
Disruption of Intranasal GnRH Neuronal Migration Route into the Brain Induced by Proinflammatory Cytokine IL-6: Ex Vivo and In Vivo Rodent Models


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. GnRH Neuron Distribution after IL-6 Prenatal Treatment in Mouse Fetuses
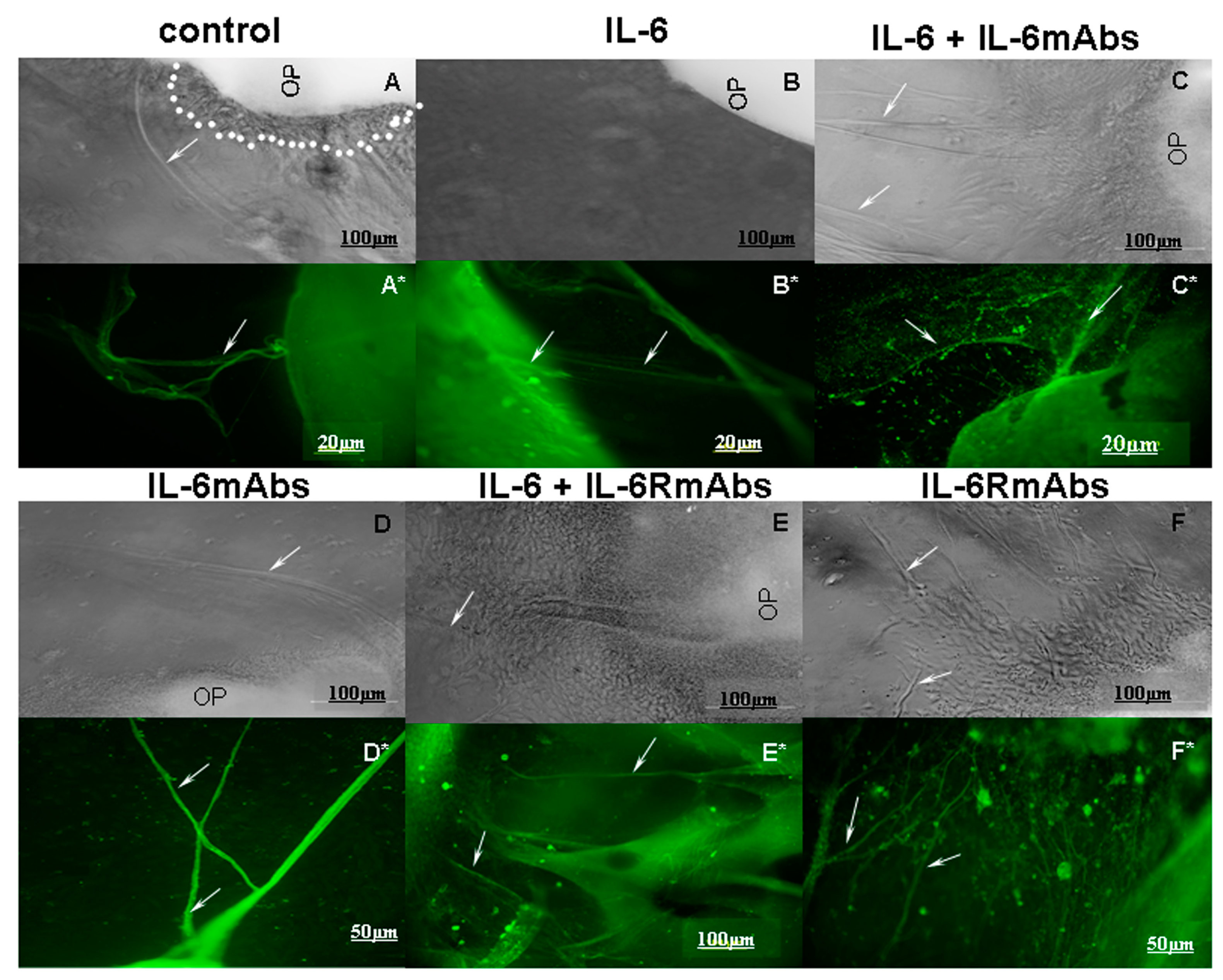
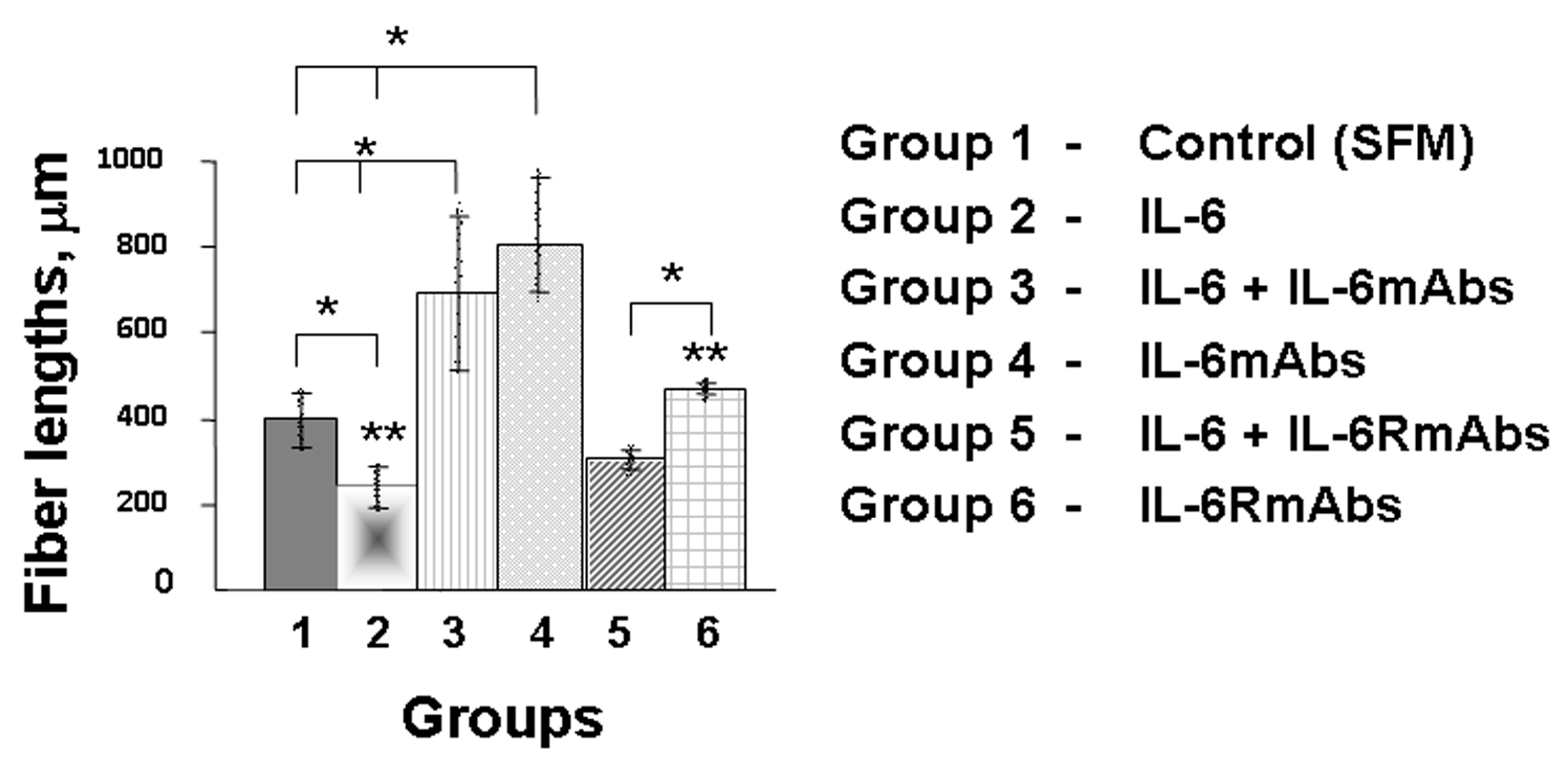
2.2. Effect of IL-6 on the Growth of Olfactory Nerve Fibers in an Organotypic Culture of Fetal Mouse Placodes
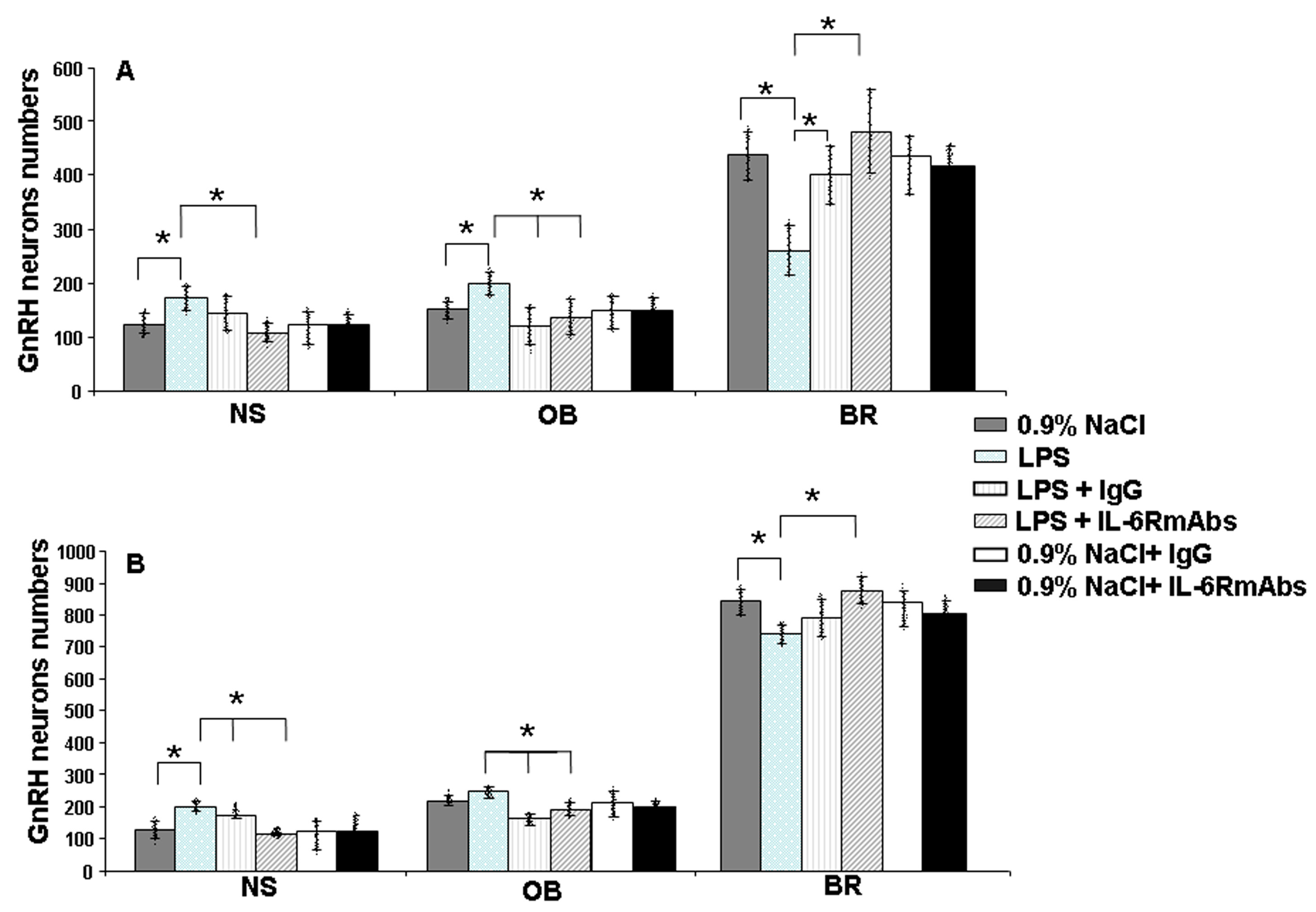
2.3. GnRH-ir Neuron Distribution after Prenatal LPS and IL-6RmAbs or IgG Treatment in Rat Fetuses
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design (In Vivo Model)
4.2. GnRH Immunohistochemical Staining in Fetuses
4.3. Preparation of Fetal Olfactory Mouse Placodes (Ex Vivo Model)
4.3.1. Immunohistochemistry
4.3.2. The Measurement of Olfactory Fiber Growth
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BR | brain |
| BSA | bovine serum albumin |
| ED | day of embryonic development (embryonic day) |
| GnRH | gonadotropin releasing hormone |
| HPG | hypothalamic–pituitary–gonadal (axis) |
| IgG | immunoglobulin G |
| IL | interleukin |
| IL-6mAbs | monoclonal anti-interleukin 6 antibodies |
| IL-6RmAbs | monoclonal anti-interleukin 6 receptor antibodies |
| ir | immunoreactive |
| LIF | leukemia inhibitory factor |
| LPS | lipopolysaccharide |
| MCP-1 | monocyte chemotactic protein 1 |
| MIA | maternal immune activation |
| NGS | normal goat serum |
| NS | nasal area |
| OB | olfactory bulbs |
| OE | olfactory epithelium |
| PAF | paraformaldehyde |
| PBS | phosphate buffered saline |
| PolyAbs | polyclonal antibodies |
| SFM | serum-free medium |
| TNFα | tumor necrosis factor α |
| VNO | vomeronasal organ |
References
- Arck, P.C.; Hecher, K. Fetomaternal immune cross-talk and its consequences for maternal and offspring’s health. Nat. Med. 2013, 19, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Estes, M.L.; Prendergast, K.; MacMahon, J.A.; Cameron, S.; Aboubechara, J.P.; Farrelly, K.; Sell, G.L.; Haapanen, L.; Schauer, J.D.; Horta, A.; et al. Baseline immunoreactivity before pregnancy and poly(I:C) dose combine to dictate susceptibility and resilience of offspring to maternal immune activation. Brain Behav. Immun. 2020, 88, 619–630. [Google Scholar] [CrossRef]
- Wu, S.; Wolfe, A. Signaling of cytokines is important in regulation of GnRH neurons. Mol. Neurobiol. 2012, 45, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Pei, D.E.; Yang, R.D.; Wan, C.L.; Ye, Y.M.; Peng, S.S.; Zeng, Q.Q.; Yu, Y. Prenatal maternal vaginal inflammation increases anxiety and alters HPA axis signalling in adult male mice. Int. J. Dev. Neurosci. 2019, 75, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Zakharova, L.; Sharova, V.; Izvolskaia, M. Mechanisms of Reciprocal Regulation of Gonadotropin-Releasing Hormone (GnRH)-Producing and Immune Systems: The Role of GnRH, Cytokines and Their Receptors in Early Ontogenesis in Normal and Pathological Conditions. Int. J. Mol. Sci. 2020, 22, 114. [Google Scholar] [CrossRef]
- Wang, H.; Yang, L.L.; Hu, Y.F.; Wang, B.W.; Huang, Y.Y.; Zhang, C.; Chen, Y.H.; Xu, D.X. Maternal LPS exposure during pregnancy impairs testicular development, steroidogenesis and spermatogenesis in male offspring. PLoS ONE 2014, 9, e106786. [Google Scholar] [CrossRef]
- Izvolskaia, M.; Ignatiuk, V.; Ismailova, A.; Sharova, V.; Zakharova, L. IgG modulation in male mice with reproductive failure after prenatal inflammation. Reproduction 2021, 161, 669–679. [Google Scholar] [CrossRef]
- Hudalla, H.; Karenberg, K.; Kuon, R.J.; Pöschl, J.; Tschada, R.; Frommhold, D. LPS-induced maternal inflammation promotes fetal leukocyte recruitment and prenatal organ infiltration in mice. Pediatr. Res. 2018, 84, 757–764. [Google Scholar] [CrossRef]
- Bernardi, M.M.; Kirsten, T.B.; Matsuoka, S.M.; Teodorov, E.; Habr, S.F.; Penteado, S.H.; Palermo-Neto, J. Prenatal lipopolysaccharide exposure affects maternal behavior and male offspring sexual behavior in adulthood. Neuroimmunomodulation 2010, 17, 47–55. [Google Scholar] [CrossRef]
- Rose, D.R.; Careaga, M.; Van de Water, J.; McAllister, K.; Bauman, M.D.; Ashwood, P. Long-term altered immune responses following fetal priming in a non-human primate model of maternal immune activation. Brain Behav. Immun. 2017, 63, 60–70. [Google Scholar] [CrossRef]
- Izvolskaia, M.; Sharova, V.; Zakharova, L. Perinatal Inflammation Reprograms Neuroendocrine, Immune, and Reproductive Functions: Profile of Cytokine Biomarkers. Inflammation 2020, 43, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Yasumatsu, K.; Nagao, J.I.; Arita-Morioka, K.I.; Narita, Y.; Tasaki, S.; Toyoda, K.; Ito, S.; Kido, H.; Tanaka, Y. Bacterial-induced maternal interleukin-17A pathway promotes autistic-like behaviors in mouse offspring. Exp. Anim. 2020, 69, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Sharova, V.S.; Izvolskaia, M.S.; Zakharova, L.A. Lipopolysaccharide-induced maternal inflammation affects the gonadotropin-releasing hormone neuron development in fetal mice. Neuroimmunomodulation 2015, 22, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Ignatiuk, V.; Izvolskaia, M.; Sharova, V.; Zakharova, L. Disruptions in Hypothalamic-Pituitary-Gonadal Axis Development and Their IgG Modulation after Prenatal Systemic Inflammation in Male Rats. Int. J. Mol. Sci. 2023, 24, 2726. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, E.Y.; Patterson, P.H. Activation of the maternal immune system induces endocrine changes in the placenta via IL-6. Brain Behav. Immun. 2011, 25, 604–615. [Google Scholar] [CrossRef]
- Choi, G.B.; Yim, Y.S.; Wong, H.; Kim, S.; Kim, H.; Kim, S.V.; Hoeffer, C.A.; Littman, D.R.; Huh, J.R. The maternal interleukin-17a pathway in mice promotes autism-like phenotypes in offspring. Science 2016, 351, 933–999. [Google Scholar] [CrossRef]
- Hwang, H.M.; Ku, R.Y.; Hashimoto-Torii, K. Prenatal Environment That Affects Neuronal Migration. Front. Cell Dev. Biol. 2019, 7, 138. [Google Scholar] [CrossRef]
- Du, M.R.; Wang, S.C.; Li, D.J. The integrative roles of chemokines at the maternal-fetal interface in early pregnancy. Cell Mol. Immunol. 2014, 11, 438–448. [Google Scholar] [CrossRef]
- Oskvig, D.B.; Elkahloun, A.G.; Johnson, K.R.; Phillips, T.M.; Herkenham, M. Maternal immune activation by LPS selectively alters specific gene expression profiles of interneuron migration and oxidative stress in the fetus without triggering a fetal immune response. Brain Behav. Immun. 2012, 26, 623–634. [Google Scholar] [CrossRef]
- Lombardo, M.V.; Moon, H.M.; Su, J.; Palmer, T.D.; Courchesne, E.; Pramparo, T. Maternal immune activation dysregulation of the fetal brain transcriptome and relevance to the pathophysiology of autism spectrum disorder. Mol. Psychiatry 2018, 23, 1001–1013. [Google Scholar] [CrossRef]
- Rasmussen, J.M.; Graham, A.M.; Entringer, S.; Gilmore, J.H.; Styner, M.; Fair, D.A.; Wadhwa, P.D.; Buss, C. Maternal Interleukin-6 concentration during pregnancy is associated with variation in frontolimbic white matter and cognitive development in early life. Neuroimage 2019, 185, 825–835. [Google Scholar] [CrossRef]
- Han, A.R.; Lee, S.K. Immune modulation of i.v. immunoglobulin in women with reproductive failure. Reprod. Med. Biol. 2018, 17, 115–124. [Google Scholar] [CrossRef]
- Domínguez-Soto, Á.; Simón-Fuentes, M.; de Las Casas-Engel, M.; Cuevas, V.D.; López-Bravo, M.; Domínguez-Andrés, J.; Saz-Leal, P.; Sancho, D.; Ardavín, C.; Ochoa-Grullón, J.; et al. IVIg Promote Cross-Tolerance against Inflammatory Stimuli In Vitro and In Vivo. J. Immunol. 2018, 201, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Sawa, T.; Kinoshita, M.; Inoue, K.; Ohara, J.; Moriyama, K. Immunoglobulin for Treating Bacterial Infections: One More Mechanism of Action. Antibodies 2019, 8, 52. [Google Scholar] [CrossRef]
- Berardicurti, O.; Ruscitti, P.; Ursini, F.; D’Andrea, S.; Ciaffi, J.; Meliconi, R.; Iagnocco, A.; Cipriani, P.; Giacomelli, R. Mortality in tocilizumab-treated patients with COVID-19: A systematic review and meta-analysis. Clin. Exp. Rheumatol. 2020, 38, 1247–1254. [Google Scholar] [PubMed]
- Noyola, D.E.; Demmler, G.J.; Nelson, C.T.; Griesser, C.; Williamson, W.D.; Atkins, J.T.; Rozelle, J.; Turcich, M.; Llorente, A.M.; Sellers-Vinson, S.; et al. Houston Congenital CMV Longitudinal Study Group. Early predictors of neurodevelopmental outcome in symptomatic congenital cytomegalovirus infection. J. Pediatr. 2001, 138, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Ignatiuk, V.M.; Izvolskaya, M.S.; Sharova, V.S.; Voronova, S.N.; Zakharova, L.A. Disruptions in the reproductive system of female rats after prenatal lipopolysaccharide-induced immunological stress: Role of sex steroids. Stress 2019, 22, 133–141. [Google Scholar] [CrossRef]
- Barabás, K.; Szabó-Meleg, E.; Ábrahám, I.M. Effect of Inflammation on Female Gonadotropin-Releasing Hormone (GnRH) Neurons: Mechanisms and Consequences. Int. J. Mol. Sci. 2020, 21, 529. [Google Scholar] [CrossRef] [PubMed]
- Schwanzel-Fukuda, M.; Pfaff, D.W. Origin of luteinizing hormone-releasing hormone neurons. Nature 1989, 338, 161–164. [Google Scholar] [CrossRef]
- Wray, S. From nose to brain: Development of gonadotrophin-releasing hormone-1 neurones. J. Neuroendocrinol. 2010, 22, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Klenke, U.; Taylor-Burds, C. Culturing embryonic nasal explants for developmental and physiological study. Curr. Protoc. Neurosci. 2012, 59, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Daikoku-Ishido, H.; Okamura, Y.; Yanaihara, N.; Daikoku, S. Development of the hypothalamic luteinizing hormone-releasing hormone-containing neuron system in the rat: In vivo and in transplantation studies. Dev. Biol. 1990, 140, 374–387. [Google Scholar] [CrossRef]
- Yoshida, K.; Tobet, S.A.; Crandall, J.E.; Jimenez, T.P.; Schwarting, G.A. The migration of luteinizing hormone-releasing hormone neurons in the developing rat is associated with a transient, caudal projection of the vomeronasal nerve. J. Neurosci. 1995, 15, 7769–7777. [Google Scholar] [CrossRef] [PubMed]
- Schwanzel-Fukuda, M. Origin and migration of luteinizing hormone-releasing hormone neurons in mammals. Microsc. Res. Tech. 1999, 44, 2–10. [Google Scholar] [CrossRef]
- Hoffman, G.E.; Finch, C.E. LHRH neurons in the female C57BL/6J mouse brain during reproductive aging: No loss up to middle age. Neurobiol. Aging 1986, 7, 45–48. [Google Scholar] [CrossRef]
- Oleari, R.; Massa, V.; Cariboni, A.; Lettieri, A. The Differential Roles for Neurodevelopmental and Neuroendocrine Genes in Shaping GnRH Neuron Physiology and Deficiency. Int. J. Mol. Sci. 2021, 22, 9425. [Google Scholar] [CrossRef]
- Chachlaki, K.; Oleari, R. Editorial: Physiological and pathological aspects of GnRH neuron system development. Front. Endocrinol. 2023, 14, 1268663. [Google Scholar] [CrossRef]
- Bornstein, S.R.; Rutkowski, H.; Vrezas, I. Cytokines and steroidogenesis. Mol. Cell Endocrinol. 2004, 215, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Li, J.; Garbett, K.; Mirnics, K.; Patterson, P.H. Maternal immune activation alters fetal brain development through interleukin-6. J. Neurosci. 2007, 27, 10695–10702. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.L.; Hsiao, E.Y.; Yan, Z.; Mazmanian, S.K.; Patterson, P.H. The placental interleukin-6 signaling controls fetal brain development and behavior. Brain Behav. Immun. 2017, 62, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Mirabella, F.; Desiato, G.; Mancinelli, S.; Fossati, G.; Rasile, M.; Morini, R.; Markicevic, M.; Grimm, C.; Amegandjin, C.; Termanini, A.; et al. Prenatal interleukin 6 elevation increases glutamatergic synapse density and disrupts hippocampal connectivity in offspring. Immunity 2021, 54, 2611–2631.e8. [Google Scholar] [CrossRef]
- Ashdown, H.; Dumont, Y.; Ng, M.; Poole, S.; Boksa, P.; Luheshi, G.N. The role of cytokines in mediating effects of prenatal infection on the fetus: Implications for schizophrenia. Mol. Psychiatry 2006, 11, 47–55. [Google Scholar] [CrossRef]
- Fueshko, S.; Wray, S. LHRH cells migrate on peripherin fibers in embryonic olfactory explant cultures: An in vitro model for neurophilic neuronal migration. Dev. Biol. 1994, 166, 331–348. [Google Scholar] [CrossRef] [PubMed]
- Kyvelidou, C.; Sotiriou, D.; Zerva, I.; Athanassakis, I. Protection Against Lipopolysaccharide-Induced Immunosuppression by IgG and IgM. Shock 2018, 49, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Chaigne, B.; Mouthon, L. Mechanisms of action of intravenous immunoglobulin. Transfus. Apher. Sci. 2017, 56, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, M.; Laskou, F.; Stapleton, P.P.; Hadavi, S.; Dasgupta, B. Tocilizumab (Actemra). Hum. Vaccin. Immunother. 2017, 13, 1972–1988. [Google Scholar] [CrossRef]
- Wakabayashi, A.; Sawada, K.; Nakayama, M.; Toda, A.; Kimoto, A.; Mabuchi, S.; Kinose, Y.; Nakamura, K.; Takahashi, K.; Kurachi, H.; et al. Targeting interleukin-6 receptor inhibits preterm delivery induced by inflammation. Mol. Hum. Reprod. 2013, 19, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Bai, C.; Liu, Y. Interleukin-6 contributes to myocardial damage in pregnant rats with reduced uterine perfusion pressure. Braz. J. Med. Biol. Res. 2018, 51, e6921. [Google Scholar] [CrossRef]
- Tzekova, N.; Heinen, A.; Bunk, S.; Hermann, C.; Hartung, H.P.; Reipert, B.; Küry, P. Immunoglobulins stimulate cultured Schwann cell maturation and promote their potential to induce axonal outgrowth. J. Neuroinflamm. 2015, 12, 107. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharova, V.; Ignatiuk, V.; Izvolskaia, M.; Zakharova, L. Disruption of Intranasal GnRH Neuronal Migration Route into the Brain Induced by Proinflammatory Cytokine IL-6: Ex Vivo and In Vivo Rodent Models. Int. J. Mol. Sci. 2023, 24, 15983. https://doi.org/10.3390/ijms242115983
Sharova V, Ignatiuk V, Izvolskaia M, Zakharova L. Disruption of Intranasal GnRH Neuronal Migration Route into the Brain Induced by Proinflammatory Cytokine IL-6: Ex Vivo and In Vivo Rodent Models. International Journal of Molecular Sciences. 2023; 24(21):15983. https://doi.org/10.3390/ijms242115983
Chicago/Turabian StyleSharova, Viktoria, Vasilina Ignatiuk, Marina Izvolskaia, and Liudmila Zakharova. 2023. "Disruption of Intranasal GnRH Neuronal Migration Route into the Brain Induced by Proinflammatory Cytokine IL-6: Ex Vivo and In Vivo Rodent Models" International Journal of Molecular Sciences 24, no. 21: 15983. https://doi.org/10.3390/ijms242115983
APA StyleSharova, V., Ignatiuk, V., Izvolskaia, M., & Zakharova, L. (2023). Disruption of Intranasal GnRH Neuronal Migration Route into the Brain Induced by Proinflammatory Cytokine IL-6: Ex Vivo and In Vivo Rodent Models. International Journal of Molecular Sciences, 24(21), 15983. https://doi.org/10.3390/ijms242115983