

Predictive Potential of RNA Polymerase B (II) Subunit 1 (RPB1) Cytoplasmic Aggregation for Neoadjuvant Chemotherapy Failure

 , , ,
, , ,  ,
,  and add
Show full author list
and add
Show full author list
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
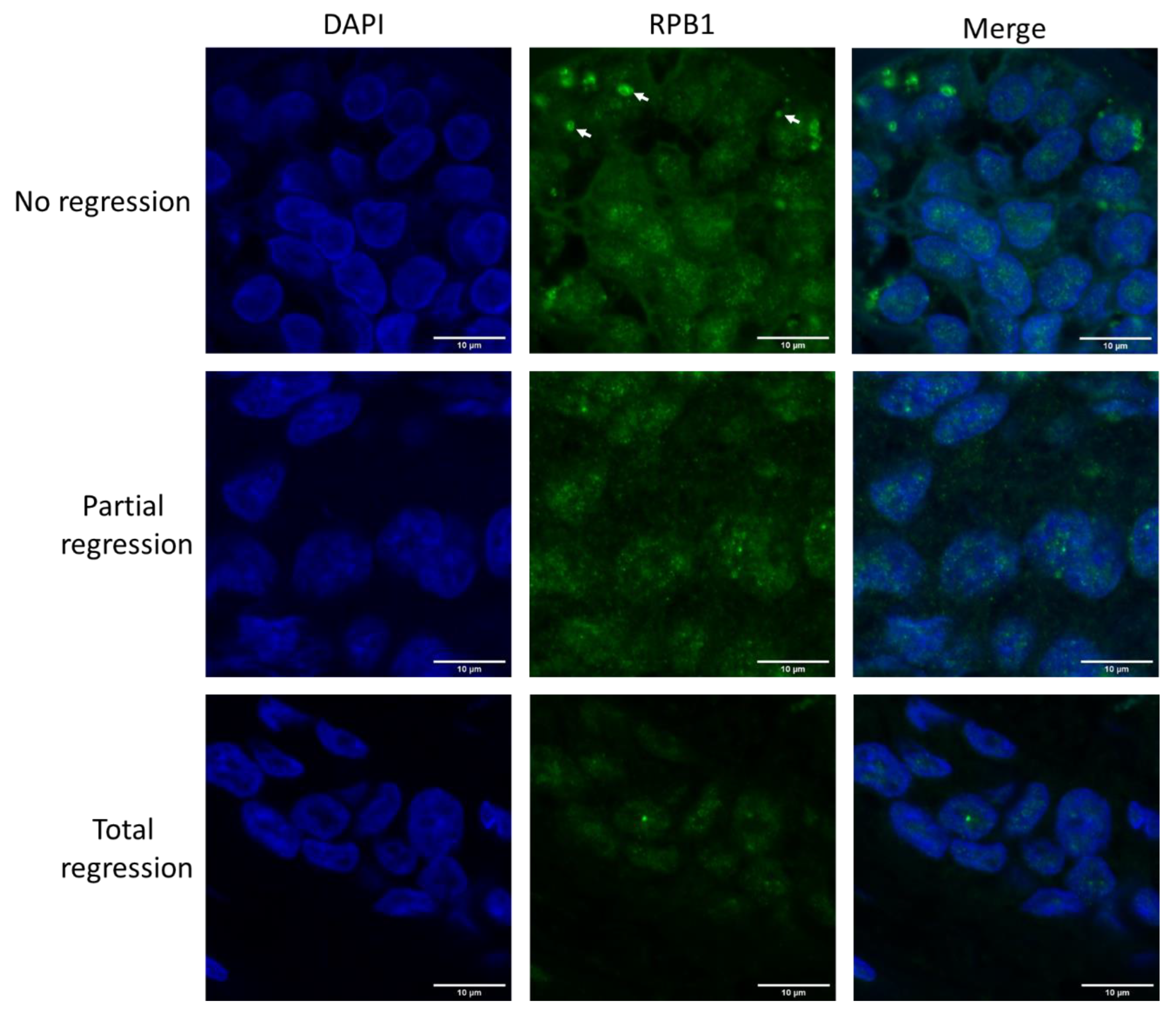
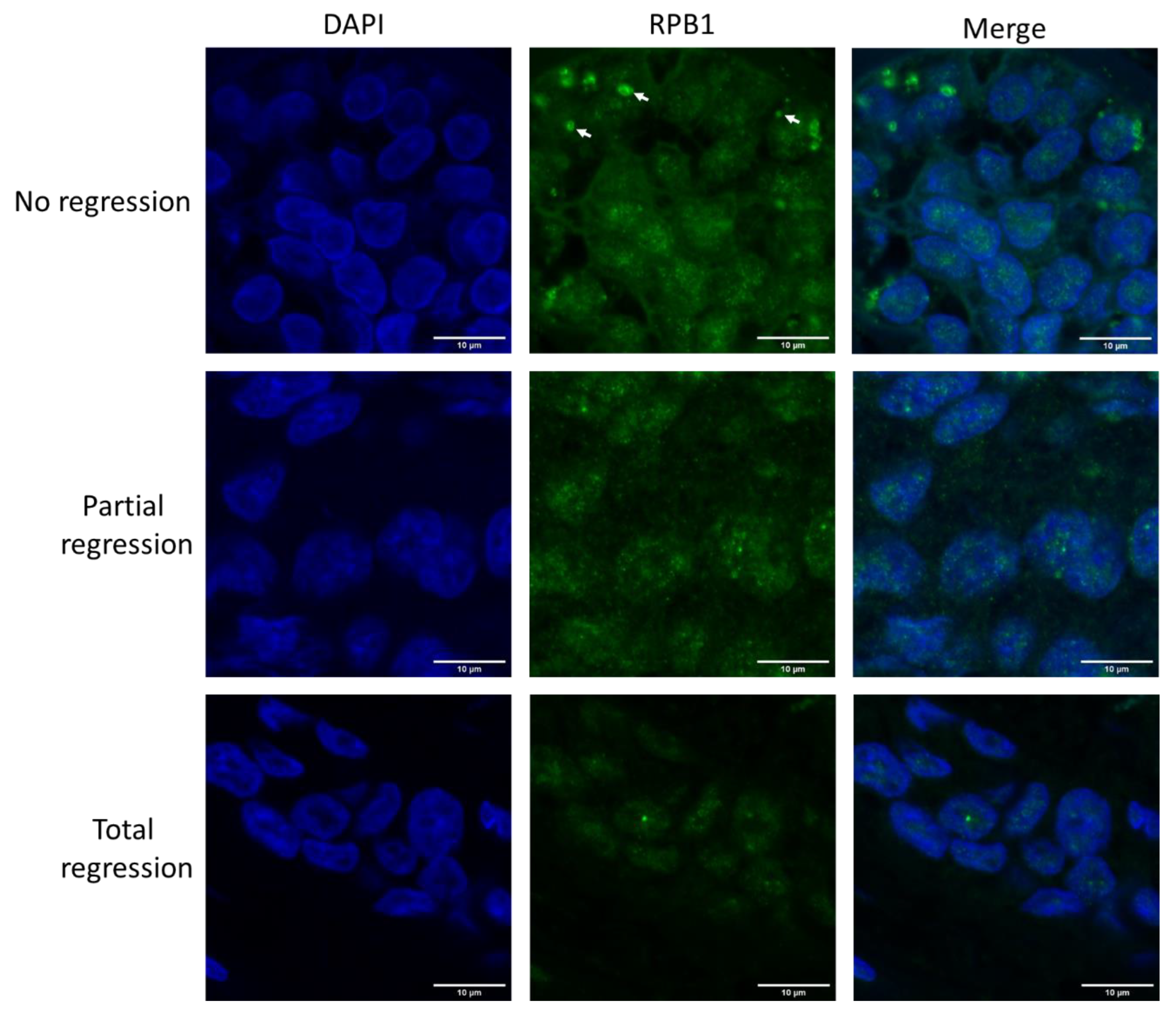
2.1. Large RPB1 Cytoplasmic Foci Are Apparent in Non-Regressive Invasive Carcinoma of NST Cells
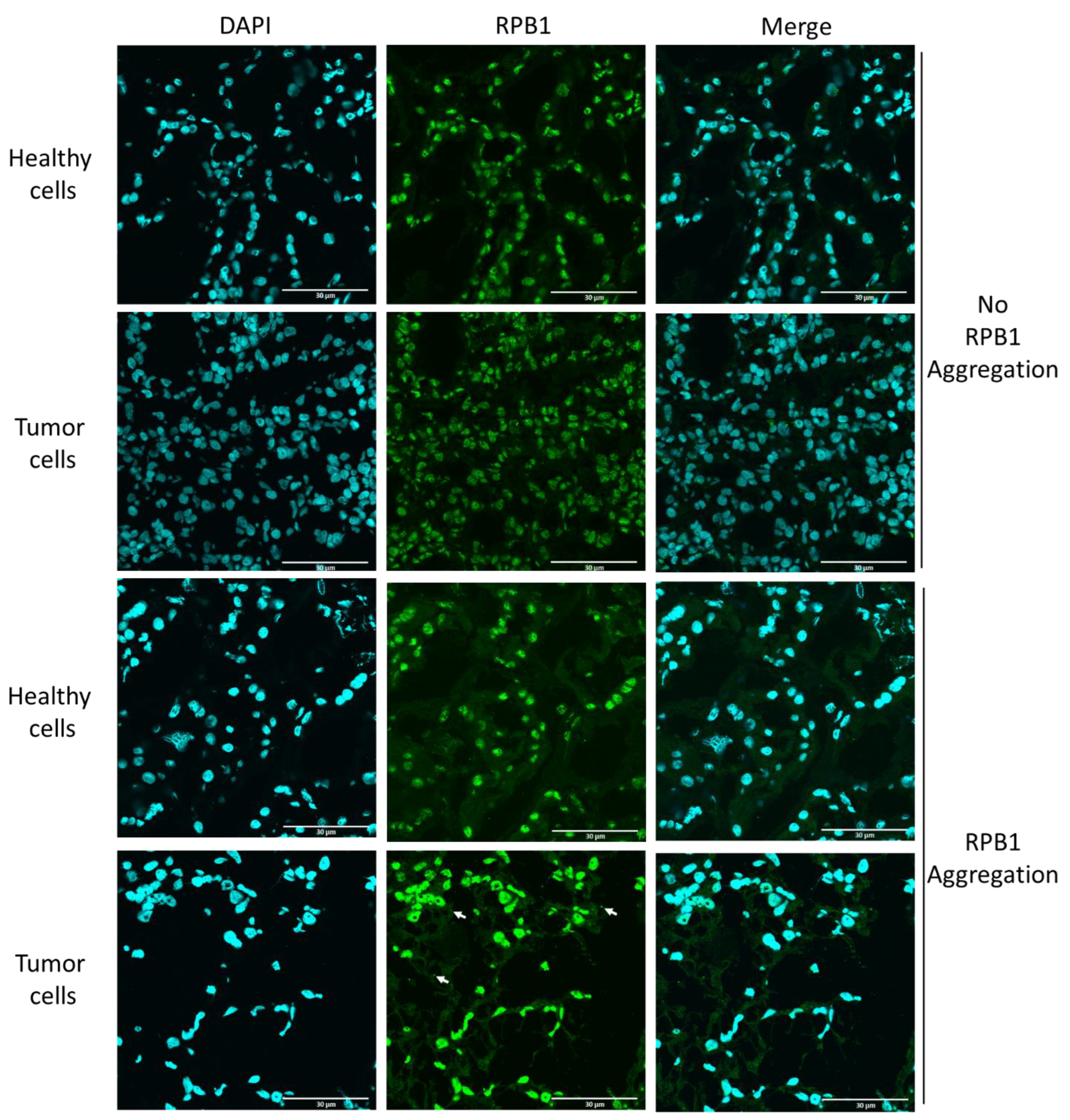
2.2. Cytoplasmic RPB1 Foci Are Apparent Sporadically in Renal-Cell Carcinoma Cells, Although Not in the Cells of Surrounding Tissues
3. Discussion
4. Materials and Methods
4.1. Cohort Selection
4.2. Preparations of Normal and Tumorous Tissues
4.3. Preparations of Formalin-Fixed Paraffin-Embedded (FFPE) Biopsy Samples
4.4. Immunostaining of Frozen Tissues
4.5. Immunostaining of FFPE Samples
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| P-body | Processing Body |
| RPB1 | RNA Polymerase B (II) subunit 1 |
| HSP90 | Heat Shock Protein 90 |
| NOT1 | Negative on TATA-less 1 |
| CCR4 | Carbon Catabolite Repression 4 |
| NST | No Special Type |
| IPA | Isopropanol |
| BSA | Bovine Serum Albumin |
| GAM | Goat Anti Mouse |
| GAR | Goat Anti Rabbit |
| PBS(T) | Phosphate Buffered Saline with Tween 20 |
| ACC | Accuracy |
| ERR | Error Rate |
| MCC | Matthews Correlation Coefficient |
| RPL5 | Ribosomal Protein L5 (L:large subunit) |
| RPL10 | Ribosomal Protein L10 (L:large subunit) |
| RP | Ribosomal Protein |
| RiBi | Ribosome Biogenesis |
References
- Masood, S. Neoadjuvant chemotherapy in breast cancers. Women’s Health 2016, 12, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Robert, J. Clinical pharmacokinetics of epirubicin. Clin. Pharmacokinet. 1994, 26, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.; Kedersha, N.; Ivanov, P. Stress granules, P-bodies and cancer. Biochim. Biophys. Acta-Gene Regul. Mech. 2015, 1849, 861–870. [Google Scholar] [CrossRef]
- Panasenko, O.O.; Somasekharan, S.P.; Villanyi, Z.; Zagatti, M.; Bezrukov, F.; Rashpa, R.; Cornut, J.; Iqbal, J.; Longis, M.; Carl, S.H.; et al. Co-translational assembly of proteasome subunits in NOT1-containing assemblysomes. Nat. Struct. Mol. Biol. 2019, 26, 110–120. [Google Scholar] [CrossRef]
- Nemeth-Szatmari, O.; Nagy-Miko, B.; Gyorkei, A.; Varga, D.; Kovacs, B.B.H.; Igaz, N.; Bognar, B.; Razga, Z.; Nagy, G.; Zsindely, N.; et al. Phase separated ribosome-nascent chain complexes in genotoxic stress response. RNA 2023, 29, 1557–1574. [Google Scholar] [CrossRef] [PubMed]
- Collart, M.A.; Panasenko, O.O. The Ccr4-not complex. Gene 2012, 492, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Collart, M.A. The Ccr4-Not complex is a key regulator of eukaryotic gene expression. WIREs RNA 2016, 7, 438–454. [Google Scholar] [CrossRef] [PubMed]
- Collart, M.A.; Weiss, B. Ribosome pausing, a dangerous necessity for co-translational events. Nucleic Acids Res. 2019, 48, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Villanyi, Z.; Ribaud, V.; Kassem, S.; Panasenko, O.O.; Pahi, Z.; Gupta, I.; Steinmetz, L.; Boros, I.; Collart, M.A. The Not5 subunit of the ccr4-not complex connects transcription and translation. PLoS Genet. 2014, 10, e1004569. [Google Scholar] [CrossRef] [PubMed]
- Cramer, P. Multisubunit RNA polymerases. Curr. Opin. Struct. Biol. 2002, 12, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Czeko, E.; Seizl, M.; Augsberger, C.; Mielke, T.; Cramer, P. Iwr1 directs RNA polymerase II nuclear import. Mol. Cell 2011, 42, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.E.; Panasenko, O.O.; Villanyi, Z.; Zagatti, M.; Weiss, B.; Pagliazzo, L.; Huch, S.; Polte, C.; Zahoran, S.; Hughes, C.S.; et al. Not4 and Not5 modulate translation elongation by Rps7A ubiquitination, Rli1 moonlighting, and condensates that exclude eIF5A. Cell Rep. 2021, 36, 109633. [Google Scholar] [CrossRef] [PubMed]
- Mosley, A.L.; Pattenden, S.G.; Carey, M.; Venkatesh, S.; Gilmore, J.M.; Florens, L.; Workman, J.L.; Washburn, M.P. Rtr1 is a CTD phosphatase that regulates RNA polymerase II during the transition from serine 5 to serine 2 phosphorylation. Mol. Cell 2009, 34, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Tharwat, A. Classification assessment methods. Appl. Comput. Inform. 2021, 17, 168–192. [Google Scholar] [CrossRef]
- Matthews, B. Comparison of the predicted and observed secondary structure of T4 phage lysozyme. Biochim. Biophys Acta-Protein Struct. 1975, 405, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Ördög, N.; Borsos, B.N.; Majoros, H.; Ujfaludi, Z.; Pankotai-Bodó, G.; Bankó, S.; Sükösd, F.; Kuthi, L.; Pankotai, T. The clinical significance of epigenetic and RNAPII variabilities occurring in clear cell renal cell carcinoma as a potential prognostic marker. Transl. Oncol. 2022, 20, 101420. [Google Scholar] [CrossRef]
- De Keersmaecker, K.; Atak, Z.K.; Li, N.; Vicente, C.; Patchett, S.; Girardi, T.; Gianfelici, V.; Geerdens, E.; Clappier, E.; Porcu, M.; et al. Exome sequencing identifies mutation in CNOT3 and ribosomal genes RPL5 and RPL10 in T-cell acute lymphoblastic leukemia. Nat. Genet. 2013, 45, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Ebright, R.Y.; Lee, S.; Wittner, B.S.; Niederhoffer, K.L.; Nicholson, B.T.; Bardia, A.; Truesdell, S.; Wiley, D.F.; Wesley, B.; Li, S.; et al. Deregulation of ribosomal protein expression and translation promotes breast cancer metastasis. Science 2020, 367, 1468–1473. [Google Scholar] [CrossRef] [PubMed]
- Gupta, I.; Villanyi, Z.; Kassem, S.; Hughes, C.; Panasenko, O.O.; Steinmetz, L.M.; Collart, M.A. Translational Capacity of a Cell Is Determined during Transcription Elongation via the Ccr4-Not Complex. Cell Rep. 2016, 15, 1782–1794. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.; Weiss, B.; Panasenko, O.O.; Huch, S.; Villanyi, Z.; Albert, B.; Dilg, D.; Zagatti, M.; Schaughency, P.; Liao, S.E.; et al. Not1 and Not4 inversely determine mRNA solubility that sets the dynamics of co-translational events. bioRxiv 2022. [Google Scholar] [CrossRef]
- Available online: https://www.proteinatlas.org/ENSG00000181222-POLR2A/subcellular#human (accessed on 9 October 2023).
- Sadeghipour, A.; Babaheidarian, P. Making Formalin-Fixed, Paraffin Embedded Blocks. In Biobanking; Yong, W., Ed.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2019; Volume 1897. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagy-Mikó, B.; Németh-Szatmári, O.; Faragó-Mészáros, R.; Csókási, A.; Bognár, B.; Ördög, N.; Borsos, B.N.; Majoros, H.; Ujfaludi, Z.; Oláh-Németh, O.; et al. Predictive Potential of RNA Polymerase B (II) Subunit 1 (RPB1) Cytoplasmic Aggregation for Neoadjuvant Chemotherapy Failure. Int. J. Mol. Sci. 2023, 24, 15869. https://doi.org/10.3390/ijms242115869
Nagy-Mikó B, Németh-Szatmári O, Faragó-Mészáros R, Csókási A, Bognár B, Ördög N, Borsos BN, Majoros H, Ujfaludi Z, Oláh-Németh O, et al. Predictive Potential of RNA Polymerase B (II) Subunit 1 (RPB1) Cytoplasmic Aggregation for Neoadjuvant Chemotherapy Failure. International Journal of Molecular Sciences. 2023; 24(21):15869. https://doi.org/10.3390/ijms242115869
Chicago/Turabian StyleNagy-Mikó, Bence, Orsolya Németh-Szatmári, Réka Faragó-Mészáros, Aliz Csókási, Bence Bognár, Nóra Ördög, Barbara N. Borsos, Hajnalka Majoros, Zsuzsanna Ujfaludi, Orsolya Oláh-Németh, and et al. 2023. "Predictive Potential of RNA Polymerase B (II) Subunit 1 (RPB1) Cytoplasmic Aggregation for Neoadjuvant Chemotherapy Failure" International Journal of Molecular Sciences 24, no. 21: 15869. https://doi.org/10.3390/ijms242115869
APA StyleNagy-Mikó, B., Németh-Szatmári, O., Faragó-Mészáros, R., Csókási, A., Bognár, B., Ördög, N., Borsos, B. N., Majoros, H., Ujfaludi, Z., Oláh-Németh, O., Nikolényi, A., Dobi, Á., Kószó, R., Sántha, D., Lázár, G., Simonka, Z., Paszt, A., Ormándi, K., Pankotai, T., ... Vörös, A. (2023). Predictive Potential of RNA Polymerase B (II) Subunit 1 (RPB1) Cytoplasmic Aggregation for Neoadjuvant Chemotherapy Failure. International Journal of Molecular Sciences, 24(21), 15869. https://doi.org/10.3390/ijms242115869