, B.L.; Zhang, Q.; Zhang, W.H. Rice shaker potassium channel OsAKT2 positively regulates salt tolerance and grain yield by mediating K+ redistribution. Plant Cell Environ. 2021, 44, 2951–2965. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Choi, E.H.; Min, M.K.; Hwang, H.; Moon, S.J.; Yoon, I.; Byun, M.O.; Kim, B.G. Differential gene expression of two outward-rectifying shaker-like potassium channels OsSKOR and OsGORK in rice. J. Plant Biol. 2015, 58, 230–235. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Huang, S.G.; Meynard, D.; Chaine, C.; Michel, R.; Roelfsema, M.; Guiderdoni, E.; Sentenac, H.; Véry, A.A. A dual role for the OsK5.2 ion channel in stomatal movements and K+ loading into xylem sap. Plant Physiol. 2017, 174, 2409–2418. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.; Yoon, J.; Kim, H.Y.; Min, M.K.; Kim, J.A.; Choie, E.H.; Lan, W.Z.; Bae, Y.M.; Luan, S.; Cho, H.; et al. Unique features of two potassium channels, OsKAT2 and OsKAT3, expressed in rice guard cells. PLoS ONE 2013, 8, e72541. [Google Scholar] [CrossRef]
- Li, J.F.; Shen, L.K.; Han, X.L.; He, G.F.; Fan, W.X.; Li, Y.; Yang, S.P.; Zhang, Z.D.; Yang, Y.Q.; Jin, W.W.; et al. Phosphatidic acid-regulated SOS2 controls sodium and potassium homeostasis in Arabidopsis under salt stress. EMBO J. 2023, 42, e112401. [Google Scholar] [CrossRef]
- Voelker, C.; Schmidt, D.; Mueller-Roeber, B.; Czempinski, K. Members of the Arabidopsis AtTPK/KCO family form homomeric vacuolar channels in planta. Plant J. 2006, 48, 296–306. [Google Scholar] [CrossRef]
- Isayenkov, S.; Isner, J.C.; Maathuis, F.J. Rice two-pore K+ channels are expressed in different types of vacuoles. Plant Cell 2011, 23, 756–768. [Google Scholar] [CrossRef]
- Ahn, S.J.; Shin, R.; Schachtman, D.P. Expression of KT/KUP genes in Arabidopsis and the role of root hairs in K+ uptake. Plant Physiol. 2004, 134, 1135–1145. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Lara, A.; Ródenas, R.; Amo, J.; Rivero, R.M.; Martínez, V.; Rubio, F. Modulation of K+ translocation by AKT1 and AtHAK5 in Arabidopsis plants. Plant Cell Environ. 2019, 42, 2357–2371. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Hampton, C.R.; Shin, R.; Barkla, B.J.; White, P.J.; Schachtman, D.P. The high affinity K+ transporter AtHAK5 plays a physiological role in planta at very low K+ concentrations and provides a caesium uptake pathway in Arabidopsis. J. Exp. Bot. 2008, 59, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Pyo, Y.J.; Gierth, M.; Schroeder, J.I.; Cho, M.H. High-affinity K+ transport in Arabidopsis: AtHAK5 and AKT1 are vital for seedling establishment and postgermination growth under low-potassium conditions. Plant Physiol. 2010, 153, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.L.; Kong, X.P.; Li, C.L.; Liu, Y.X.; Ding, Z.J. Potassium retention under salt stress is associated with natural variation in salinity tolerance among Arabidopsis accessions. PLoS ONE 2015, 10, e0124032. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Yamane, S.; Yamaguchi, M.; Kato, K.; Shinmyo, A.; Tsunemitsu, Y.; Iwasaki, K.; Ueno, D.; Demura, T. Characterization of rice KT/HAK/KUP potassium transporters and K+ uptake by HAK1 from Oryza sativa. Plant Biotechnol. 2018, 35, 101–111. [Google Scholar] [CrossRef]
- Chen, G.; Hu, Q.D.; Luo, L.; Yang, T.Y.; Zhang, S.; Hu, Y.B.; Yu, L.; Xu, G.H. Rice potassium transporter OsHAK1 is essential for maintaining potassium-mediated growth and functions in salt tolerance over low and high potassium concentration ranges. Plant Cell Environ. 2015, 38, 2747–2765. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhang, Y.; Ruan, B.P.; Guo, L.B.; Zeng, D.L.; Gao, Z.Y.; Zhu, L.; Hu, J.; Ren, D.Y.; Yu, L.; et al. OsHAK1 controls the vegetative growth and panicle fertility of rice by its effect on potassium-mediated sugar metabolism. Plant Sci. 2018, 274, 261–270. [Google Scholar] [CrossRef]
- Yang, T.Y.; Zhang, S.; Hu, Y.B.; Wu, F.C.; Hu, Q.D.; Chen, G.; Cai, J.; Wu, T.; Moran, N.; Yu, L.; et al. The role of a potassium transporter OsHAK5 in potassium acquisition and transport from roots to shoots in rice at low potassium supply levels. Plant Physiol. 2014, 166, 945–959. [Google Scholar] [CrossRef]
- Horie, T.; Sugawara, M.; Okada, T.; Taira, K.; Kaothien-Nakayama, P.; Katsuhara, M.; Shinmyo, A.; Nakayama, H. Rice sodium-insensitive potassium transporter, OsHAK5, confers increased salt tolerance in tobacco BY2 cells. J. Biosci. Bioeng. 2011, 111, 346–356. [Google Scholar] [CrossRef]
- Bañuelos, M.A.; Garciadeblas, B.; Cubero, B.; Rodrıguez-Navarro, A. Inventory and functional characterization of the HAK potassium transporters of rice. Plant Physiol. 2002, 130, 784–795. [Google Scholar] [CrossRef]
- Wang, X.H.; Li, J.F.; Li, F.; Pan, Y.; Cai, D.; Mao, D.D.; Chen, L.B.; Luan, S. Rice potassium transporter OsHAK8 mediates K+ uptake and translocation in response to low K+ stress. Front. Plant Sci. 2021, 12, 730002. [Google Scholar] [CrossRef]
- Zhang, L.N.; Sun, X.Y.; Li, Y.F.; Luo, X.; Song, S.W.; Chen, Y.; Wang, X.H.; Mao, D.D.; Chen, L.B.; Luan, S. Rice Na+-permeable transporter OsHAK12 mediates shoots Na+ exclusion in response to salt stress. Front. Plant Sci. 2021, 12, 771746. [Google Scholar] [CrossRef]
- Feng, H.M.; Tang, Q.; Cai, J.; Xu, B.C.; Xu, G.H.; Yu, L. Rice OsHAK16 functions in potassium uptake and translocation in shoot, maintaining potassium homeostasis and salt tolerance. Planta 2019, 250, 549–561. [Google Scholar] [CrossRef]
- Peng, L.R.; Xiao, H.J.; Li, R.; Zeng, Y.; Gu, M.; Moran, N.; Yu, L.; Xu, G.H. Potassium transporter OsHAK18 mediates potassium and sodium circulation and sugar translocation in rice. Plant Physiol. 2023, 193, 2003–2020. [Google Scholar] [CrossRef]
- Shen, Y.; Shen, L.K.; Shen, Z.X.; Jing, W.; Ge, H.L.; Zhao, J.Z.; Zhang, W.H. The potassium transporter OsHAK21 functions in the maintenance of ion homeostasis and tolerance to salt stress in rice. Plant Cell Environ. 2015, 38, 2766–2779. [Google Scholar] [CrossRef]
- Li, W.H.; Li, M.Q.; Li, S.; Zhang, Y.F.; Li, X.; Xu, G.H.; Yu, L. Function of rice high-affinity potassium transporters in pollen development and fertility. Plant Cell Physiol. 2022, 63, 967–980. [Google Scholar] [CrossRef]
- Almeida, P.; Katschnig, D.; de Boer, A.H. HKT transporters--state of the art. Int. J. Mol. Sci. 2013, 14, 20359–20385. [Google Scholar] [CrossRef]
- Wang, R.; Jing, W.; Xiao, L.Y.; Jin, Y.K.; Shen, L.K.; Zhang, W.H. The rice high-affinity potassium transporter1;1 is involved in salt tolerance and regulated by an MYB-Type transcription factor. Plant Physiol. 2015, 168, 1076–1090. [Google Scholar] [CrossRef]
- Garciadeblás, B.; Senn, M.E.; Bañuelos, M.A.; Navarro, A.R. Sodium transport and HKT transporters: The rice model. Plant J. 2003, 34, 788–801. [Google Scholar] [CrossRef]
- Suzuki, K.; Yamaji, N.; Costa, A.; Okuma, E.; Kobayashi, N.I.; Kashiwagi, T.; Katsuhara, M.; Wang, C.; Tanoi, K.; Murata, Y.; et al. OsHKT1;4-mediated Na+ transport in stems contributes to Na+ exclusion from leaf blades of rice at the reproductive growth stage upon salt stress. BMC Plant Biol. 2016, 16, 22. [Google Scholar] [CrossRef]
- Ren, Z.H.; Gao, J.P.; Li, L.G.; Cai, X.L.; Huang, W.; Chao, D.Y.; Zhu, M.Z.; Wang, Z.Y.; Luan, S.; Lin, H.X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef]
- Horie, T.; Costa, A.; Kim, T.H.; Han, M.J.; Horie, R.; Leung, H.Y.; Miyao, A.; Hirochika, H.; An, G.; Schroeder, J.I. Rice OsHKT2;1 transporter mediates large Na+ influx component into K+-starved roots for growth. EMBO J. 2007, 26, 3003–3014. [Google Scholar] [CrossRef]
- Lan, W.Z.; Wang, W.; Wang, S.M.; Li, L.G.; Buchanan, B.B.; Lin, H.X.; Gao, J.P.; Luan, S. A rice high-affinity potassium transporter (HKT) conceals a calcium-permeable cation channel. Proc. Natl. Acad. Sci. USA 2010, 107, 7089–7094. [Google Scholar] [CrossRef]
- Yokoi, S.; Quintero, F.J.; Cubero, B.; Ruiz, M.T.; Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M. Differential expression and function of Arabidopsis thaliana NHX Na+/H+ antiporters in the salt stress response. Plant J. 2002, 30, 529–539. [Google Scholar] [CrossRef]
- Fukuda, A.; Nakamura, A.; Hara, N.; Toki, S.; Tanaka, Y. Molecular and functional analyses of rice NHX-type Na+ /H+ antiporter genes. Planta 2011, 233, 175–188. [Google Scholar] [CrossRef]
- Steudle, E. The cohesion-tension mechanism and the acquisition of water by plant roots. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 847–875. [Google Scholar] [CrossRef]
- Hosmani, P.S.; Kamiya, T.; Danku, J.; Naseer, S.; Geldner, N.; Guerinot, M.L.; Salt, D.E. Dirigent domain-containing protein is part of the machinery required for formation of the lignin-based Casparian strip in the root. Proc. Natl. Acad. Sci. USA 2013, 110, 14498–14503. [Google Scholar] [CrossRef]
- Kamiya, T.; Borghi, M.; Wang, P.; Danku, J.M.C.; Kalmbach, L.; Hosmani, P.S.; Naseer, S.; Fujiwara, T.; Geldner, N.; Salt, D.E. The MYB36 transcription factor orchestrates Casparian strip formation. Proc. Natl. Acad. Sci. USA 2015, 112, 10533–10538. [Google Scholar] [CrossRef]
- Wu, Q.; Feng, Z.; Tsukagoshi, H.; Yang, M.; Ao, Y.; Fujiwara, T.; Kamiya, T. Early differentiation of Casparian strip mediated by nitric oxide is required for efficient K transport under low K conditions in Arabidopsis. Plant J. 2023, 116, 467–477. [Google Scholar] [CrossRef]
- Geldner, N. The endodermis. Annu. Rev. Plant Biol. 2013, 64, 531–558. [Google Scholar] [CrossRef]
- Tagliani, A.; Tran, A.N.; Novi, G.; Mambro, R.D.; Pesenti, M.; Sacchi, G.A.; Perata, P.; Pucciariello, C. The calcineurin β-like interacting protein kinase CIPK25 regulates potassium homeostasis under low oxygen in Arabidopsis. J. Exp. Bot. 2020, 71, 2678–2689. [Google Scholar] [CrossRef] [PubMed]
- Epstein, E.; Rains, D.W.; Elzam, O.E. Resolution of dual mechanisms of potassium absorption by barley roots. Proc. Natl. Acad. Sci. USA 1963, 49, 684–692. [Google Scholar] [CrossRef]
- Gireth, M.; Maser, P.; Schroeder, J.I. The potassium transpoters AtHAK5 functions in K+ deprivation-induced high-affinity K+ uptake and AKT1 K+ channel contribution to K+ uptake kinetics in Arabidopsis roots. Plant Physiol. 2005, 137, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Kochian, L.V.; Lucas, W.J. Potassium transport in corn roots: I. resolution of kinetics into a saturable and linear component. Plant Physiol. 1982, 70, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Maathuis, F.J.M. Physiological functions of mineral macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, D.; Basset, M.; Lepetit, M.; Conejero, G.; Gaymard, F.; Astruc, S.; Grignon, C. Tissue-specific expression of Arabidopsis AKT1 gene is consistent with a role in K+ nutrition. Plant J. 1996, 9, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Maathuis, F.J.M. Cellular and tissue distribution of potassium: Physiological relevance, mechanisms and regulation. J. Plant Physiol. 2014, 171, 708–714. [Google Scholar] [CrossRef]
- Liu, K.; Li, L.G.; Luan, S. Intracellular K+ sensing of SKOR, a Shaker-type K+ channel from Arabidopsis. Plant J. 2006, 46, 260–268. [Google Scholar] [CrossRef]
- Drechsler, N.; Zheng, Y.; Bohner, A.; Nobmann, B.; von Wirén, N.; Kunze, R.; Rausch, C. Nitrate-dependent control of shoot K homeostasis by the nitrate transporter1/peptide transporter family member NPF7.3/NRT1.5 and the stelar K+ outward rectifier SKOR in Arabidopsis. Plant Physiol. 2015, 169, 2832–2847. [Google Scholar] [CrossRef]
- Han, M.; Wu, W.; Wu, W.H.; Wang, Y. Potassium transporter KUP7 is involved in K+ acquisition and translocation in Arabidopsis root under K+-limited conditions. Mol. Plant. 2016, 9, 437–446. [Google Scholar] [CrossRef]
- Xia, X.D.; Fan, X.R.; Wei, J.; Feng, H.M.; Qu, H.Y.; Xie, D.; Miller, A.J.; Xu, G.H. Rice nitrate transporter OsNPF2.4 functions in low-affinity acquisition and long-distance transport. J. Exp. Bot. 2015, 66, 317–331. [Google Scholar] [CrossRef]
- Cuin, T.A.; Dreyer, I.; Michard, E. The role of potassium channels in Arabidopsis thaliana long distance electrical signalling: AKT2 modulates tissue excitability while GORK shapes action potentials. Int. J. Mol. Sci. 2018, 19, 926. [Google Scholar] [CrossRef] [PubMed]
- Held, K.; Pascaud, F.; Eckert, C.; Gajdanowicz, P.; Hashimoto, K.; Corratgé-Faillie, C.; Offenborn, J.N.; Lacombe, B.; Dreyer, I.; Thibaud, J.B.; et al. Calcium-dependent modulation and plasma membrane targeting of the AKT2 potassium channel by the CBL4/CIPK6 calcium sensor/protein kinase complex. Cell Res. 2011, 21, 1116–1130. [Google Scholar] [CrossRef]
- Kwak, J.M.; Murata, Y.; Baizabal-Aguirre, V.M.; Merrill, J.; Wang, M.; Kemper, A.; Hawke, S.D.; Tallman, G.; Schroeder, J.I. Dominant negative guard cell K+ channel mutants reduce inward-rectifying K+ currents and light-induced stomatal opening in Arabidopsis. Plant Physiol. 2001, 127, 473–485. [Google Scholar] [CrossRef]
- Pilot, G.; Lacombe, B.; Gaymard, F.; Chérel, I.; Boucherez, J.; Thibaud, J.B.; Sentenac, H. Guard cell inward K+ channel activity in Arabidopsis involves expression of the twin channel subunits KAT1 and KAT2. J. Biol. Chem. 2001, 276, 3215–3221. [Google Scholar] [CrossRef] [PubMed]
- Hosy, E.; Vavasseur, A.; Mouline, K.; Dreyer, I.; Gaymard, F.; Porée, F.; Boucherez, J.; Lebaudy, A.; Bouchez, D.; Véry, A.A.; et al. The Arabidopsis outward K+ channel GORK is involved in regulation of stomatal movements and plant transpiration. Proc. Natl. Acad. Sci. USA 2003, 100, 5549–5554. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Dobritsa, A.A. Exine and aperture patterns on the pollen surface: Their formation and roles in plant reproduction. Annu. Plant Rev. 2018, 1, 589–628. [Google Scholar] [CrossRef]
- Lu, Y.X.; Chanroj, S.; Zulkifli, L.; Johnson, M.A.; Uozumi, N.; Sez, C.H. Pollen tubes lacking a pair of K+ transporters fail to target ovules in Arabidopsis. Plant Cell 2011, 23, 81–93. [Google Scholar] [CrossRef]
- Padmanaban, S.; Czerny, D.D.; Levin, K.A.; Leydon, A.R.; Su, R.T.; Maugel, T.K.; Zou, Y.; Chanroj, S.; Cheung, A.Y.; Johnson, M.A.; et al. Transporters involved in pH and K+ homeostasis affect pollen wall formation, male fertility, and embryo development. J. Exp. Bot. 2017, 68, 3165–3178. [Google Scholar] [CrossRef]
- Zhao, L.N.; Shen, L.K.; Zhang, W.Z.; Zhang, W.; Wang, Y.; Wu, W.H. Ca2+-dependent protein kinase11 and 24 modulate the activity of the inward rectifying K+ channels in Arabidopsis pollen tubes. Plant Cell 2013, 25, 649–661. [Google Scholar] [CrossRef]
- Li, D.D.; Guan, H.; Li, F.; Liu, C.Z.; Dong, Y.X.; Zhang, X.S.; Gao, X.Q. Arabidopsis shaker pollen inward K+ channel SPIK functions in SnRK1 complex-regulated pollen hydration on the stigma. J. Interg. Plant Biol. 2017, 59, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Amien, S.; Kliwer, I.; Marton, M.L.; Debener, T.; Geiger, D.; Becker, D.; Dresselhaus, T. Defensin-like ZmES4 mediates pollen tube burst in maize via opening of the potassium channel KZM1. PLoS Biol. 2010, 8, e1000388. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, T.; Liu, L.T. Pollen germination is impaired by disruption of a Shaker K+ channel OsAKT1.2 in rice. J. Plant Physiol. 2020, 248, 153140. [Google Scholar] [CrossRef]
- Becker, D.; Geiger, D.; Dunkel, M.; Roller, A.; Bertl, A.; Latz, A.; Carpaneto, A.; Dietrich, P.; Roelfsema, M.R.G.; Voelker, C.; et al. AtTPK4, an Arabidopsis tandem-pore K+ channel, poised to control the pollen membrane voltage in a pH-and Ca2+-dependent manner. Proc. Natl. Acad. Sci. USA 2004, 101, 15621–15626. [Google Scholar] [CrossRef] [PubMed]
- Haque, U.S.; Elias, S.M.; Jahan, I.; Seraj, Z.I. Functional genomic analysis of K+ related salt-responsive transporters in tolerant and sensitive genotypes of rice. Front. Plant Sci. 2023, 13, 1089109. [Google Scholar] [CrossRef]
- Liu, L.T.; Zheng, C.H.; Kuang, B.J.; Wei, L.Q.; Yan, L.F.; Wang, T. Receptor-like kinase RUPO interacts with potassium transporters to regulate pollen tube growth and integrity in rice. PLoS Genet. 2016, 12, e1006085. [Google Scholar] [CrossRef] [PubMed]
- Li, L.G.; Kim, B.G.; Cheong, Y.H.; Pandey, G.K.; Luan, S. A Ca2+ signaling pathway regulates a K+ channel for low-K response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 12625–12630. [Google Scholar] [CrossRef]
- Xu, J.; Li, H.D.; Chen, L.Q.; Wang, Y.; Liu, L.L.; He, L.; Wu, W.H. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 in Arabidopsis. Cell 2006, 125, 1347–1360. [Google Scholar] [CrossRef]
- Tang, R.J.; Zhao, F.G.; Yang, Y.; Wang, C.; Li, K.L.; Kleist, T.J.; Lemaux, P.G.; Luan, S. A calcium signalling network activates vacuolar K+ remobilization to enable plant adaptation to low-K environments. Nat. Plants 2020, 6, 384–393. Available online: https://www.nature.com/articles/s41477-020-0621-7 (accessed on 1 January 2020). [CrossRef]
- Ragel, P.; Ródenas, R.; García-Martín, E.; Andrés, Z.; Villalta, I.; Nieves-Cordones, M.; Rivero, R.M.; Martínez, V.; Pardo, J.M.; Quintero, F.J.; et al. CIPK23 regulates HAK5-mediated high-affinity K+ uptake in Arabidopsis roots. Plant Physiol. 2015, 169, 2863–2873. [Google Scholar] [CrossRef]
- Santa-María, G.E.; Oliferuk, S.; Moriconi, J.I. KT-HAK-KUP transporters in major terrestrial photosynthetic organisms: A twenty years tale. J. Plant Physiol. 2018, 226, 77–90. [Google Scholar] [CrossRef]
- Lhamo, D.; Wang, C.; Gao, Q.F.; Luan, S. Recent advances in genome-wide analyses of plant potassium transporter families. Curr. Genom. 2021, 22, 164–180. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, M.L.; Ma, T.L.; Wang, Y. Phosphorylation of ARF2 relieves its repression of transcription of the K+ transporter gene HAK5 in response to low potassium stress. Plant Cell 2016, 28, 3005–3019. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.Z.; Luo, Y.X.; Wang, P.D.; Gilliham, M.; Long, Y. MYB77 regulates high-affinity potassium uptake by promoting expression of HAK5. New Phytol. 2021, 232, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Du, X.Q.; Wang, F.L.; Li, H.; Jing, S.; Yu, M.; Li, J.G.; Wu, W.H.; Kudla, J.; Wang, Y. The transcription factor MYB59 regulates K+/ translocation in the Arabidopsis response to low K+ stress. Plant Cell 2019, 31, 699–714. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Z.; Liu, J.; Chen, S.J.; Wang, P.F.; Liu, H.T.; Dong, J.; Zheng, Y.X.; Xie, Y.X.; Wang, C.Y.; Guo, T.C.; et al. Melatonin promotes potassium deficiency tolerance by regulating HAK1 transporter and its upstream transcription factor NAC71 in wheat. J. Pineal Res. 2021, 70, e12727. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Cong, D.L.; Ju, H.Y. Functional characterization of ZmHAK1 promoter and its regulatory transcription factors in maize. Mol. Biol. 2020, 54, 374–388. [Google Scholar] [CrossRef]
- Huang, Y.N.; Yang, S.Y.; Li, J.L.; Wang, S.F.; Wang, J.J.; Hao, D.L.; Su, Y.H. The rectification control and physiological relevance of potassium channel OsAKT2. Plant Physiol. 2021, 187, 2296–2310. [Google Scholar] [CrossRef] [PubMed]
- Song, T.Z.; Shi, Y.Y.; Shen, L.K.; Cao, C.J.; Shen, Y.; Jing, W.; Tian, Q.X.; Lin, F.; Li, W.Y.; Zhang, W.H. An endoplasmic reticulum-localized cytochrome b5 regulates high-affinity K+ transport in response to salt stress in rice. Proc. Natl. Acad. Sci. USA 2021, 118, e2114347118. [Google Scholar] [CrossRef]
- Riedelsberger, J.; Miller, J.K.; Valdebenito-Maturana, B.; Piñeros, M.A.; González, W.; Dreyer, I. Plant HKT channels: An updated view on structure, function and gene regulation. Int. J. Mol. Sci. 2021, 22, 1892. [Google Scholar] [CrossRef]
- Wang, J.; Nan, N.; Li, N.; Liu, Y.T.; Wang, T.J.; Hwang, I.; Liu, B.; Xu, Z.Y. A DNA methylation reader-chaperone regulator-transcription factor complex activates OsHKT1;5 expression during salinity stress. Plant Cell 2020, 32, 3535–3558. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.Y.; Shi, Y.Y.; Wang, R.; Feng, Y.; Wang, L.S.; Zhang, H.S.; Shi, X.Y.; Jing, G.Q.; Deng, P.; Song, T.Z.; et al. The factor OsMYBc and an E3 ligase regulate expression of a K+ transporter during salt stress. Plant Physiol. 2022, 190, 843–859. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Anschutz, U.; Becker, D.; Shabala, S. Going beyond nutrition: Regulation of potassium homoeostasis as a common denominator of plant adaptive responses to environment. J. Plant Physiol. 2014, 171, 670–687. [Google Scholar] [CrossRef]
- Ismail, A.M.; Horie, T. Genomics, physiology, and molecular breeding approaches for improving salt tolerance. Annu. Rev. Plant Biol. 2017, 68, 405–434. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liang, X.Y.; Wang, L.M.; Cao, Y.B.; Song, W.B.; Shi, J.P.; Lai, J.S.; Jiang, C.F. A HAK family Na+ transporter confers natural variation of salt tolerance in maize. Nat. Plants 2019, 5, 1297–1308. Available online: https://www.nature.com/articles/s41477-019-0565-y (accessed on 1 December 2019). [CrossRef] [PubMed]
- Cuin, T.A.; Bose, J.; Stefano, G.; Jha, D.; Tester, D.; Mancuso, S.; Shabala, S. Assessing the role of root plasma membrane and tonoplast Na+/H+ exchangers in salinity tolerance in wheat: In planta quantification methods. Plant Cell Environ. 2011, 34, 947–961. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Wang, L.L.; Liu, Y.L.; Zhang, Q.; Wei, Q.P.; Zhang, W.H. Nitric oxide enhances salt tolerance in maize seedlings through increasing activities of proton-pump and Na+/H+ antiport in the tonoplast. Planta 2006, 224, 545–555. [Google Scholar] [CrossRef]
- Mian, A.; Oomen, R.J.F.J.; Isayenkov, S.; Sentenac, I.H.; Maathuis, F.J.M.; Véry, A.A. Over-expression of an Na+- and K+-permeable HKT transporter in barley improves salt tolerance. Plant J. 2011, 68, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.M.; Waner, D.A.; Horie, T.; Li, S.L.; Horie, R.; Abid, K.B.; Schroeder, J.I. Microarray-based rapid cloning of an ion accumulation deletion mutant in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2004, 101, 15404–15409. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, X.L.; He, Y.Q.; Xu, H.; Wang, L. Clock component OsPRR73 positively regulates rice salt tolerance by modulating OsHKT2;1-mediated sodium homeostasis. EMBO J. 2021, 40, e105086. [Google Scholar] [CrossRef]
- Davenport, R.J.; Muñoz-Mayor, A.; Jha, D.; Essah, P.A.; Rus, A.; Tester, M. The Na+ transporter AtHKT1;1 controls retrieval of Na+ from the xylem in Arabidopsis. Plant Cell Environ. 2007, 30, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Sunarpi, X.; Horie, T.; Motoda, J.; Kubo, M.; Yang, H.; Yoda, K.; Horie, R.; Chan, W.Y.; Leung, H.Y.; Hattori, K.; et al. Enhanced salt tolerance mediated by AtHKT1 transporter-induced Na unloading from xylem vessels to xylem parenchyma cells. Plant J. 2005, 44, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Pruthi, R.; Chapagain, S.; Coronejo, S.; Singh, L.; Subudhi, P.K. Quantitative trait loci, candidate genes, and breeding lines to improve salt tolerance at the flowering and seedling stages in rice. Food Energy Secur. 2022, 00, e433. [Google Scholar] [CrossRef]
- Obata, T.; Kitamoto, H.K.; Nakamura, A.; Fukuda, A.; Tanaka, Y. Rice shaker potassium channel OsKAT1 confers tolerance to salinity stress on yeast and rice cells. Plant Physiol. 2007, 144, 1978–1985. [Google Scholar] [CrossRef]
- Leidi, E.O.; Barragán, V.; Rubio, L.; El-Hamdaoui, A.; Ruiz, M.T.; Cubero, B.; Fernández, J.A.; Bressan, R.A.; Hasegawa, P.M.; Quintero, F.J.; et al. The AtNHX1 exchanger mediates potassium compartmentation in vacuoles of transgenic tomato. Plant J. 2010, 61, 495–506. [Google Scholar] [CrossRef]
- Barragán, V.; Leidi, E.O.; Andrés, Z.; Rubio, L.; Luca, A.D.; Fernández, J.A.; Cubero, B.; Pardo, J.M. Ion exchangers NHX1 and NHX2 mediate active potassium uptake into vacuoles to regulate cell turgor and stomatal function in Arabidopsis. Plant Cell 2012, 24, 1127–1142. [Google Scholar] [CrossRef]
- Bassil, E.; Ohto, M.; Esumi, T.; Tajima, H.; Zhu, Z.; Cagnac, O.; Belmonte, M.; Peleg, Z.; Yamaguchi, T.; Blumwald, E. The Arabidopsis intracellular Na+/H+ antiporters NHX5 and NHX6 are endosome associated and necessary for plant growth and development. Plant Cell 2011, 23, 224–239. [Google Scholar] [CrossRef]
- Zelm, E.V.; Zhang, Y.X.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Halfter, U.; Ishitani, M.; Zhu, J.K. The Arabidopsis SOS2 protein kinase physically interacts with and is activated by the calcium-binding protein SOS3. Proc. Natl. Acad. Sci. USA 2000, 97, 3735–3740. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.K.; Song, R.F.; Guo, J.X.; Zhang, Y.; Zuo, J.X.; Chen, H.H.; Liao, C.Y.; Hu, X.Y.; Ren, F.; Lu, Y.T.; et al. CycC1;1-WRKY75 complex-mediated transcriptional regulation of SOS1 controls salt stress tolerance in Arabidopsis. Plant Cell 2023, 35, 2570–2591. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Tang, L.H.; Nie, J.W.; Zhang, C.R.; Han, X.N.; Li, Q.Y.; Qin, L.; Wang, M.H.; Huang, X.H.; Yu, F.F.; et al. Structure and activation mechanism of the rice Salt Overly Sensitive 1 (SOS1) Na+/H+ antiporter. Nat. Plants 2023, 9, 1924–1936. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.J.; Li, G.H.; Lu, W.P.; Lu, D.L. Interactive effects of nitrogen and potassium on grain yield and quality of waxy maize. Plants 2022, 11, 2528. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Peng, J.S.; He, Y.N.; Zhang, G.B.; Yi, H.Y.; Fu, Y.L.; Gong, J.M. Arabidopsis NRT1.5 mediates the suppression of nitrate starvation-induced leaf senescence by modulating foliar potassium level. Mol. Plant. 2016, 9, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Straub, T.; Ludewig, U.; Neuhäuser, B. The Kinase CIPK23 inhibits ammonium transport in Arabidopsis thaliana. Plant Cell 2017, 29, 409–422. [Google Scholar] [CrossRef]
- Bao, A.L.; Liang, Z.J.; Zhao, Z.Q.; Cai, H.M. Overexpressing of OsAMT1-3, a high affinity ammonium transporter gene, modifies rice growth and carbon-nitrogen metabolic status. Int. J. Mol. Sci. 2015, 16, 9037–9063. [Google Scholar] [CrossRef]
- Wei, J.; Zheng, Y.; Feng, H.M.; Qu, H.Y.; Fan, X.R.; Yamaji, N.; Ma, J.F.; Xu, G.H. OsNRT2.4 encodes a dual-affinity nitrate transporter and functions in nitrate-regulated root growth and nitrate distribution in rice. J. Exp. Bot. 2018, 69, 1095–1107. [Google Scholar] [CrossRef]
- Dai, S.H.; Wu, H.C.; Chen, H.Y.; Wang, Z.H.; Yu, X.; Wang, L.; Jia, X.Q.; Qin, C.; Zhu, Y.Y.; Yi, K.K.; et al. Comparative transcriptome analyses under individual and combined nutrient starvations provide insights into N/P/K interactions in rice. Plant Physiol. Bioch. 2023, 197, 107642. [Google Scholar] [CrossRef]
- Wang, Y.H.; Garvin, D.F.; Kochian, L.V. Rapid induction of regulatory and transporter genes in response to phosphorus, potassium, and iron deficiencies in tomato roots. Evidence for cross talk and root/rhizosphere-mediated signals. Plant Physiol. 2002, 130, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Sun, Y.F.; Pei, W.X.; Jain, A.; Sun, R.; Cao, Y.; Wu, X.N.; Jiang, T.T.; Zhang, L.; Fan, X.R.; et al. Involvement of OsPht1;4 in phosphate acquisition and mobilization facilitates embryo development in rice. Plant J. 2015, 82, 556–569. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Gene Family | Gene | Gene ID | Expression Tissues | References |
|---|---|---|---|---|
| Shaker K+ channels in Arabidopsis and rice | AtAKT1 | AT2G26650 | Root | [16,17] |
| AtAKT2 | AT4G22200 | Shoot | [18] | |
| AtSKOR | AT3G02850 | Root | [19] | |
| AtGORK | AT5G37500 | Root | [19] | |
| AtKAT1 | AT5G46240 | Shoot | [20] | |
| AtKAT2 | AT4G18290 | Shoot | [20] | |
| AtKAT3 | AT4G32650 | Shoot | [20] | |
| OsAKT1 | Os01g45990 | Root | [21,22] | |
| OsAKT2 | Os05g35410 | Shoot | [23] | |
| OsSKOR | Os04g36740 | Root and flower | [24] | |
| OsGORK | Os06g14030 | Root and flower | [24,25] | |
| OsKAT1 | Os01g55200 | / | [26] | |
| OsKAT2 | Os01g11250 | Shoot and leaf sheath | [26] | |
| OsKAT3 | Os02g14840 | Shoot and leaf sheath | [26] | |
| TPK-type K+ channels in Arabidopsis and rice | AtTPK1 | AT5G55630 | Root and pollen | [28] |
| AtTPK3 | AT4G18160 | Root and pollen | [28] | |
| AtTPK4 | AT1G02510 | Pollen | [28] | |
| AtTPK5 | AT4G01840 | Vascular tissue | [28] | |
| AtKCO3 | AT5G46360 | Vascular tissue | [28] | |
| OsTPKa | Os03g54100 | Root and shoot | [29] | |
| OsTPKb | Os07g01810 | Root and shoot | [29] | |
| KUP/HAK/KT transporters in Arabidopsis and rice | AtKUP2 | AT2G40540 | Flower | [30] |
| AtKUP3 | AT3G02050 | Silique | [30] | |
| AtKUP4 | AT4G23640 | Silique | [30] | |
| AtKUP5 | AT4G33530 | Root and shoot | [30] | |
| AtKUP6 | AT1G70300 | Root and shoot | [30] | |
| AtKUP7 | AT5G09400 | Root and shoot | [30] | |
| AtKUP8 | AT5G14880 | Root and shoot | [30] | |
| AtKUP11 | AT2G35060 | Shoot | [30] | |
| AtKUP12 | At1G60160 | Root and shoot | [30] | |
| AtHAK5 | AT4G13420 | Root | [31,32,33,34] | |
| OsHAK1 | Os04g32920 | Root | [35,36,37] | |
| OsHAK5 | Os01g70490 | Root | [38,39] | |
| OsHAK7 | Os07g47350 | Root and stem | [40] | |
| OsHAK8 | Os03g21890 | Root | [41] | |
| OsHAK12 | Os08g10550 | Root | [42] | |
| OsHAK16 | Os03g37840 | Root | [43] | |
| OsHAK18 | Os09g38960 | Root | [44] | |
| OsHAK21 | Os03g37930 | Root and shoot | [45] | |
| OsHAK26 | Os08g39950 | Anther and seed coat | [46] | |
| HKT transporters in Arabidopsis and rice | AtHKT1;1 | AT4G10310 | Root and shoot | [47] |
| OsHKT1;1 | Os04g51820 | Shoot | [48] | |
| OsHKT1;3 | Os02g07830 | Bud | [49] | |
| OsHKT1;4 | Os04g51830 | Leaf sheath | [50] | |
| OsHKT1;5 | Os01g20160 | Root | [51] | |
| OsHKT2;1 | Os06g48810 | Root | [52] | |
| OsHKT2;4 | Os06g48800 | Root and shoot | [53] | |
| Vacuolar Na+/H+ antiporters in Arabidopsis and rice | AtNHX1 | AT5G27150 | Root and shoot | [54] |
| AtNHX2 | AT3G05030 | Root and shoot | [54] | |
| AtNHX3 | AT5G55470 | Root and shoot | [54] | |
| AtNHX4 | AT3G06370 | Root and shoot | [54] | |
| AtNHX6 | AT1G79610 | Root and shoot | [54] | |
| OsNHX1 | Os07g47100 | Panicle and leaf sheath | [55] | |
| OsNHX2 | Os05g05590 | Panicle and leaf sheath | [55] | |
| OsNHX5 | Os09g11450 | Flag leaf | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lian, W.; Geng, A.; Wang, Y.; Liu, M.; Zhang, Y.; Wang, X.; Chen, G. The Molecular Mechanism of Potassium Absorption, Transport, and Utilization in Rice. Int. J. Mol. Sci. 2023, 24, 16682. https://doi.org/10.3390/ijms242316682
Lian W, Geng A, Wang Y, Liu M, Zhang Y, Wang X, Chen G. The Molecular Mechanism of Potassium Absorption, Transport, and Utilization in Rice. International Journal of Molecular Sciences. 2023; 24(23):16682. https://doi.org/10.3390/ijms242316682
Chicago/Turabian StyleLian, Wenli, Anjing Geng, Yihan Wang, Minghao Liu, Yue Zhang, Xu Wang, and Guang Chen. 2023. "The Molecular Mechanism of Potassium Absorption, Transport, and Utilization in Rice" International Journal of Molecular Sciences 24, no. 23: 16682. https://doi.org/10.3390/ijms242316682
APA StyleLian, W., Geng, A., Wang, Y., Liu, M., Zhang, Y., Wang, X., & Chen, G. (2023). The Molecular Mechanism of Potassium Absorption, Transport, and Utilization in Rice. International Journal of Molecular Sciences, 24(23), 16682. https://doi.org/10.3390/ijms242316682