Endothelial Cell Dysfunction and Hypoxia as Potential Mediators of Pain in Fabry Disease: A Human-Murine Translational Approach

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Investigations in Patient-Derived EC
2.1.1. Clinical Characteristics
2.1.2. FD-Patient-Derived EC Carry a Characteristic Cellular Phenotype
2.1.3. Hypoxia-Associated Gene Expression Is Modified in P1 Compared to Control EC
2.1.4. Reduced Nuclear HIF1a and Increased HIF2 Signals in EC of P1 and P2 Compared to Control EC
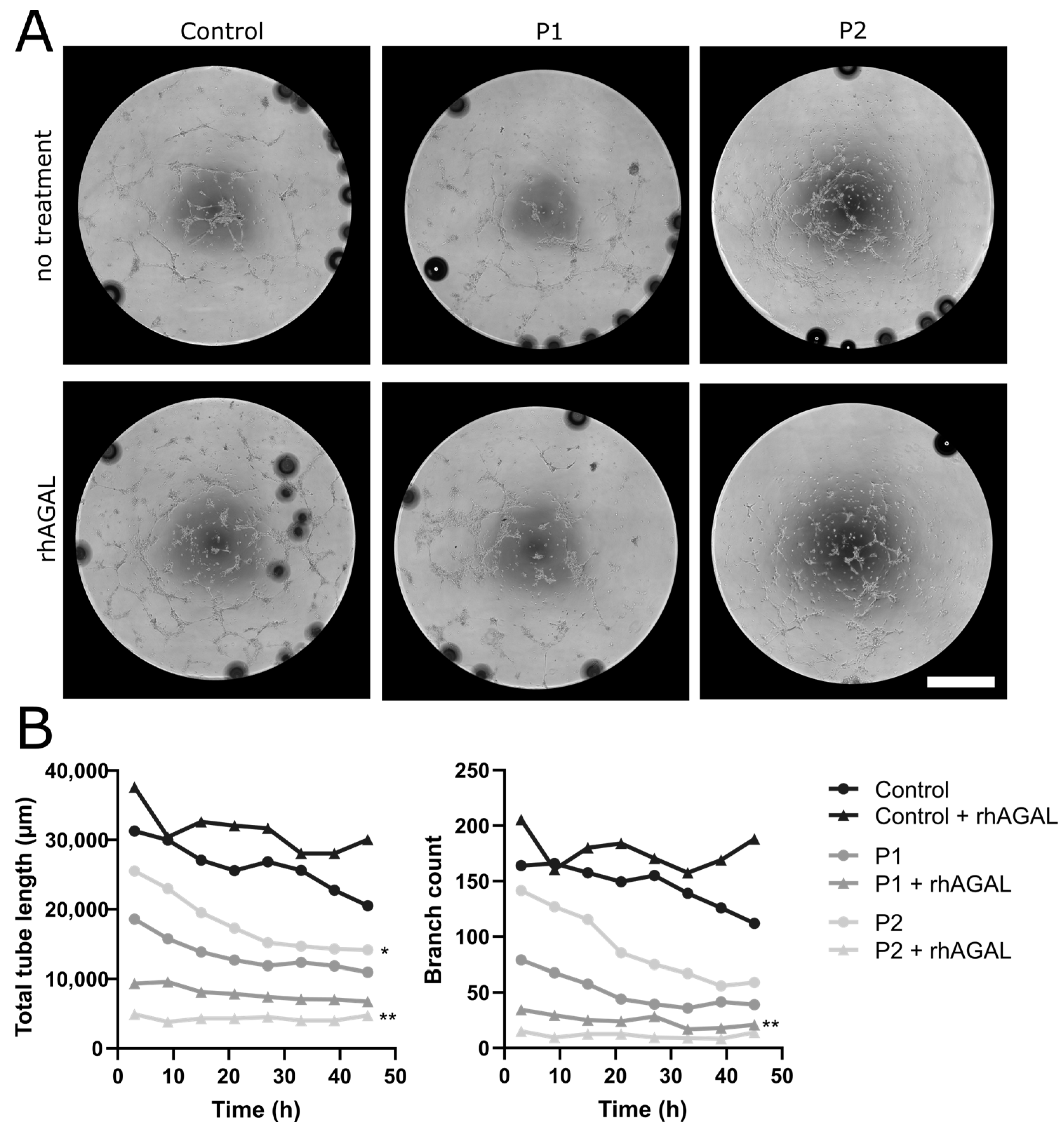
2.1.5. Impaired Tube Formation Properties in FD-Patient-Derived EC under Normal and Hypoxic Conditions
2.1.6. Selective TGFβ Pathway Block Recovers FD-Patient-Derived EC Function
2.1.7. Selective TGFβ Pathway Inhibition Revealed Differential Expression of ALK5 Downstream Target Genes
2.1.8. Mitochondrial Membrane Potential and Form Are Altered in FD Compared to Control EC
2.2. Investigations in Murine DRG Tissue and Neuronal Cell Culture
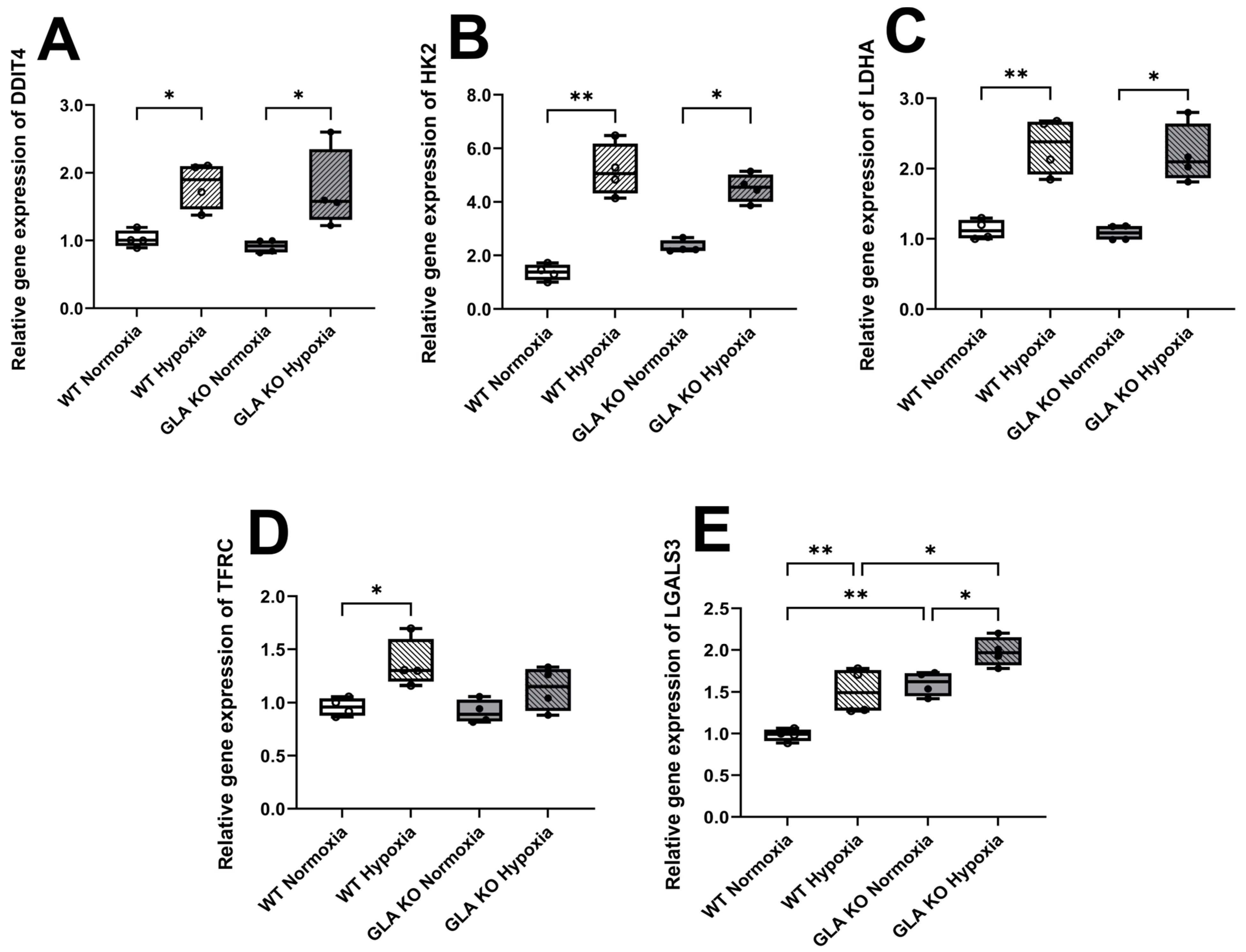
2.2.1. Higher Expression Levels of the Hypoxia-Associated Target Genes in the DRG of Old GLA KO Than Old WT Mice
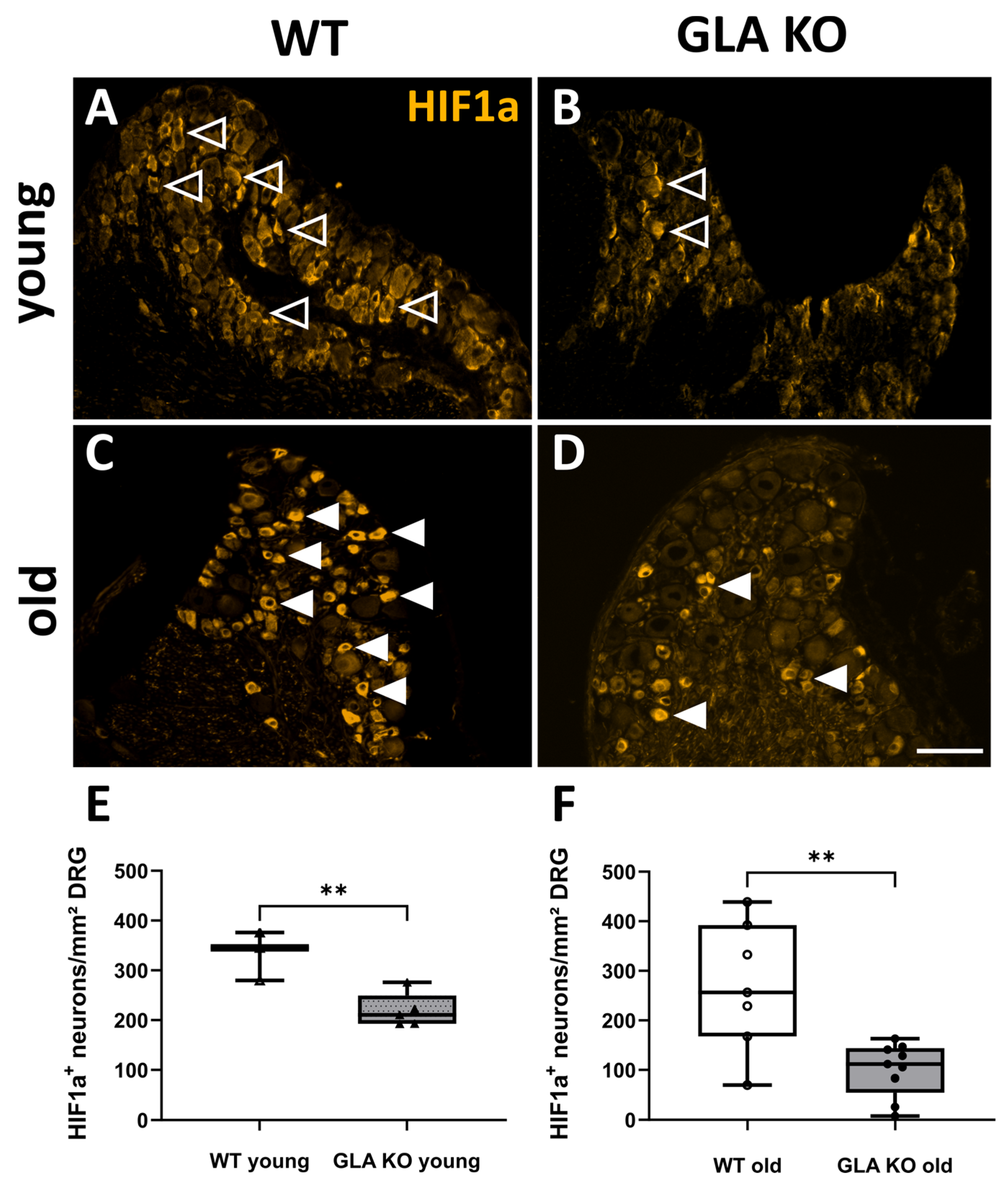
2.2.2. Increased Nuclear HIF1a Intensity in Old GLA KO DRG Compared to Old WT DRG
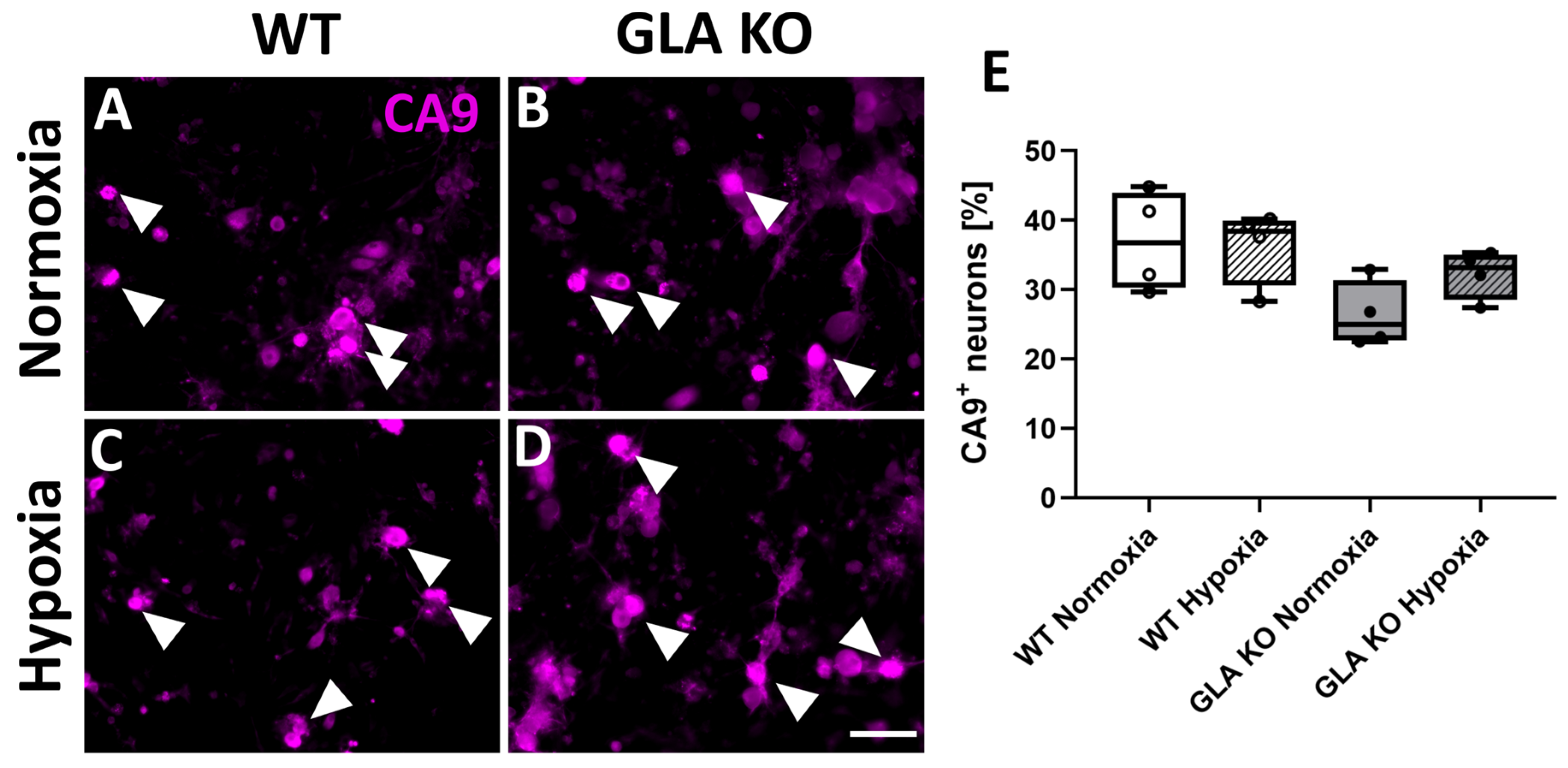
2.2.3. Higher Numbers of CA9+ DRG Neurons in Old GLA KO Mice Compared to Old WT Mice
2.2.4. Higher Expression Levels of Hypoxia-Associated Target Genes in Cultivated DRG Neurons of WT and GLA KO Mice under Hypoxia Compared to Normoxia
2.2.5. Similar HIF1a Protein Distribution in DRG Neurons of Old WT and GLA KO Mice Cultivated under Normoxia and Hypoxia
2.2.6. Similar Number of CA9+ DRG Neurons in WT and GLA KO Mice Cultivated under Normoxia and Hypoxia
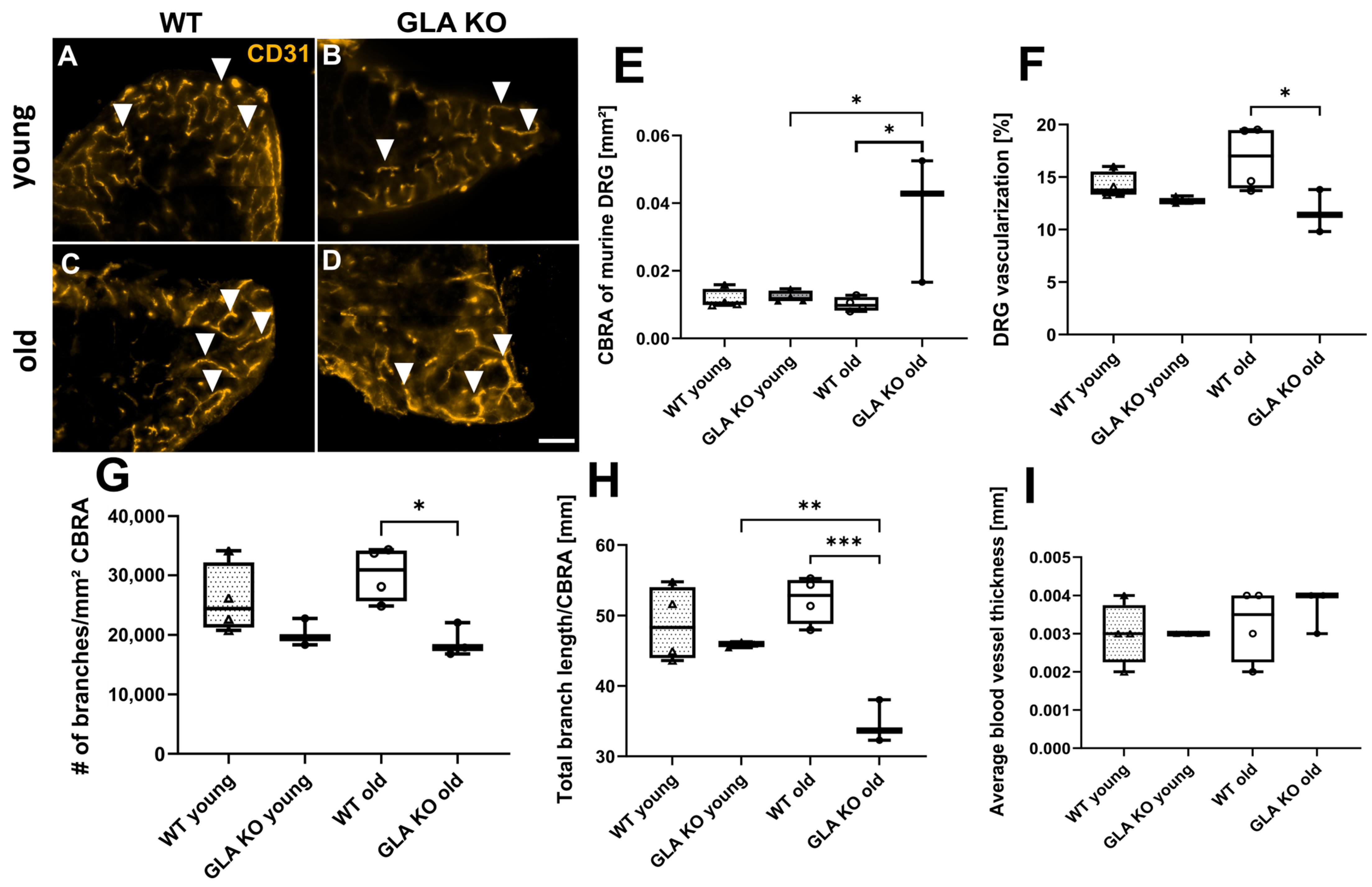
2.2.7. Reduced Vascularization in Old GLA KO Compared to Old WT Mice
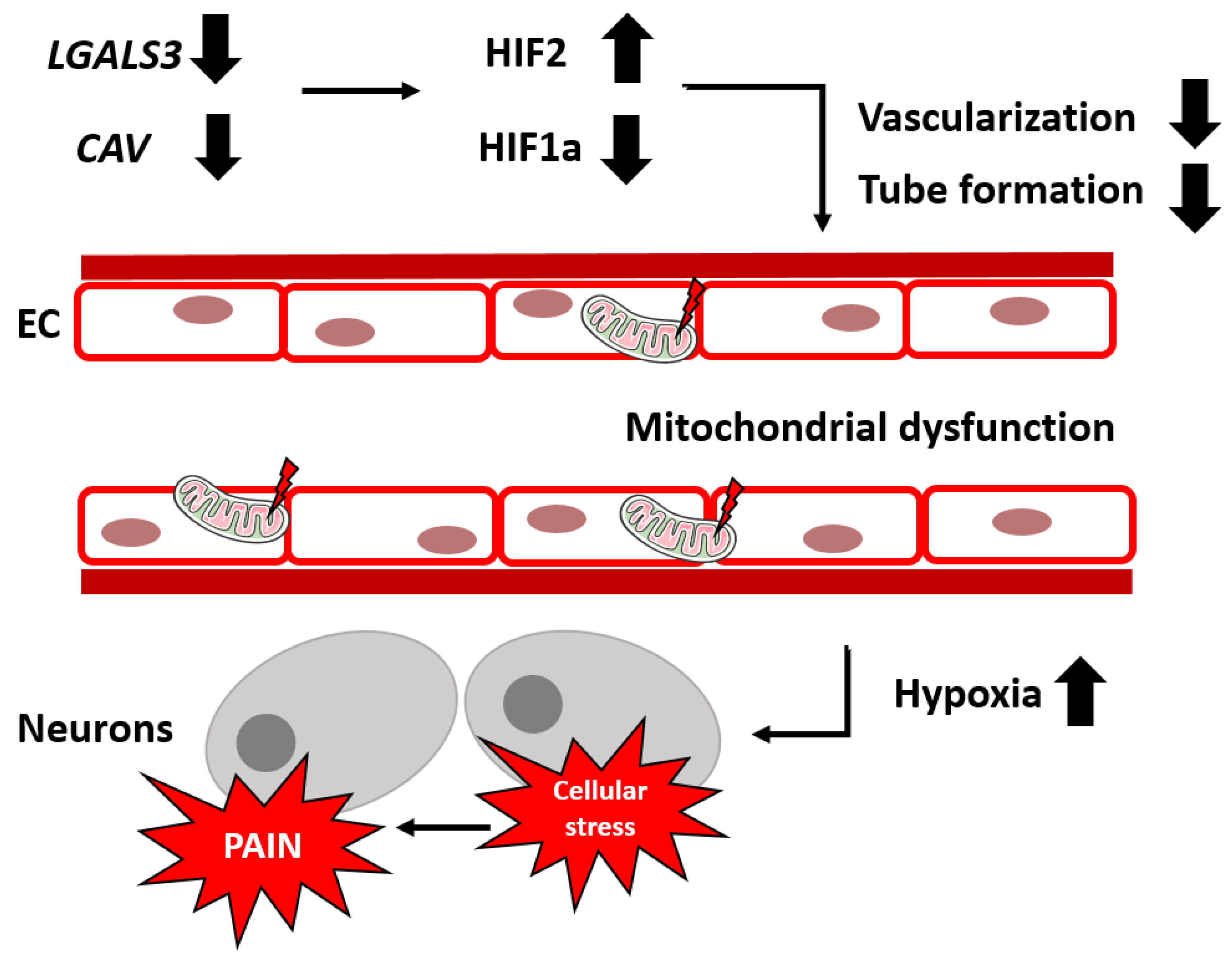
3. Discussion
4. Materials and Methods
4.1. Experiments on Human Biomaterial
4.1.1. Patients and Clinical Examination
4.1.2. Generation of Subject-Derived EC
4.1.3. Acetylated-Low-Density Lipoprotein (Ac-LDL) Uptake Assay
4.1.4. AGAL Enzyme Activity Assay
4.1.5. EC Tube Formation Assay
4.1.6. EC Cultivation under Hypoxic Conditions
4.1.7. Immunolabeling
4.1.8. Expansion Microscopy
4.1.9. Mitochondrial Morphology
4.1.10. Mitochondrial Membrane Potential
4.1.11. EC Gene Expression Analysis
4.2. Experiments on GLA KO Mouse Model
4.2.1. Mouse Colony
4.2.2. Tissue Collection
4.2.3. DRG Neuronal Cell Culture
4.2.4. In Vitro Hypoxia Experiments with DRG Neuronal Cell Cultures
4.2.5. Immunolabeling
4.2.6. Gene Expression Analysis
4.3. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Germain, D.P.; Levade, T.; Hachulla, E.; Knebelmann, B.; Lacombe, D.; Seguin, V.L.; Nguyen, K.; Noel, E.; Rabes, J.P. Challenging the traditional approach for interpreting genetic variants: Lessons from Fabry disease. Clin. Genet. 2022, 101, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Weissmann, C.; Albanese, A.A.; Contreras, N.E.; Gobetto, M.N.; Castellanos, L.C.S.; Uchitel, O.D. Ion channels and pain in Fabry disease. Mol. Pain. 2021, 17, 17448069211033172. [Google Scholar] [CrossRef] [PubMed]
- Do, H.S.; Park, S.W.; Im, I.; Seo, D.; Yoo, H.W.; Go, H.; Kim, Y.H.; Koh, G.Y.; Lee, B.H.; Han, Y.M. Enhanced thrombospondin-1 causes dysfunction of vascular endothelial cells derived from Fabry disease-induced pluripotent stem cells. EBioMedicine 2020, 52, 102633. [Google Scholar] [CrossRef] [PubMed]
- Pollmann, S.; Scharnetzki, D.; Manikowski, D.; Lenders, M.; Brand, E. Endothelial Dysfunction in Fabry Disease Is Related to Glycocalyx Degradation. Front. Immunol. 2021, 12, 789142. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.S.; Meng, X.L.; Schiffmann, R.; Brady, R.O.; Kaneski, C.R. Establishment and characterization of Fabry disease endothelial cells with an extended lifespan. Mol. Genet. Metab. 2007, 92, 137–144. [Google Scholar] [CrossRef]
- Ohshima, T.; Murray, G.J.; Swaim, W.D.; Longenecker, G.; Quirk, J.M.; Cardarelli, C.O.; Sugimoto, Y.; Pastan, I.; Gottesman, M.M.; Brady, R.O.; et al. alpha-Galactosidase A deficient mice: A model of Fabry disease. Proc. Natl. Acad. Sci. USA 1997, 94, 2540–2544. [Google Scholar] [CrossRef]
- Park, J.L.; Shu, L.; Shayman, J.A. Differential involvement of COX1 and COX2 in the vasculopathy associated with the alpha-galactosidase A-knockout mouse. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1133–H1140. [Google Scholar] [CrossRef]
- Park, J.L.; Whitesall, S.E.; D’Alecy, L.G.; Shu, L.; Shayman, J.A. Vascular dysfunction in the alpha-galactosidase A-knockout mouse is an endothelial cell-, plasma membrane-based defect. Clin. Exp. Pharmacol. Physiol. 2008, 35, 1156–1163. [Google Scholar] [CrossRef]
- Godel, T.; Baumer, P.; Pham, M.; Kohn, A.; Muschol, N.; Kronlage, M.; Kollmer, J.; Heiland, S.; Bendszus, M.; Mautner, V.F. Human dorsal root ganglion in vivo morphometry and perfusion in Fabry painful neuropathy. Neurology 2017, 89, 1274–1282. [Google Scholar] [CrossRef]
- Chen, F.W.; Davies, J.P.; Calvo, R.; Chaudhari, J.; Dolios, G.; Taylor, M.K.; Patnaik, S.; Dehdashti, J.; Mull, R.; Dranchack, P.; et al. Activation of mitochondrial TRAP1 stimulates mitochondria-lysosome crosstalk and correction of lysosomal dysfunction. iScience 2022, 25, 104941. [Google Scholar] [CrossRef]
- Gambardella, J.; Fiordelisi, A.; Sorriento, D.; Cerasuolo, F.; Buonaiuto, A.; Avvisato, R.; Pisani, A.; Varzideh, F.; Riccio, E.; Santulli, G.; et al. Mitochondrial microRNAs Are Dysregulated in Patients with Fabry Disease. J. Pharmacol. Exp. Ther. 2023, 384, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Schumann, A.; Schaller, K.; Belche, V.; Cybulla, M.; Grunert, S.C.; Moers, N.; Sass, J.O.; Kaech, A.; Hannibal, L.; Spiekerkoetter, U. Defective lysosomal storage in Fabry disease modifies mitochondrial structure, metabolism and turnover in renal epithelial cells. J. Inherit. Metab. Dis. 2021, 44, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Ivashkiv, L.B. The hypoxia-lactate axis tempers inflammation. Nat. Rev. Immunol. 2020, 20, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.T.; Colgan, S.P. Regulation of immunity and inflammation by hypoxia in immunological niches. Nat. Rev. Immunol. 2017, 17, 774–785. [Google Scholar] [CrossRef]
- Klein, T.; Gruener, J.; Breyer, M.; Schlegel, J.; Schottmann, N.M.; Hofmann, L.; Gauss, K.; Mease, R.; Erbacher, C.; Finke, L.; et al. Small fibre neuropathy in Fabry disease: A human-derived neuronal in vitro disease model. bioRxiv 2023. [Google Scholar] [CrossRef]
- Rohani, A.; Kashatus, J.A.; Sessions, D.T.; Sharmin, S.; Kashatus, D.F. Mito Hacker: A set of tools to enable high-throughput analysis of mitochondrial network morphology. Sci. Rep. 2020, 10, 18941. [Google Scholar] [CrossRef]
- Schito, L.; Rey, S. Hypoxia: Turning vessels into vassals of cancer immunotolerance. Cancer Lett. 2020, 487, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.; Johnson, R.S. Hypoxia: A key regulator of angiogenesis in cancer. Cancer Metastasis Rev. 2007, 26, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Biancini, G.B.; Vanzin, C.S.; Rodrigues, D.B.; Deon, M.; Ribas, G.S.; Barschak, A.G.; Manfredini, V.; Netto, C.B.; Jardim, L.B.; Giugliani, R.; et al. Globotriaosylceramide is correlated with oxidative stress and inflammation in Fabry patients treated with enzyme replacement therapy. Biochim. Biophys. Acta 2012, 1822, 226–232. [Google Scholar] [CrossRef]
- Chimenti, C.; Scopelliti, F.; Vulpis, E.; Tafani, M.; Villanova, L.; Verardo, R.; De Paulis, R.; Russo, M.A.; Frustaci, A. Increased oxidative stress contributes to cardiomyocyte dysfunction and death in patients with Fabry disease cardiomyopathy. Hum. Pathol. 2015, 46, 1760–1768. [Google Scholar] [CrossRef]
- Ho, K.H.; Chen, P.H.; Chou, C.M.; Shih, C.M.; Lee, Y.T.; Cheng, C.H.; Chen, K.C. A Key Role of DNA Damage-Inducible Transcript 4 (DDIT4) Connects Autophagy and GLUT3-Mediated Stemness To Desensitize Temozolomide Efficacy in Glioblastomas. Neurotherapeutics 2020, 17, 1212–1227. [Google Scholar] [CrossRef]
- Simonson, B.; Subramanya, V.; Chan, M.C.; Zhang, A.; Franchino, H.; Ottaviano, F.; Mishra, M.K.; Knight, A.C.; Hunt, D.; Ghiran, I.; et al. DDiT4L promotes autophagy and inhibits pathological cardiac hypertrophy in response to stress. Sci. Signal. 2017, 10, eaaf5967. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S. Role and mechanism of the action of angiopoietin-like protein ANGPTL4 in plasma lipid metabolism. J. Lipid Res. 2021, 62, 100150. [Google Scholar] [CrossRef]
- Sylvers-Davie, K.L.; Davies, B.S.J. Regulation of lipoprotein metabolism by ANGPTL3, ANGPTL4, and ANGPTL8. Am. J. Physiol. Endocrinol. Metab. 2021, 321, E493–E508. [Google Scholar] [CrossRef] [PubMed]
- Dijk, W.; Heine, M.; Vergnes, L.; Boon, M.R.; Schaart, G.; Hesselink, M.K.; Reue, K.; van Marken Lichtenbelt, W.D.; Olivecrona, G.; Rensen, P.C.; et al. ANGPTL4 mediates shuttling of lipid fuel to brown adipose tissue during sustained cold exposure. Elife 2015, 4, e08428. [Google Scholar] [CrossRef]
- Garcia-Ponce, A.; Chanez Paredes, S.; Castro Ochoa, K.F.; Schnoor, M. Regulation of endothelial and epithelial barrier functions by peptide hormones of the adrenomedullin family. Tissue Barriers 2016, 4, e1228439. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, J.M.; Dietrich, B.; Fiedler, J.; Jazbutyte, V.; Fleissner, F.; Karpinski, N.; Weidemann, F.; Wanner, C.; Asan, E.; Caprio, M.; et al. Pathologic endothelial response and impaired function of circulating angiogenic cells in patients with Fabry disease. Basic. Res. Cardiol. 2013, 108, 311. [Google Scholar] [CrossRef]
- van Lier, D.; Kox, M.; Pickkers, P. Promotion of vascular integrity in sepsis through modulation of bioactive adrenomedullin and dipeptidyl peptidase 3. J. Intern. Med. 2021, 289, 792–806. [Google Scholar] [CrossRef]
- Wu, X.; Zheng, W.; Jin, P.; Hu, J.; Zhou, Q. Role of IGFBP1 in the senescence of vascular endothelial cells and severity of aging-related coronary atherosclerosis. Int. J. Mol. Med. 2019, 44, 1921–1931. [Google Scholar] [CrossRef]
- Lin, Q.; Cong, X.; Yun, Z. Differential hypoxic regulation of hypoxia-inducible factors 1alpha and 2alpha. Mol. Cancer Res. 2011, 9, 757–765. [Google Scholar] [CrossRef]
- Reiterer, M.; Colaco, R.; Emrouznejad, P.; Jensen, A.; Rundqvist, H.; Johnson, R.S.; Branco, C. Acute and chronic hypoxia differentially predispose lungs for metastases. Sci. Rep. 2019, 9, 10246. [Google Scholar] [CrossRef] [PubMed]
- Saxena, K.; Jolly, M.K. Acute vs. Chronic vs. Cyclic Hypoxia: Their Differential Dynamics, Molecular Mechanisms, and Effects on Tumor Progression. Biomolecules 2019, 9, 339. [Google Scholar] [CrossRef] [PubMed]
- Biancini, G.B.; Moras, A.M.; Reinhardt, L.S.; Busatto, F.F.; de Moura Sperotto, N.D.; Saffi, J.; Moura, D.J.; Giugliani, R.; Vargas, C.R. Globotriaosylsphingosine induces oxidative DNA damage in cultured kidney cells. Nephrology 2017, 22, 490–493. [Google Scholar] [CrossRef]
- Biancini, G.B.; Moura, D.J.; Manini, P.R.; Faverzani, J.L.; Netto, C.B.; Deon, M.; Giugliani, R.; Saffi, J.; Vargas, C.R. DNA damage in Fabry patients: An investigation of oxidative damage and repair. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2015, 784–785, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Atiskova, Y.; Wildner, J.; Spitzer, M.S.; Aries, C.; Muschol, N.; Dulz, S. Retinal vessel tortuosity as a prognostic marker for disease severity in Fabry disease. Orphanet J. Rare Dis. 2021, 16, 485. [Google Scholar] [CrossRef]
- Mishra, V.; Banerjee, A.; Gandhi, A.B.; Kaleem, I.; Alexander, J.; Hisbulla, M.; Kannichamy, V.; Valaiyaduppu Subas, S.; Hamid, P. Stroke and Fabry Disease: A Review of Literature. Cureus 2020, 12, e12083. [Google Scholar] [CrossRef]
- Tapia, D.; Floriolli, D.; Han, E.; Lee, G.; Paganini-Hill, A.; Wang, S.; Zandihaghighi, S.; Kimonis, V.; Fisher, M. Prevalence of cerebral small vessel disease in a Fabry disease cohort. Mol. Genet. Metab. Rep. 2021, 29, 100815. [Google Scholar] [CrossRef]
- Choi, J.B.; Seol, D.W.; Do, H.S.; Yang, H.Y.; Kim, T.M.; Byun, Y.G.; Park, J.M.; Choi, J.; Hong, S.P.; Chung, W.S.; et al. Fasudil alleviates the vascular endothelial dysfunction and several phenotypes of Fabry disease. Mol. Ther. 2023, 31, 1002–1016. [Google Scholar] [CrossRef]
- Ivanova, M.M.; Dao, J.; Slayeh, O.A.; Friedman, A.; Goker-Alpan, O. Circulated TGF-beta1 and VEGF-A as Biomarkers for Fabry Disease-Associated Cardiomyopathy. Cells 2023, 12, 2102. [Google Scholar] [CrossRef]
- Sorriento, D.; Iaccarino, G. The Cardiovascular Phenotype in Fabry Disease: New Findings in the Research Field. Int. J. Mol. Sci. 2021, 22, 1331. [Google Scholar] [CrossRef]
- Tebani, A.; Mauhin, W.; Abily-Donval, L.; Lesueur, C.; Berger, M.G.; Nadjar, Y.; Berger, J.; Benveniste, O.; Lamari, F.; Laforet, P.; et al. A Proteomics-Based Analysis Reveals Predictive Biological Patterns in Fabry Disease. J. Clin. Med. 2020, 9, 1325. [Google Scholar] [CrossRef]
- Zampetti, A.; Gnarra, M.; Borsini, W.; Giurdanella, F.; Antuzzi, D.; Piras, A.; Smaldone, C.; Pieroni, M.; Cadeddu, C.; de Waure, C.; et al. Vascular endothelial growth factor (VEGF-a) in Fabry disease: Association with cutaneous and systemic manifestations with vascular involvement. Cytokine 2013, 61, 933–939. [Google Scholar] [CrossRef]
- Lee, M.H.; Choi, E.N.; Jeon, Y.J.; Jung, S.C. Possible role of transforming growth factor-beta1 and vascular endothelial growth factor in Fabry disease nephropathy. Int. J. Mol. Med. 2012, 30, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- DaCosta Byfield, S.; Major, C.; Laping, N.J.; Roberts, A.B. SB-505124 is a selective inhibitor of transforming growth factor-beta type I receptors ALK4, ALK5, and ALK7. Mol. Pharmacol. 2004, 65, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Yang, M.; Zhao, H.; Han, Y.; Jiang, N.; Yang, J.; Chen, W.; Li, C.; Liu, Y.; Zhao, C.; et al. Caveolin-1 Regulates Cellular Metabolism: A Potential Therapeutic Target in Kidney Disease. Front. Pharmacol. 2021, 12, 768100. [Google Scholar] [CrossRef]
- Shu, L.; Shayman, J.A. Caveolin-associated accumulation of globotriaosylceramide in the vascular endothelium of alpha-galactosidase A null mice. J. Biol. Chem. 2007, 282, 20960–20967. [Google Scholar] [CrossRef]
- Monticelli, M.; Liguori, L.; Allocca, M.; Bosso, A.; Andreotti, G.; Lukas, J.; Monti, M.C.; Morretta, E.; Cubellis, M.V.; Hay Mele, B. Drug Repositioning for Fabry Disease: Acetylsalicylic Acid Potentiates the Stabilization of Lysosomal Alpha-Galactosidase by Pharmacological Chaperones. Int. J. Mol. Sci. 2022, 23, 5105. [Google Scholar] [CrossRef] [PubMed]
- Elsaid, H.O.A.; Rivedal, M.; Skandalou, E.; Svarstad, E.; Tondel, C.; Birkeland, E.; Eikrem, O.; Babickova, J.; Marti, H.P.; Furriol, J. Proteomic analysis unveils Gb3-independent alterations and mitochondrial dysfunction in a gla(-/-) zebrafish model of Fabry disease. J. Transl. Med. 2023, 21, 591. [Google Scholar] [CrossRef]
- Adebayo, M.; Singh, S.; Singh, A.P.; Dasgupta, S. Mitochondrial fusion and fission: The fine-tune balance for cellular homeostasis. FASEB J. 2021, 35, e21620. [Google Scholar] [CrossRef]
- Vasquez-Trincado, C.; Garcia-Carvajal, I.; Pennanen, C.; Parra, V.; Hill, J.A.; Rothermel, B.A.; Lavandero, S. Mitochondrial dynamics, mitophagy and cardiovascular disease. J. Physiol. 2016, 594, 509–525. [Google Scholar] [CrossRef]
- Youle, R.J.; van der Bliek, A.M. Mitochondrial fission, fusion, and stress. Science 2012, 337, 1062–1065. [Google Scholar] [CrossRef]
- Qu, K.; Yan, F.; Qin, X.; Zhang, K.; He, W.; Dong, M.; Wu, G. Mitochondrial dysfunction in vascular endothelial cells and its role in atherosclerosis. Front. Physiol. 2022, 13, 1084604. [Google Scholar] [CrossRef]
- Dong, F.; Zhu, M.; Zheng, F.; Fu, C. Mitochondrial fusion and fission are required for proper mitochondrial function and cell proliferation in fission yeast. FEBS J. 2022, 289, 262–278. [Google Scholar] [CrossRef] [PubMed]
- Knights, A.J.; Yik, J.J.; Mat Jusoh, H.; Norton, L.J.; Funnell, A.P.; Pearson, R.C.; Bell-Anderson, K.S.; Crossley, M.; Quinlan, K.G. Kruppel-like Factor 3 (KLF3/BKLF) Is Required for Widespread Repression of the Inflammatory Modulator Galectin-3 (Lgals3). J. Biol. Chem. 2016, 291, 16048–16058. [Google Scholar] [CrossRef]
- Pierzynowska, K.; Rintz, E.; Gaffke, L.; Wegrzyn, G. Ferroptosis and Its Modulation by Autophagy in Light of the Pathogenesis of Lysosomal Storage Diseases. Cells 2021, 10, 365. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Berdini, L.; Ferrero, G.O.; Bustos Plonka, F.; Cardozo Gizzi, A.M.; Prucca, C.G.; Quiroga, S.; Caputto, B.L. The moonlighting protein c-Fos activates lipid synthesis in neurons, an activity that is critical for cellular differentiation and cortical development. J. Biol. Chem. 2020, 295, 8808–8818. [Google Scholar] [CrossRef]
- Da Vitoria Lobo, M.E.; Weir, N.; Hardowar, L.; Al Ojaimi, Y.; Madden, R.; Gibson, A.; Bestall, S.M.; Hirashima, M.; Schaffer, C.B.; Donaldson, L.F.; et al. Hypoxia-induced carbonic anhydrase mediated dorsal horn neuron activation and induction of neuropathic pain. Pain 2022, 163, 2264–2279. [Google Scholar] [CrossRef] [PubMed]
- Watzky, M.; Huard, S.; Juricek, L.; Dairou, J.; Chauvet, C.; Coumoul, X.; Letessier, A.; Miotto, B. Hexokinase 2 is a transcriptional target and a positive modulator of AHR signalling. Nucleic Acids Res. 2022, 50, 5545–5564. [Google Scholar] [CrossRef]
- Burbidge, K.; Rademacher, D.J.; Mattick, J.; Zack, S.; Grillini, A.; Bousset, L.; Kwon, O.; Kubicki, K.; Simon, A.; Melki, R.; et al. LGALS3 (galectin 3) mediates an unconventional secretion of SNCA/alpha-synuclein in response to lysosomal membrane damage by the autophagic-lysosomal pathway in human midbrain dopamine neurons. Autophagy 2022, 18, 1020–1048. [Google Scholar] [CrossRef]
- Hofmann, L.; Hose, D.; Griesshammer, A.; Blum, R.; Doring, F.; Dib-Hajj, S.; Waxman, S.; Sommer, C.; Wischmeyer, E.; Uceyler, N. Characterization of small fiber pathology in a mouse model of Fabry disease. Elife 2018, 7, e39300. [Google Scholar] [CrossRef]
- Le Moine, C.M.; Morash, A.J.; McClelland, G.B. Changes in HIF-1alpha protein, pyruvate dehydrogenase phosphorylation, and activity with exercise in acute and chronic hypoxia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1098–R1104. [Google Scholar] [CrossRef] [PubMed]
- Burand, A.J., Jr.; Stucky, C.L. Fabry disease pain: Patient and preclinical parallels. Pain 2021, 162, 1305–1321. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, M.; Ma, Z.; Yuan, D.; Zhu, J.; Tuo, B.; Li, T.; Liu, X. Alteration and dysfunction of ion channels/transporters in a hypoxic microenvironment results in the development and progression of gastric cancer. Cell. Oncol. 2021, 44, 739–749. [Google Scholar] [CrossRef]
- Miyake, T.; Nakamura, S.; Zhao, M.; So, K.; Inoue, K.; Numata, T.; Takahashi, N.; Shirakawa, H.; Mori, Y.; Nakagawa, T.; et al. Cold sensitivity of TRPA1 is unveiled by the prolyl hydroxylation blockade-induced sensitization to ROS. Nat. Commun. 2016, 7, 12840. [Google Scholar] [CrossRef]
- Tran, H.; Gupta, M.; Gupta, K. Targeting novel mechanisms of pain in sickle cell disease. Blood 2017, 130, 2377–2385. [Google Scholar] [CrossRef] [PubMed]
- Lakoma, J.; Rimondini, R.; Ferrer Montiel, A.; Donadio, V.; Liguori, R.; Caprini, M. Increased expression of Trpv1 in peripheral terminals mediates thermal nociception in Fabry disease mouse model. Mol. Pain. 2016, 12, 1744806916663729. [Google Scholar] [CrossRef]
- Namer, B.; Orstavik, K.; Schmidt, R.; Mair, N.; Kleggetveit, I.P.; Zeidler, M.; Martha, T.; Jorum, E.; Schmelz, M.; Kalpachidou, T.; et al. Changes in Ionic Conductance Signature of Nociceptive Neurons Underlying Fabry Disease Phenotype. Front. Neurol. 2017, 8, 335. [Google Scholar] [CrossRef] [PubMed]
- Spitzel, M.; Wagner, E.; Breyer, M.; Henniger, D.; Bayin, M.; Hofmann, L.; Mauceri, D.; Sommer, C.; Uceyler, N. Dysregulation of Immune Response Mediators and Pain-Related Ion Channels Is Associated with Pain-like Behavior in the GLA KO Mouse Model of Fabry Disease. Cells 2022, 11, 1730. [Google Scholar] [CrossRef] [PubMed]
- Beasley, N.J.; Wykoff, C.C.; Watson, P.H.; Leek, R.; Turley, H.; Gatter, K.; Pastorek, J.; Cox, G.J.; Ratcliffe, P.; Harris, A.L. Carbonic anhydrase IX, an endogenous hypoxia marker, expression in head and neck squamous cell carcinoma and its relationship to hypoxia, necrosis, and microvessel density. Cancer Res. 2001, 61, 5262–5267. [Google Scholar]
- Shin, K.H.; Diaz-Gonzalez, J.A.; Russell, J.; Chen, Q.; Burgman, P.; Li, X.F.; Ling, C.C. Detecting changes in tumor hypoxia with carbonic anhydrase IX and pimonidazole. Cancer Biol. Ther. 2007, 6, 70–75. [Google Scholar] [CrossRef]
- Mizukami, Y.; Kohgo, Y.; Chung, D.C. Hypoxia inducible factor-1 independent pathways in tumor angiogenesis. Clin. Cancer Res. 2007, 13, 5670–5674. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Li, L.; Liu, X.; Feng, Q.; Zhang, Y.; Zheng, P.; Cui, N. Hexokinases 2 promoted cell motility and distant metastasis by elevating fibronectin through Akt1/p-Akt1 in cervical cancer cells. Cancer Cell Int. 2021, 21, 600. [Google Scholar] [CrossRef] [PubMed]
- Seretny, M.; Colvin, L.A. Pain management in patients with vascular disease. Br. J. Anaesth. 2016, 117 (Suppl. S2), ii95–ii106. [Google Scholar] [CrossRef]
- Uceyler, N.; Magg, B.; Thomas, P.; Wiedmann, S.; Heuschmann, P.; Sommer, C. A comprehensive Fabry-related pain questionnaire for adult patients. Pain 2014, 155, 2301–2305. [Google Scholar] [CrossRef] [PubMed]
- Karl, F.; Wussmann, M.; Kress, L.; Malzacher, T.; Fey, P.; Groeber-Becker, F.; Uceyler, N. Patient-derived in vitro skin models for investigation of small fiber pathology. Ann. Clin. Transl. Neurol. 2019, 6, 1797–1806. [Google Scholar] [CrossRef] [PubMed]
- Gu, M. Efficient Differentiation of Human Pluripotent Stem Cells to Endothelial Cells. Curr. Protoc. Hum. Genet. 2018, 98, e64. [Google Scholar] [CrossRef] [PubMed]
- Üçeyler, N.; Biko, L.; Hose, D.; Hofmann, L.; Sommer, C. Comprehensive and differential long-term characterization of the alpha-galactosidase A deficient mouse model of Fabry disease focusing on the sensory system and pain development. Mol. Pain 2016, 12, 1744806916646379. [Google Scholar] [CrossRef]
- Sleigh, J.N.; Weir, G.A.; Schiavo, G. A simple, step-by-step dissection protocol for the rapid isolation of mouse dorsal root ganglia. BMC Res. Notes 2016, 9, 82. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P1 | P2 | Ctrl | |
|---|---|---|---|
| Age | 28 | 18 | 59 |
| Sex | M | M | M |
| Genotype | c.1069C > T//p.Q357X | c.568delG//p.A190Pfs*2 | NA |
| FD-associated pain character | Attacks | No pain | NA |
| Cardiomyopathy | Yes | No | NA |
| Nephropathy | No | No | NA |
| FD-specific treatment | Agalsidase-β | None | NA |
| AGAL activity | 0.5 (ref.: 3.4–13.0 nmol/h/mL) | 0.04 (ref.: 0.4–1.0 nmol/min/mg/protein) | NA |
| Lyso-Gb3 | 57.7 (ref.: <0.9 ng/mL) | 241 (ref.: <20.1 ng/mL) | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klug, K.; Spitzel, M.; Hans, C.; Klein, A.; Schottmann, N.M.; Erbacher, C.; Üçeyler, N. Endothelial Cell Dysfunction and Hypoxia as Potential Mediators of Pain in Fabry Disease: A Human-Murine Translational Approach. Int. J. Mol. Sci. 2023, 24, 15422. https://doi.org/10.3390/ijms242015422
Klug K, Spitzel M, Hans C, Klein A, Schottmann NM, Erbacher C, Üçeyler N. Endothelial Cell Dysfunction and Hypoxia as Potential Mediators of Pain in Fabry Disease: A Human-Murine Translational Approach. International Journal of Molecular Sciences. 2023; 24(20):15422. https://doi.org/10.3390/ijms242015422
Chicago/Turabian StyleKlug, Katharina, Marlene Spitzel, Clara Hans, Alexandra Klein, Nicole Michelle Schottmann, Christoph Erbacher, and Nurcan Üçeyler. 2023. "Endothelial Cell Dysfunction and Hypoxia as Potential Mediators of Pain in Fabry Disease: A Human-Murine Translational Approach" International Journal of Molecular Sciences 24, no. 20: 15422. https://doi.org/10.3390/ijms242015422
APA StyleKlug, K., Spitzel, M., Hans, C., Klein, A., Schottmann, N. M., Erbacher, C., & Üçeyler, N. (2023). Endothelial Cell Dysfunction and Hypoxia as Potential Mediators of Pain in Fabry Disease: A Human-Murine Translational Approach. International Journal of Molecular Sciences, 24(20), 15422. https://doi.org/10.3390/ijms242015422