HSP90AB1 Is a Host Factor Required for Transmissible Gastroenteritis Virus Infection

 , and
, and
Abstract
:1. Introduction
2. Results
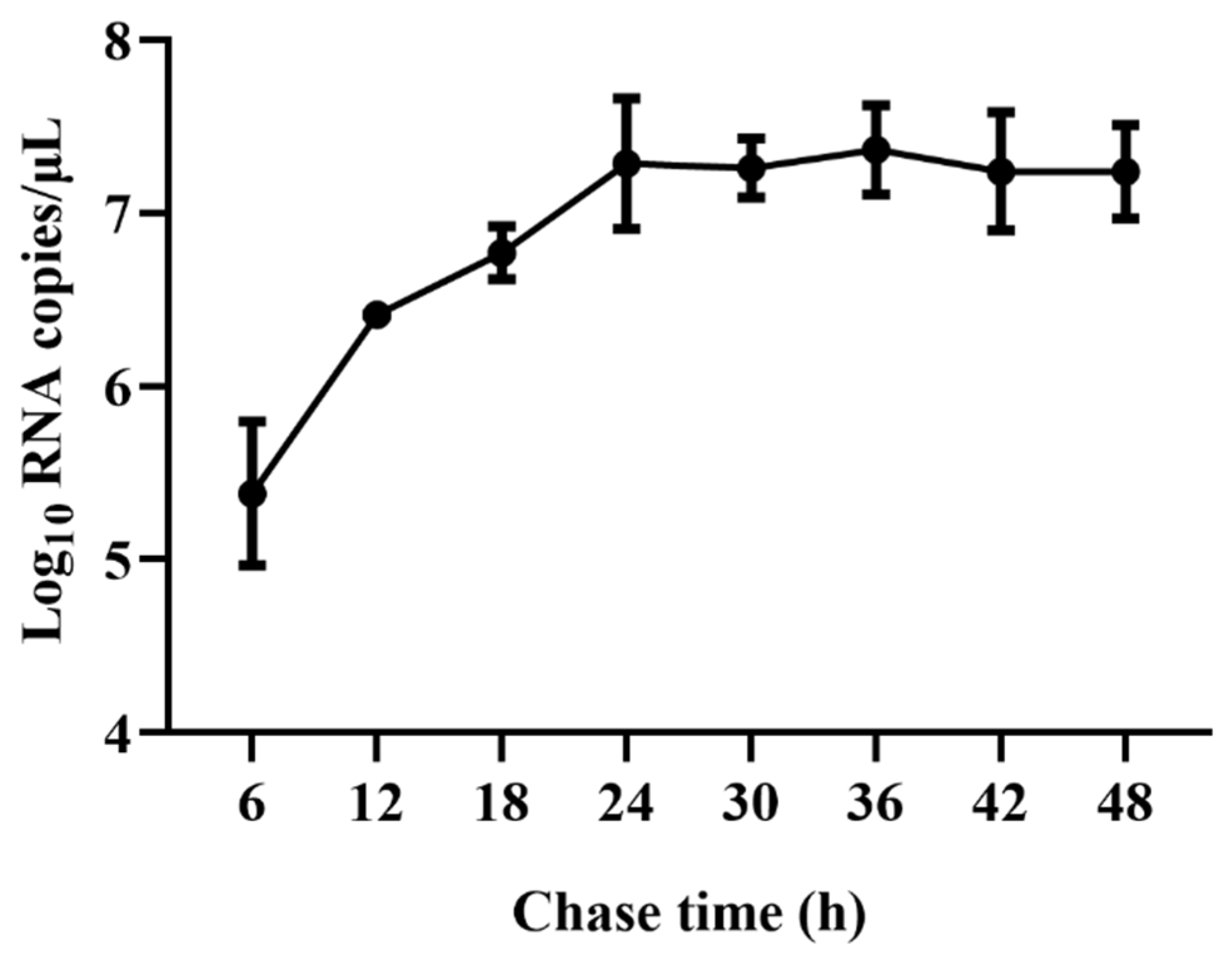
2.1. Viral Replication Kinetics of TGEV in ST Cells
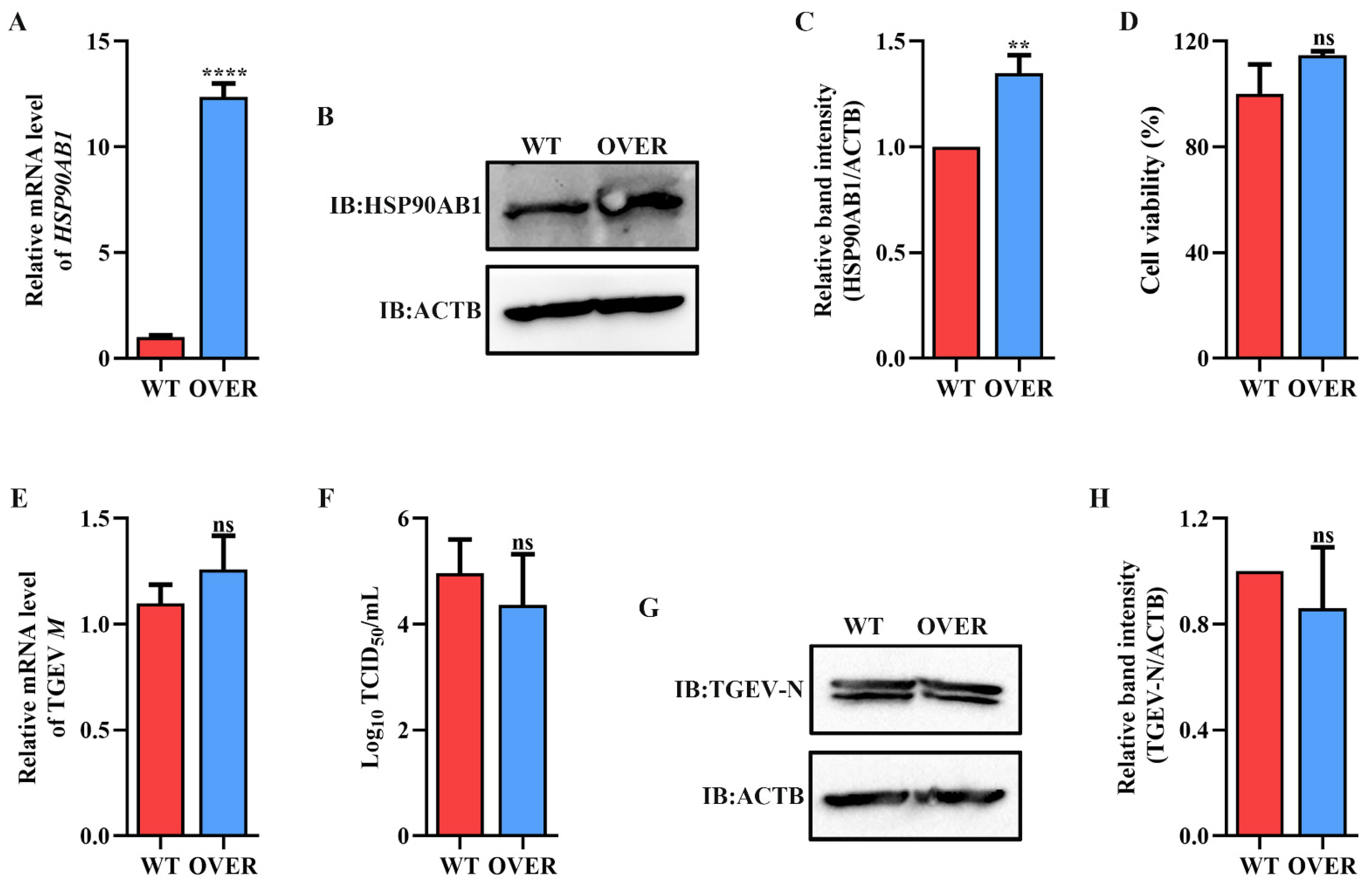
2.2. HSP90AB1 Enhances TGEV Infection
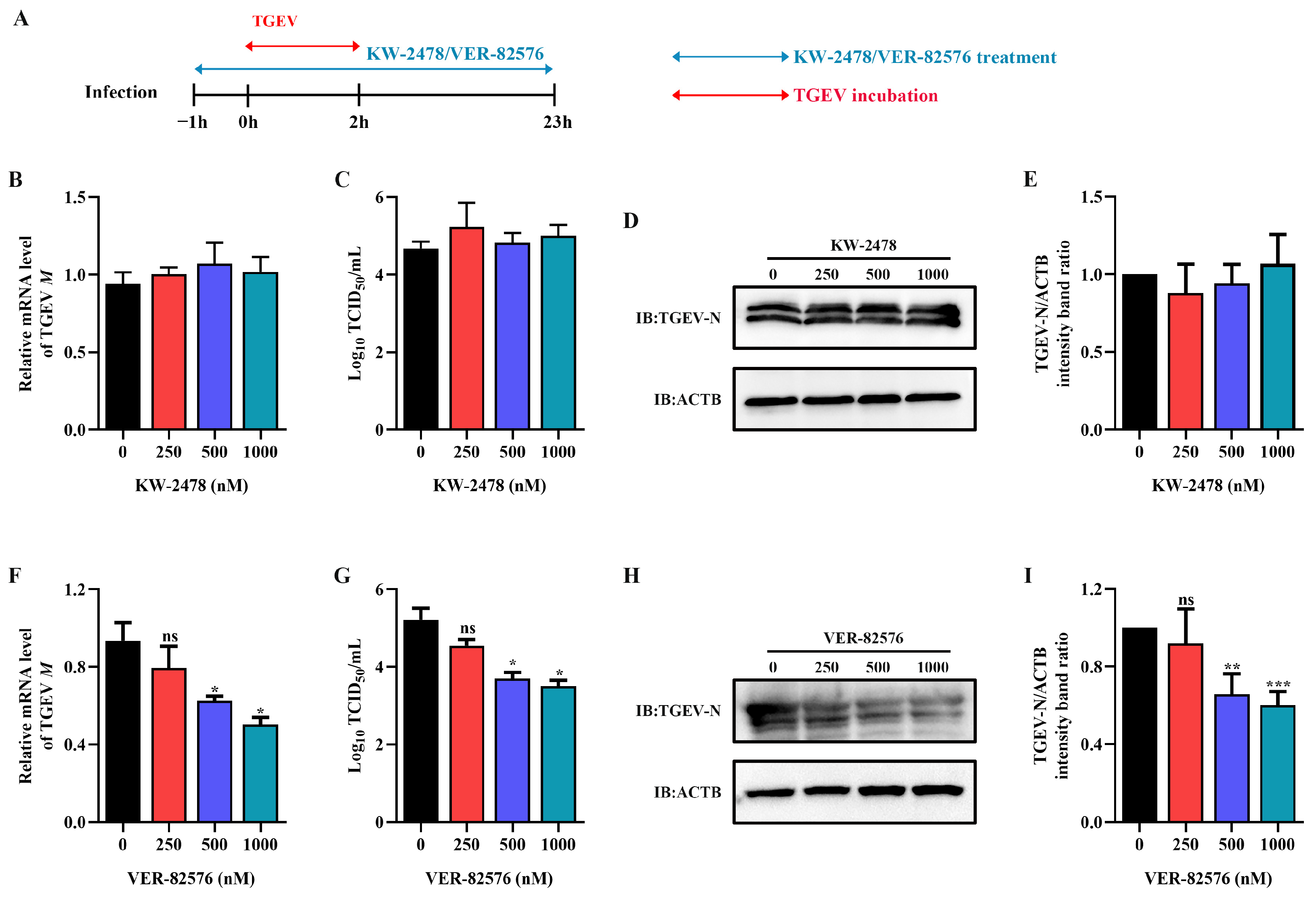
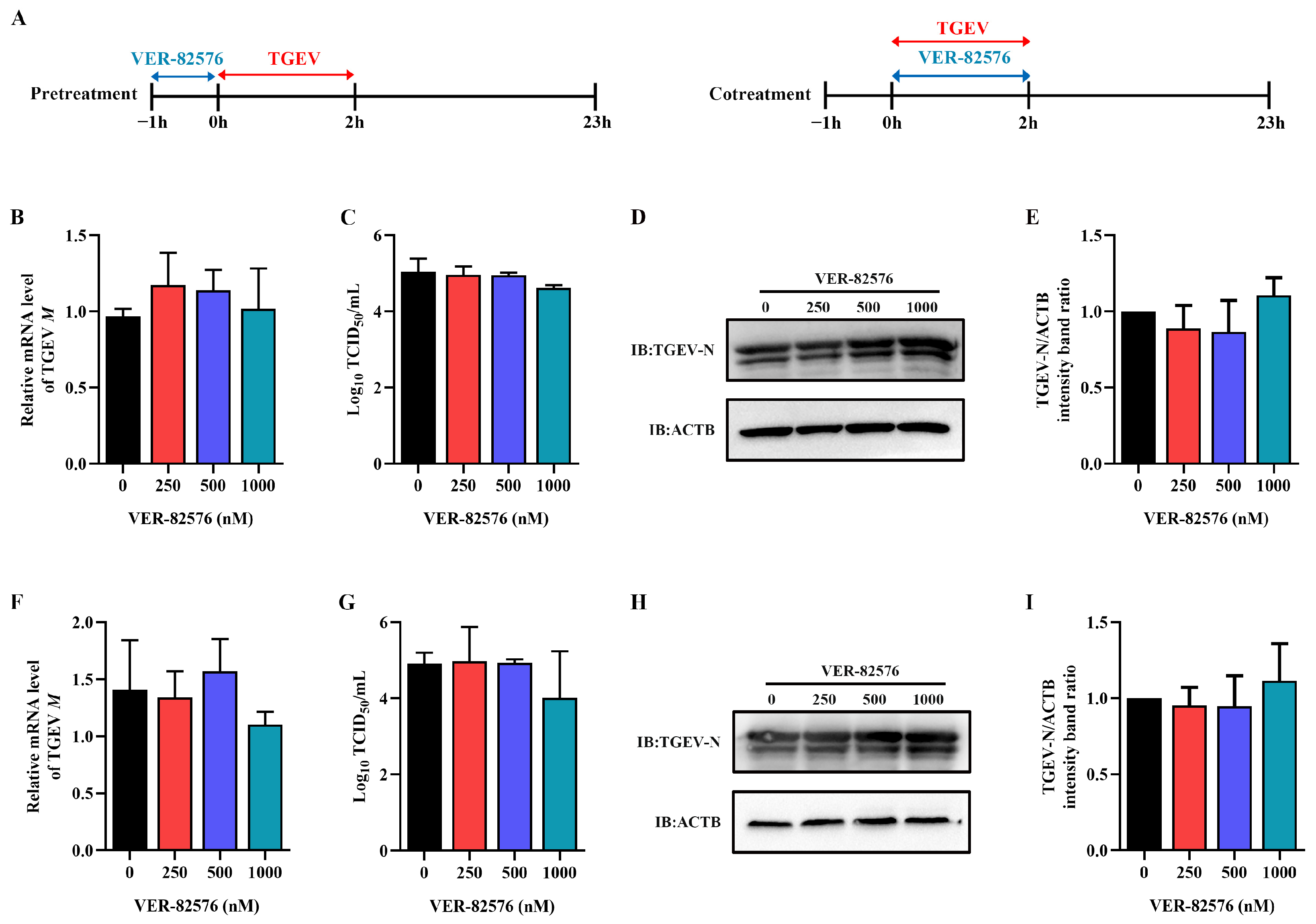
2.3. HSP90 Inhibitor VER-82576 Could Inhibit TGEV Infection
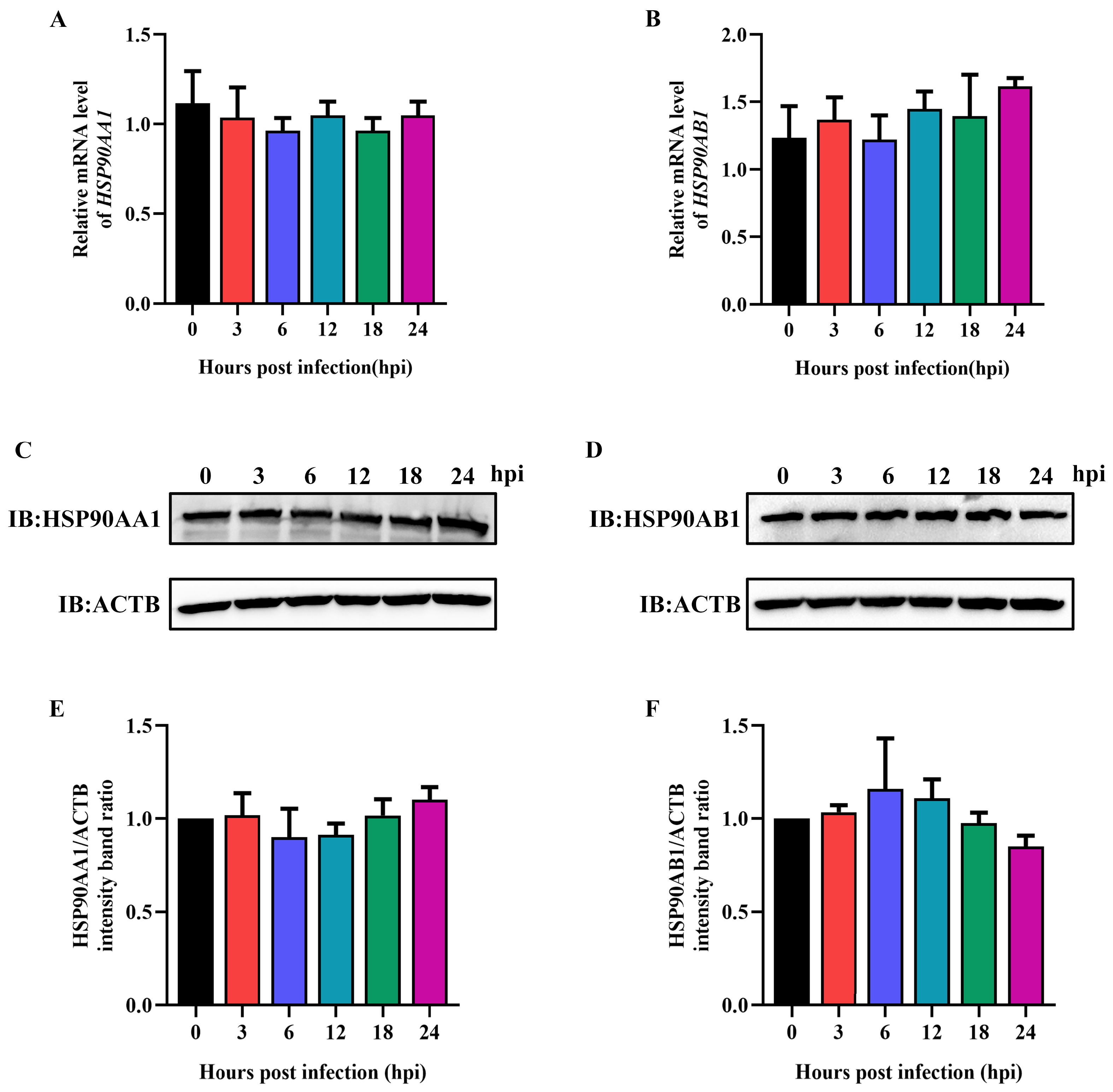
2.4. TGEV Infection Had No Influence on HSP90 Expression
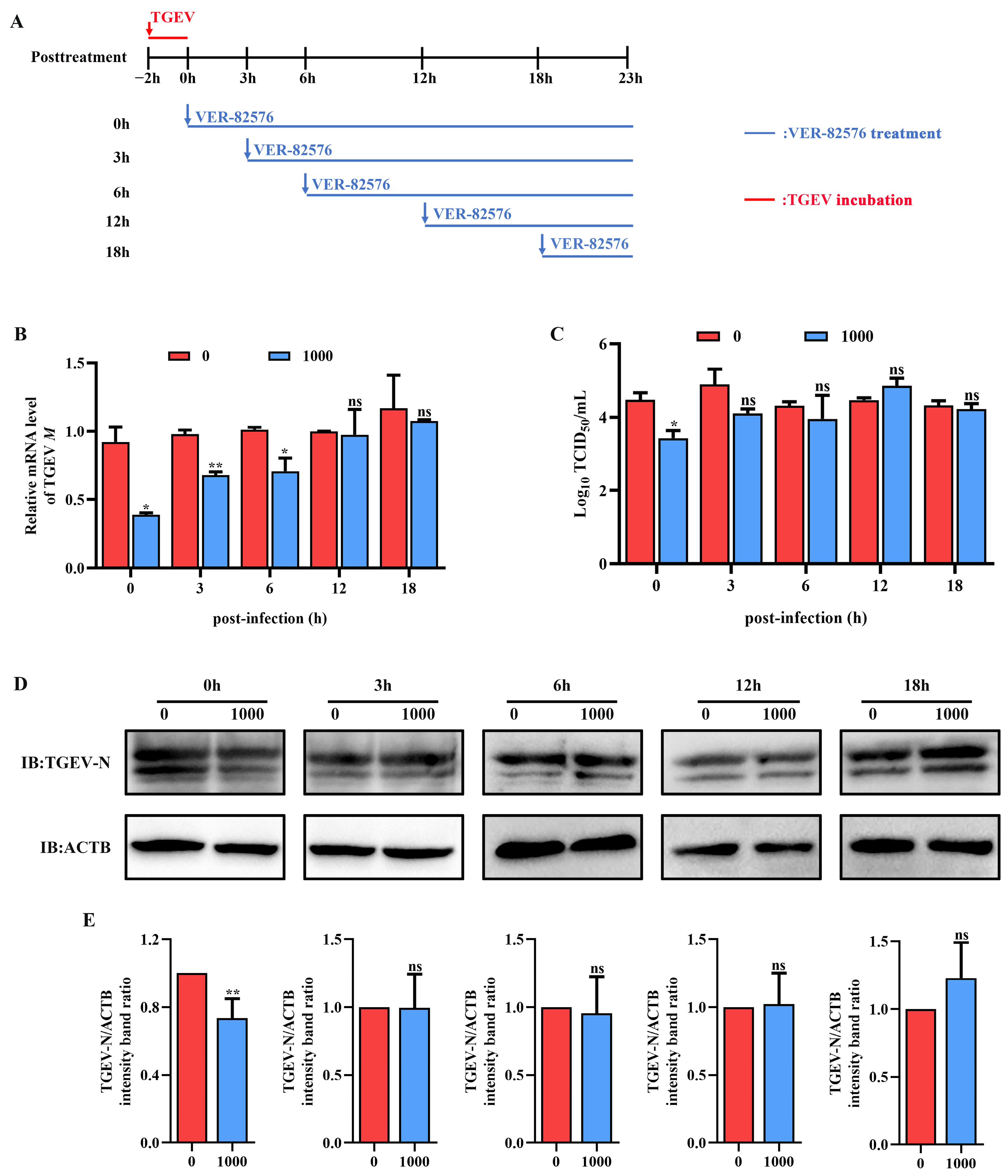
2.5. VER-82576 Inhibits TGEV Infection at the Early Stage of Replication
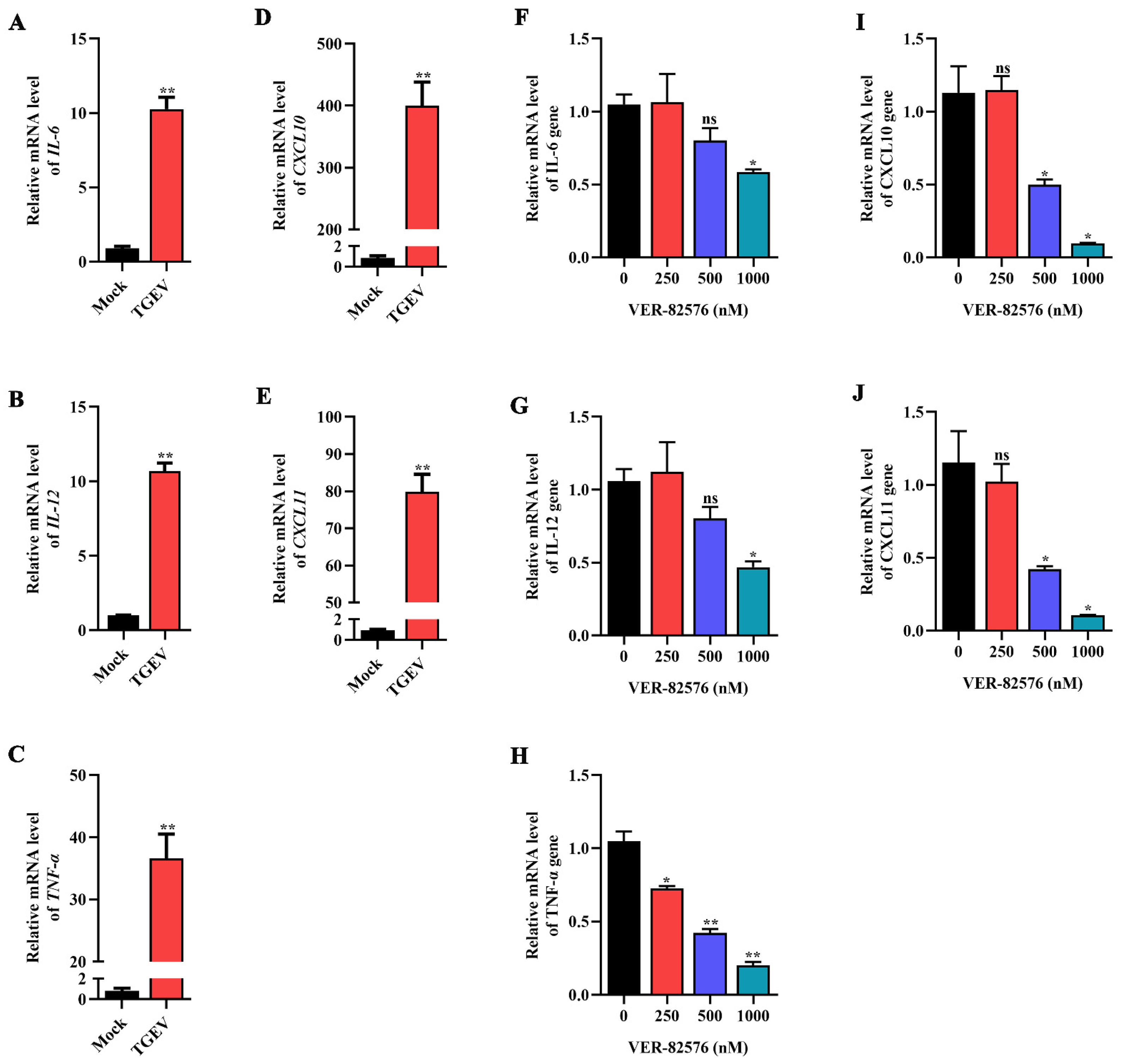
2.6. Effect of VER-82576 on TGEV-Induced Proinflammatory Cytokine Production
3. Discussion
4. Materials and Methods
4.1. Cells, Virus, and Inhibitors
4.2. Viral Replication Kinetics of TGEV in ST Cells
4.3. Effect of the HSP90 on TGEV Infection
4.4. Cell Viability Assay
4.5. Effect of VER-82576 on Different Life Cycles of TGEV Infection
4.6. Effect of VER-82576 on TGEV-Induced Proinflammatory Cytokine Production
4.7. qRT-PCR
4.8. Western Blot
4.9. TCID50
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, S.; Wu, H.; Chen, Z. Evolution of transmissible gastroenteritis virus (TGEV): A codon usage perspective. Int. J. Mol. Sci. 2020, 21, 7898. [Google Scholar] [CrossRef]
- Valkó, A.; Bálint, Á.; Bozsa, Á.; Cságola, A. Prevalence of antibodies against transmissible gastroenteritis virus (TGEV) in Hungary. Vet. Anim. Sci. 2019, 7, 100042. [Google Scholar] [CrossRef]
- Guo, R.; Fan, B.; Chang, X.; Zhou, J.; Zhao, Y.; Shi, D.; Yu, Z.; He, K.; Li, B. Characterization and evaluation of the pathogenicity of a natural recombinant transmissible gastroenteritis virus in China. Virology 2020, 545, 24–32. [Google Scholar] [CrossRef]
- Wang, X.; Fang, L.; Liu, S.; Ke, W.; Wang, D.; Peng, G.; Xiao, S. Susceptibility of porcine IPI-2I intestinal epithelial cells to infection with swine enteric coronaviruses. Vet. Microbiol. 2019, 233, 21–27. [Google Scholar] [CrossRef]
- Du, J.; Chen, D.; Yu, B.; He, J.; Yu, J.; Mao, X.; Luo, Y.; Zheng, P.; Luo, J. L-Leucine promotes STAT1 and ISGs expression in TGEV-infected IPEC-J2 cells via mTOR activation. Front. Immunol. 2021, 12, 656573. [Google Scholar] [CrossRef]
- Doyle, L.P.; Hutchings, L.M. A transmissible gastroenteritis in pigs. J. Am. Vet. Med. Assoc. 1946, 108, 257–259. [Google Scholar]
- Yuan, D.; Yan, Z.; Li, M.; Wang, Y.; Su, M.; Sun, D. Isolation and characterization of a porcine transmissible gastroenteritis coronavirus in northeast China. Front. Vet. Sci. 2021, 8, 611721. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, Y.; Cao, Y. The roles of apoptosis in swine response to viral infection and pathogenesis of swine enteropathogenic coronaviruses. Front. Vet. Sci. 2020, 7, 572425. [Google Scholar] [CrossRef]
- Turlewicz-Podbielska, H.; Pomorska-Mol, M. Porcine Coronaviruses: Overview of the State of the Art. Virol. Sin. 2021, 36, 833–851. [Google Scholar] [CrossRef]
- Zheng, L.; Zhao, F.; Ru, J.; Liu, L.; Wang, Z.; Wang, N.; Shu, X.; Wei, Z.; Guo, H. Evaluation of the Effect of Inactivated Transmissible Gastroenteritis Virus Vaccine with Nano Silicon on the Phenotype and Function of Porcine Dendritic Cells. Viruses 2021, 13, 2158. [Google Scholar] [CrossRef]
- Wißing, M.H.; Brüggemann, Y.; Steinmann, E.; Todt, D. Virus-host cell interplay during hepatitis E virus infection. Trends Microbiol. 2021, 29, 309–319. [Google Scholar] [CrossRef]
- Zhang, S.; Hu, W.; Yuan, L.; Yang, Q. Transferrin receptor 1 is a supplementary receptor that assists transmissible gastroenteritis virus entry into porcine intestinal epithelium. Cell Commun. Signal. CCS 2018, 16, 69. [Google Scholar] [CrossRef]
- Sun, L.; Zhao, C.; Fu, Z.; Fu, Y.; Su, Z.; Li, Y.; Zhou, Y.; Tan, Y.; Li, J.; Xiang, Y.; et al. Genome-scale CRISPR screen identifies TMEM41B as a multi-function host factor required for coronavirus replication. PLoS Pathog. 2021, 17, e1010113. [Google Scholar] [CrossRef]
- Song, Z.; Yang, Y.; Wang, L.; Wang, K.; Ran, L.; Xie, Y.; Huang, L.; Yang, Z.; Yuan, P.; Yu, Q. EIF4A2 interacts with the membrane protein of transmissible gastroenteritis coronavirus and plays a role in virus replication. Res. Vet. Sci. 2019, 123, 39–46. [Google Scholar] [CrossRef]
- Yuan, P.; Huang, S.; Yang, Z.; Xie, L.; Wang, K.; Yang, Y.; Ran, L.; Yu, Q.; Song, Z. UBXN1 interacts with the S1 protein of transmissible gastroenteritis coronavirus and plays a role in viral replication. Vet. Res. 2019, 50, 28. [Google Scholar] [CrossRef]
- Dong, W.; Xie, W.; Liu, Y.; Sui, B.; Zhang, H.; Liu, L.; Tan, Y.; Tong, X.; Fu, Z.F.; Yin, P.; et al. Receptor tyrosine kinase inhibitors block proliferation of TGEV mainly through p38 mitogen-activated protein kinase pathways. Antivir. Res. 2020, 173, 104651. [Google Scholar] [CrossRef]
- Yang, C.-W.; Hsu, H.-Y.; Chang, H.-Y.; Lee, Y.-Z.; Lee, S.-J. Natural cardenolides suppress coronaviral replication by downregulating JAK1 via a Na(+)/K(+)-ATPase independent proteolysis. Biochem. Pharmacol. 2020, 180, 114122. [Google Scholar] [CrossRef]
- Yang, C.-W.; Chang, H.-Y.; Lee, Y.-Z.; Hsu, H.-Y.; Lee, S.-J. The cardenolide ouabain suppresses coronaviral replication via augmenting a Na(+)/K(+)-ATPase-dependent PI3K_PDK1 axis signaling. Toxicol. Appl. Pharmacol. 2018, 356, 90–97. [Google Scholar] [CrossRef]
- Shan, L.; Fu, F.; Xue, M.; Zhu, X.; Li, L.; Feng, L.; Liu, P. Interferon gamma inhibits transmissible gastroenteritis virus infection mediated by an IRF1 signaling pathway. Arch. Virol. 2019, 164, 2659–2669. [Google Scholar] [CrossRef]
- Gao, D.M.; Yu, H.Y.; Zhou, W.; Xia, B.B.; Li, H.Z.; Wang, M.L.; Zhao, J. Inhibitory effects of recombinant porcine interferon-α on porcine transmissible gastroenteritis virus infections in TGEV-seronegative piglets. Vet. Microbiol. 2021, 252, 108930. [Google Scholar] [CrossRef]
- Zhao, J.; Yi, Y.X.; Yu, H.Y.; Wang, M.L.; Lai, P.F.; Gan, L.; Zhao, Y.; Fu, X.; Chen, J. Antiviral activity of recombinant porcine interferon-α against porcine transmissible gastroenteritis virus in PK-15 Cells. Kafkas Univ. Vet. Fak. Derg. 2017, 23, 603–611. [Google Scholar] [CrossRef]
- Shan, Q.; Ma, F.; Wei, J.; Li, H.; Ma, H.; Sun, P. Physiological functions of heat shock proteins. Curr. Protein Pept. Sci. 2020, 21, 751–760. [Google Scholar] [CrossRef]
- Geller, R.; Taguwa, S.; Frydman, J. Broad action of Hsp90 as a host chaperone required for viral replication. Biochim. et Biophys. Acta 2012, 1823, 698–706. [Google Scholar] [CrossRef]
- Zhang, W.; Jia, K.; Jia, P.; Xiang, Y.; Lu, X.; Liu, W.; Yi, M. Marine medaka heat shock protein 90ab1 is a receptor for red-spotted grouper nervous necrosis virus and promotes virus internalization through clathrin-mediated endocytosis. PLoS Pathog. 2020, 16, e1008668. [Google Scholar] [CrossRef]
- Li, F.; Jin, F.; Wang, Y.; Zheng, D.; Liu, J.; Zhang, Z.; Wang, R.; Dong, D.; Zheng, K.; Wang, Y. Hsp90 inhibitor AT-533 blocks HSV-1 nuclear egress and assembly. J. Biochem. 2018, 164, 397–406. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, R.; Li, F.; Wang, Y.; Zhang, Z.; Wang, Q.; Ren, Z.; Jin, F.; Kitazato, K.; Wang, Y. Heat-shock protein 90α is involved in maintaining the stability of VP16 and VP16-mediated transactivation of α genes from herpes simplex virus-1. Mol. Med. 2018, 24, 65. [Google Scholar] [CrossRef]
- Li, C.; Chu, H.; Liu, X.; Chiu, M.C.; Zhao, X.; Wang, D.; Wei, Y.; Hou, Y.; Shuai, H.; Cai, J.; et al. Human coronavirus dependency on host heat shock protein 90 reveals an antiviral target. Emerg. Microbes Infect. 2020, 9, 2663–2672. [Google Scholar] [CrossRef]
- Zhang, S.; Huang, W.; Ren, L.; Ju, X.; Gong, M.; Rao, J.; Sun, L.; Li, P.; Ding, Q.; Wang, J.; et al. Comparison of viral RNA-host protein interactomes across pathogenic RNA viruses informs rapid antiviral drug discovery for SARS-CoV-2. Cell Res. 2022, 32, 9–23. [Google Scholar] [CrossRef]
- Wyler, E.; Mösbauer, K.; Franke, V.; Diag, A.; Gottula, L.T.; Arsiè, R.; Klironomos, F.; Koppstein, D.; Hönzke, K.; Ayoub, S.; et al. Transcriptomic profiling of SARS-CoV-2 infected human cell lines identifies HSP90 as target for COVID-19 therapy. iScience 2021, 24, 102151. [Google Scholar] [CrossRef]
- Qu, H.; Wen, Y.; Hu, J.; Xiao, D.; Li, S.; Zhang, L.; Liao, Y.; Chen, R.; Zhao, Y.; Wen, Y.; et al. Study of the inhibitory effect of STAT1 on PDCoV infection. Vet. Microbiol. 2022, 266, 109333. [Google Scholar] [CrossRef]
- Zhao, Y.; Xiao, D.; Zhang, L.; Song, D.; Chen, R.; Li, S.; Liao, Y.; Wen, Y.; Liu, W.; Yu, E.; et al. HSP90 inhibitors 17-AAG and VER-82576 inhibit porcine deltacoronavirus replication in vitro. Vet. Microbiol. 2022, 265, 109316. [Google Scholar] [CrossRef]
- Delmas, B.; Gelfi, J.; Sjöström, H.; Noren, O.; Laude, H. Further characterization of aminopeptidase-N as a receptor for coronaviruses. Adv. Exp. Med. Biol. 1993, 342, 293–298. [Google Scholar] [CrossRef]
- Ma, R.; Zhang, Y.; Liu, H.; Ning, P. Proteome profile of swine testicular cells infected with porcine transmissible gastroenteritis coronavirus. PLoS ONE 2014, 9, e110647. [Google Scholar] [CrossRef]
- Zhai, X.; Wang, S.; Zhu, M.; He, W.; Pan, Z.; Su, S. Antiviral effect of lithium chloride and diammonium glycyrrhizinate on porcine deltacoronavirus in vitro. Pathogens 2019, 8, 144. [Google Scholar] [CrossRef]
- Martinez, J.P.; Sasse, F.; Brönstrup, M.; Diez, J.; Meyerhans, A. Antiviral drug discovery: Broad-spectrum drugs from nature. Nat. Prod. Rep. 2015, 32, 29–48. [Google Scholar] [CrossRef]
- Srisutthisamphan, K.; Jirakanwisal, K.; Ramphan, S.; Tongluan, N.; Kuadkitkan, A.; Smith, D.R. Hsp90 interacts with multiple dengue virus 2 proteins. Sci. Rep. 2018, 8, 4308. [Google Scholar] [CrossRef]
- Lee, K.H.; Lee, J.H.; Han, S.W.; Im, S.A.; Kim, T.Y.; Oh, D.Y.; Bang, Y.J. Antitumor activity of NVP-AUY922, a novel heat shock protein 90 inhibitor, in human gastric cancer cells is mediated through proteasomal degradation of client proteins. Cancer Sci. 2011, 102, 1388–1395. [Google Scholar] [CrossRef]
- Massey, A.J.; Schoepfer, J.; Brough, P.A.; Brueggen, J.; Chène, P.; Drysdale, M.J.; Pfaar, U.; Radimerski, T.; Ruetz, S.; Schweitzer, A.; et al. Preclinical antitumor activity of the orally available heat shock protein 90 inhibitor NVP-BEP800. Mol. Cancer Ther. 2010, 9, 906–919. [Google Scholar] [CrossRef]
- Nakashima, T.; Ishii, T.; Tagaya, H.; Seike, T.; Nakagawa, H.; Kanda, Y.; Akinaga, S.; Soga, S.; Shiotsu, Y. New molecular and biological mechanism of antitumor activities of KW-2478, a novel nonansamycin heat shock protein 90 inhibitor, in multiple myeloma cells. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 2792–2802. [Google Scholar] [CrossRef]
- Mshaik, R.; Simonet, J.; Georgievski, A.; Jamal, L.; Bechoua, S.; Ballerini, P.; Bellaye, P.S.; Mlamla, Z.; Pais de Barros, J.P.; Geissler, A.; et al. HSP90 inhibitor NVP-BEP800 affects stability of SRC kinases and growth of T-cell and B-cell acute lymphoblastic leukemias. Blood Cancer J. 2021, 11, 61. [Google Scholar] [CrossRef]
- Sakata, M.; Katoh, H.; Otsuki, N.; Okamoto, K.; Mori, Y. Heat shock protein 90 ensures the integrity of rubella virus p150 protein and supports viral replication. J. Virol. 2019, 93, e01142-19. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.Y.; Kuo, R.L.; Ma, W.C.; Huang, H.I.; Yu, J.S.; Yen, S.M.; Huang, C.R.; Shih, S.R. Heat shock protein-90-beta facilitates enterovirus 71 viral particles assembly. Virology 2013, 443, 236–247. [Google Scholar] [CrossRef]
- Kumar, P.; Gaur, P.; Kumari, R.; Lal, S.K. Influenza A virus neuraminidase protein interacts with Hsp90, to stabilize itself and enhance cell survival. J. Cell. Biochem. 2019, 120, 6449–6458. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Yang, Y.; Wang, J.; Jing, Y.; Yang, Q. Impact of TGEV infection on the pig small intestine. Virol. J. 2018, 15, 102. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Gao, Z.; Cao, R.; Yang, K.; Cui, Y.; Li, S.; Meng, X.; He, Q.; Li, Z. Transmissible gastroenteritis virus infection up-regulates FcRn expression via nucleocapsid protein and secretion of TGF-β in porcine intestinal epithelial cells. Front. Immunol. 2019, 10, 3085. [Google Scholar] [CrossRef]
- Zhao, S.; Gao, Q.; Lin, J.; Yan, M.; Yu, Q.; Yang, Q. Differential response of porcine immature monocyte-derived dendritic cells to virulent and inactivated transmissible gastroenteritis virus. Res. Vet. Sci. 2014, 97, 623–630. [Google Scholar] [CrossRef]
- Xia, L.; Dai, L.; Zhu, L.; Hu, W.; Yang, Q. Proteomic analysis of IPEC-J2 cells in response to coinfection by porcine transmissible gastroenteritis virus and enterotoxigenic escherichia coli K88. Proteomics. Clin. Appl. 2017, 11, 1600137. [Google Scholar] [CrossRef]
- Akhtar, M.; Guo, S.; Guo, Y.F.; Zahoor, A.; Shaukat, A.; Chen, Y.; Umar, T.; Deng, P.G.; Guo, M. Upregulated-gene expression of pro-inflammatory cytokines (TNF-α, IL-1β and IL-6) via TLRs following NF-κB and MAPKs in bovine mastitis. Acta Trop. 2020, 207, 105458. [Google Scholar] [CrossRef]
- Liu, D.; Wu, A.; Cui, L.; Hao, R.; Wang, Y.; He, J.; Guo, D. Hepatitis B virus polymerase suppresses NF-κB signaling by inhibiting the activity of IKKs via interaction with Hsp90β. PLoS ONE 2014, 9, e91658. [Google Scholar] [CrossRef]
- Li, F.; Song, X.; Su, G.; Wang, Y.; Wang, Z.; Qing, S.; Jia, J.; Wang, Y.; Huang, L.; Zheng, K.; et al. AT-533, a Hsp90 inhibitor, attenuates HSV-1-induced inflammation. Biochem. Pharmacol. 2019, 166, 82–92. [Google Scholar] [CrossRef]
- Liu, J.; You, J.; Zhang, X.; Ma, C.; Dong, M.; Chen, L.; Jiang, P.; Yun, S. The Hsp90 inhibitor 17-DMAG decreases infection of porcine circovirus type 2 in mice. Microb. Pathog. 2017, 109, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sgRNA | Sequences (5′-3′) |
|---|---|
| HSP90AA1 | F: CACCGCGACGAGATGGTTTCCCTCA |
| R: AAACTGAGGGAAACCATCTCGTCGC | |
| HSP90AB1 | F: CACCGAGGTCAAAAGGAGCCCGACG |
| R: AAACCGTCGGGCTCCTTTTGACCTC |
| Genes | Sequences (5′-3′) | Size (bp) |
|---|---|---|
| TGEV M | F: GTGGAGAACGCTATTGTGCTAT | 258 |
| R: AAATCGTAAGAGCCAAAACAAC | ||
| IL-6 | F: GATGCTTCCAATCTGGGTTC | 218 |
| R: ATTTGTGGTGGGGTTAGGG | ||
| IL12 | F: AACCACCTGGACCATCTCA | 211 |
| R: CCTCCACTGTGCTGGTTTT | ||
| TNF-α | F: TCCTCACTCACACCATCAGC | 222 |
| R: GCCCAGATTCAGCAAAGTCC | ||
| CXCL10 | F: AATCTACCTCTGCCATCATCTC | 373 |
| R: AGTAGAAGCCCACGGAGTAAAG | ||
| CXCL11 | F: AACTATTCAAGGCTTCCCCAT | 201 |
| R: ACATTTGCTTGCTTTGATTTG | ||
| ACTB | F: CTTCCTGGGCATGGAGTCC | 201 |
| R: GGCGCGATGATCTTGATCTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, D.; Zhao, Y.; Sun, Y.; Liang, Y.; Chen, R.; Wen, Y.; Wu, R.; Zhao, Q.; Du, S.; Yan, Q.; et al. HSP90AB1 Is a Host Factor Required for Transmissible Gastroenteritis Virus Infection. Int. J. Mol. Sci. 2023, 24, 15971. https://doi.org/10.3390/ijms242115971
Song D, Zhao Y, Sun Y, Liang Y, Chen R, Wen Y, Wu R, Zhao Q, Du S, Yan Q, et al. HSP90AB1 Is a Host Factor Required for Transmissible Gastroenteritis Virus Infection. International Journal of Molecular Sciences. 2023; 24(21):15971. https://doi.org/10.3390/ijms242115971
Chicago/Turabian StyleSong, Daili, Yujia Zhao, Ying Sun, Yixiao Liang, Rui Chen, Yiping Wen, Rui Wu, Qin Zhao, Senyan Du, Qigui Yan, and et al. 2023. "HSP90AB1 Is a Host Factor Required for Transmissible Gastroenteritis Virus Infection" International Journal of Molecular Sciences 24, no. 21: 15971. https://doi.org/10.3390/ijms242115971
APA StyleSong, D., Zhao, Y., Sun, Y., Liang, Y., Chen, R., Wen, Y., Wu, R., Zhao, Q., Du, S., Yan, Q., Han, X., Cao, S., & Huang, X. (2023). HSP90AB1 Is a Host Factor Required for Transmissible Gastroenteritis Virus Infection. International Journal of Molecular Sciences, 24(21), 15971. https://doi.org/10.3390/ijms242115971