
Molecular Action of Tamoxifen in the Ovaries of Rats with Mammary Neoplasia
 , ,
, ,
Abstract
:1. Introduction
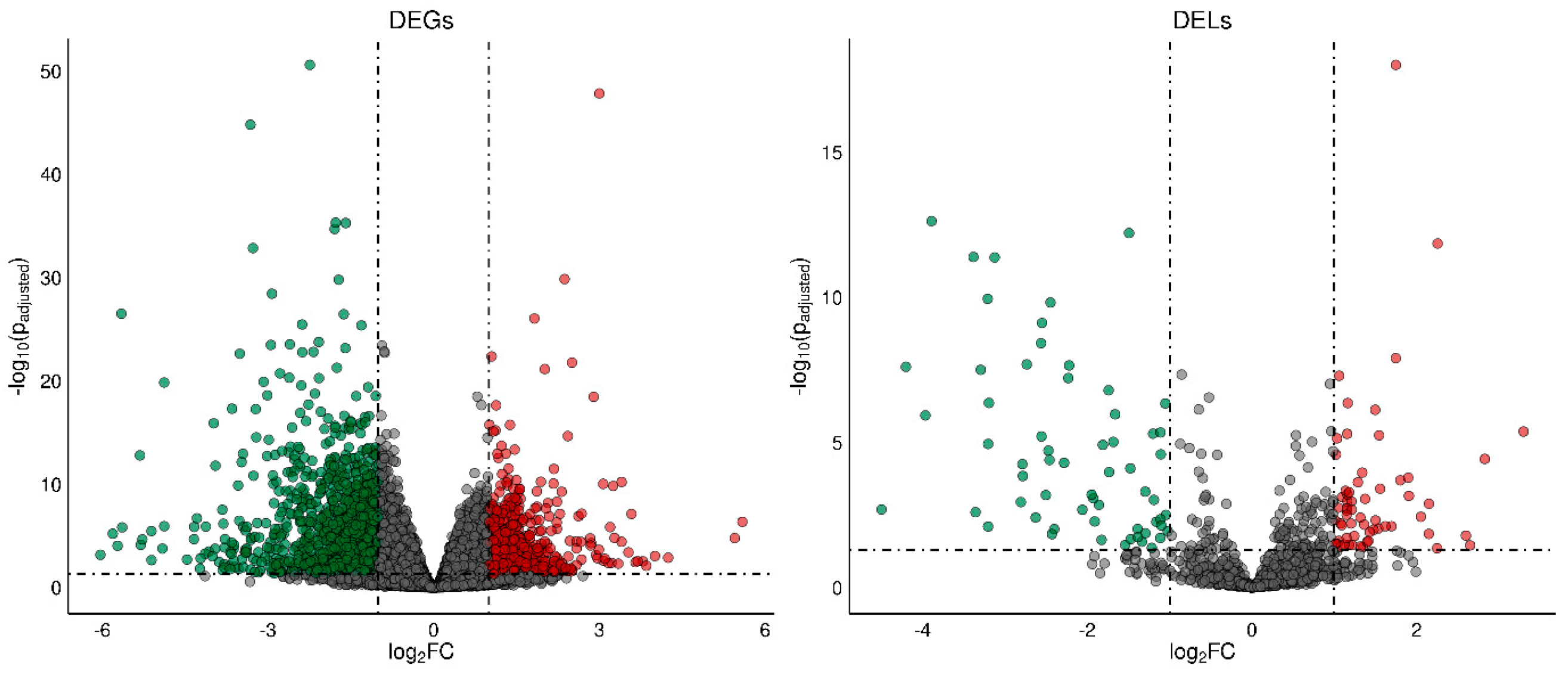
2. Results
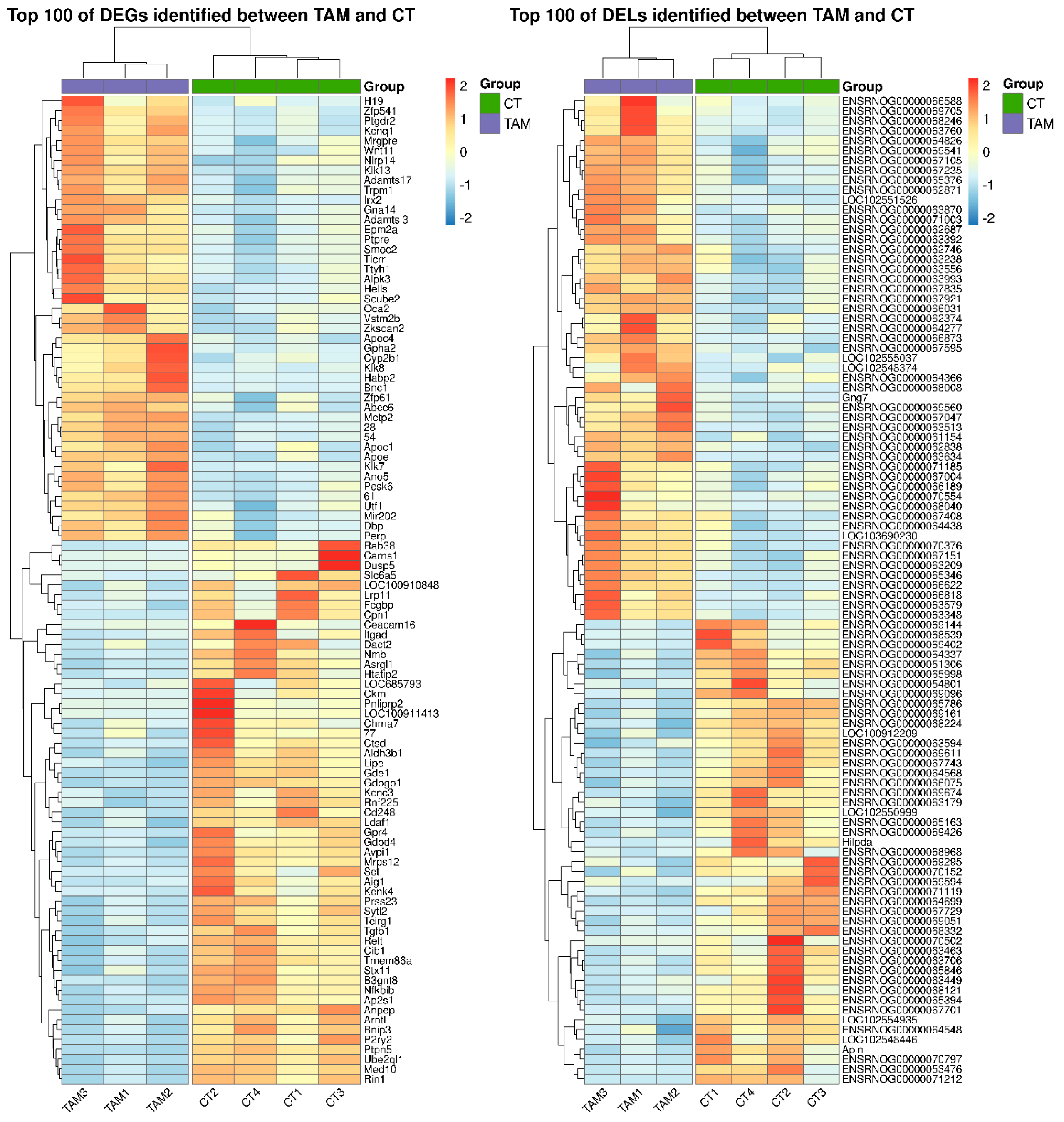
2.1. Effects of TAM on the mRNA Expression Profile of Rat Ovaries
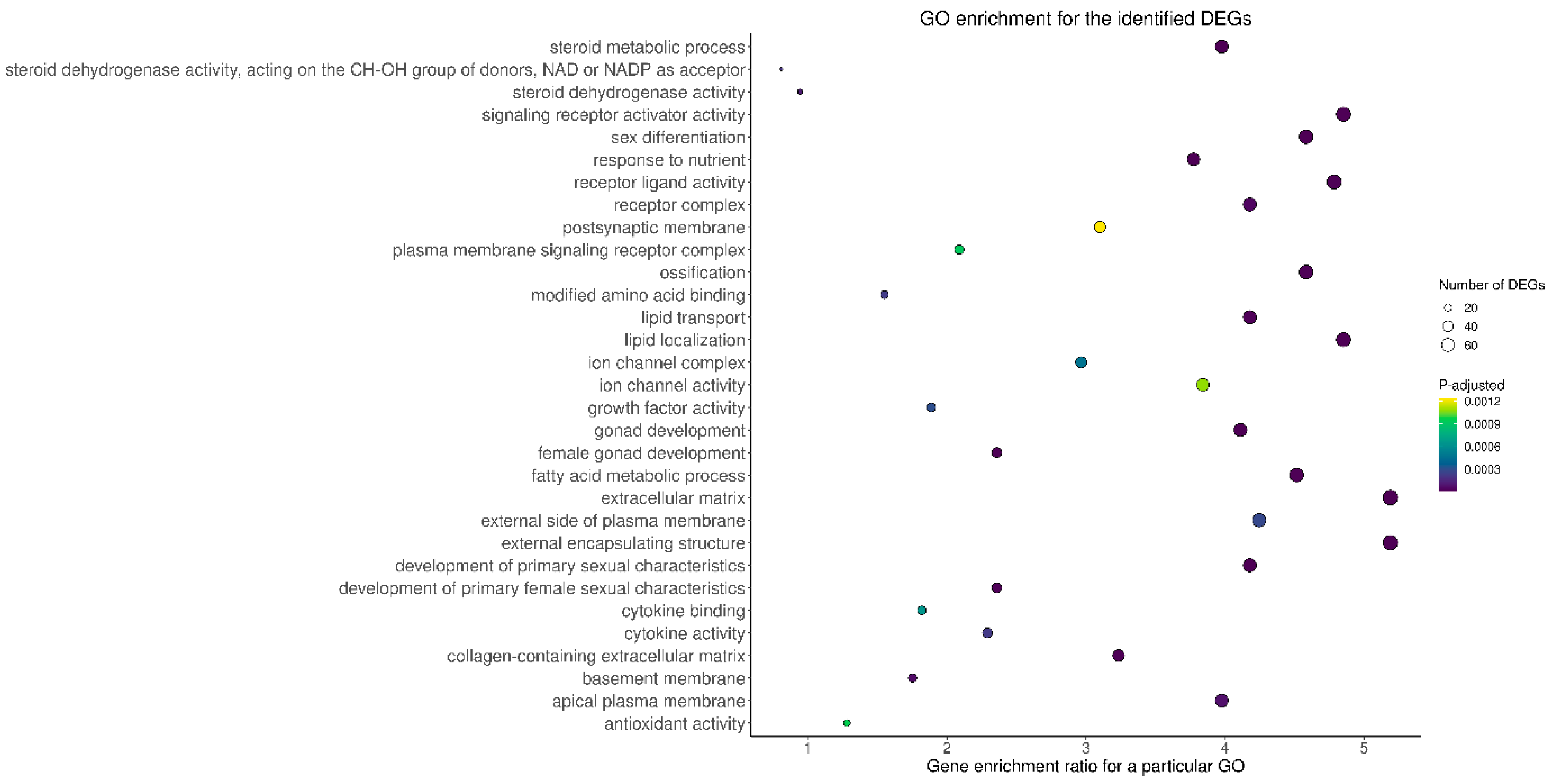
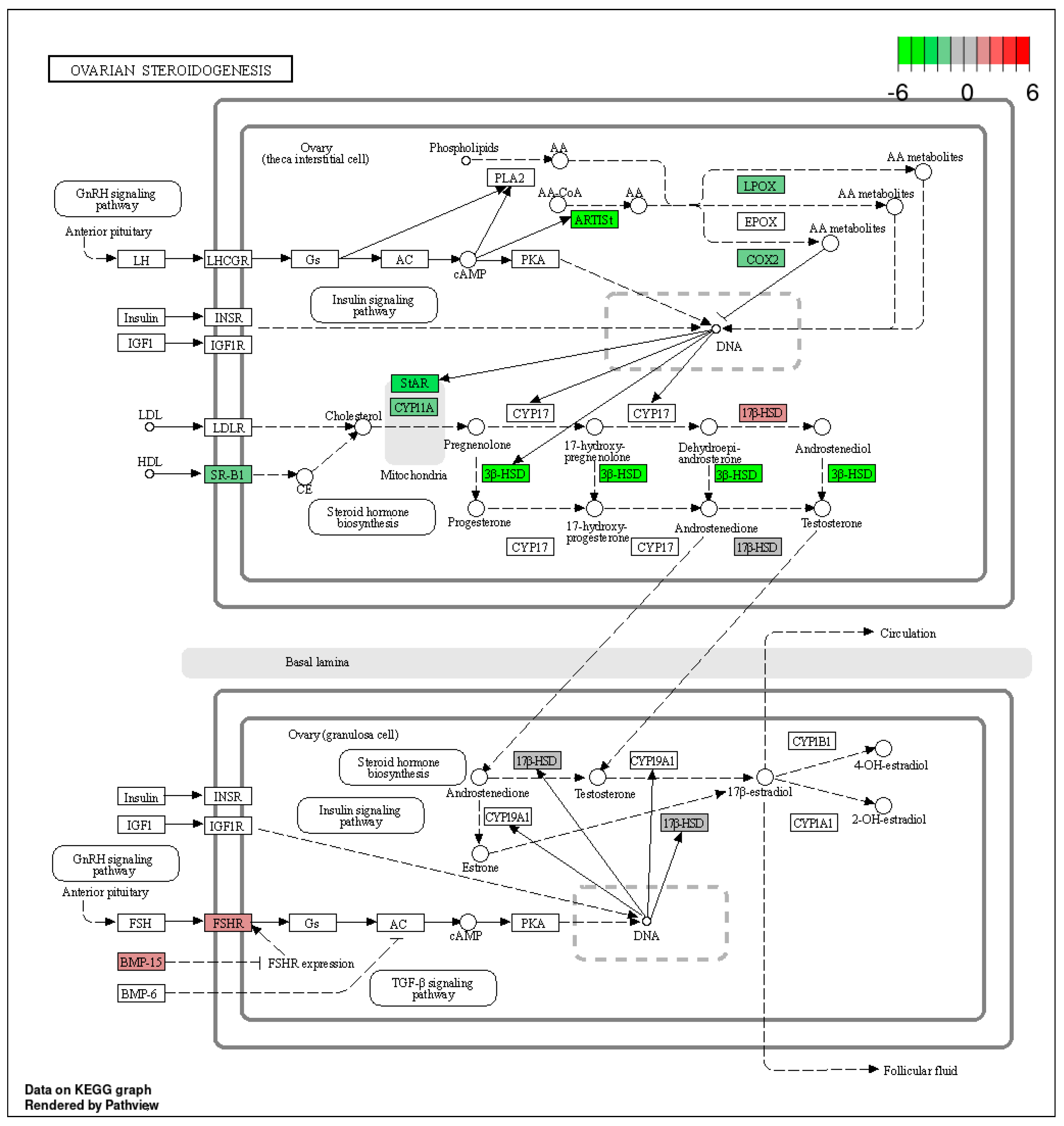
2.2. Functional Enrichment of the Identified DEGs
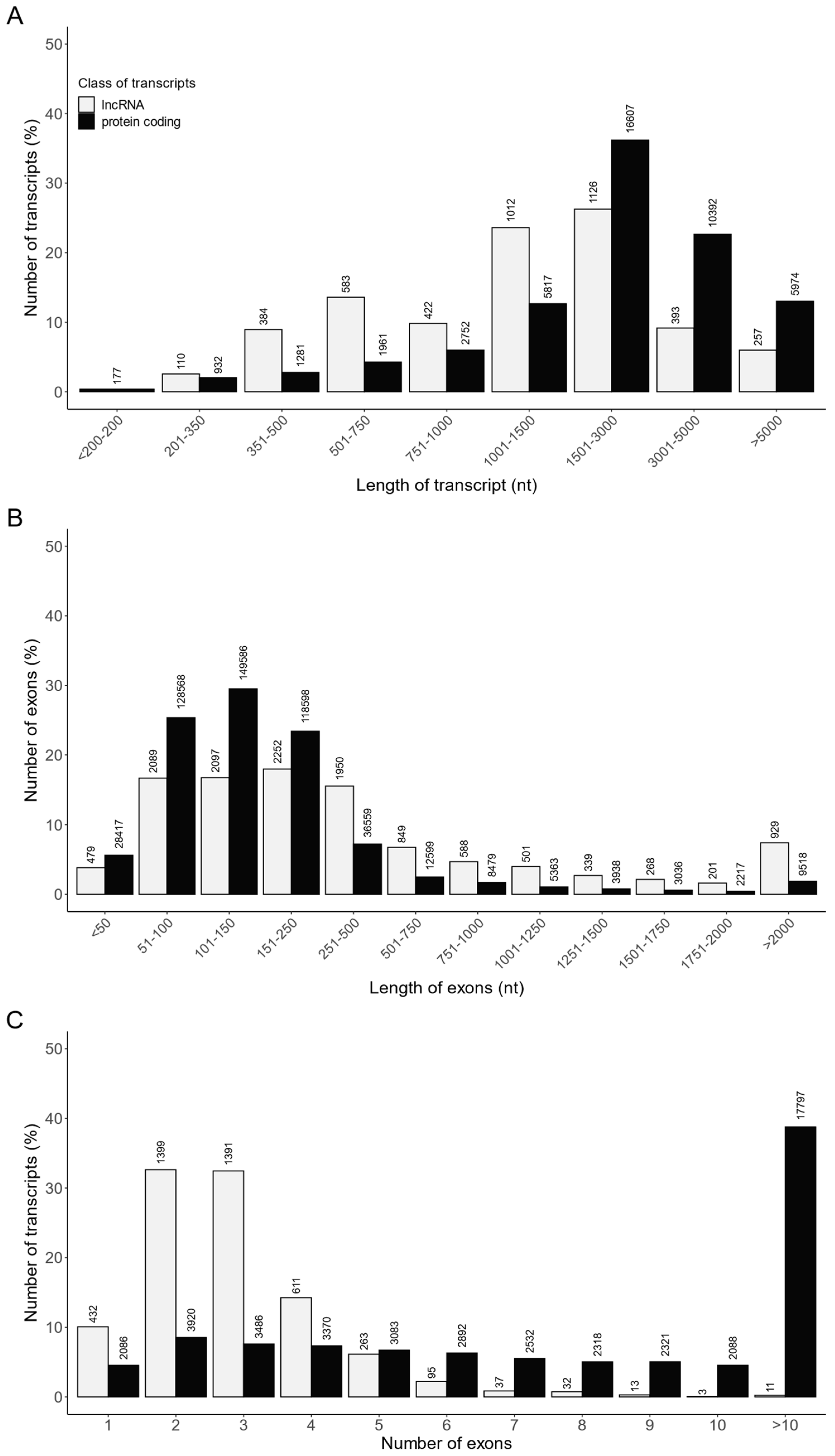
2.3. Effects of TAM on the lncRNA Expression Profile of Rat Ovaries
2.4. Validation of RNA-Seq Data by Real-Time PCR
2.5. Effects of TAM on the Proteome of Rat Ovaries
3. Discussion
4. Material and Methods
4.1. Animals and Treatments
4.2. Total RNA Isolation and Sequencing
4.3. Identification of lncRNA
4.4. Identification of Differentially Expressed lncRNAs (DELs) and Genes (DEGs)
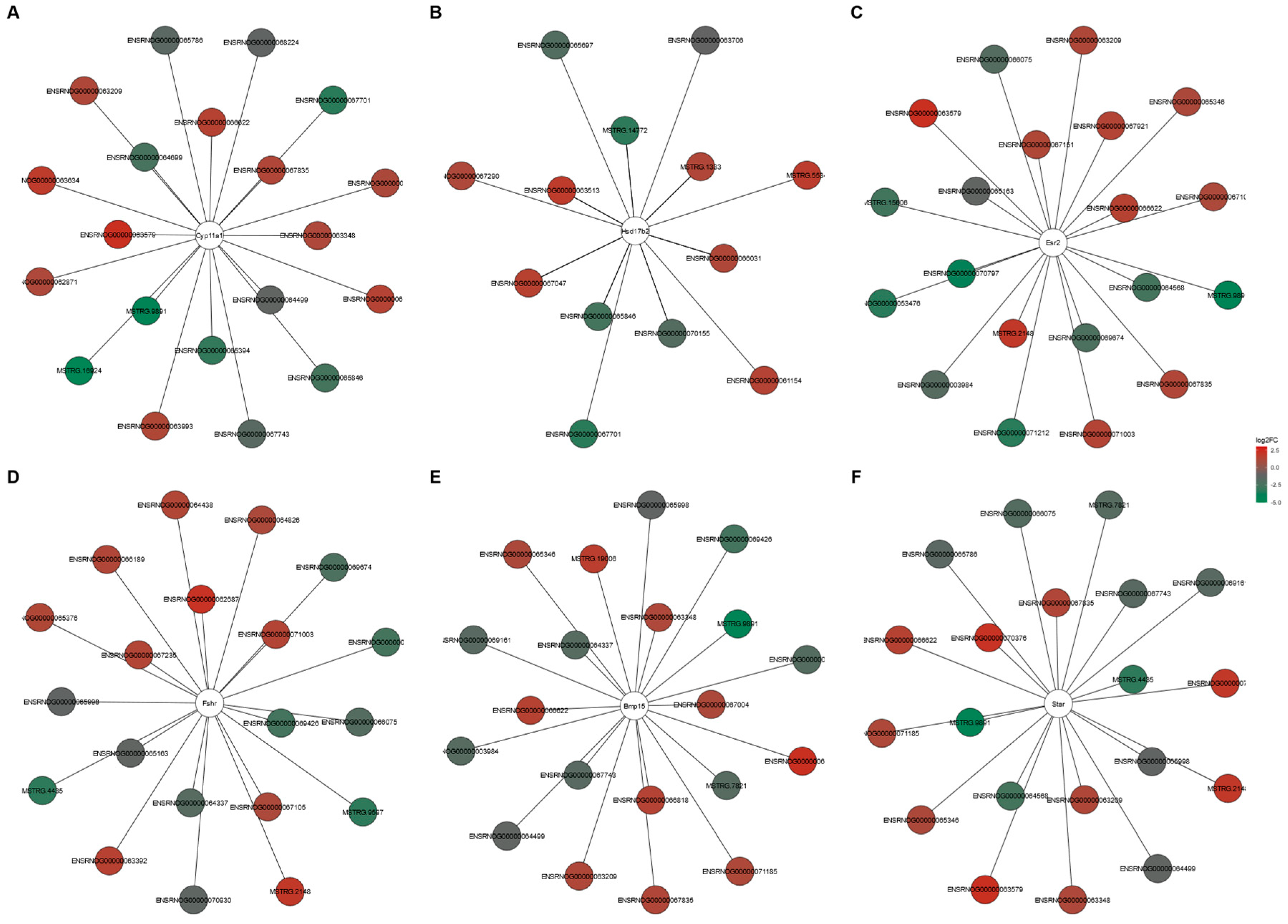
4.5. Cis- and Trans-Regulated Target Gene Prediction of lncRNAs
4.6. Functional Enrichment Analysis
4.7. Real-Time PCR
4.8. Protein Isolation and 2D-DIGE
4.9. Protein Digestion and MALDI-TOF/TOF Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Howell, A.; Howell, S.J. Tamoxifen evolution. Br. J. Cancer 2023, 128, 421–425. [Google Scholar] [CrossRef]
- Yen, C.; Zhao, F.; Yu, Z.; Zhu, X.; Li, C.G. Interactions Between Natural Products and Tamoxifen in Breast Cancer: A Comprehensive Literature Review. Front. Pharmacol. 2022, 13, 847113. [Google Scholar] [CrossRef]
- Mata, D.G.d.M.M.; Amir Carmona, C.; Eisen, A.; Trudeau, M. Appraising Adjuvant Endocrine Therapy in Hormone Receptor Positive HER2-Negative Breast Cancer—A Literature Review. Curr. Oncol. 2022, 29, 4956–4969. [Google Scholar] [CrossRef]
- Schiff, R.; Massarweh, S.A.; Shou, J.; Bharwani, L.; Arpino, G.; Rimawi, M.; Osborne, C.K. Advanced concepts in estrogen receptor biology and breast cancer endocrine resistance: Implicated role of growth factor signaling and estrogen receptor coregulators. Cancer Chemother. Pharmacol. 2005, 56 (Suppl. 1), 10–20. [Google Scholar] [CrossRef]
- Clemons, M.; Danson, S.; Howell, A. Tamoxifen (“Nolvadex”): A review. Cancer Treat. Rev. 2002, 28, 165–180. [Google Scholar] [CrossRef]
- Jordan, V.C. New insights into the metabolism of tamoxifen and its role in the treatment and prevention of breast cancer. Steroids 2007, 72, 829–842. [Google Scholar] [CrossRef]
- Polin, S.A.; Ascher, S.M. The effect of tamoxifen on the genital tract. Cancer Imaging 2008, 8, 135–145. [Google Scholar] [CrossRef]
- Cohen, I.; Figer, A.; Tepper, R.; Shapira, J.; Altaras, M.M.; Yigael, D.; Beyth, Y. Ovarian overstimulation and cystic formation in premenopausal tamoxifen exposure: Comparison between tamoxifen-treated and nontreated breast cancer patients. Gynecol. Oncol. 1999, 72, 202–207. [Google Scholar] [CrossRef]
- Kojima, N.; Yamasaki, Y.; Koh, H.; Miyashita, M.; Morita, H. Long-Acting Luteinizing Hormone-Releasing Hormone Agonist for Ovarian Hyperstimulation Induced by Tamoxifen for Breast Cancer. Case Rep Obstet Gynecol. 2018, 2018, 4931852. [Google Scholar] [CrossRef]
- Metindir, J.; Aslan, S.; Bilir, G. Ovarian cyst formation in patients using tamoxifen for breast cancer. Jpn. J. Clin. Oncol. 2005, 35, 607–611. [Google Scholar] [CrossRef]
- Yamazaki, R.; Inokuchi, M.; Ishikawa, S.; Myojo, S.; Iwadare, J.; Bono, Y.; Mizumoto, Y.; Nakamura, M.; Takakura, M.; Iizuka, T.; et al. Tamoxifen-induced ovarian hyperstimulation during premenopausal hormonal therapy for breast cancer in Japanese women. SpringerPlus 2015, 4, 425. [Google Scholar] [CrossRef] [PubMed]
- Groom, G.V.; Griffiths, K. Effect of the anti-oestrogen tamoxifen on plasma levels of luteinizing hormone, follicle-stimulating hormone, prolactin, oestradiol and progesterone in normal pre-menopausal women. J. Endocrinol. 1976, 70, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Komatsu, K.; Osuka, S.; Murase, T.; Bayasula, B.; Nakanishi, N.; Nakamura, T.; Goto, M.; Iwase, A.; Masubuchi, S.; et al. Tamoxifen Activates Dormant Primordial Follicles in Mouse Ovaries. Reprod. Sci. 2022, 29, 3404–3412. [Google Scholar] [CrossRef]
- Zık, B.; Kurnaz, H.; Güler, S.; Asmaz, E.D. Effect of tamoxifen on the Notch signaling pathway in ovarian follicles of mice. Biotech. Histochem. 2019, 94, 410–419. [Google Scholar] [PubMed]
- Akduman, A.T.; Özerkan, K.; Zik, B.; Peker, S.; Avcı, B.; Ata, B. Effect of tamoxifen on ovarian reserve: A randomized controlled assessor-blind trial in a mouse model. J. Turk. Ger. Gynecol. Assoc. 2014, 15, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Tsujioka, S.; Ban, Y.; Wise, L.D.; Tsuchiya, T.; Sato, T.; Matsue, K.; Ikeda, T.; Sasaki, M.; Nishikibe, M. Collaborative work on evaluation of ovarian toxicity. 3) Effects of 2- or 4- week repeated-dose toxicity and fertility studies with tamoxifen in female rats. J. Toxicol. Sci. 2009, 34, 43–51. [Google Scholar]
- Nynca, A.; Swigonska, S.; Ruszkowska, M.; Sadowska, A.; Orlowska, K.; Molcan, T.; Myszczynski, K.; Otrocka-Domagala, I.; Paździor-Czapula, K.; Kurowicka, B.; et al. Tamoxifen decreases ovarian toxicity without compromising cancer treatment in a rat model of mammary cancer. BMC Genom. 2023, 24, 325. [Google Scholar] [CrossRef]
- Roness, H.; Kalich-Philosoph, L.; Meirow, D. Prevention of chemotherapy-induced ovarian damage: Possible roles for hormonal and non-hormonal attenuating agents. Hum. Reprod. Update 2014, 20, 759–774. [Google Scholar] [CrossRef]
- Ting, A.Y.; Petroff, B.K. Challenges and potential for ovarian preservation with SERMs. Biol. Reprod. 2015, 92, 133. [Google Scholar]
- Ting, A.Y.; Petroff, B.K. Tamoxifen decreases ovarian follicular loss from experimental toxicant DMBA and chemotherapy agents cyclophosphamide and doxorubicin in the rat. J. Assist. Reprod. Genet. 2010, 27, 591–597. [Google Scholar]
- Mahran, Y.F.; El-Demerdash, E.; Nada, A.S.; Ali, A.A.; Abdel-Naim, A.B. Insights into the protective mechanisms of tamoxifen in radiotherapy-induced ovarian follicular loss: Impact on insulin-like growth factor 1. Endocrinology 2013, 154, 3888–3899. [Google Scholar] [PubMed]
- Jordan, V.C. Tamoxifen as the first targeted long-term adjuvant therapy for breast cancer. Endocrinol. Relat. Cancer 2014, 21, R235–R246. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Shin, H.C. Risk Factors for Tamoxifen-Induced Ovarian Hyperstimulation in Breast Cancer Patients. Clin. Breast Cancer 2020, 20, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Noh, W.C.; Nam, S.J.; Park, B.W.; Lee, E.S.; Im, S.A.; Jung, Y.S.; Yoon, J.H.; Kang, S.S.; Park, K.H.; et al. Five-year changes in ovarian function restoration in premenopausal patients with breast cancer taking tamoxifen after chemotherapy: An ASTRRA study report. Eur. J. Cancer 2021, 151, 190–200. [Google Scholar] [CrossRef]
- Pafili, K.; Paschou, S.A.; Armeni, E.; Polyzos, S.A.; Goulis, D.G.; Lambrinoudaki, I. Non-alcoholic fatty liver disease through the female lifespan: The role of sex hormones. J. Endocrinol. Investig. 2022, 45, 1609–1623. [Google Scholar] [CrossRef]
- Yoo, J.J.; Lim, Y.S.; Kim, M.S.; Lee, B.; Kim, B.Y.; Kim, Z.; Lee, J.E.; Lee, M.H.; Kim, S.G.; Kim, Y.S. Risk of fatty liver after long-term use of tamoxifen in patients with breast cancer. PLoS ONE 2020, 15, e0236506. [Google Scholar] [CrossRef]
- Lorizio, W.; Wu, A.H.; Beattie, M.S.; Rugo, H.; Tchu, S.; Kerlikowske, K.; Ziv, E. Clinical and biomarker predictors of side effects from tamoxifen. Breast Cancer Res. Treat. 2012, 132, 1107–1118. [Google Scholar] [CrossRef]
- Selvaraj, V.; Stocco, D.M.; Clark, B.J. Current knowledge on the acute regulation of steroidogenesis. Biol. Reprod. 2018, 99, 13–26. [Google Scholar] [CrossRef]
- Connan-Perrot, S.; Leger, T.; Lelandais, P.; Desdoits-Lethimonier, C.; David, A.; Fowler, P.A.; Mazaud-Guittot, S. Six decades of research on human fetal gonadal steroids. Int. J. Mol. Sci. 2021, 22, 6681. [Google Scholar] [CrossRef]
- Xiao, C.; Wang, J.; Zhang, C. Synthesis, Regulatory Factors, and Signaling Pathways of Estrogen in the Ovary. Reprod. Sci. 2023, 30, 350–360. [Google Scholar] [CrossRef]
- Gao, X.; Petroff, B.K.; Oluola, O.; Georg, G.; Terranova, P.F.; Rozman, K.K. Endocrine disruption by indole-3-carbinol and tamoxifen: Blockage of ovulation. Toxicol. Appl. Pharmacol. 2002, 183, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; He, M.; Zhang, J.; Tong, Y.; Chen, T.; Wang, C.; Pan, W.; Xiao, Z. Mechanisms of primordial follicle activation and new pregnancy opportunity for premature ovarian failure patients. Front. Physiol. 2023, 14, 1113684. [Google Scholar] [CrossRef] [PubMed]
- Kezele, P.R.; Ague, J.M.; Nilsson, E.; Skinner, M.K. Alterations in the ovarian transcriptome during primordial follicle assembly and development. Biol. Reprod. 2005, 72, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Chakravarthi, V.; Ghosh, S.; Roby, K.F.; Wolfe, M.W.; Rumi, M.A.K. A Gatekeeping Role of ESR2 to Maintain the Primordial Follicle Reserve. Endocrinology 2020, 161, bqaa037. [Google Scholar] [CrossRef]
- Gou, R.; Hu, Y.; Liu, O.; Dong, H.; Gao, L.; Wang, S.; Zheng, M.; Li, X.; Lin, B. PGK1 Is a Key Target for Anti-Glycolytic Therapy of Ovarian Cancer: Based on the Comprehensive Analysis of Glycolysis-Related Genes. Front. Oncol. 2021, 11, 682461. [Google Scholar] [CrossRef]
- Li, R.; Wang, W.; Yang, Y.; Gu, C. Exploring the role of glucose-6-phosphate dehydrogenase in cancer (Review). Oncol. Rep. 2020, 44, 2325–2336. [Google Scholar] [CrossRef]
- Johansson, H.K.L.; Svingen, T.; Boberg, J.; Fowler, P.A.; Stead, D.; Vinggaard, A.M.; Filis, P. Calretinin is a novel candidate marker for adverse ovarian effects of early life exposure to mixtures of endocrine disruptors in the rat. Arch. Toxicol. 2020, 94, 1241–1250. [Google Scholar] [CrossRef]
- Hoter, A.; Naim, H.Y. Heat Shock Proteins and Ovarian Cancer: Important Roles and Therapeutic Opportunities. Cancers 2019, 11, 1389. [Google Scholar] [CrossRef]
- Chew, G.L.; Huo, C.W.; Huang, D.; Blick, T.; Hill, P.; Cawson, J.; Frazer, H.; Southey, M.C.; Hopper, J.L.; Britt, K.; et al. Effects of tamoxifen and oestrogen on histology and radiographic density in high and low mammographic density human breast tissues maintained in murine tissue engineering chambers. Breast Cancer Res. Treat. 2014, 148, 303–314. [Google Scholar] [CrossRef]
- Yager, J.D.; Roebuck, B.D.; Paluszcyk, T.L.; Memoli, V.A. Effects of ethinyl estradiol and tamoxifen on liver DNA turnover and new synthesis and appearance of gamma glutamyl transpeptidase-positive foci in female rats. Carcinogenesis 1986, 7, 2007–2014. [Google Scholar] [CrossRef]
- Swigonska, S.; Nynca, A.; Molcan, T.; Jablonska, M.; Ciereszko, R.E. Knock-down of aryl hydrocarbon receptor (AhR) affects the lncRNA-mediated response of porcine granulosa cells (AVG-16 cell line) to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Anim. Reprod. Sci. 2023, 255, 107277. [Google Scholar] [CrossRef] [PubMed]
- Snel, B.; Lehmann, G.; Bork, P.; Huynen, M.A. STRING: A web-server to retrieve and display the repeatedly occurring neighbourhood of a gene. Nucleic Acids Res. 2000, 28, 3442–3444. [Google Scholar] [CrossRef] [PubMed]
- Sadowska, A.; Nynca, A.; Ruszkowska, M.; Paukszto, L.; Myszczynski, K.; Orlowska, K.; Swigonska, S.; Molcan, T.; Jastrzebski, J.P.; Ciereszko, R.E. Transcriptional profiling of porcine granulosa cells exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Chemosphere 2017, 178, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Orlowska, K.; Swigonska, S.; Sadowska, A.; Ruszkowska, M.; Nynca, A.; Molcan, T.; Ciereszko, R.E. The effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin on the proteome of porcine granulosa cells. Chemosphere 2018, 212, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Ramagli, L.S.; Rodriguez, L.V. Quantitation of microgram amounts of protein in two-dimensional polyacrylamide gel electrophoresis sample buffer. Electrophoresis 1985, 6, 559–563. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identified Proteins | MASCOT Protein Score | Sequence Coverage (%) | Number of Peptides | Fold Change | Accession Number |
|---|---|---|---|---|---|
| 60 kDa heat shock protein, mitochondrial (CH60) | 228 | 29 | 11 | −4.1 | gi ǀ206597443 |
| Vimentin (VIM) | 287 | 26 | 12 | −2.3 | gi ǀ38197662 |
| Calretinin (CALB2) | 113 | 14 | 4 | −2.1 | gi ǀ16758892 |
| Phosphoglycerate kinase 1 (PGK1) | 52 | 3 | 1 | −1.8 | gi ǀ56585024 |
| Glucose-6-phosphate 1-dehydrogenase (G6PD) | 69 | 12 | 5 | −1.8 | gi ǀ8393381 |
| Tropomyosin alpha-3 chain (TPM3) | 722 | 49 | 22 | −1.8 | gi ǀ669633260 |
| Fructose-bisphosphate aldolase A (ALDOA) | 108 | 16 | 4 | −1.7 | gi ǀ408772019 |
| Prohibitin (PHB) | 83 | 21 | 5 | −1.4 | gi ǀ13937353 |
| Actin, cytoplasmic 1 (ACTB) | 105 | 35 | 1 | 2.5 | gi ǀ13592133 |
| Aldehyde dehydrogenase, mitochondrial (ALDH2) | 263 | 16 | 8 | 2.3 | gi ǀ1820958497 |
| Heat shock cognate 71 kDa protein (HSP7C) | 107 | 10 | 5 | 1.9 | gi ǀ13242237 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nynca, A.; Swigonska, S.; Molcan, T.; Petroff, B.K.; Ciereszko, R.E. Molecular Action of Tamoxifen in the Ovaries of Rats with Mammary Neoplasia. Int. J. Mol. Sci. 2023, 24, 15767. https://doi.org/10.3390/ijms242115767
Nynca A, Swigonska S, Molcan T, Petroff BK, Ciereszko RE. Molecular Action of Tamoxifen in the Ovaries of Rats with Mammary Neoplasia. International Journal of Molecular Sciences. 2023; 24(21):15767. https://doi.org/10.3390/ijms242115767
Chicago/Turabian StyleNynca, Anna, Sylwia Swigonska, Tomasz Molcan, Brian K. Petroff, and Renata E. Ciereszko. 2023. "Molecular Action of Tamoxifen in the Ovaries of Rats with Mammary Neoplasia" International Journal of Molecular Sciences 24, no. 21: 15767. https://doi.org/10.3390/ijms242115767
APA StyleNynca, A., Swigonska, S., Molcan, T., Petroff, B. K., & Ciereszko, R. E. (2023). Molecular Action of Tamoxifen in the Ovaries of Rats with Mammary Neoplasia. International Journal of Molecular Sciences, 24(21), 15767. https://doi.org/10.3390/ijms242115767