
Contribution of the LAC Genes in Fruit Quality Attributes of the Fruit-Bearing Plants: A Comprehensive Review

Abstract
:1. Introduction
2. Molecular Regulation of the LAC Gene Family Involved in Fruit Lignification
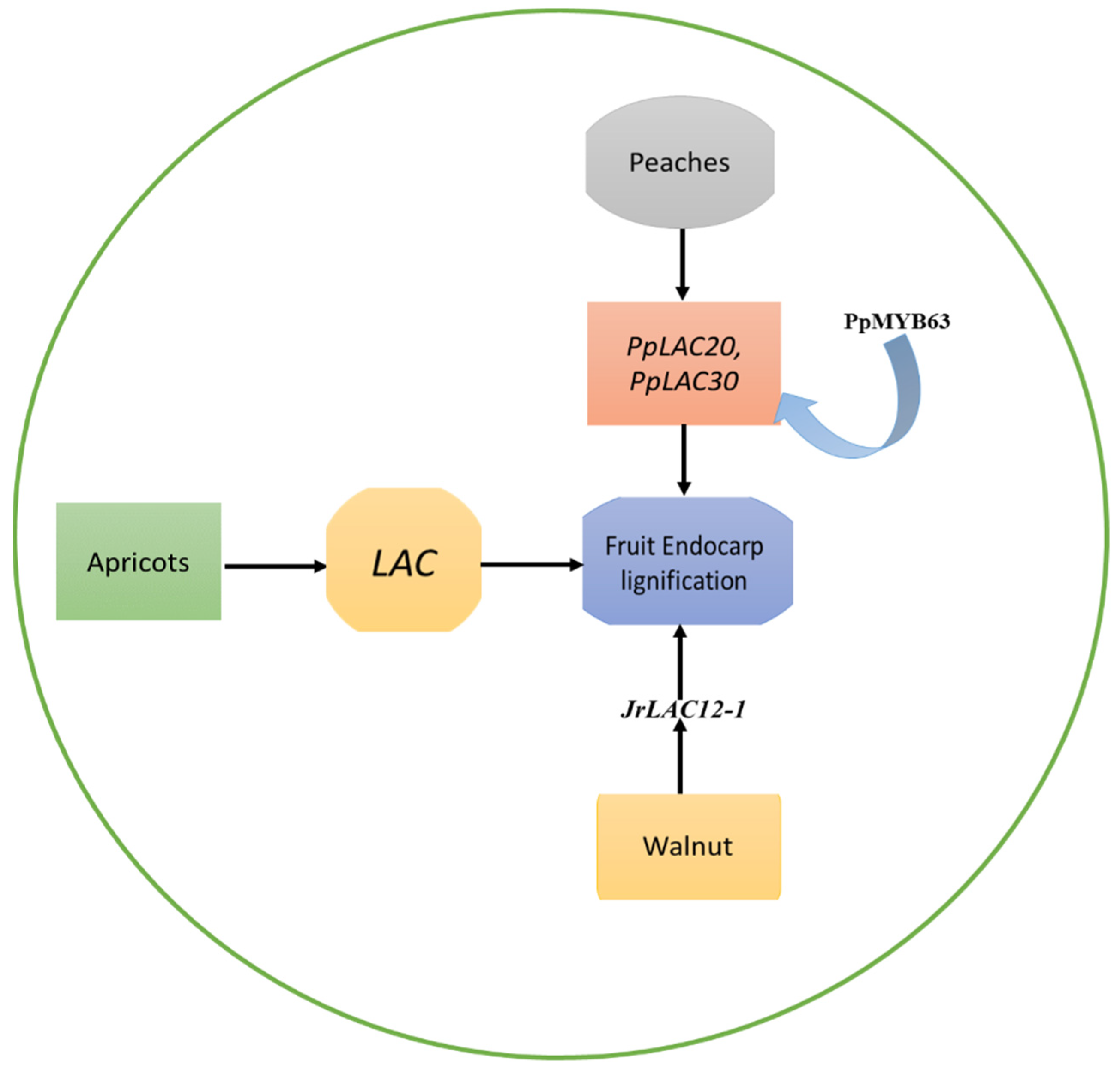
2.1. Molecular Mechanism of LAC Genes Involved in Fruit Endocarp Lignification
2.2. Molecular Mechanism of LAC Genes Involved in Fruit Flesh Lignification
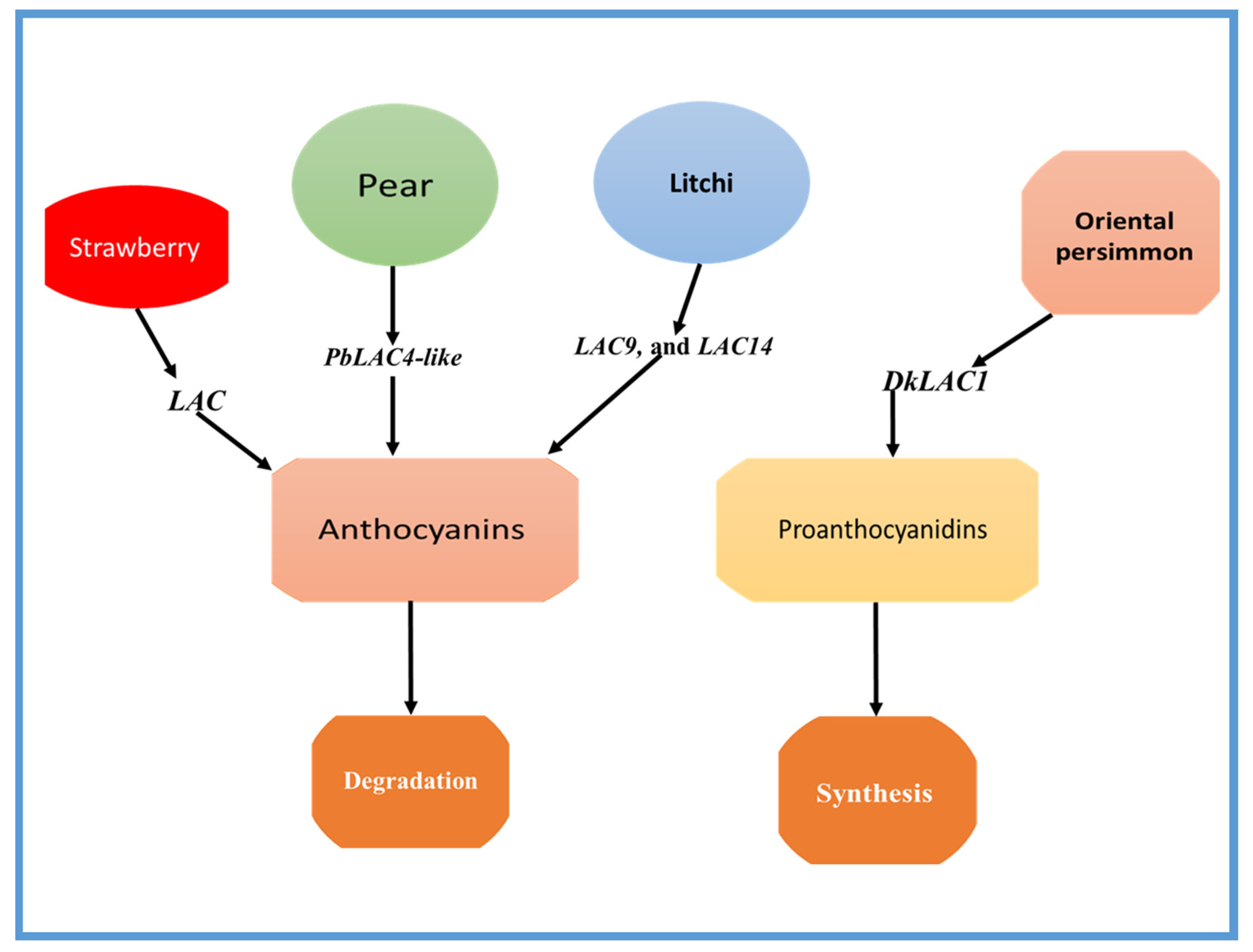
3. Molecular Regulation of LAC Gene Family Involved in Anthocyanin and PA Degradation/Biosynthesis in Fruit
4. Molecular Regulation of the LAC Gene Family Involved in the Abiotic Stress Response in Fruits
5. Molecular Regulation of the LAC Gene Family Involved in the Protection of Fruits from Diseases
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muhammad, N.; Zhao, Z.H.; Meng, X.W.; Wang, J.R.; Yang, M.; Liu, M.J. Stoneless fruit cultivars: Past, present and future perspectives with special reference to economically important Ziziphus species. Acta Hortic. 2022, 1350, 79–90. [Google Scholar]
- Shi, J.; Yao, J.; Tong, R.; Wang, S.; Li, M.; Song, C.; Wan, R.; Jiao, J.; Zheng, X. Genome-Wide Identification of Laccase Gene Family from Punica granatum and Functional Analysis towards Potential Involvement in Lignin Biosynthesis. Horticulturae 2023, 9, 918. [Google Scholar]
- Khan, M.K.U.; Muhammad, N.; Jia, Z.; Peng, J.; Liu, M. Mechanism of Stone (Hardened Endocarp) Formation in Fruits: An Attempt toward Pitless Fruits, and Its Advantages and Disadvantages. Genes 2022, 13, 2123. [Google Scholar] [PubMed]
- Hu, Q.; Luo, C.; Zhang, Q.; Luo, Z. Isolation and characterization of a Laccase gene potentially involved in proanthocyanidin polymerization in oriental persimmon (Diospyros kaki Thunb.) fruit. Mol. Biol. Rep. 2013, 40, 2809–2820. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; He, Y.; Lv, S.; Zhu, H.; Wang, T.; Wang, G.; Hong, N.; Wang, L. The PcMYB44-mediated miR397-PcLACs module regulates defence-induced lignification in pear resistance to fungal disease. Mol. Plant Pathol. 2023, 24, 1107–1125. [Google Scholar] [CrossRef]
- Hu, Q.; Min, L.; Yang, X.; Jin, S.; Zhang, L.; Li, Y. Laccase GhLac1 modulates broad-spectrum biotic stress tolerance via manipulating phenylpropanoid pathway and jasmonic acid synthesis. Plant Physiol. 2018, 176, 1808–1823. [Google Scholar] [CrossRef]
- Zhukhlistova, N.E.; Zhukova, Y.N.; Lyashenko, A.V.; Zaĭtsev, V.N.; Mikhaĭlov, A.M. Three-dimensional organization of three-domain copper oxidases: A review. Crystallogr. Rep. 2008, 53, 92–109. [Google Scholar]
- Xue, C.; Yao, J.L.; Xue, Y.S.; Su, G.Q.; Wang, L.; Lin, L.K.; Allan, A.C.; Zhang, S.L.; Wu, J. PbrMYB169 positively regulates lignification of stone cells in pear fruit. J. Exp. Bot. 2019, 70, 1801–1814. [Google Scholar]
- Ranocha, P.; Mcdougall, G.; Hawkins, S.; Sterjiades, R.; Borderies, G.; Stewart, D.; Cabanes-Macheteau, M. Biochemical characterization, molecular cloning and expression of laccases—A divergent gene family—In poplar. Eur. J. Biochem. 1999, 259, 485–495. [Google Scholar] [CrossRef]
- Yihui, G.; Song, J.; Du, L.; Vinqvist, M.; Palmer, L.C.; Fillmore, S.; Pang, X.; Zhang, Z. Characterization of laccase from apple fruit during postharvest storage and its response to diphenylamine and 1-methylcyclopropene treatments. Food Chem. 2018, 253, 314–321. [Google Scholar] [CrossRef]
- Zhao, G.; Xiang, F.; Zhang, S.; Song, J.; Li, X.; Song, L.; Zhai, R.; Yang, C.; Wang, Z.; Ma, F.; et al. PbLAC4-like, activated by PbMYB26, related to the degradation of anthocyanin during color fading in pear. BMC Plant Biol. 2021, 21, 469. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Luo, L.; Wang, X.; Shen, Z.; Zheng, L. Comprehensive analysis of rice laccase gene (OsLAC) family and ectopic expression of OsLAC10 enhances tolerance to copper stress in Arabidopsis. Int. J. Mol. Sci. 2017, 18, 209. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.H.; Zhang, L.Y.; Lin, X.J.; Gao, Y.; Zhang, J.; Huang, W.L.; Zhao, D.; Ferrarezi, R.S.; Fan, G.C.; Chen, L.S. CsiLAC4 modulates boron flow in Arabidopsis and Citrus via high-boron-dependent lignification of cell walls. New Phytol. 2022, 233, 1257–1273. [Google Scholar] [CrossRef] [PubMed]
- Dardick, C.; Callahan, A.M. Evolution of the fruit endocarp: Molecular mechanisms underlying adaptations in seed protection and dispersal strategies. Front. Plant Sci. 2014, 5, 284. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant develppment and plant-environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, M.; Tuskan, G.; Muchero, W.; Chen, J.G. Recent advances in the transcriptional regulation of secondary cell wall biosynthesis in the woody plants. Front. Plant Sci. 2018, 9, 1535. [Google Scholar] [CrossRef]
- Barros, J.; Serk, H.; Granlundz, I.; Pesquet, E. The cell biology of lignification in higher plants. Ann. Bot. 2015, 115, 1053–1074. [Google Scholar]
- Vanholme, R.; De Meester, B.; Ralph, J.; Boerjan, W. Lignin biosynthesis and its integration into metabolism. Curr. Opin. Biotech. 2019, 56, 230–239. [Google Scholar]
- Zhang, Q.; Ma, X.; Liu, W.; Liu, N.; Zhang, Y.; Xu, M.; Liu, S.; Zhang, Y.; Zhao, H.; Liu, J. Metabolomics Data Revealed Metabolite Changes during Endocarp Lignification in Kernel-Using Apricot. Horticulturae 2022, 8, 967. [Google Scholar]
- Zhang, X.; Zhang, L.J.; Zhang, Q.P.; Xu, J.Y.; Liu, W.S.; Dong, W.X. Comparative transcriptome profiling and morphology provide insights into endocarp cleaving of apricot cultivar (Prunus armeniaca L.). BMC Plant Biol. 2017, 17, 72. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Q.P.; Sun, X.Y.; Du, X.; Liu, W.S.; Dong, W.X. Differential expression of genes encoding phenylpropanoid enzymes in an apricot cultivar (Prunus armeniaca L.) with cleavable endocarp. Trees 2019, 33, 1695–1710. [Google Scholar] [CrossRef]
- Li, S.M.; Su, X.Q.; Abdullah, M.; Sun, Y.M.; Li, G.H.; Cheng, X.; Lin, Y.; Cai, Y.P.; Jin, Q. Effects of different pollens on primary metabolism and lignin biosynthesis in pear. Int. J. Mol. Sci. 2018, 19, 2273. [Google Scholar] [CrossRef] [PubMed]
- Dardick, C.D.; Callahan, A.M.; Chiozzotto, R.; Schaffer, R.J.; Piagnani, M.C.; Scorza, R. Stone formation in peach fruit exhibits spatial coordination of the lignin and flavonoid pathways and similarity to Arabidopsisdehiscence. BMC Biol. 2010, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Ryugo, K. The rate of dry weight accumulation by the peach pit during the hardening process. Am. Soc. Horticult. Sci. 1961, 78, 132. [Google Scholar]
- Ryugo, K. Changes in methoxyl content in the peach endocarp and some of its soluble phenolic constituents during lignification. Am. Soc. Horticult. Sci. 1963, 84, 110–115. [Google Scholar]
- Mendu, V.; Harman-Ware, A.E.; Crocker, M.; Jae, J.; Stork, J.; Morton, S., III; Placido, A.; Huber, G.; DeBolt, S. Identification and thermochemical analysis of high-lignin feedstocks for biofuel and biochemical production. Biotechnol. Biofuels 2011, 4, 43. [Google Scholar] [CrossRef]
- Qui, K.; Zhou, H.; Pan, H.; Sheng, Y.; Yu, H.; Xie, Q.; Chen, H.; Cai, Y.; Zhang, J.; He, J. Genome-wide identification and functional analysis of the peach (P. persica) laccase gene family reveal members potentially involved in endocarp lignification. Trees 2022, 36, 1477–1496. [Google Scholar] [CrossRef]
- Abdallah, I.B.; Tlili, N.; Martinez-Force, E.; Rubio, A.G.; Perez-Camino, M.C.; Albouchi, A.; Boukhchina, S. Content of carotenoids, tocopherols, sterols, triterpenic and aliphatic alcohols, and volatile compounds in six walnuts (Juglans regia L.) varieties. Food Chem. 2015, 173, 972–978. [Google Scholar] [CrossRef]
- Yu, A.; Zou, H.; Li, P.; Yao, X.; Guo, J.; Sun, R.; Wang, G.; Xi, X.; Liu, A. Global Transcriptomic Analyses Provide New Insight into the Molecular Mechanisms of Endocarp Formation and Development in Iron Walnut (Juglans sigillata Dode). Int. J. Mol. Sci. 2023, 24, 6543. [Google Scholar] [CrossRef]
- Li, P.; Wang, H.; Liu, P.; Li, Y.; Liu, K.; An, X.; Zhang, Z.; Zhao, S. The role of JrLACs in the lignification of walnut endocarp. BMC Plant Biol. 2021, 21, 511. [Google Scholar] [CrossRef]
- Xue, C.; Yao, J.L.; Qin, M.F.; Zhang, M.Y.; Allan, A.C.; Wang, D.F.; Wu, J. PbrmiR397a regulates lignification during stone cell development in pear fruit. Plant Biotechnol. J. 2019, 17, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Feng, X.; Lai, R.; Hu, Y.; Chen, W.; Wu, R.; Chen, Y. Cloning and expression analysis of the laccase genes from kiwifruit (Actinidia chinensis var. deliciosa ‘Miliang-1’). J. Agric. Biotechnol. 2018, 26, 64–76. [Google Scholar]
- Mi, L.; Ma, D.; Lv, S.; Xu, S.; Zhong, B.; Peng, T.; Liu, D.; Liu, Y. Comparative Transcriptome and sRNAome Analyses Reveal the Regulatory Mechanisms of Fruit Ripening in a Spontaneous Early-Ripening Navel Orange Mutant and Its Wild Type. Genes 2022, 13, 1706. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, S.; Fu, Q.; Wang, Z.; Liu, X.; Sun, L.; Zhao, Z. Transcriptomic and metabolomic analysis reveals a protein module involved in preharvest apple peel browning. Plant Physiol. 2023, 192, 2102–2122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, L.; Zeng, X.; Chen, R.; Yang, S.; Pan, S. Comparative transcriptome analysis reveals fruit discoloration mechanisms in postharvest strawberries in response to high ambient temperature. Food Chem. X 2019, 2, 100025. [Google Scholar] [CrossRef] [PubMed]
- Berni, R.; Piasecki, E.; Legay, S.; Hausman, J.F.; Siddiqui, K.S.; Cai, G.; Guerriero, G. Identification of the laccase-like multicopper oxidase gene family of sweet cherry (Prunus avium L.) and expression analysis in six ancient Tuscan varieties. Sci. Rep. 2019, 9, 3557. [Google Scholar] [CrossRef]
- Gong, X.; Xie, Z.; Qi, K.; Zhao, L.; Yuan, Y.; Xu, J.; Rui, W.; Shiratake, K.; Bao, J.; Khanizadeh, S.; et al. PbMC1a/1b regulates lignification during stone cell development in pear (Pyrus bretschneideri) fruit. Hortic. Res. 2020, 7, 59. [Google Scholar] [CrossRef]
- Fang, F.; Zhang, X.L.; Luo, H.H.; Zhou, J.J.; Gong, Y.H.; Li, W.J.; Shi, Z.W.; He, Q.; Wu, Q.; Li, L.; et al. An intracellular laccase is responsible for epicatechin-mediated anthocyanin degradation in litchi fruit pericarp. Plant Physiol. 2015, 169, 2391–2408. [Google Scholar] [CrossRef]
- Wang, W.; Wang, C.; Xie, Z.; Jia, H.; Tang, W.; Cui, M.; Fang, J. Function analysis of VvmiR397a and its target genes VvLACs in grape berry development. Acta Hortic. Sin. 2018, 45, 1441–1455. [Google Scholar]
- Cho, H.Y.; Lee, C.; Hwang, S.G.; Park, Y.C.; Lim, H.L.; Jang, C.S. Overexpression of the OsChI1 gene, encoding a putative laccase precursor, increases tolerance to drought and salinity stress in transgenic Arabidopsis. Gene 2014, 552, 98–105. [Google Scholar] [CrossRef]
- Ma, J.; Xu, Z.S.; Wang, F.; Xiong, A.S. Isolation, purification and characterization of two laccases from carrot (Daucus carota L.) and their response to abiotic and metal ions stresses. Protein J. 2015, 34, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Zhang, L.; Tan, M.; Wang, X.; Wang, G.L.; Qi, M.; Liu, B.; Gao, J.; Pan, Y.; Wang, Y. Genome-wide identification and characterization of laccase family members in eggplant (Solanum melongena L.). PeerJ 2022, 10, 12922. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhou, Y.; Wang, B.; Ding, L.; Wang, Y.; Luo, L.; Zhang, Y.; Kong, W. Genome-wide identification and characterization of laccase gene family in Citrus sinensis. Gene 2019, 689, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Yang, T.; Zhang, Y.; Miao, X.; Jin, C.; Xu, X. Genome-wide analyses and expression patterns under abiotic stress of LAC gene family in pear (Pyrus bretschneideri). Plant Biotechnol. Rep. 2021, 15, 403–416. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, Y.; Liang, M.; Kong, W.; Liu, J. The citrus laccase gene CsLAC18 contributes to cold tolerance. Int. J. Mol. Sci. 2022, 23, 14509. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, L.; Wang, X.; Chen, B.; Zhao, J.; Cui, J. The cotton laccase gene GhLAC15 enhances Verticillium wilt resistance via an increase in defence-induced lignification and lignin components in the cell walls of plants. Mol. Plant Pathol. 2019, 20, 309–322. [Google Scholar] [CrossRef]
- Hou, Y.; Yu, X.; Chen, W.; Zhuang, W.; Wang, S.; Sun, C. MdWRKY75e enhances resistance to Alternaria alternata in Malus domestica. Hortic. Res. 2021, 8, 225. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Species | Genes | Function | Reference |
|---|---|---|---|
| Endocarp lignification | |||
| Peach | PpLAC7, 19, 20, 21, 27, 28, and 30 | Contribute to lignin synthesis and endocarp hardness in peach fruit. | [27] |
| PpLAC20 and PpLAC30 | PpMYB63, which is similar to AtMYB58 and AtMYB63, have the ability to activate the promoters of PpLAC20 and PpLAC30. | [27] | |
| Pomegranate | PgLAC16 | Increased expression of PgLAC16 may impede the accumulation of storage material during seed development, leading to the prevention of seed hardness in soft-seed pomegranate. | [2] |
| PgLAC37 and PgLAC50 | Hard-seed pomegranate cultivar exhibited more significant amounts of PgLAC37 and PgLAC50. | [2] | |
| Walnut | JrLAC12-1, JrLAC12-2, and JrLAC16 | Play a role in lignin biosynthesis. | [30] |
| Flesh lignification | |||
| Pear | LAC genes | The reduction in lignin content and stone cell number in pear fruit was observed when PbrmiR397a was overexpressed and three LAC genes were simultaneously silenced. | [8] |
| LAC18 | PbrMYB169 protein was found to activate the promoters of several lignin genes including LAC18 through binding with AC elements [ACC(T/A)ACC]. | [31] | |
| Kiwifruit | LAC genes | The expression of LAC genes was correlated with the production of lignin, which contributes to the firmness of the fruit. | [32] |
| Fruit ripening | |||
| Oranges | LAC4, 17, 22 | Fruit ripening and quality development. | [33] |
| Anthocyanin and PA degradation/ Biosynthesis | |||
| Apple | MdLAC7 | Apple peel browning by producing pigments. | [34] |
| Pear | PbLAC4-like | Anthocyanin degradation. | [11] |
| Strawberry | LAC9, and LAC14 | Anthocyanin degradation. | [35] |
| Oriental persimmon (Diospyros kaki Thunb.) | DkLAC1 | The DkLAC1 is a laccase gene found in plants and is closely related to AtLAC15, an enzyme known for its involvement in the polymerization of PAs. | [4] |
| Protection of fruits from diseases | |||
| Chinese pear | PbrLAC | PcmiR397 and overexpression of a single PcLAC in pear increased resistance to pathogens through enhanced lignin synthesis. | [5] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.K.U.; Zhang, X.; Ma, Z.; Huang, M.; Yang, C.; Wang, X.; Liu, M.; Peng, J. Contribution of the LAC Genes in Fruit Quality Attributes of the Fruit-Bearing Plants: A Comprehensive Review. Int. J. Mol. Sci. 2023, 24, 15768. https://doi.org/10.3390/ijms242115768
Khan MKU, Zhang X, Ma Z, Huang M, Yang C, Wang X, Liu M, Peng J. Contribution of the LAC Genes in Fruit Quality Attributes of the Fruit-Bearing Plants: A Comprehensive Review. International Journal of Molecular Sciences. 2023; 24(21):15768. https://doi.org/10.3390/ijms242115768
Chicago/Turabian StyleKhan, Muhammad Khalil Ullah, Xiaojie Zhang, Zitan Ma, Mingxia Huang, Ce Yang, Xiaoming Wang, Mengjun Liu, and Jianying Peng. 2023. "Contribution of the LAC Genes in Fruit Quality Attributes of the Fruit-Bearing Plants: A Comprehensive Review" International Journal of Molecular Sciences 24, no. 21: 15768. https://doi.org/10.3390/ijms242115768
APA StyleKhan, M. K. U., Zhang, X., Ma, Z., Huang, M., Yang, C., Wang, X., Liu, M., & Peng, J. (2023). Contribution of the LAC Genes in Fruit Quality Attributes of the Fruit-Bearing Plants: A Comprehensive Review. International Journal of Molecular Sciences, 24(21), 15768. https://doi.org/10.3390/ijms242115768