
Bioactive Polyketides from the Natural Complex of the Sea Urchin-Associated Fungi Penicillium sajarovii KMM 4718 and Aspergillus protuberus KMM 4747

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
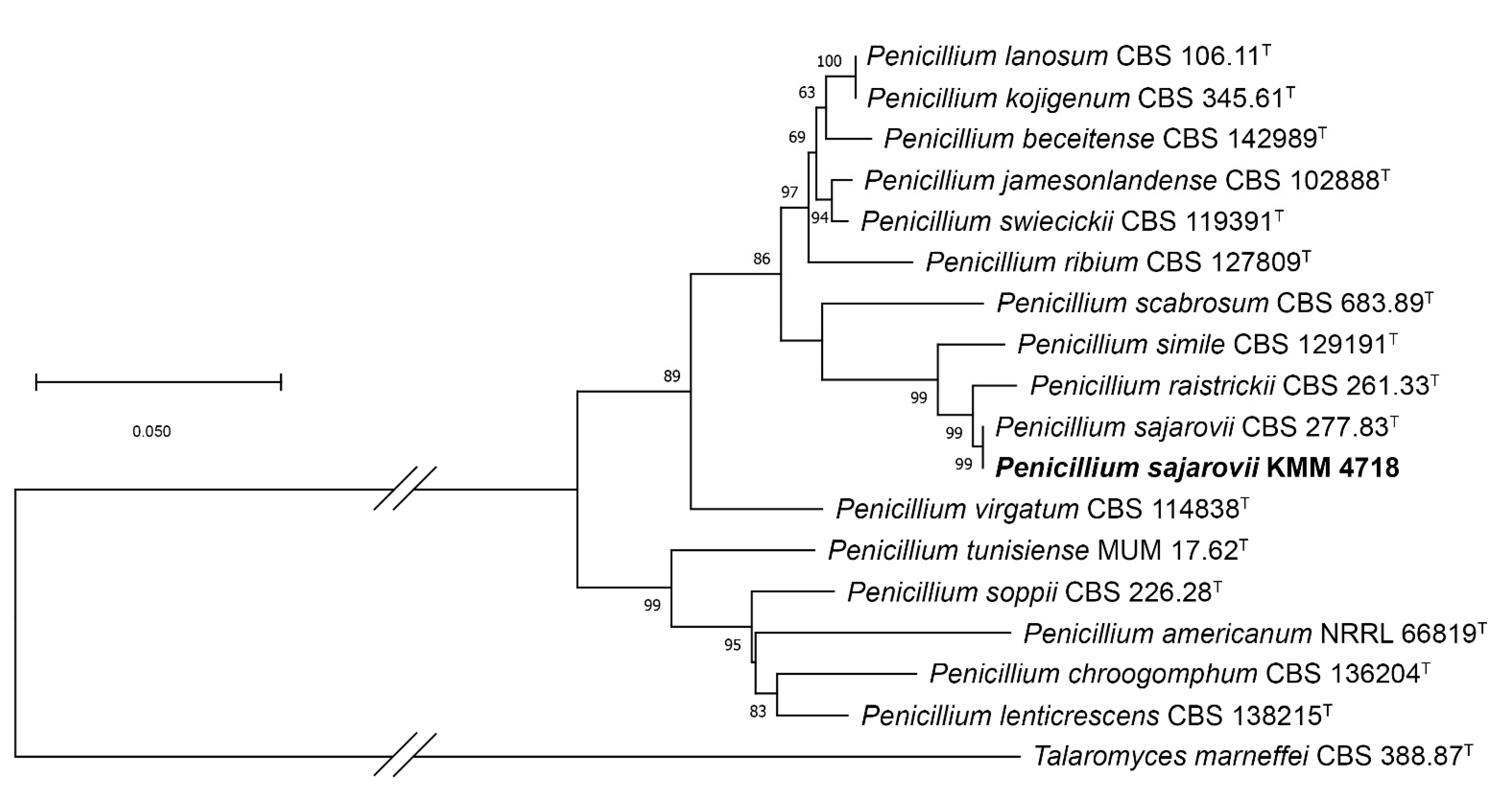
2.1. Molecular Identification of the Fungal Strains
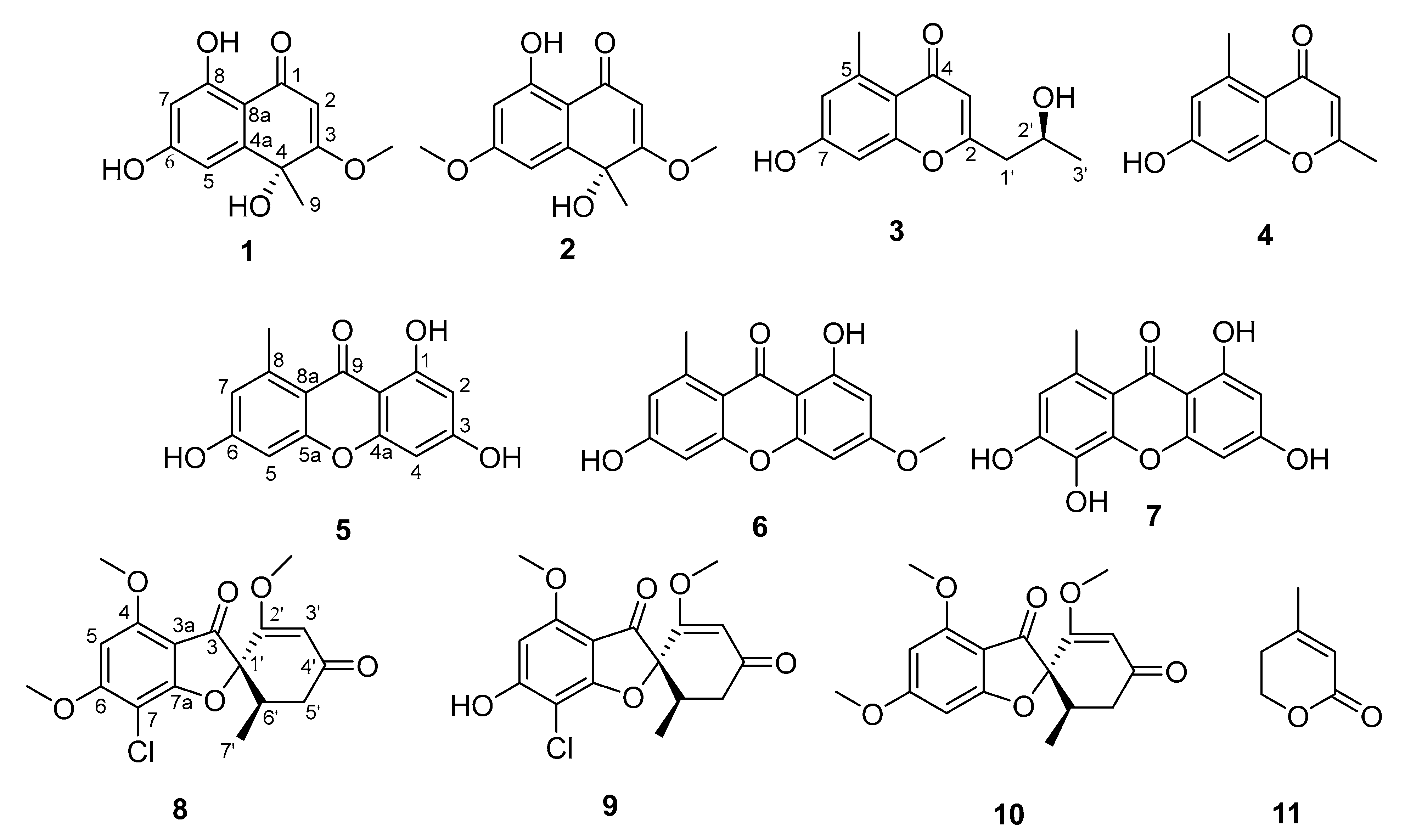
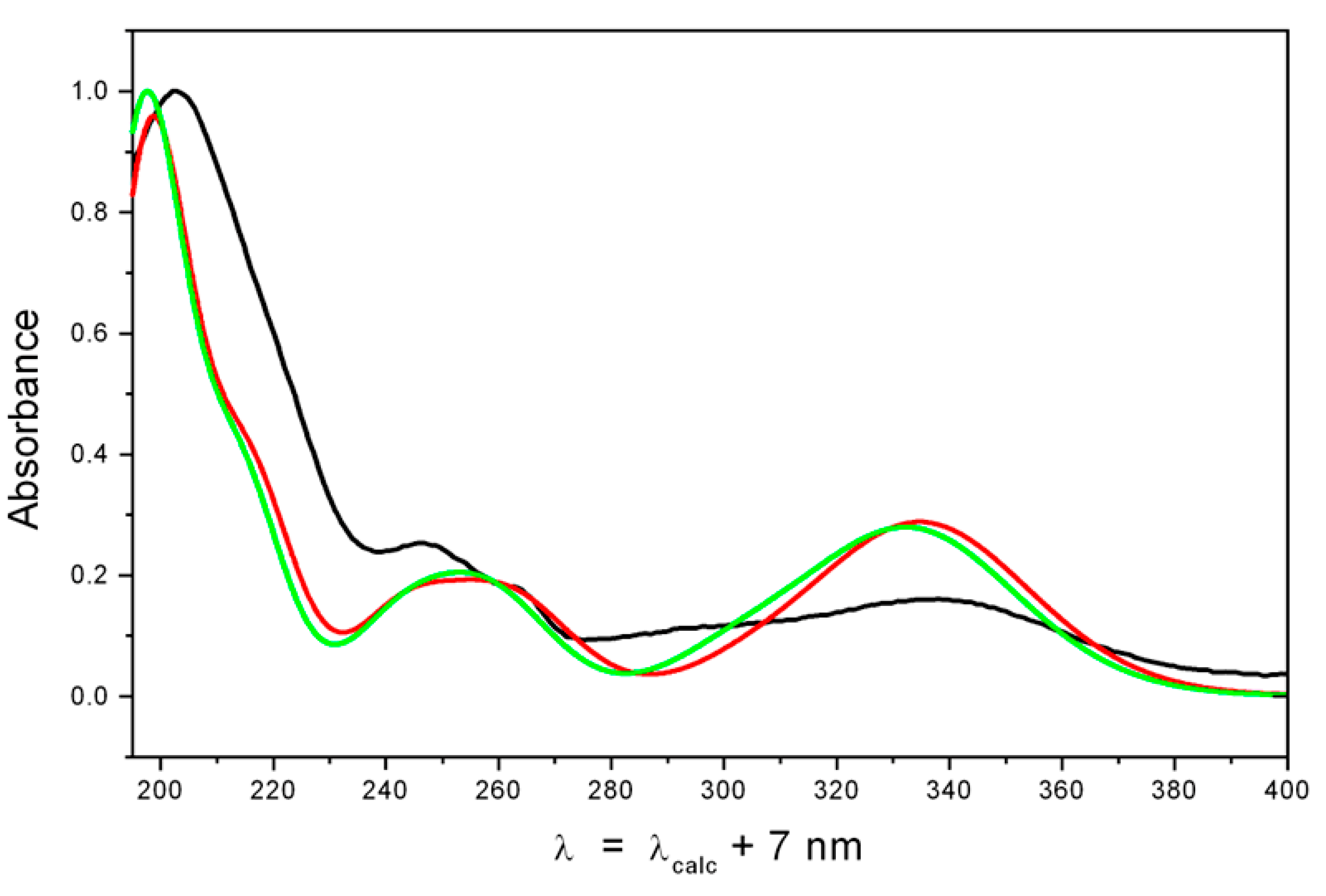
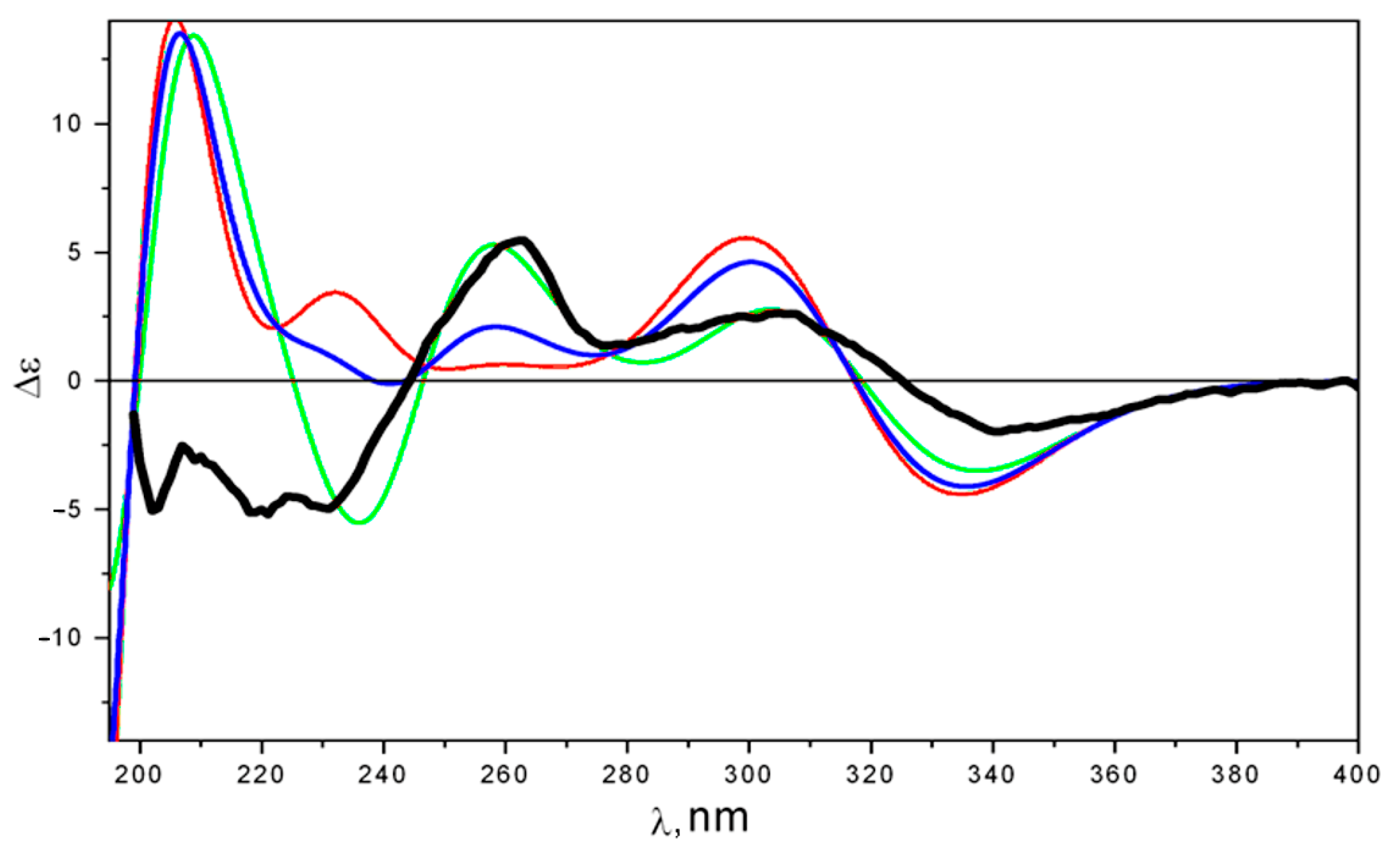
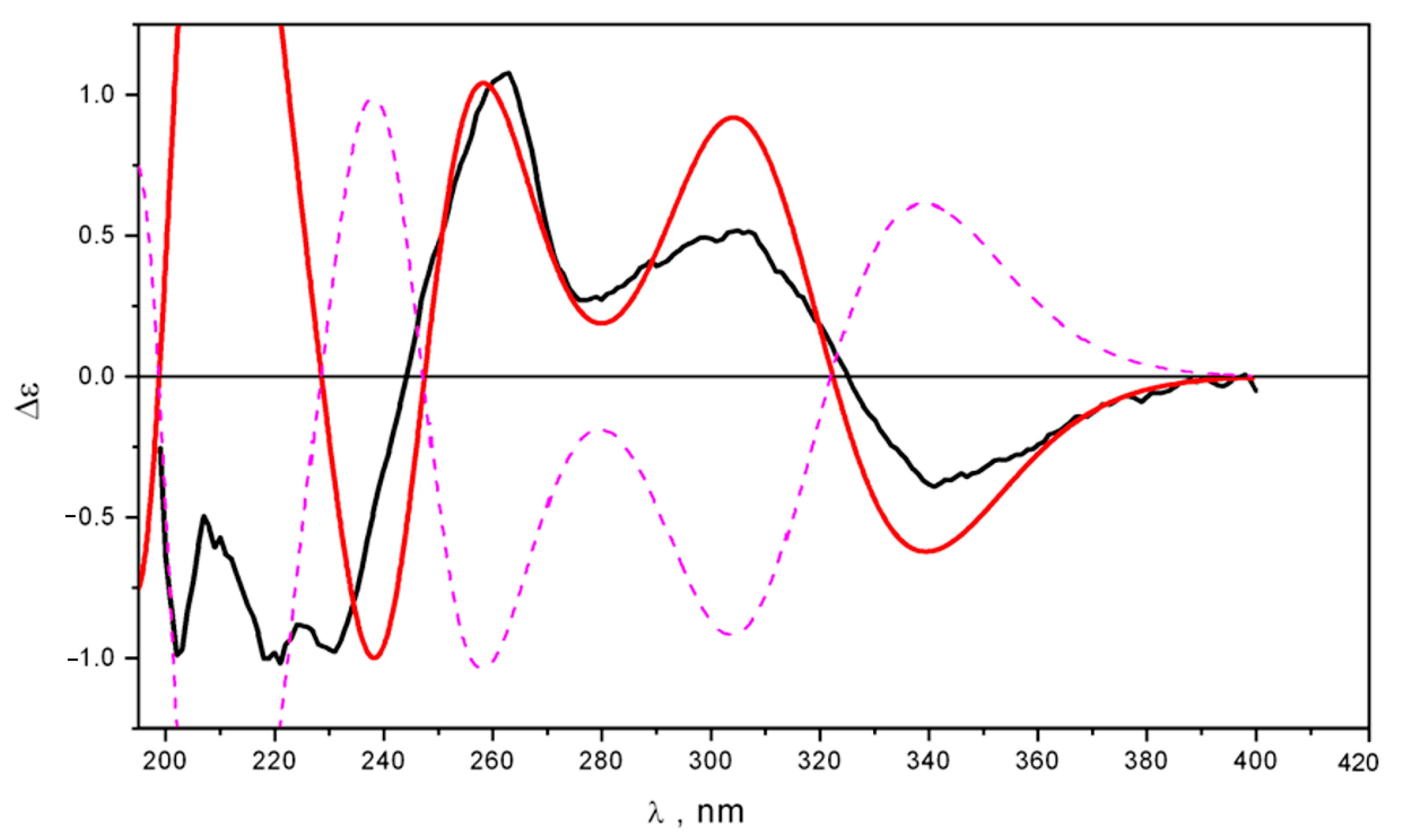
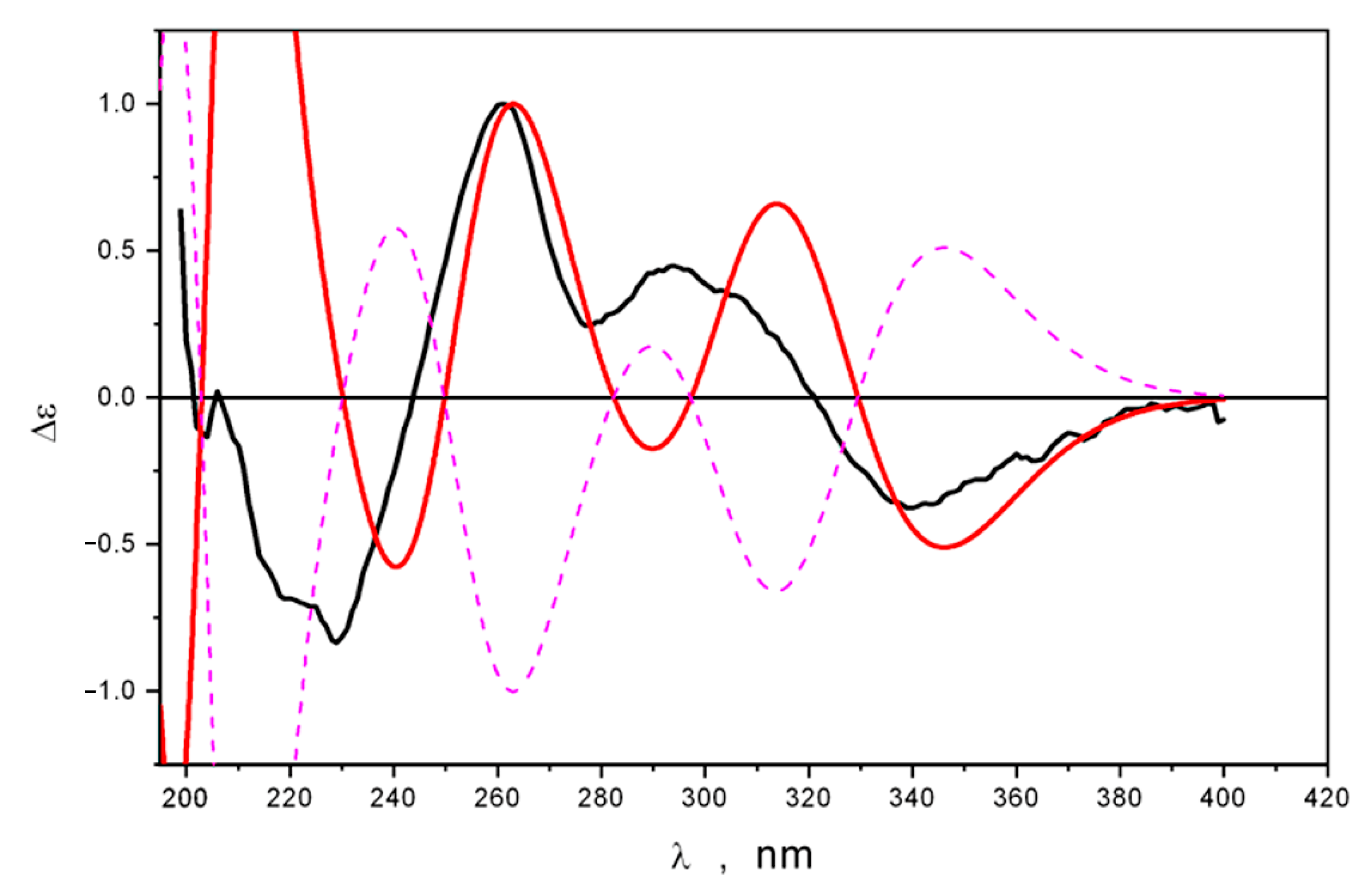
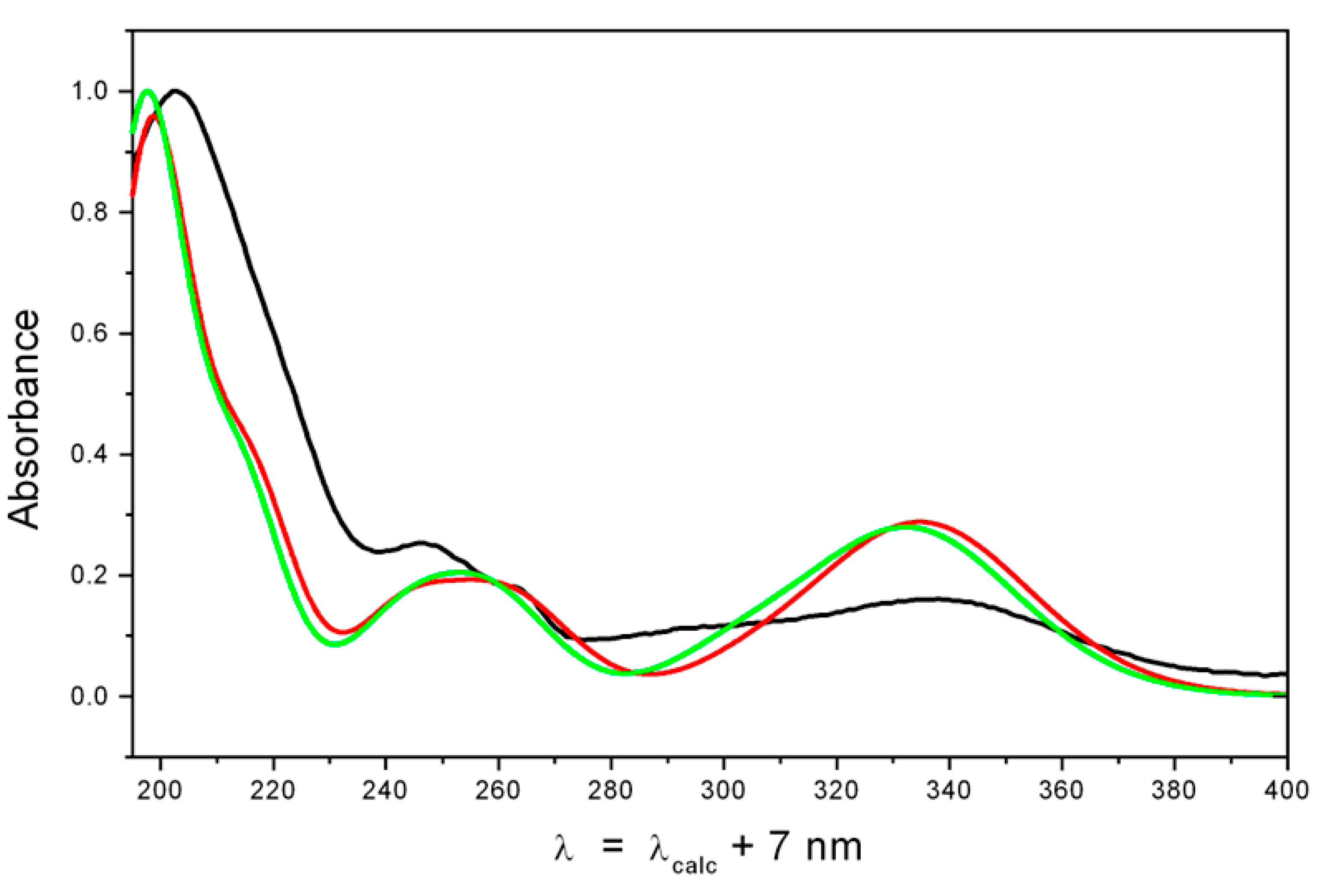
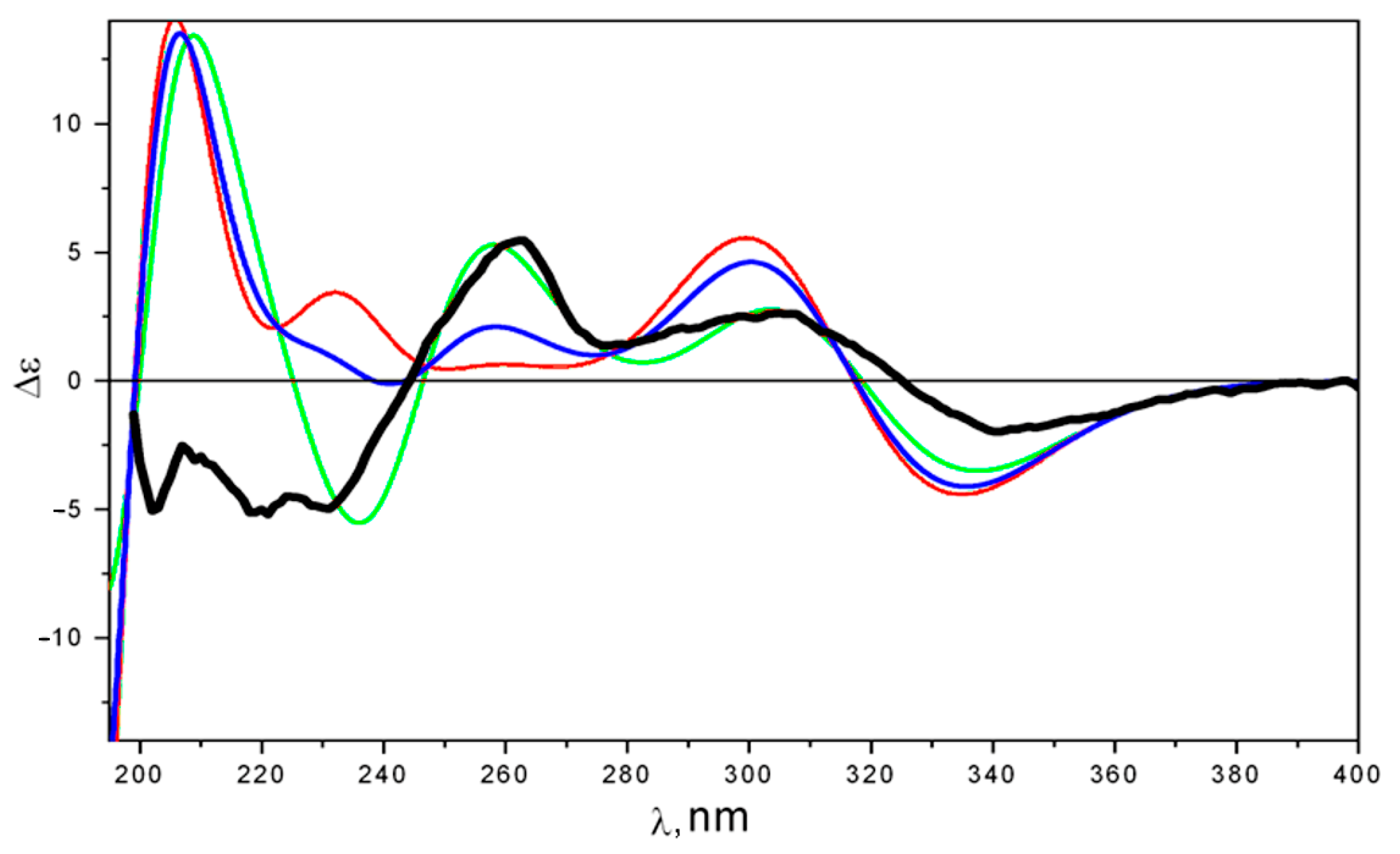
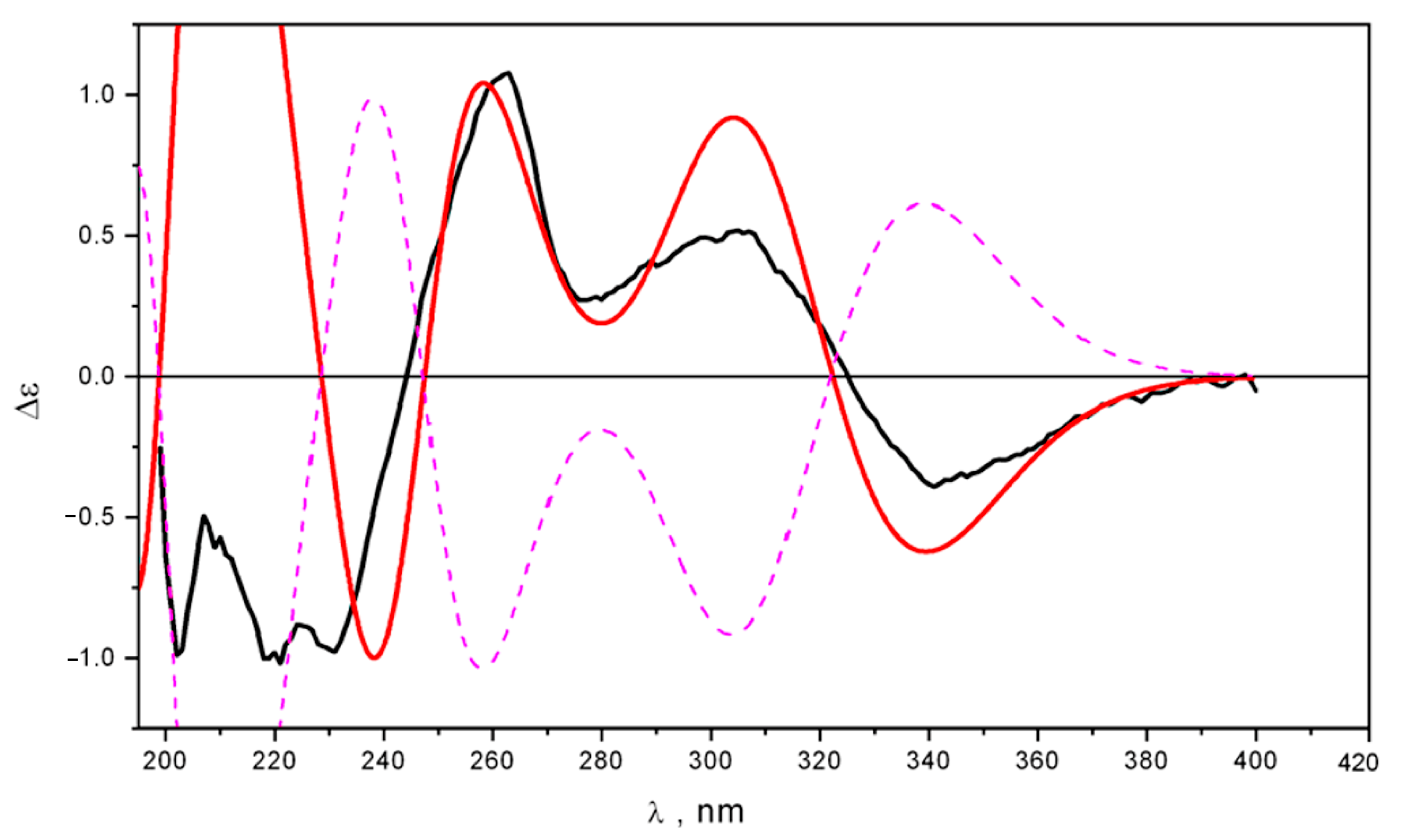
2.2. Structure Elucidation
2.3. Bioactivity of Isolated Compounds
2.3.1. Antimicrobial Activity
2.3.2. Cytotoxic Activity
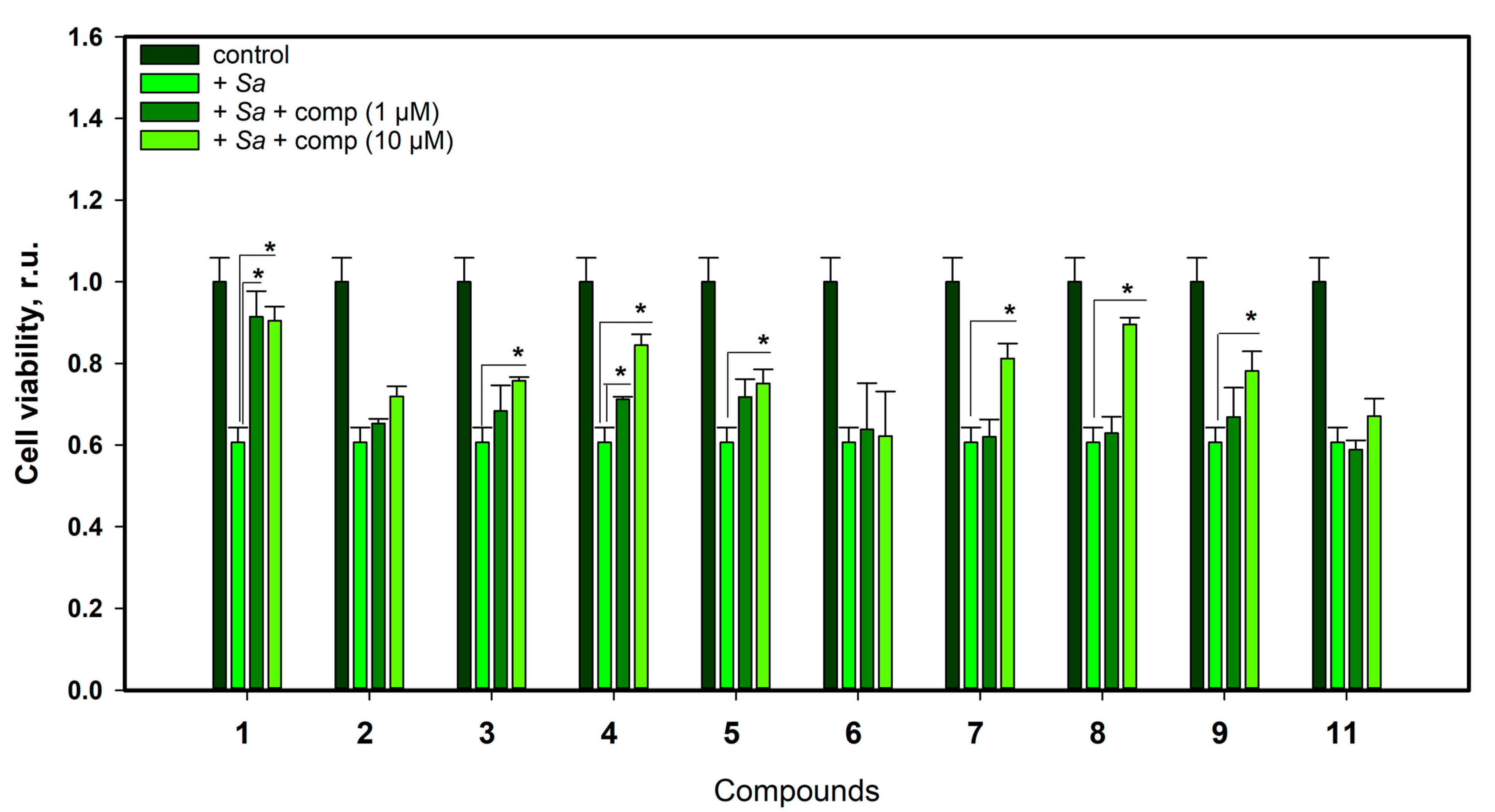
2.3.3. The Effect of Compounds against S. aureus Infection Damage of H9c2
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Fungal Strains
4.3. DNA Extraction and Amplification
4.4. Phylogenetic Analysis
4.5. Cultivation of Penicillium sajarovii KMM 4718 and Aspergillus protuberus KMM 4747
4.6. Extraction and Isolation
4.7. Spectral Data
4.8. Quantum-Chemical Modeling
4.9. Urease Inhibition Assay
4.10. Antimicrobial Activity
4.11. Cell Culture
4.12. Cell Viability Assay
4.13. Staphylococcus aureus-Induced Infection of H9c2 Cardiomyocytes
4.14. Statistical Data Evaluation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, J.; Lao, Y.-M.; Song, J.-T.; Jin, H.; Zhu, J.-M.; Cai, Z.-H. Temporal heterogeneity of microbial communities and metabolic activities during a natural algal bloom. Water Res. 2020, 183, 116020. [Google Scholar] [CrossRef]
- Liu, C.; Kakeya, H. Cryptic Chemical communication: Secondary metabolic responses revealed by microbial co-culture. Chem. Asian J. 2020, 15, 327–337. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, P.; Ye, X.; Wei, B.; Emam, M.; Zhang, H.; Wang, H. The structural diversity of marine microbial secondary metabolites based on co-culture strategy: 2009–2019. Mar. Drugs 2020, 18, 449. [Google Scholar] [CrossRef]
- Li, X.; Xu, H.; Li, Y.; Liao, S.; Liu, Y. Exploring diverse bioactive secondary metabolites from marine microorganisms using co-culture strategy. Molecules 2023, 28, 6371. [Google Scholar] [CrossRef]
- Zhuravleva, O.I.; Belousova, E.B.; Oleinikova, G.K.; Antonov, A.S.; Khudyakova, Y.V.; Rasin, A.B.; Popov, R.S.; Menchinskaya, E.S.; Trinh, P.T.H.; Yurchenko, A.N.; et al. Cytotoxic drimane-type sesquiterpenes from co-culture of the marine-derived fungi Aspergillus carneus KMM 4638 and Beauveria felina (=Isaria felina) KMM 4639. Mar. Drugs 2022, 20, 584. [Google Scholar] [CrossRef]
- Afiyatullov, S.S.; Zhuravleva, O.I.; Antonov, A.S.; Berdyshev, D.V.; Pivkin, M.V.; Denisenko, V.A.; Popov, R.S.; Gerasimenko, A.V.; von Amsberg, G.; Dyshlovoy, S.A.; et al. Prenylated indole alkaloids from co-culture of marine-derived fungi Aspergillus sulphureus and Isaria felina. J. Antibiot. 2018, 71, 846–853. [Google Scholar] [CrossRef]
- Zhuravleva, O.I.; Kirichuk, N.N.; Denisenko, V.A.; Dmitrenok, P.S.; Yurchenko, E.A.; Min′ko, E.M.; Ivanets, E.V.; Afiyatullov, S.S. New diorcinol J produced by co-cultivation of marine fungi Aspergillus sulphureus and Isaria felina. Chem. Nat. Comp. 2016, 52, 227–230. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsube, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Liu, W.Z.; Ma, L.Y.; Liu, D.S.; Huang, Y.L.; Wang, C.H.; Shi, S.S.; Pan, X.H.; Song, X.D.; Zhu, R.X. Peniciketals A-C, new spiroketals from saline soil derived Penicillium raistrichii. Org. Lett. 2014, 16, 90–93. [Google Scholar] [CrossRef]
- Belofsky, G.N.; Gloer, K.B.; Gloer, J.B.; Wicklow, D.T.; Dowd, P.F. New p-terphenyl and polyketide metabolites from the sclerotia of Penicillium raistrickii. J. Nat. Prod. 1998, 61, 1115–1119. [Google Scholar] [CrossRef]
- Li, J.; Hu, Y.; Hao, X.; Tan, J.; Li, F.; Qiao, X.; Chen, S.; Xiao, C.; Chen, M.; Peng, Z.; et al. Raistrickindole A, an anti-HCV oxazinoindole alkaloid from Penicillium raistrickii IMB17-034. J. Nat. Prod. 2019, 82, 1391–1395. [Google Scholar] [CrossRef]
- Liu, D.-S.; Rong, X.-G.; Kang, H.-H.; Ma, L.-Y.; Hamann, M.T.; Liu, W.-Z. Raistrickiones A−E from a highly productive strain of Penicillium raistrickii generated through thermo change. Mar. Drugs 2018, 16, 213. [Google Scholar] [CrossRef]
- Ma, L.-Y.; Liu, W.; Shen, L.; Huang, Y.-L.; Rong, X.-G.; Xu, Y.-Y.; Xue Dong, G. Spiroketals, isocoumarin, and indoleformic acid derivatives from saline soil derived fungus Penicillium raistrickii. Tetrahedron 2012, 68, 2276–2282. [Google Scholar] [CrossRef]
- Miao, F.-P.; Li, X.-D.; Liu, X.-H.; Cichewicz, R.H.; Ji, N.-Y. Secondary metabolites from an algicolous Aspergillus versicolor strain. Mar. Drugs 2012, 10, 131–139. [Google Scholar] [CrossRef]
- Ebada, S.; El-Neketi, M.; Ebrahim, W.; Mándi, A.; Kurtán, T.; Kalscheuer, R.; Müller, W.; Chaidir, C. Cytotoxic secondary metabolites from the endophytic fungus Aspergillus versicolor KU258497. Phytochem. Lett. 2018, 24, 88–93. [Google Scholar] [CrossRef]
- Elsbaey, M.; Tanaka, C.; Miyamoto, T. New secondary metabolites from the mangrove endophytic fungus Aspergillus versicolor. Phytochem. Lett. 2019, 32, 70–76. [Google Scholar] [CrossRef]
- Kirichuk, N.N.; Pivkin, M.V.; Matveeva, T.V. Three new Penicillium species from marine subaqueous soils. Mycol. Prog. 2017, 16, 15–26. [Google Scholar] [CrossRef]
- Jones, E.; Suetrong, S.; Sakayaroj, J.; Bahkali, A.; Abdel-Wahab, M.; Boekhout, T.; Pang, K.-L. Classification of marine Ascomycota, Basidiomycota, Blastocladiomycota and Chytridiomycota. Fungal Divers. 2015, 73, 1–72. [Google Scholar] [CrossRef]
- McGovern, E.P.; Bentley, R. Biosynthesis of flaviolin and 5,8-dihydroxy-2,7-dimethoxy-1,4-naphthoquinone. Biochemistry 1975, 14, 3138–3143. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16 Rev. A.03; Gaussian, Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- He, Z.-H.; Wu, J.; Xu, L.; Hu, M.-Y.; Xie, M.-M.; Hao, Y.-J.; Li, S.-J.; Shao, Z.-Z.; Yang, X.-W. Chemical constituents of the deep-sea-derived Penicillium solitum. Mar. Drugs 2021, 19, 580. [Google Scholar] [CrossRef]
- Kashiwada, Y.; Nonaka, G.; Nishioka, I. Studies on Rhubarb (Rhei Rhizoma). V. Isolation and characterization of chromone and chromanone berivatives. Chem. Pharm. Bull. 1984, 32, 3493–3500. [Google Scholar] [CrossRef]
- Zhan, F.; Yang, T.; Han, Y.-p.; Li, G.-y. A new chlorinated diphenyl ether and five known polyketide metabolites from Penicillium griseofulvum cib-119. Nat. Prod. Res. 2013, 27, 1393–1397. [Google Scholar] [CrossRef]
- Khamthong, N.; Rukachaisirikul, V.; Tadpetch, K.; Kaewpet, M.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Tetrahydroanthraquinone and xanthone derivatives from the marine-derived fungus Trichoderma aureoviride PSU-F95. Arch. Pharmacal Res. 2012, 35, 461–468. [Google Scholar] [CrossRef]
- Königs, P.; Rinker, B.; Maus, L.; Nieger, M.; Rheinheimer, J.; Waldvogel, S.R. Structural revision and synthesis of altechromone A. J. Nat. Prod. 2010, 73, 2064–2066. [Google Scholar] [CrossRef]
- Kawakami, H.; Suzuki, C.; Yamaguchi, H.; Hara, K.; Komine, M.; Yamamoto, Y. Norlichexanthone produced by cultured endolichenic fungus induced from Pertusaria laeviganda and its antioxidant activity. Biosci. Biotechnol. Biochem. 2019, 83, 996–999. [Google Scholar] [CrossRef]
- Wang, Q.X.; Bao, L.; Yang, X.L.; Guo, H.; Yang, R.N.; Ren, B.; Zhang, L.X.; Dai, H.Q.; Guo, L.D.; Liu, H.W. Polyketides with antimicrobial activity from the solid culture of an endolichenic fungus Ulocladium sp. Fitoterapia 2012, 83, 209–214. [Google Scholar] [CrossRef]
- Li, Y.; Tan, Y.; Liu, J.; Zhou, X.; Zong, S.; Dong, J.; Liu, Y.; Yang, B. A new griseofulvin derivative from a soft coral-derived fungus Eupenicillium sp. Nat. Prod. Res. 2020, 34, 2971–2975. [Google Scholar] [CrossRef]
- Rego, Y.F.; Queiroz, M.P.; Brito, T.O.; Carvalho, P.G.; de Queiroz, V.T.; de Fátima, Â.; Macedo Jr, F. A review on the development of urease inhibitors as antimicrobial agents against pathogenic bacteria. J. Adv. Res. 2018, 13, 69–100. [Google Scholar] [CrossRef]
- Finkelstein, E.; Amichai, B.; Grunwald, M.H. Griseofulvin and its uses. Int. J. Antimicrob. Agents 1996, 6, 189–194. [Google Scholar] [CrossRef]
- Aris, P.; Wei, Y.; Mohamadzadeh, M.; Xia, X. Griseofulvin: An updated overview of old and current knowledge. Molecules 2022, 27, 7034. [Google Scholar] [CrossRef]
- Rathinasamy, K.; Jindal, B.; Asthana, J.; Singh, P.; Balaji, P.V.; Panda, D. Griseofulvin stabilizes microtubule dynamics, activates p53 and inhibits the proliferation of MCF-7 cells synergistically with vinblastine. BMC Cancer 2010, 10, 213. [Google Scholar] [CrossRef]
- Gu, W. Bioactive metabolites from Alternaria brassicicola ML-P08, an endophytic fungus residing in Malus halliana. World J. Microbiol. Biotechnol. 2009, 25, 1677–1683. [Google Scholar] [CrossRef]
- Abbafati, C.; Machado, D.B.; Cislaghi, B.; Salman, O.M.; Karanikolos, M.; McKee, M.; Abbas, K.M.; Brady, O.J.; Larson, H.J.; Trias-Llimós, S.; et al. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef]
- Scholin, C.A.; Herzog, M.; Sogin, M.; Anderson, D.M. Identification of group- and strain-specific genetic markers for globally distributed Alexandrium (Dinophyceae). II. Sequence analysis of a fragment of the LSU rRNA gene. J. Phycol. 1994, 30, 999–1011. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Tavare, S. Some probabilistic and statistical problems in the analysis of DNA sequences. Lect. Math. Life Sci. 1986, 17, 57–86. [Google Scholar]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | 1 | 2 | ||
|---|---|---|---|---|
| δC, Type | δH, Mult. (J in Hz) | δC, Type | δH, Mult. (J in Hz) | |
| 1 | 190.3, C | 190.4, C | ||
| 2 | 99.6, CH | 5.52, s | 99.6, CH | 5.55, s |
| 3 | 179.3, C | 179.6, C | ||
| 4 | 70.7, C | 70.9, C | ||
| 4a | 151.5, C | 151.1, C | ||
| 5 | 106.1, CH | 6.77, d (2.5) | 105.2, CH | 6.81, d (2.5) |
| 6 | 164.1, C | 166.1, C | ||
| 7 | 102.4, CH | 6.25, d (2.5) | 100.6, CH | 6.35, d (2.5) |
| 8 | 164.4, C | 164.7, C | ||
| 8a | 107.5, C | 108.0, C | ||
| 9 | 32.5, CH3 | 1.60, s | 32.6, CH3 | 1.62, s |
| 3-OMe | 56.9, CH3 | 3.89, s | 57.0, CH3 | 3.90, s |
| 6-OMe | 55.9, CH3 | 3.87, s | ||
| 8-OH | 13.2, s | |||
| Compounds | Inhibition of Urease Activity IC50, µM | Inhibition of Microbial Growth 1, % | ||
|---|---|---|---|---|
| S. aureus | E. coli | C. albicans | ||
| 1 | >100.0 | 35.1 ± 0.9 | 25.4 ± 1.0 | - |
| 2 | 98.5 ± 0.8 | - | - | - |
| 3 | >100.0 | 17.7 ± 0.3 | - | - |
| 4 | >100.0 | 30.1 ± 1.2 | 13.6 ± 0.3 | 11.1 ± 1.3 |
| 5 | 15.3 ± 2.1 | 20.8 ± 6.5 | 27.9 ± 3.3 | - |
| 6 | 97.3 ± 1.6 | - | - | - |
| 7 | 100.0 | - | - | 15.4 ± 2.1 |
| 8 | 10.1 ± 0.6 | 11.5 ± 4.3 | 16.8 ± 4.5 | - |
| 9 | >100.0 | 23.0 ± 0.3 | 23.4 ± 1.8 | - |
| 11 | 11.4 ± 1.0 | - | - | - |
| Compound | HepG2 | H9c2 | ||||
|---|---|---|---|---|---|---|
| Cell Viability, % | IC50 | Cell Viability, % | IC50 | |||
| 100 µM | 10 µM | 100 µM | 10 µM | |||
| 1 | 106.8 ± 3.9 | 107.5 ± 3.2 | >100 | 65.4 ± 4.8 | 83.6 ± 4.4 | >100 |
| 2 | 76.9 ± 2.1 | 87.3 ± 4.1 | >100 | 79.0 ± 3.4 | 96.0 ± 1.9 | >100 |
| 3 | 101.4 ± 3.9 | 91.7 ± 1.5 | >100 | 74.3 ± 3.2 | 96.4 ± 2.1 | >100 |
| 4 | 92.6 ± 4.1 | 99.2 ± 6.5 | >100 | 96.5 ± 3.6 | 110.3 ± 2.9 | >100 |
| 5 | 52.6 ± 3.0 | 88.0 ± 4.9 | >100 | 49.0 ± 1.6 | 86.3 ± 6.3 | 97.4 ± 2.2 |
| 6 | 27.1 ± 2.4 | 85.7 ± 5.7 | 64.7 ± 0.8 | 63.3 ± 1.4 | 86.0 ± 4.8 | >100 |
| 7 | 57.4 ± 4.2 | 77.1 ± 1.4 | >100 | 52.9 ± 1.5 | 98.0 ± 0.9 | >100 |
| 8 | 39.9 ± 0.7 | 81.0 ± 2.1 | 77.8 ± 1.3 | 71.6 ± 3.7 | 89.8 ± 2.7 | >100 |
| 9 | 84.2 ± 3.2 | 91.5 ± 6.8 | >100 | 92.9 ± 3.7 | 97.4 ± 8.1 | >100 |
| 11 | 76.6 ± 2.4 | 87.5 ± 5.3 | >100 | 89.0 ± 2.9 | 95.9 ± 0.6 | >100 |
| Species | Strain Number | GenBank Accession Number | |||
|---|---|---|---|---|---|
| ITS | BenA | CaM | RPB2 | ||
| Penicillium simile Davolos et al. Davolos et al., 2012 | CBS 129191 T | FJ376592 | FJ376595 | GQ979710 | – |
| Penicillium sajarovii Quintan | CBS 277.83 T | KC411724 | MN969397 | MN969295 | – |
| Penicillium sajarovii | KMM 4718 | OR431843 | OQ466617 | OR451997 | – |
| Penicillium raistrickii G. Sm | CBS 261.33 T | AY373927 | KJ834485 | KJ867006 | – |
| Penicillium lanosum Westling | CBS 106.11 T | DQ304540 | DQ285627 | FJ530974 | – |
| Penicillium beceitense Guevara-Suarez et al. | CBS 142989 T | LT899780 | LT898229 | LT899764 | – |
| Penicillium jamesonlandense Frisvad & Overy | CBS 102888 T | DQ267912 | DQ309448 | KJ866985 | – |
| Penicillium swiecickii Zaleski | CBS 119391 T | AF033490 | KJ834494 | KJ866993 | – |
| Penicillium ribium Frisvad & Overy | CBS 127809 T | DQ267916 | MN969395 | KJ866995 | – |
| Penicillium scabrosum Frisvad et al. | CBS 683.89 T | DQ267906 | DQ285610 | FJ530987 | – |
| Penicillium chroogomphum Xu et al. | CBS 136204 T | KC594043 | KP684056 | KP684057 | – |
| Penicillium americanum Jurjevic et al. | NRRL 66819 T | MK791278 | MK803427 | MK803428 | – |
| Penicillium lenticrescens Visagie et al. | CBS 138215 T | KJ775675 | KJ775168 | KJ775404 | – |
| Penicillium soppii Zaleski | CBS 226.28 T | AF033488 | MN969399 | KJ867002 | – |
| Penicillium tunisiense Ouhibi et al. | MUM 17.62 T | MG586956 | MG586970 | MG586974 | – |
| Penicillium virgatum Nirenberg & Kwasna | CBS 114838 T | AJ748692 | KJ834500 | KJ866992 | – |
| Penicillium kojigenum Sm | CBS 345.61 T | AF033489 | KJ834463 | KJ867011 | – |
| Aspergillus versicolor (Vuill.) Tirab | CBS 583.65 T | EF652442 | EF652266 | EF652354 | EF652178 |
| Aspergillus fructus Jurjevic et al. | NRRL 239 T | EF652449 | EF652273 | EF652361 | EF652185 |
| Aspergillus tabacinus Nakaz et al. | CBS 122718 T | EF652478 | EF652302 | EF652390 | EF652214 |
| Aspergillus amoenus Roberg | NRRL 4838 T | EF652480 | JN853946 | JN854035 | JN853824 |
| Aspergillus griseoaurantiacus Visagie et al. | CBS 138191 T | KJ775553 | KJ775086 | KJ775357 | KU866988 |
| Aspergillus austroafricanus Jurjevic et al. | CBS 145748 T | JQ301891 | JN853963 | JN854025 | JN853814 |
| Aspergillus hongkongensis Tsang et al. | CBS 145671 T | AB987907 | LC000552 | MN969320 | LC000578 |
| Aspergillus protuberus Munt.-Cvetk | CBS 602.74 T | EF652460 | EF652284 | EF652372 | EF652196 |
| Aspergillus protuberus | KMM 4747 | OR431842 | OQ466615 | OR451996 | OR451998 |
| Aspergillus tennesseensis Jurjevic et al. | CBS 145752 T | JQ301895 | JN853976 | JN854017 | JN853806 |
| Aspergillus cvjetkovicii Jurjevic et al. | NRRL 227 T | EF652440 | EF652264 | EF652352 | EF652176 |
| Aspergillus jensenii Jurjevic et al. | NRRL 58600 T | JQ301892 | JN854007 | JN854046 | JN853835 |
| Aspergillus puulaauensis Jurjevic et al. | CBS 145750 T | JQ301893 | JN853979 | JN854034 | JN853823 |
| Aspergillus venenatus Jurjevic et al. | CBS 145753 T | JQ301896 | JN854003 | JN854014 | JN853803 |
| Aspergillus creber Jurjevic et al. | CBS 145749 T | JQ301889 | JN853980 | JN854043 | JN853832 |
| Aspergillus sydowii (Bainier & Sartory) Thom & Church | CBS 593.65 T | EF652450 | EF652274 | EF652362 | EF652186 |
| Aspergillus subversicolor Jurjevic et al. | CBS 145751 T | JQ301894 | JN853970 | JN854010 | JN853799 |
| Talaromyces marneffei (Segretain) Samson et al. | CBS 388.87 T | JN899344 | JX091389 | KF741958 | KM023283 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leshchenko, E.V.; Berdyshev, D.V.; Yurchenko, E.A.; Antonov, A.S.; Borkunov, G.V.; Kirichuk, N.N.; Chausova, V.E.; Kalinovskiy, A.I.; Popov, R.S.; Khudyakova, Y.V.; et al. Bioactive Polyketides from the Natural Complex of the Sea Urchin-Associated Fungi Penicillium sajarovii KMM 4718 and Aspergillus protuberus KMM 4747. Int. J. Mol. Sci. 2023, 24, 16568. https://doi.org/10.3390/ijms242316568
Leshchenko EV, Berdyshev DV, Yurchenko EA, Antonov AS, Borkunov GV, Kirichuk NN, Chausova VE, Kalinovskiy AI, Popov RS, Khudyakova YV, et al. Bioactive Polyketides from the Natural Complex of the Sea Urchin-Associated Fungi Penicillium sajarovii KMM 4718 and Aspergillus protuberus KMM 4747. International Journal of Molecular Sciences. 2023; 24(23):16568. https://doi.org/10.3390/ijms242316568
Chicago/Turabian StyleLeshchenko, Elena V., Dmitrii V. Berdyshev, Ekaterina A. Yurchenko, Alexandr S. Antonov, Gleb V. Borkunov, Natalya N. Kirichuk, Viktoria E. Chausova, Anatoly I. Kalinovskiy, Roman S. Popov, Yuliya V. Khudyakova, and et al. 2023. "Bioactive Polyketides from the Natural Complex of the Sea Urchin-Associated Fungi Penicillium sajarovii KMM 4718 and Aspergillus protuberus KMM 4747" International Journal of Molecular Sciences 24, no. 23: 16568. https://doi.org/10.3390/ijms242316568
APA StyleLeshchenko, E. V., Berdyshev, D. V., Yurchenko, E. A., Antonov, A. S., Borkunov, G. V., Kirichuk, N. N., Chausova, V. E., Kalinovskiy, A. I., Popov, R. S., Khudyakova, Y. V., Chingizova, E. A., Chingizov, A. R., Isaeva, M. P., & Yurchenko, A. N. (2023). Bioactive Polyketides from the Natural Complex of the Sea Urchin-Associated Fungi Penicillium sajarovii KMM 4718 and Aspergillus protuberus KMM 4747. International Journal of Molecular Sciences, 24(23), 16568. https://doi.org/10.3390/ijms242316568