
Multi-Omics Analysis Reveals Synergistic Enhancement of Nitrogen Assimilation Efficiency via Coordinated Regulation of Nitrogen and Carbon Metabolism by Co-Application of Brassinolide and Pyraclostrobin in Arabidopsis thaliana


Abstract
:1. Introduction
2. Results
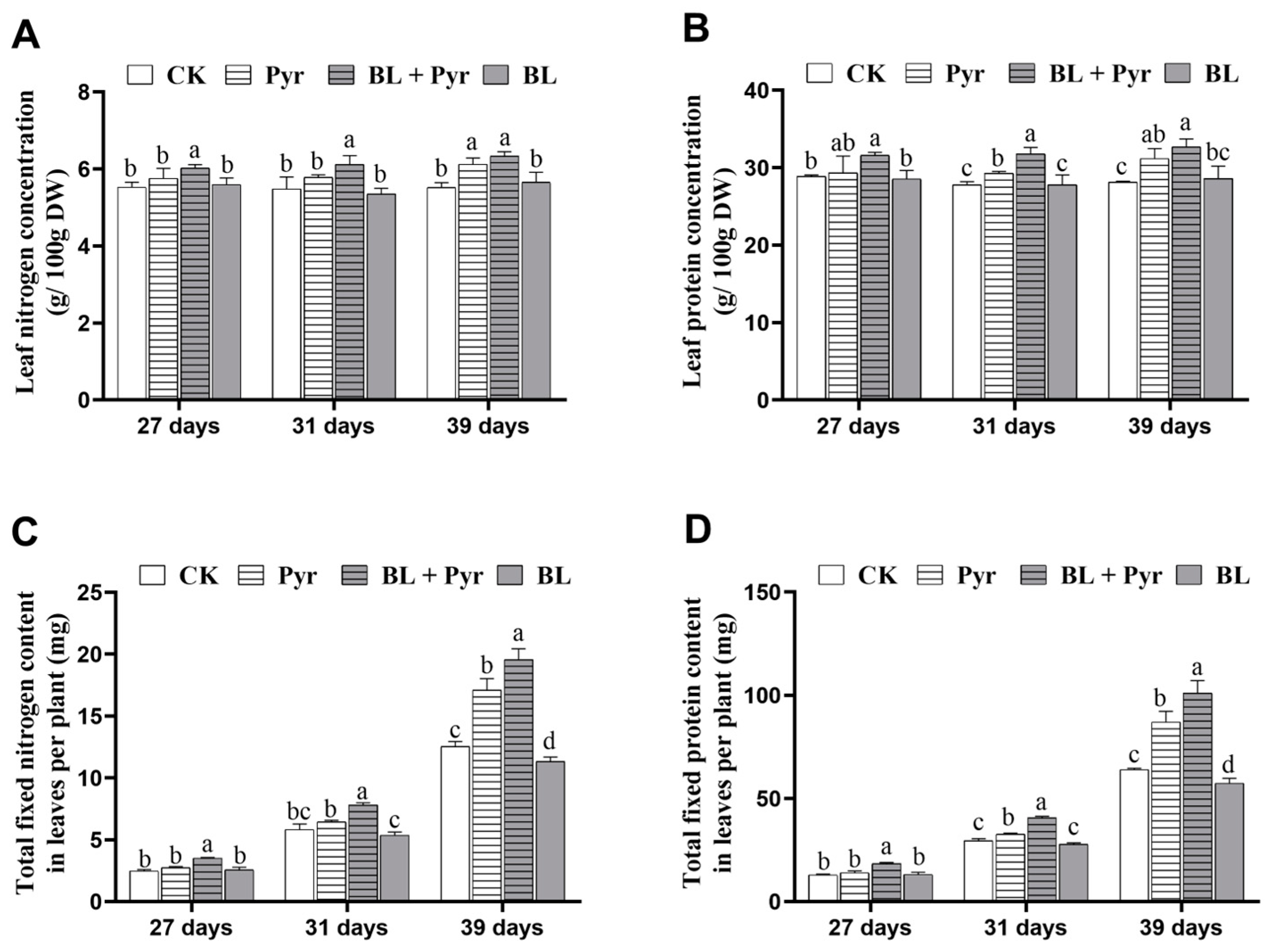
2.1. The Co-Treatment of BL and Pyr Improved N Content and Protein Content
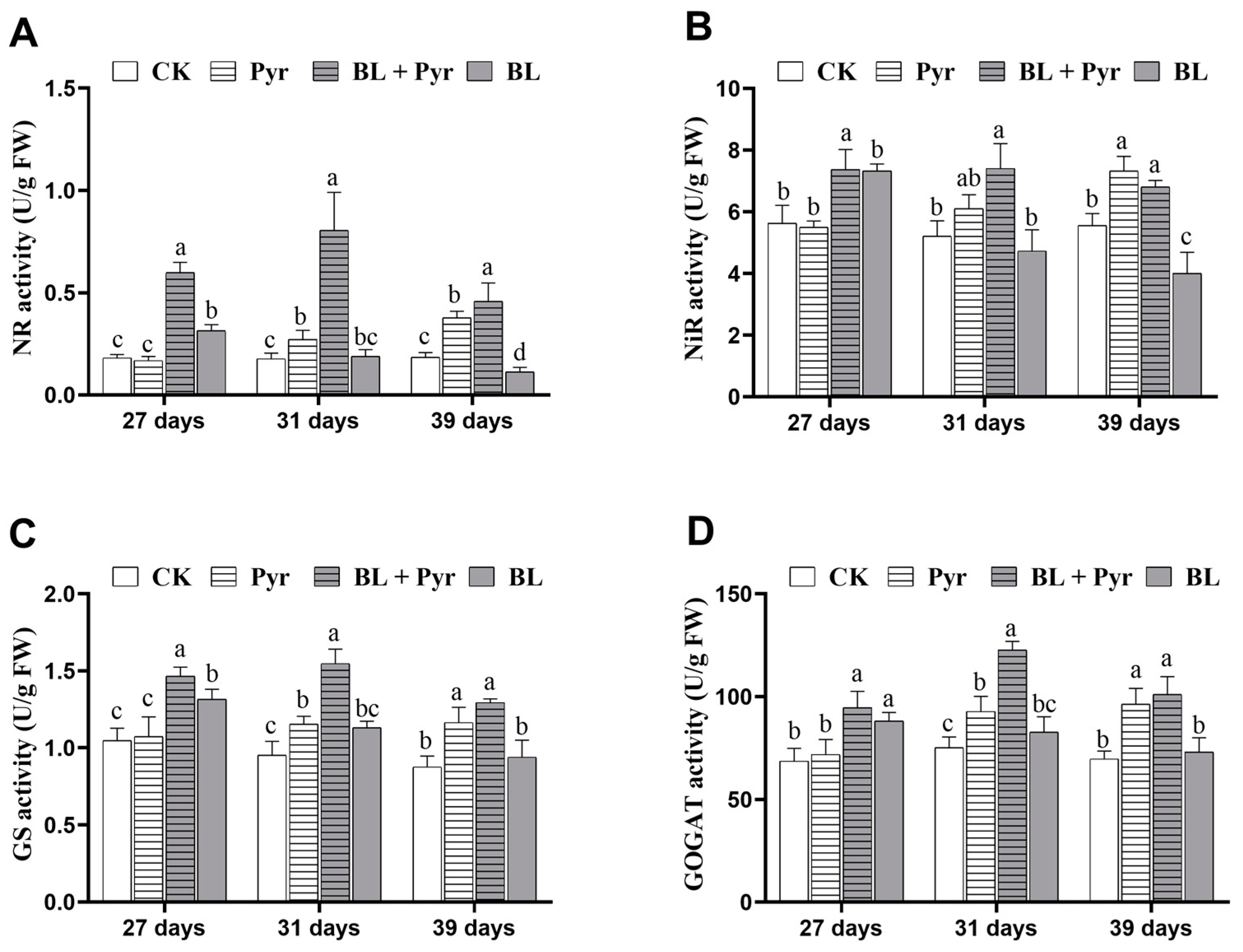
2.2. The Enzyme Activity of NR, NiR, GS, and GOGAT Were All Enhanced by the Co-Application of BL and Pyr
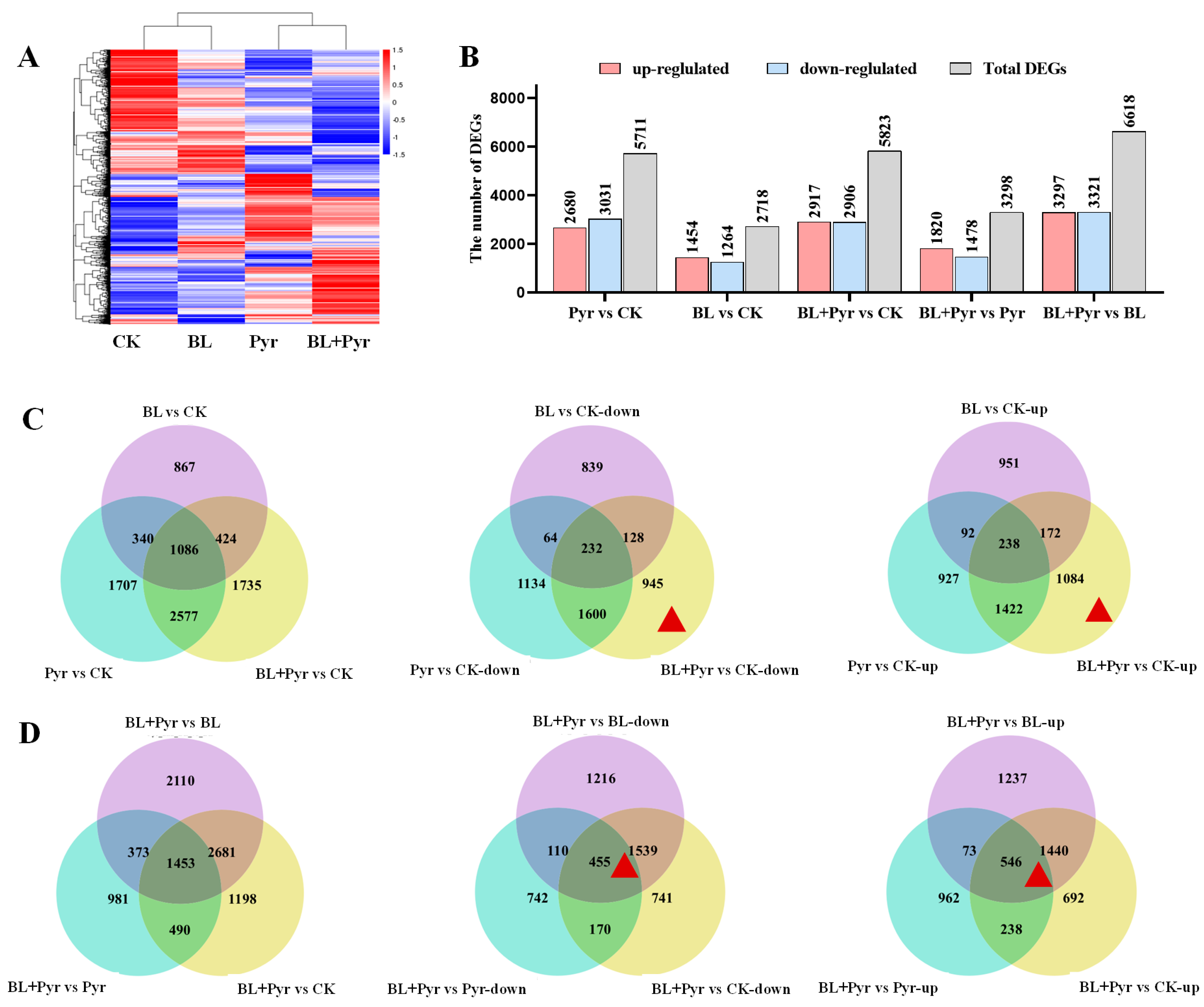
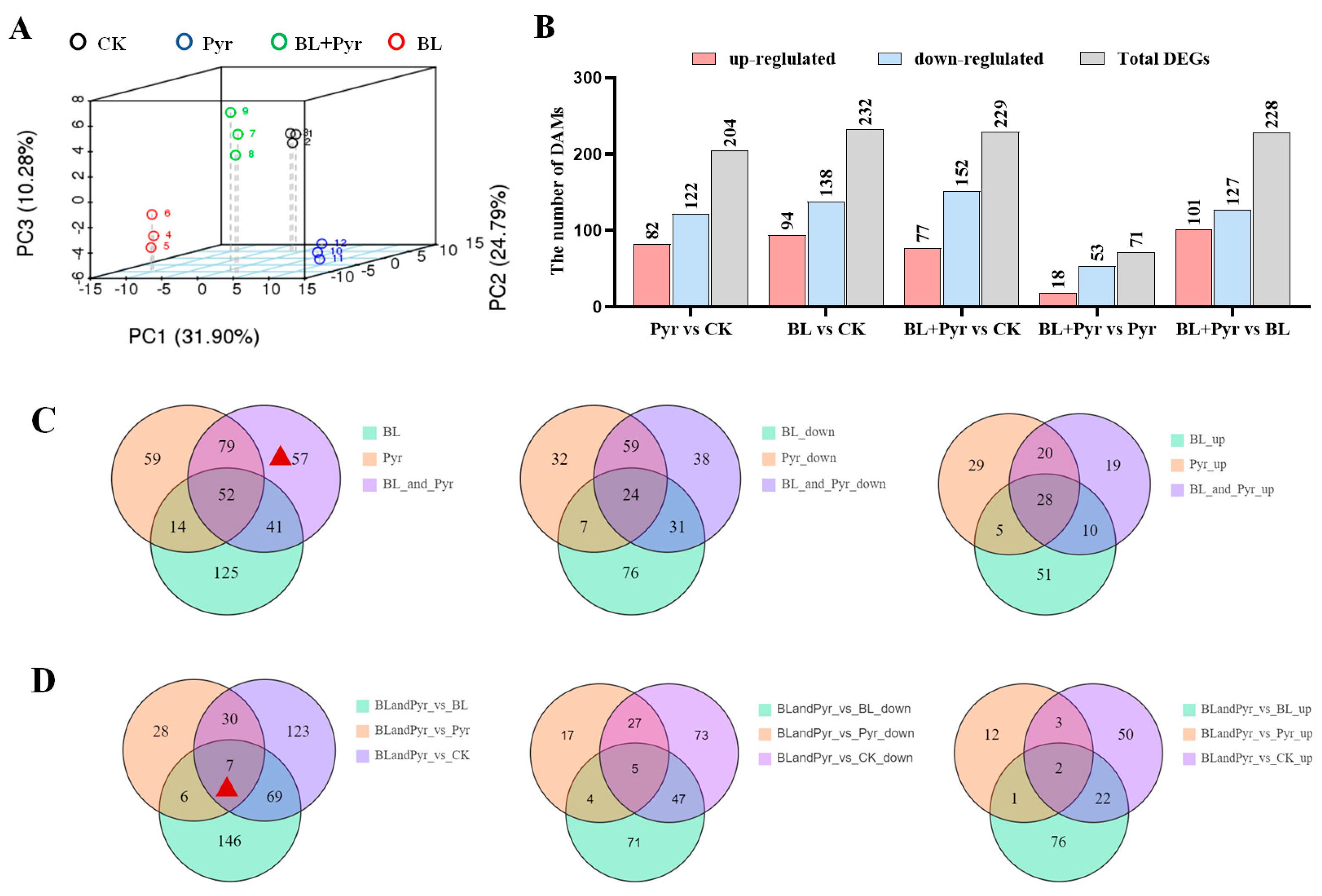
2.3. Genes Related to Protein Synthesis, Photosynthesis, and Carbohydrate Metabolism Were Uniquely Regulated by the Co-Application of BL and Pyr
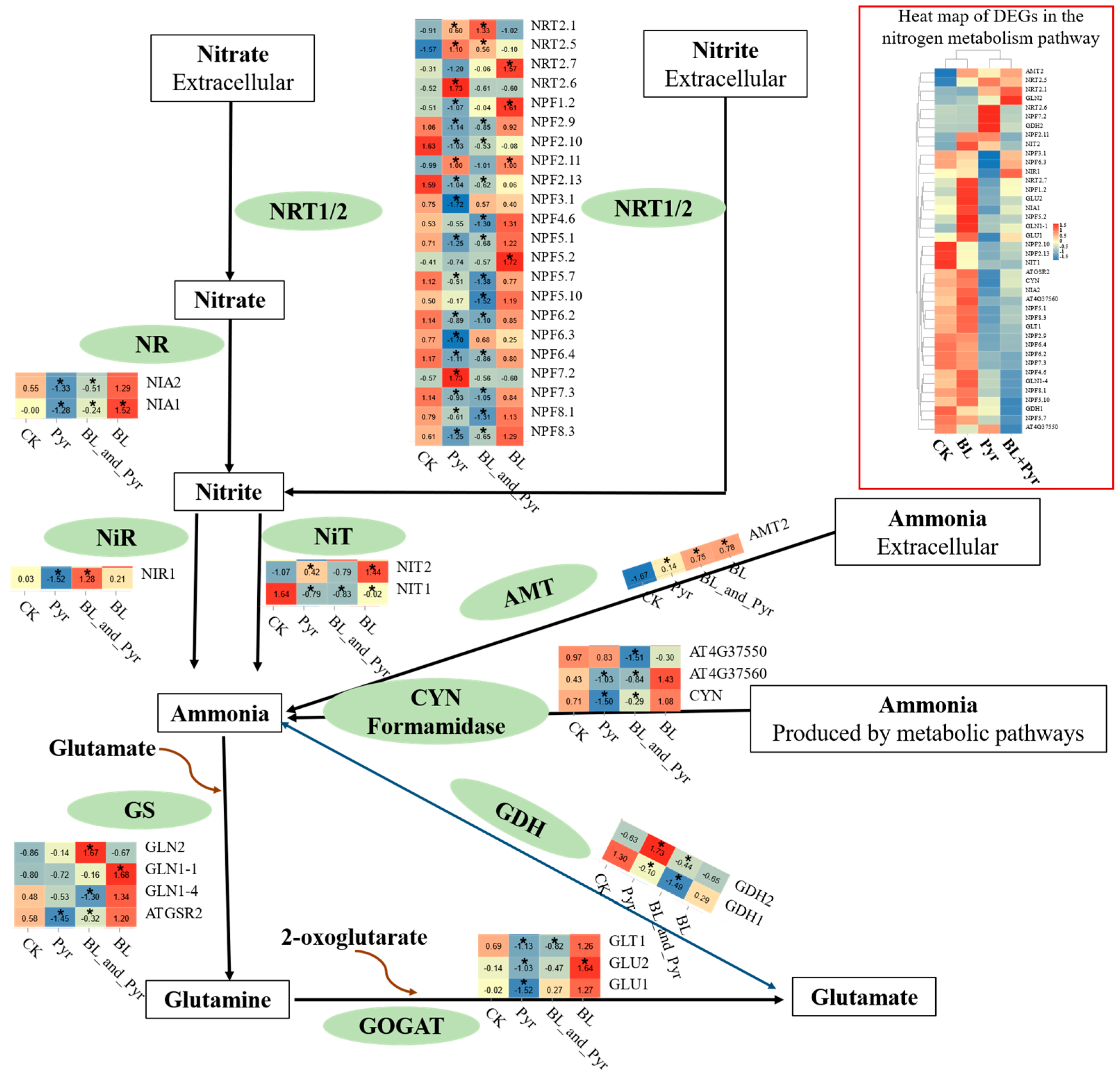
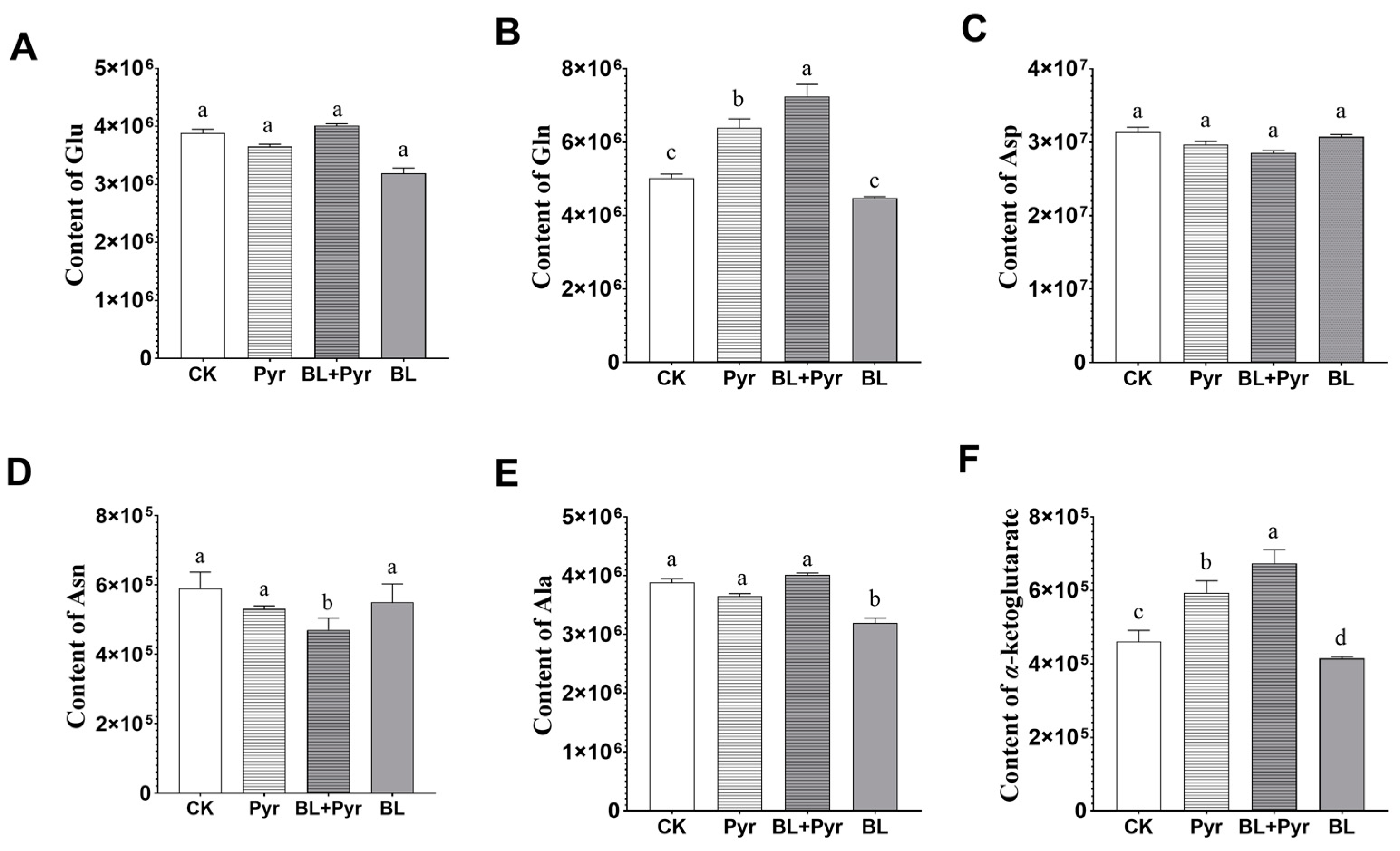
2.4. The Metabolites Related to N Metabolism and Carbon Metabolism Were Coordinately Regulated by the Co-Application of BL and Pyr
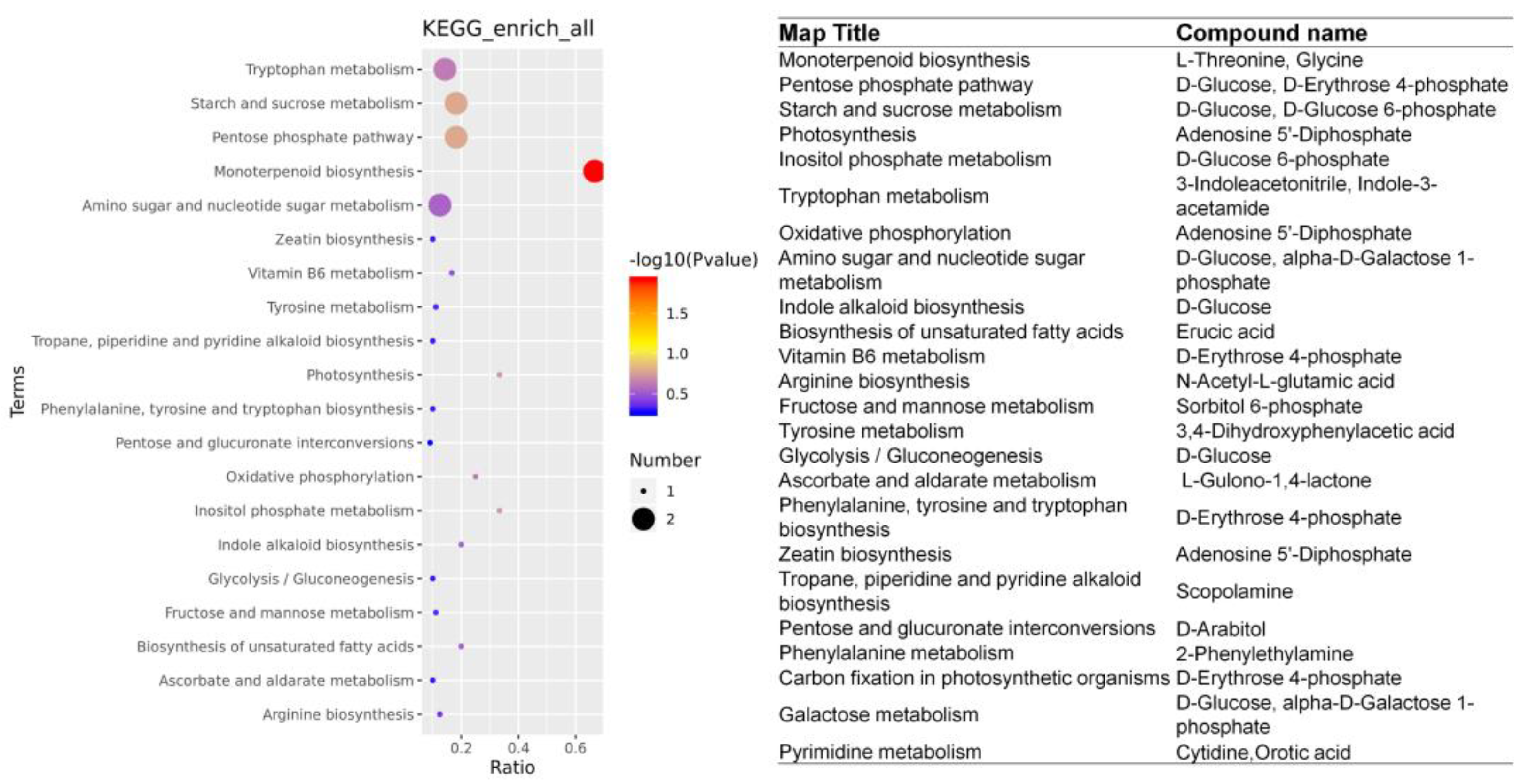
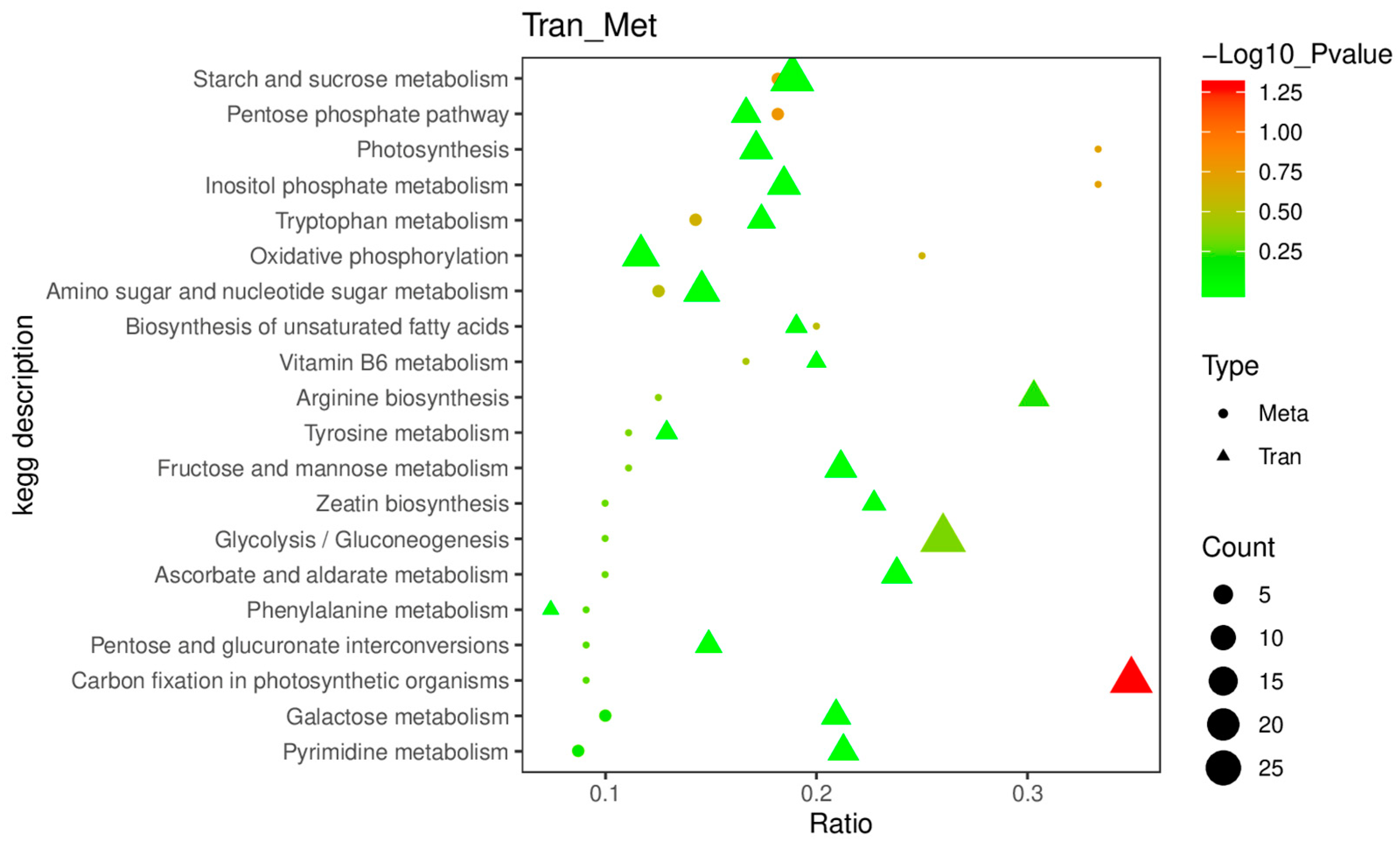
2.5. The Accumulation of DAMs Was Highly Correlated with the Regulation of DEGs
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Measurement of Leaf N Content and Protein Content
4.3. Measurement of N Assimilating Enzyme Activity
4.3.1. Measurement of Nitrate Reductase (NR) Enzyme Activity
4.3.2. Measurement of Nitrite Reductase (NiR) Enzyme Activity
4.3.3. Measurement of Glutamine Synthetase (GS) Enzyme Activity
4.3.4. Measurement of Glutamate Synthase (GOGAT) Enzyme Activity
4.4. Confirmation of RNA-Seq Data by qRT-PCR
4.5. RNA-Seq Analysis
4.6. Metabolite Profiling
4.7. Integrated Transcriptome and Metabolome Analyses
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qi, J.; Yu, L.; Ding, J.; Ji, C.; Wang, S.; Wang, C.; Ding, G.; Shi, L.; Xu, F.; Cai, H. Transcription factor OsSNAC1 positively regulates nitrate transporter gene expression in rice. Plant Physiol. 2023, 192, 2923–2942. [Google Scholar] [PubMed]
- Vivia, S.; Snyder, R.; Tegeder, M. Targeting nitrogen metabolism and transport processes to improve plant nitrogen use efficiency. Front. Plant Sci. 2021, 11, 628366. [Google Scholar]
- Liu, Q.; Wu, K.; Song, W.; Zhong, N.; Wu, Y.; Fu, X. Improving crop nitrogen use efficiency toward sustainable green revolution. Annu. Rev. Plant Biol. 2022, 73, 523–551. [Google Scholar] [CrossRef] [PubMed]
- Krapp, A. Plant nitrogen assimilation and its regulation: A complex puzzle with missing pieces. Curr. Opin. Plant Biol. 2015, 25, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Ali, A. Nitrate assimilation pathway in higher plants: Critical role in nitrogen signalling and utilization. Plant Sci. Today 2020, 7, 182–192. [Google Scholar] [CrossRef]
- Rosado-Souza, L.; Yokoyama, R.; Sonnewald, U.; Fernie, A.R. Understanding source-sink interactions: Progress in model plants and translational research to crops. Mol. Plant 2023, 16, 96–121. [Google Scholar] [CrossRef]
- Waqas, M.; Hawkesford, M.J.; Geilfus, C.-M. Feeding the world sustainably: Efficient nitrogen use. Trends Plant Sci. 2023, 28, 505–508. [Google Scholar] [CrossRef]
- Zhang, Z.; Gao, S.; Chu, C. Improvement of nutrient use efficiency in rice: Current toolbox and future perspectives. Theor. Appl. Genet. 2020, 133, 1365–1384. [Google Scholar] [CrossRef]
- Kasemsap, P.; Bloom, A.J. Breeding for higher yields of wheat and rice through modifying nitrogen metabolism. Plants 2022, 12, 85. [Google Scholar] [CrossRef]
- Lebedev, V.G.; Popova, A.A.; Shestibratov, K.A. Genetic engineering and genome editing for improving nitrogen use efficiency in plants. Cell 2021, 10, 3303. [Google Scholar] [CrossRef]
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: Current status and future prospects. J. Genet. Genom. 2022, 49, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Wang, Y.; Chen, G.; Zhang, A.; Yang, S.; Shang, L.; Wang, D.; Ruan, B.; Liu, C.; Jiang, H.; et al. The indica nitrate reductase gene OsNR2 allele enhances rice yield potential and nitrogen use efficiency. Nat. Commun. 2019, 10, 5207. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Xuan, W.; Tian, Y.; Fan, L.; Sun, J.; Tang, W.; Chen, G.; Wang, B.; Liu, Y.; Wu, W.; et al. Enhanced OsNLP4-OsNiR cascade confers nitrogen use efficiency by promoting tiller number in rice. Plant Biotechnol. J. 2021, 19, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Marmagne, A.; Park, J.; Fabien, C.; Yim, Y.; Kim, S.-J.; Kim, T.-H.; Lim, P.O.; Masclaux-Daubresse, C.; Nam, H.G. Concurrent activation of OsAMT1;2 and OsGOGAT1 in rice leads to enhanced nitrogen use efficiency under nitrogen limitation. Plant J. 2020, 103, 7–20. [Google Scholar] [CrossRef]
- Yang, J.; Wang, M.; Li, W.; He, X.; Teng, W.; Ma, W.; Zhao, X.; Hu, M.; Li, H.; Zhang, Y.; et al. Reducing expression of a nitrate-responsive bZIP transcription factor increases grain yield and N use in wheat. Plant Biotechnol. J. 2019, 17, 1823–1833. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Zhao, X.; Liu, Q.; Hong, X.; Zhang, W.; Zhang, Y.; Sun, L.; Li, H.; Tong, Y. Transgenic expression of plastidic glutamine synthetase increases nitrogen uptake and yield in wheat. Plant Biotechnol. J. 2018, 16, 1858–1867. [Google Scholar] [CrossRef]
- Martin, A.; Lee, J.; Kichey, T.; Gerentes, D.; Zivy, M.; Tatout, C.; Dubois, F.D.R.; Balliau, T.; Valot, B.T.; Davanture, M.; et al. Two cytosolic glutamine synthetase isoforms of maize are specifically involved in the control of grain production. Plant Cell 2006, 18, 3252–3274. [Google Scholar] [CrossRef]
- Cai, H.; Zhou, Y.; Xiao, J.; Li, X.; Zhang, Q.; Lian, X. Overexpressed glutamine synthetase gene modifies nitrogen metabolism and abiotic stress responses in rice. Plant Cell Rep. 2009, 28, 527–537. [Google Scholar] [CrossRef]
- Amaro, A.C.E.; Baron, D.; Ono, E.O.; Rodrigues, J.D. Physiological effects of strobilurin and carboxamides on plants: An overview. Acta Physiol. Plant. 2019, 42, 4. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, Y.; Xie, Q.; Tong, H.; Chu, C. Crosstalk between brassinosteroid signaling and variable nutrient environments. Sci. China Life Sci. 2023, 66, 1231–1244. [Google Scholar] [CrossRef]
- Glaab, J.; Kaiser, W.M. Increased nitrate reductase activity in leaf tissue after application of the fungicide Kresoxim-methyl. Planta 1999, 207, 442–448. [Google Scholar] [CrossRef]
- Fagan, E.B.; Dourado Neto, D.; Vivian, R.; Franco, R.B.; Yeda, M.P.; Massignam, L.F.; Ferraz De Oliveira, R.; Martins, K.V. Effect of pyraclostrobin application on the photosynthesis rate, respiration, nitrate reductase activity and productivity of soybean crops. Bragantia 2010, 69, 771–777. [Google Scholar] [CrossRef]
- Nelson, K.A.; Meinhardt, C. Foliar boron and pyraclostrobin effects on corn yield. Agron. J. 2011, 103, 1352–1358. [Google Scholar] [CrossRef]
- Lima, J.D.; Moraes, W.d.S.; Modenese-Gorla da Silva, S.H. Physiological responses in banana plantlets treated with strobilurins. Semina Cienc. Agrar. 2012, 33, 77–86. [Google Scholar] [CrossRef]
- Mesara, S.N.; Dave, K.P.; Subramanian, R.B. Comparative transcriptome analysis elucidates positive physiological effects of foliar application of pyraclostrobin on tomato (Solanum lycopersicum L.). Physiol. Mol. Biol. Plants 2022, 28, 971–986. [Google Scholar] [CrossRef] [PubMed]
- Amaro, A.C.E.; Ramos, A.R.P.; Macedo, A.C.; Ono, E.O.; Rodrigues, J.D. Effects of the fungicides azoxystrobin, pyraclostrobin and boscalid on the physiology of Japanese cucumber. Sci. Hortic. 2018, 228, 66–75. [Google Scholar] [CrossRef]
- Ruske, R.E.; Gooding, M.J.; Jones, S.A. The effects of triazole and strobilurin fungicide programmes on nitrogen uptake, partitioning, remobilization and grain N accumulation in winter wheat cultivars. J. Agric. Sci. 2003, 140, 395–407. [Google Scholar] [CrossRef]
- Martinazzo, E.G.; Perboni, A.T.; Posso, D.A.; Aumonde, T.Z.; Bacarin, M.A. Growth analysis partitioning of assimilate in tomato plants cv. Micro-Tom submitted to nitrogen and pyraclostrobin. Semina Cienc. Agrar. 2015, 36, 3001–3012. [Google Scholar] [CrossRef]
- Joshi, J.; Harma, S.; Guruprasad, K. Foliar application of pyraclostrobin fungicide enhances the growth, rhizobial-nodule formation and nitrogenase activity in soybean (var. JS-335). Pestic. Biochem. Physiol. 2014, 114, 61–66. [Google Scholar] [CrossRef]
- Han, C.; Wang, L.; Lyu, J.; Shi, W.; Yao, L.; Fan, M.; Bai, M.-Y. Brassinosteroid signaling and molecular crosstalk with nutrients in plants. J. Genet. Genom. 2023, 50, 541–553. [Google Scholar] [CrossRef]
- Jia, Z.; Giehl, R.F.; Meyer, R.C.; Altmann, T.; von Wirén, N. Natural variation of BSK3 tunes brassinosteroid signaling to regulate root foraging under low nitrogen. Nat. Commun. 2019, 10, 2378. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.; Chen, J.; Yue, X.; Li, C.; Zhang, Q.; Resco de Dios, V.; Yao, Y.; Tan, W. Interaction of BES1 and LBD37 transcription factors modulates brassinosteroid-regulated root forging response under low nitrogen in Arabidopsis. Front. Plant Sci. 2022, 13, 998961. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, J.-J.; Wang, K.-X.; Xia, X.-J.; Shi, K.; Zhou, Y.-H.; Yu, J.-Q.; Zhou, J. BZR1 mediates brassinosteroid-induced autophagy and nitrogen starvation in tomato. Plant Physiol. 2019, 179, 671–685. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Giehl, R.F.; von Wirén, N. The root foraging response under low nitrogen depends on DWARF1-mediated brassinosteroid biosynthesis. Plant Physiol. 2020, 183, 998–1010. [Google Scholar] [CrossRef]
- Wang, T.; Li, M.; Yang, J.; Li, M.; Zhang, Z.; Gao, H.; Wang, C.; Tian, H. Brassinosteroid transcription factor BES1 modulates nitrate deficiency by promoting NRT2.1 and NRT2.2 transcription in Arabidopsis. Plant J. 2023, 114, 1443–1457. [Google Scholar] [CrossRef]
- Xuan, Y.H.; Duan, F.Y.; Je, B.I.; Kim, C.M.; Li, T.Y.; Liu, J.M.; Park, S.J.; Cho, J.H.; Kim, T.H.; von Wiren, N.; et al. Related to ABI3/VP1-Like 1 (RAVL1) regulates brassinosteroid-mediated activation of AMT1;2 in rice (Oryza sativa). J. Exp. Bot. 2017, 68, 727–737. [Google Scholar]
- Cai, H.; Chai, M.; Chen, F.; Huang, Y.; Zhang, M.; He, Q.; Liu, L.; Yan, M.; Qin, Y. HBI1 acts downstream of ERECTA and SWR1 in regulating inflorescence architecture through the activation of the brassinosteroid and auxin signaling pathways. New Phytol. 2021, 229, 414–428. [Google Scholar] [CrossRef]
- Chu, X.; Li, M.; Zhang, S.; Fan, M.; Han, C.; Xiang, F.; Li, G.; Wang, Y.; Xiang, C.B.; Wang, J.G. HBI1-TCP20 interaction positively regulates the CEPs-mediated systemic nitrate acquisition. J. Integr. Plant Biol. 2021, 63, 902–912. [Google Scholar] [CrossRef]
- An, Y.-Q.; Qin, Z.-T.; Li, D.-D.; Zhao, R.-Q.; Bi, B.-S.; Wang, D.-W.; Ma, D.-J.; Xi, Z. The combined formulation of brassinolide and pyraclostrobin increases biomass and seed yield by improving photosynthetic capacity in Arabidopsis thaliana. Front. Plant Sci. 2023, 14, 1138563. [Google Scholar] [CrossRef]
- Wei, S.; Li, X.; Lu, Z.; Zhang, H.; Ye, X.; Zhou, Y.; Li, J.; Yan, Y.; Pei, H.; Duan, F.; et al. A transcriptional regulator that boosts grain yields and shortens the growth duration of rice. Science 2022, 377, eabi8455. [Google Scholar] [CrossRef]
- Li, S.; Tian, Y.; Wu, K.; Ye, Y.; Yu, J.; Zhang, J.; Liu, Q.; Hu, M.; Li, H.; Tong, Y.; et al. Modulating plant growth-metabolism coordination for sustainable agriculture. Nature 2018, 560, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R.; Clarke, V.C. The nitrogen cost of photosynthesis. J. Exp. Bot. 2019, 70, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Muller, O.; Hirose, T.; Werger, M.J.A.; Hikosaka, K. Optimal use of leaf nitrogen explains seasonal changes in leaf nitrogen content of an understorey evergreen shrub. Ann. Bot. 2011, 108, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Chen, Y. The physiological response of photosynthesis to nitrogen deficiency. Plant Physiol. Biochem. 2021, 158, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Ookawa, T.; Naruoka, Y.; Yamazaki, T.; Suga, J.; Hirasawa, T. A comparison of the accumulation and partitioning of nitrogen in plants between two rice cultivars, Akenohoshi and Nipponbare, at the Ripening Stage. Plant Prod. Sci. 2003, 6, 172–178. [Google Scholar] [CrossRef]
- Uribelarrea, M.; Crafts-Brandner, S.J.; Below, F.E. Physiological N response of field-grown maize hybrids (Zea mays L.) with divergent yield potential and grain protein concentration. Plant Soil 2009, 316, 151–160. [Google Scholar] [CrossRef]
- Zhang, Q.; Tang, W.; Peng, S.; Li, Y. Limiting factors for panicle photosynthesis at the anthesis and grain filling stages in rice (Oryza sativa L.). Plant J. 2022, 109, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.-Z.; Zhang, H.-Q.; Li, P.; Zhang, D.-S.; Hao, X.-Y.; Gao, Z.-Q. Leaf nitrogen have a better relationship with photosynthesis performance across wheat species under elevated CO2 and drought. Plant Physiol. Biochem. 2021, 166, 964–973. [Google Scholar]
- Gaudinier, A.; Rodriguez-Medina, J.; Zhang, L.; Olson, A.; Liseron-Monfils, C.; Bågman, A.-M.; Foret, J.; Abbitt, S.; Tang, M.; Li, B.; et al. Transcriptional regulation of nitrogen-associated metabolism and growth. Nature 2018, 563, 259–264. [Google Scholar] [CrossRef]
- Hu, B.; Wang, W.; Chen, J.; Liu, Y.; Chu, C. Genetic improvement toward nitrogen-use efficiency in rice: Lessons and perspectives. Mol. Plant 2023, 16, 64–74. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate reductase regulates plant nitric oxide homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed]
- Dellero, Y. Manipulating amino acid metabolism to improve crop nitrogen use efficiency for a sustainable agriculture. Front. Plant Sci. 2020, 11, 602548. [Google Scholar] [CrossRef] [PubMed]
- Matejovic, I. Total nitrogen in plant material determinated by means of dry combustion: A possible alternative to determination by Kjeldahl digestion. Commun. Soil Sci. Plant Anal. 1995, 26, 2217–2229. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, Q.; Ding, L.; Sun, Y. Effect of cadmium toxicity on nitrogen metabolism in leaves of Solanum nigrum L. as a newly found cadmium hyperaccumulator. J. Hazard. Mater. 2008, 154, 818–825. [Google Scholar] [CrossRef]
- Aslam, M.; Huffaker, R.C. Role of nitrate and nitrite in the induction of nitrite reductase in leaves of barley seedlings. Plant Physiol. 1989, 91, 1152–1156. [Google Scholar] [CrossRef]
- Groat, R.G.; Vance, C.P. Root nodule enzymes of ammonia assimilation in alfalfa (Medicago sativa L.) developmental patterns and response to applied nitrogen. Plant Physiol. 1981, 67, 1198–1203. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | ID | Name | Log2FC | Family | ID | Name | Log2FC |
|---|---|---|---|---|---|---|---|
| ARF | AT1G19220 | ARF19 | −0.26 | MYB | AT1G09770 | CDC5 | 0.12 |
| AT1G59750 | ARF1 | −0.17 | AT1G18570 | MYB51 | −0.41 | ||
| AT3G61830 | ARF18 | 0.26 | AT1G18710 | MYB47 | 0.78 | ||
| ARR-B | AT2G01760 | ARR14 | −0.36 | AT3G11450 | - | 0.22 | |
| AT2G25180 | ARR12 | −0.26 | AT4G21440 | MYB102 | 0.60 | ||
| AT3G16857 | ARR1 | −0.19 | AT4G37260 | MYB73 | −0.71 | ||
| AT4G31920 | ARR10 | −0.33 | AT4G38620 | MYB4 | 0.43 | ||
| B3 | AT1G01030 | NGA3 | 0.89 | AT5G06110 | - | 0.46 | |
| AT1G16640 | - | 0.70 | MYB-related | AT1G19000 | - | −0.58 | |
| AT2G36080 | NGAL1 | −0.63 | AT1G70000 | - | −0.57 | ||
| AT2G46870 | NGA1 | 0.41 | AT1G74840 | - | −0.73 | ||
| BBR/BPC | AT2G21240 | BPC4 | −0.19 | AT3G21430 | ALY3 | −0.25 | |
| C2H2 zinc-finger protein | AT1G24625 | ZFP7 | −0.54 | AT4G01280 | RVE5 | −0.44 | |
| AT1G27730 | ZAT10 | −0.52 | AT5G17300 | RVE1 | −1.09 | ||
| AT1G55110 | IDD7 | −0.36 | AT5G45420 | MAMYB | 0.21 | ||
| AT1G72050 | TFIIIA | 0.24 | NAC | AT2G33480 | NAC041 | 0.30 | |
| AT3G13810 | IDD11 | −0.41 | AT3G10500 | NAC053 | 0.32 | ||
| AT5G14140 | - | −0.45 | AT3G15500 | NAC055 | −1.74 | ||
| AT5G25160 | ZFP3 | −0.63 | AT3G49530 | NAC062 | 0.17 | ||
| AT5G52010 | - | 0.45 | AT5G08790 | NAC081 | −0.43 | ||
| C3H zinc-finger protien | AT3G02830 | ZFN1 | −0.18 | AT5G24590 | NAC091 | 0.23 | |
| AT3G08505 | MKRN | −0.41 | RAV | AT1G13260 | RAV1 | −1.17 | |
| AT3G55980 | SZF1 | −0.45 | AT1G25560 | TEM1 | −1.17 | ||
| AT5G51980 | - | 0.27 | AT1G68840 | RAV2 | −0.79 | ||
| CO-like | AT3G02380 | COL2 | −0.48 | SBP | AT1G20980 | SPL14 | −0.18 |
| AT5G57660 | COL5 | −0.11 | AT1G53160 | SPL4 | −0.65 | ||
| AT5G59990 | - | −0.58 | AT5G43270 | SPL2 | 0.26 | ||
| DBB | AT1G06040 | BBX24 | −0.15 | SRS | AT4G36260 | SRS2 | 0.90 |
| AT4G38960 | - | −0.41 | TALE | AT4G36870 | BLH2 | −0.31 | |
| Dof | AT1G51700 | DOF1.7 | −0.30 | TCP | AT1G72010 | TCP22 | −0.23 |
| AT3G47500 | CDF3 | −0.20 | AT3G15030 | TCP4 | 0.50 | ||
| AT5G60200 | DOF5.3 | −0.47 | AT3G47620 | TCP14 | −0.28 | ||
| ERF | AT1G33760 | ERF022 | −0.87 | AT4G18390 | TCP2 | −0.26 | |
| AT1G64380 | ERF061 | 0.57 | Trihelix | AT1G13450 | GT−1 | −0.25 | |
| AT2G20880 | ERF053 | 0.77 | AT1G33240 | AT-GTL1 | −0.13 | ||
| AT2G44940 | ERF034 | 0.98 | AT5G28300 | - | 0.67 | ||
| AT2G46310 | CRF5 | 0.85 | AT5G63420 | emb2746 | 0.64 | ||
| AT3G15210 | ERF4 | −0.26 | WRKY | AT1G69310 | WRKY57 | −0.40 | |
| AT4G06746 | RAP2−9 | −1.79 | AT2G03340 | WRKY3 | −0.20 | ||
| AT4G17490 | ERF6 | −0.41 | AT2G38470 | WRKY33 | −0.41 | ||
| AT4G23750 | CRF2 | 0.98 | AT3G01970 | WRKY45 | −1.27 | ||
| AT5G44210 | ERF9 | 0.64 | AT5G07100 | WRKY26 | −0.90 | ||
| AT5G47230 | ERF5 | −0.40 | AT5G13080 | WRKY75 | −0.70 | ||
| AT5G53290 | CRF3 | 0.68 | AT5G45260 | RRS1 | −0.25 | ||
| FAR13 | AT3G06250 | FRS7 | 0.28 | AT5G52830 | WRKY27 | −0.54 | |
| G2-like | AT1G14600 | - | −0.46 | YABBY | AT1G08465 | YAB2 | −0.29 |
| AT1G25550 | HHO3 | −0.77 | ZF-HD | AT2G18350 | ZHD6 | −0.39 | |
| AT1G68670 | HHO2 | −0.83 | AT3G28920 | ZHD9 | 0.37 | ||
| AT3G46640 | PCL1 | −0.37 | bHLH | AT1G06170 | BHLH89 | 0.60 | |
| GATA | AT3G24050 | GATA1 | 0.24 | AT1G18400 | BEE1 | 0.38 | |
| GRAS | AT1G07530 | SCL14 | −0.19 | AT2G20180 | PIF1 | −0.30 | |
| AT1G66350 | RGL1 | 0.52 | AT3G07340 | BHLH62 | −0.41 | ||
| AT3G46600 | SCL30 | −0.66 | AT3G23690 | BHLH77 | −0.45 | ||
| GeBP | AT4G00238 | STKL1 | 0.43 | AT4G29100 | BHLH68 | −0.38 | |
| AT4G25210 | GEBPL | 0.27 | AT4G30980 | LRL2 | 0.63 | ||
| HD-ZIP | AT4G37790 | HAT22 | 0.24 | AT5G38860 | BIM3 | −0.41 | |
| AT5G65310 | ATHB5 | 0.16 | AT5G39860 | PRE1 | 0.38 | ||
| HSF | AT3G51910 | HSFA7A | −0.61 | AT5G50915 | BHLH137 | 0.24 | |
| AT5G03720 | HSFA3 | 0.55 | AT5G61270 | BHLH72 | −0.46 | ||
| MIKC_MADS | AT2G45660 | SOC1 | −0.43 | bZIP | AT1G43700 | VIP1 | −0.23 |
| AT3G57230 | AGL16 | −0.25 | AT3G51960 | BZIP24 | −0.61 | ||
| AT5G15800 | 1-Sep | −1.09 | AT3G62420 | BZIP53 | −0.41 | ||
| PLATZ | AT4G17900 | - | −0.20 | AT4G02640 | BZO2H1 | −0.24 | |
| BTB/POZ | AT4G37610 | BT5 | −0.92 | AT4G36730 | GBF1 | 0.20 | |
| AT5G63160 | BT1 | −1.19 | AT5G24800 | BZIP9 | −0.28 | ||
| AT3G48360 | BT2 | −0.96 | AT5G28770 | BZIP63 | −0.22 | ||
| AT5G67480 | BT4 | −0.83 |
| Class | Compound Name | log2FC |
|---|---|---|
| Flavonoids and its derivatives | Idaein chloride | −0.50 |
| Tangeretin | −0.74 | |
| Sakuranetin | −0.49 | |
| Apigenin C-pentoside | −0.50 | |
| O-methylnaringenin C-pentoside | −0.55 | |
| (-)-Gallocatechin | −0.93 | |
| Selgin C-hexoside | −0.45 | |
| Methyl-Naringenin C-pentoside | −0.55 | |
| Apigenin 4-O-rhamnoside | 0.62 | |
| Carbohydrates and its derivatives | D-Glucose | −0.88 |
| alpha-D-Galactose 1-phosphate | 0.28 | |
| Sorbitol 6-phosphate | 0.28 | |
| D-Glucose 6-phosphate | 0.33 | |
| D-Erythrose 4-phosphate | 0.55 | |
| L-Gulono−1,4-lactone | −1.43 | |
| D-arabitol | 0.38 | |
| Amino acid and its derivatives | L-arginine | 0.56 |
| N-Acetyl-L-glutamic acid | 0.29 | |
| Nepsilon-acetyl-L-lysine | −0.84 | |
| Aminomalonic acid | 0.38 | |
| Aspartic acid di-O-glucoside | 1.92 | |
| D-asparagine | −0.33 | |
| Terpenoids | Dihydroartemisinic acid | −0.95 |
| (-)-Menthol | −4.29 | |
| Artemether | −0.39 | |
| (-)-trans-carveol | −0.68 | |
| 18-Nor−4,15-dihydroxyabieta−8,11,13-tried−7-one | 0.84 | |
| Indoles and derivatives | Indole-3-carboxaldehyde | −1.10 |
| 3-indoleacetonitrile | −3.52 | |
| Indole-3-acetamide | 0.49 | |
| Nucleotide and its derivates | Adenosine 5′-Diphosphate | 0.81 |
| Cytidine | −0.98 | |
| N-(9H-Purin−6-ylcarbamoyl)threonine | 0.35 | |
| Alkaloids and derivatives | vasicine | −0.42 |
| Arecoline | −0.86 | |
| Dihydroxy tomatidine-O-hexosyl-O-rhamnoside | −1.16 | |
| Amines | Methylguanidine | −0.53 |
| 2-Phenylethylamine | −0.64 | |
| Dl-Dihydrosphingosine | −0.38 | |
| Glycerolipids | Monopalmitin | −0.65 |
| MAG (18:4) | −0.84 | |
| Organic acid and its derivatives | Scopolamine | −1.26 |
| Adipic acid | 0.55 | |
| Fatty acyls | Erucic acid | 0.51 |
| Punicic acid | −0.58 | |
| Phenylpropanoids | Calceolarioside B | −0.63 |
| Isoscopoletin | −0.69 | |
| Benzoic acids and derivatives | 3,4-Dihydroxyphenylacetic acid | −0.90 |
| Methyl vanillate | −1.21 | |
| Cinnamic acids and derivatives | N-p-Coumaroyl putrescine | −1.16 |
| N′-p-Coumaroyl agmatine | −0.52 | |
| Phenolic acids | 2,6-Di-tert-butylphenol | −0.56 |
| Denin | 2.64 | |
| Phospholipid | LysoPC 18:1 | −0.73 |
| Ethers | Ambroxane | 0.58 |
| Organoheterocyclic | Allantoin | 0.29 |
| Vitamins | Orotic acid | −1.21 |
| Name | Class | log2FC | ||
|---|---|---|---|---|
| BL + Pyr vs. CK | BL + Pyr vs. BL | BL + Pyr vs. Pyr | ||
| Monopalmitin | Glycerolipids | −0.65 | −0.65 | −0.67 |
| Dihydroartemisinic acid | Terpenoids | −0.95 | −0.95 | −0.69 |
| Indole−3-Carboxaldehyde | Indoles and derivatives | −0.99 | −0.99 | −0.58 |
| D-Glucose | Carbohydrates And Its Derivatives | −0.88 | −0.88 | −0.63 |
| (-)-Menthol | Terpenoids | −4.29 | −4.29 | −2.19 |
| Indole−3-acetamide | Indoles and derivatives | 0.60 | 0.60 | 0.82 |
| L-arginine | Amino Acid And Derivatives | 0.56 | 0.56 | 0.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, Y.-Q.; Ma, D.-J.; Xi, Z. Multi-Omics Analysis Reveals Synergistic Enhancement of Nitrogen Assimilation Efficiency via Coordinated Regulation of Nitrogen and Carbon Metabolism by Co-Application of Brassinolide and Pyraclostrobin in Arabidopsis thaliana. Int. J. Mol. Sci. 2023, 24, 16435. https://doi.org/10.3390/ijms242216435
An Y-Q, Ma D-J, Xi Z. Multi-Omics Analysis Reveals Synergistic Enhancement of Nitrogen Assimilation Efficiency via Coordinated Regulation of Nitrogen and Carbon Metabolism by Co-Application of Brassinolide and Pyraclostrobin in Arabidopsis thaliana. International Journal of Molecular Sciences. 2023; 24(22):16435. https://doi.org/10.3390/ijms242216435
Chicago/Turabian StyleAn, Ya-Qi, De-Jun Ma, and Zhen Xi. 2023. "Multi-Omics Analysis Reveals Synergistic Enhancement of Nitrogen Assimilation Efficiency via Coordinated Regulation of Nitrogen and Carbon Metabolism by Co-Application of Brassinolide and Pyraclostrobin in Arabidopsis thaliana" International Journal of Molecular Sciences 24, no. 22: 16435. https://doi.org/10.3390/ijms242216435
APA StyleAn, Y.-Q., Ma, D.-J., & Xi, Z. (2023). Multi-Omics Analysis Reveals Synergistic Enhancement of Nitrogen Assimilation Efficiency via Coordinated Regulation of Nitrogen and Carbon Metabolism by Co-Application of Brassinolide and Pyraclostrobin in Arabidopsis thaliana. International Journal of Molecular Sciences, 24(22), 16435. https://doi.org/10.3390/ijms242216435