
Ubiquitin Proteasome System Role in Diabetes-Induced Cardiomyopathy

 ,
,
Abstract
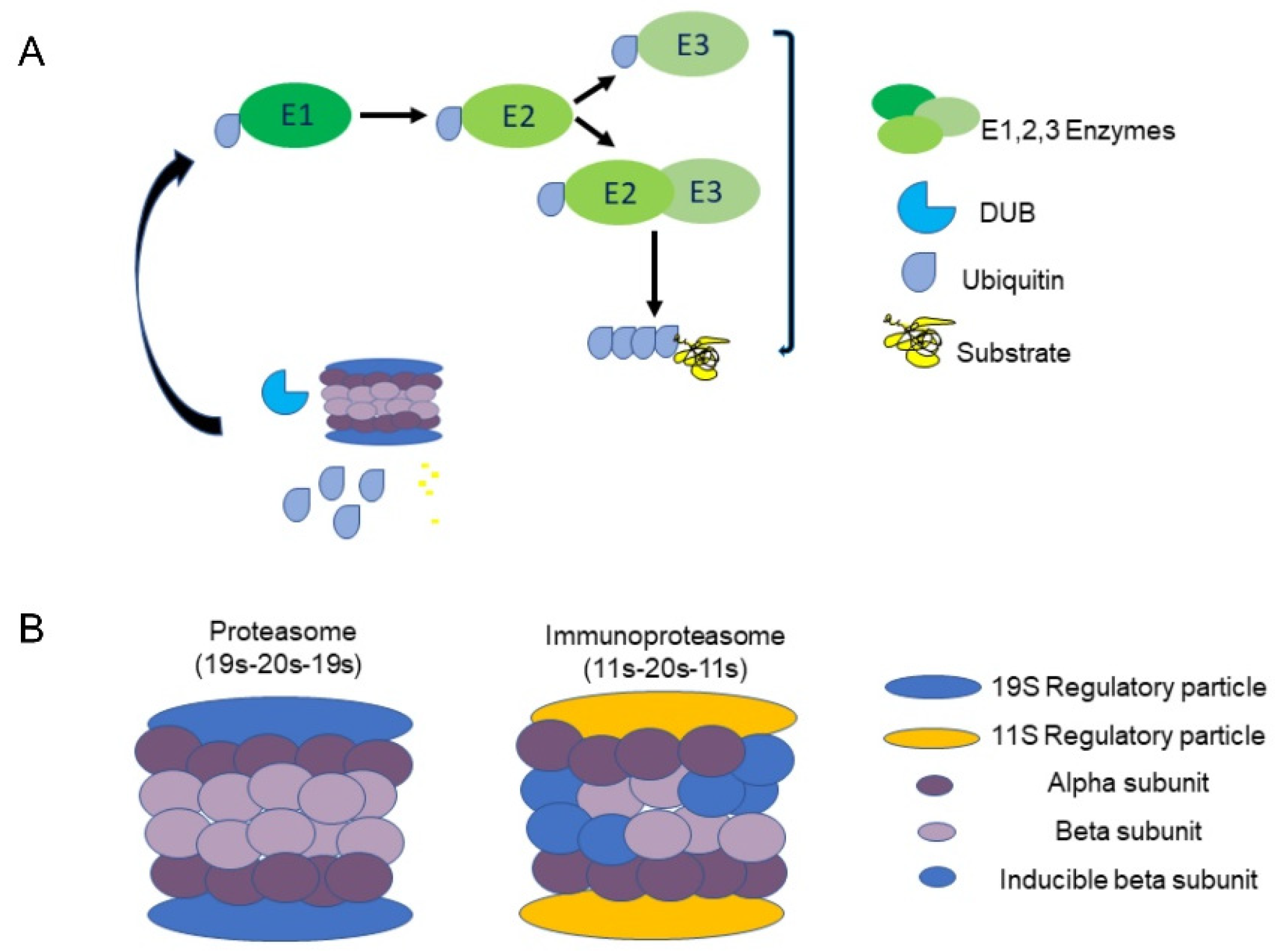
:1. Introduction
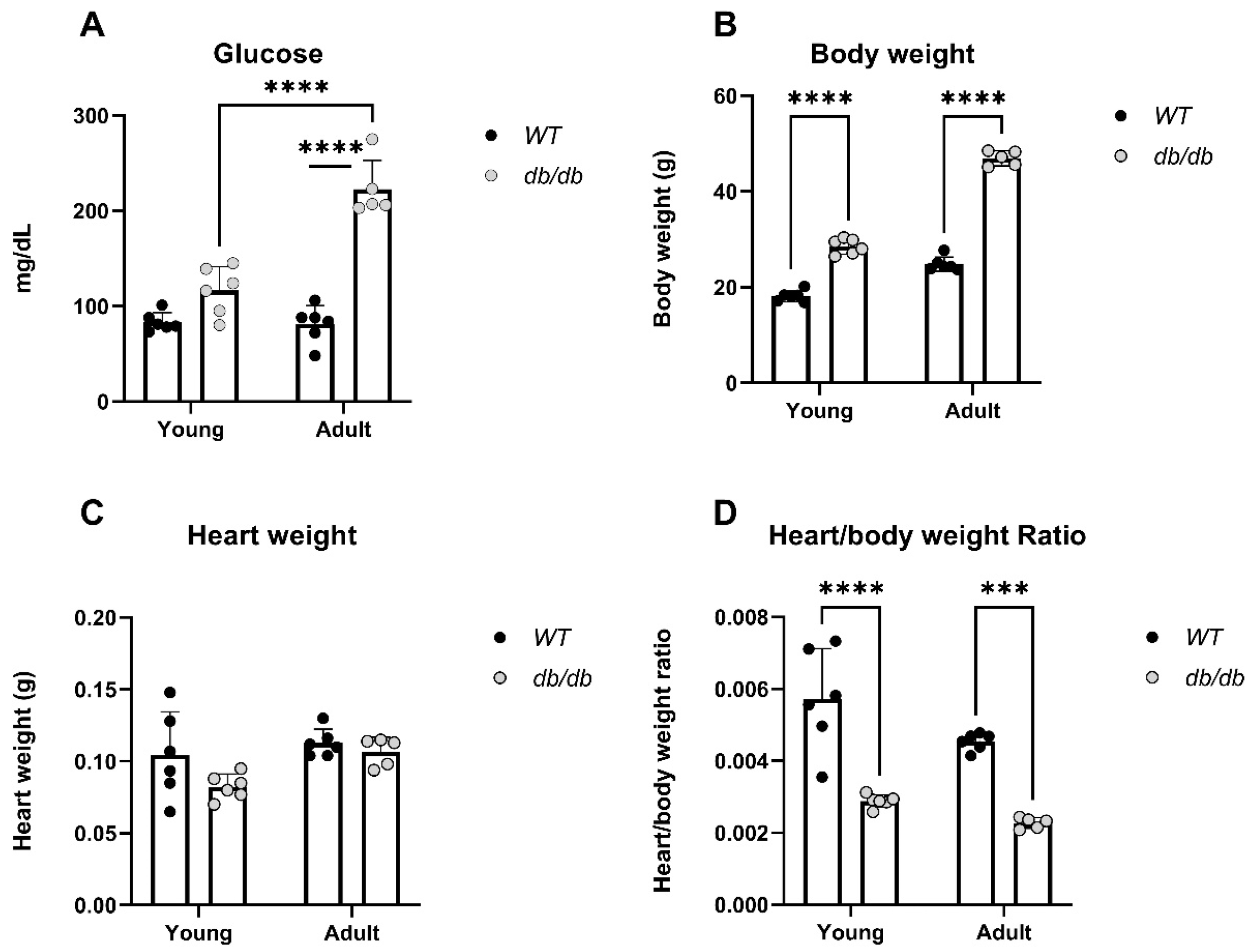
2. Results
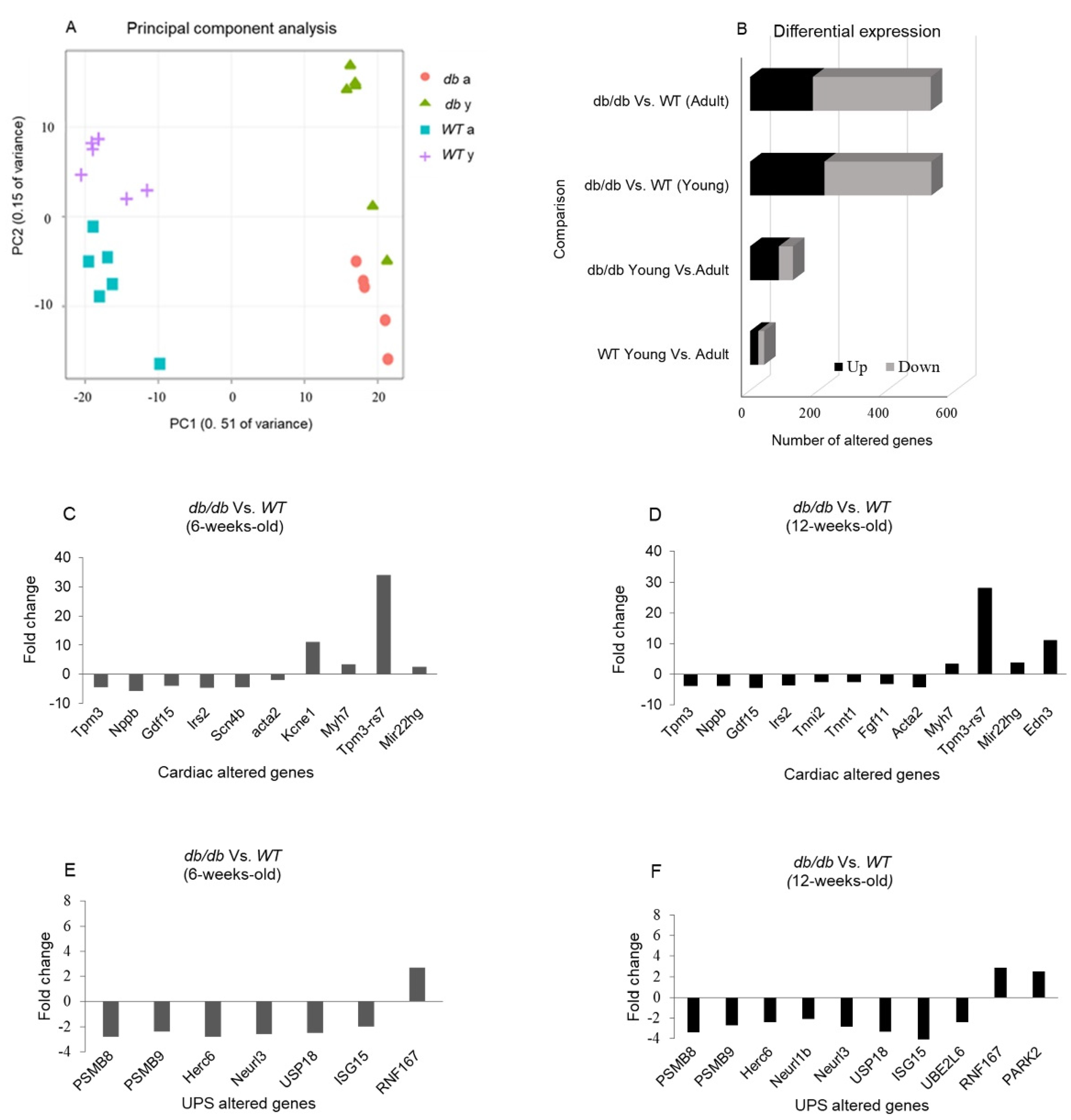
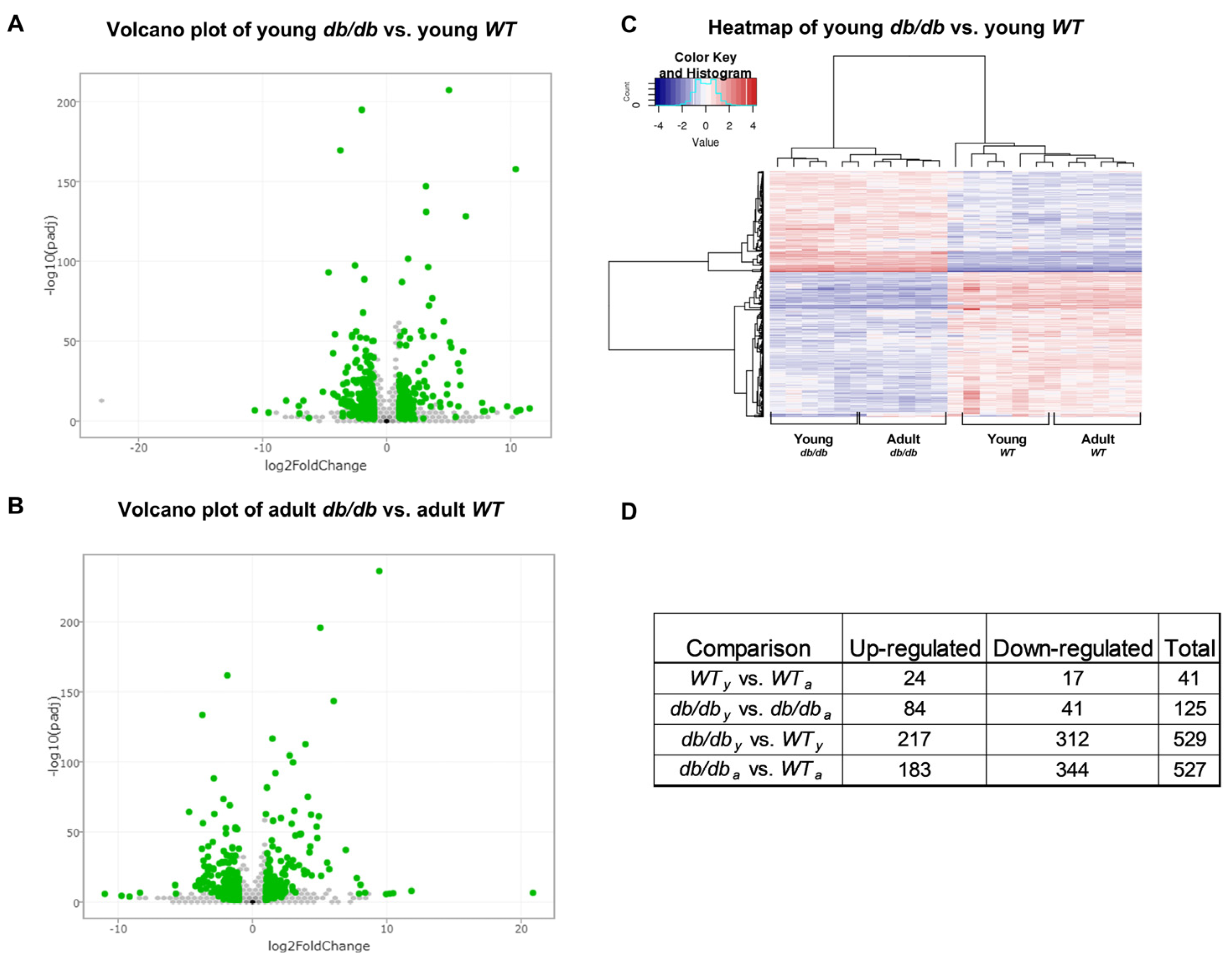
2.1. RNA Sequencing Emphasizes the Differences between db/db and WT Mice
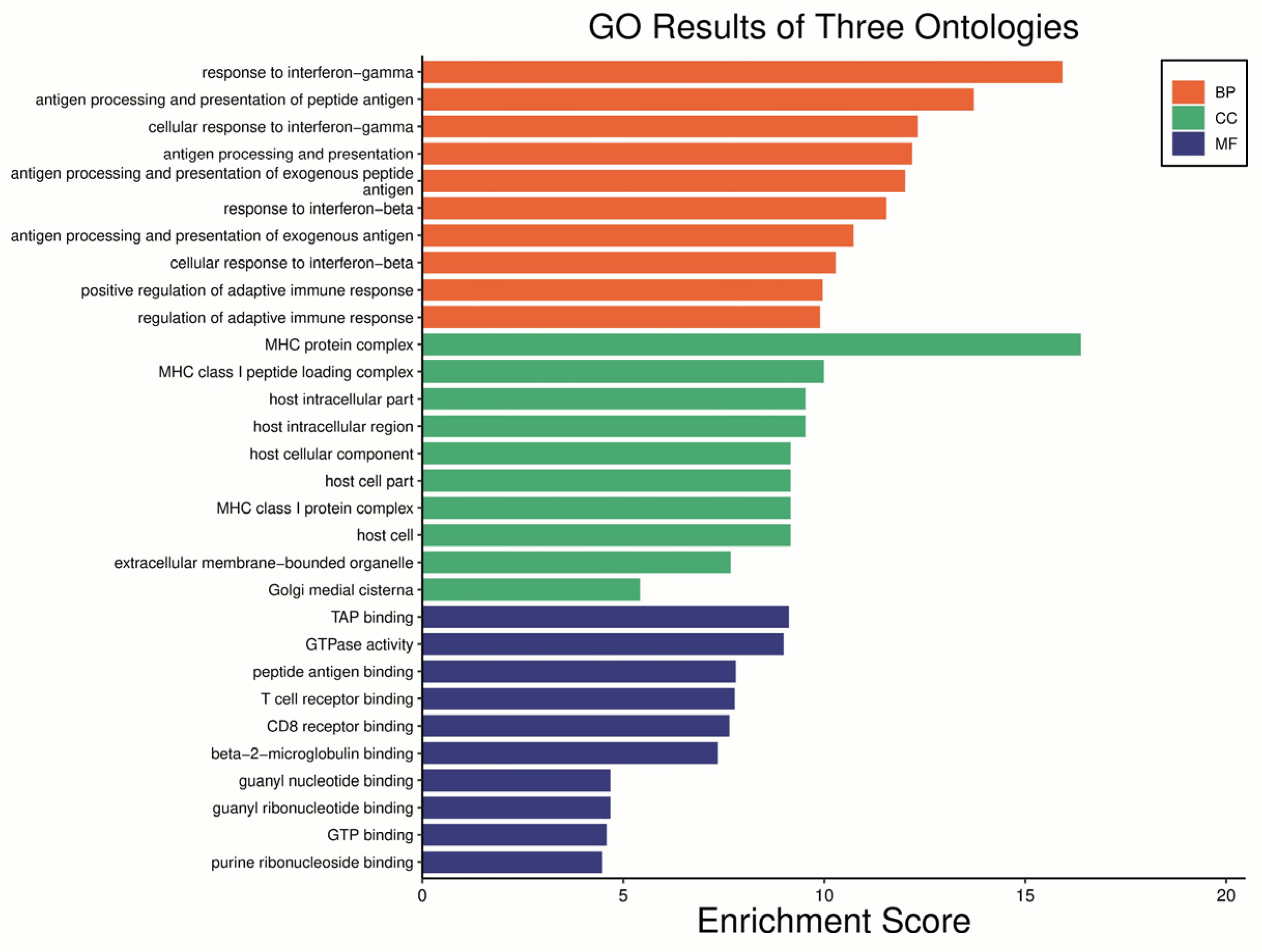
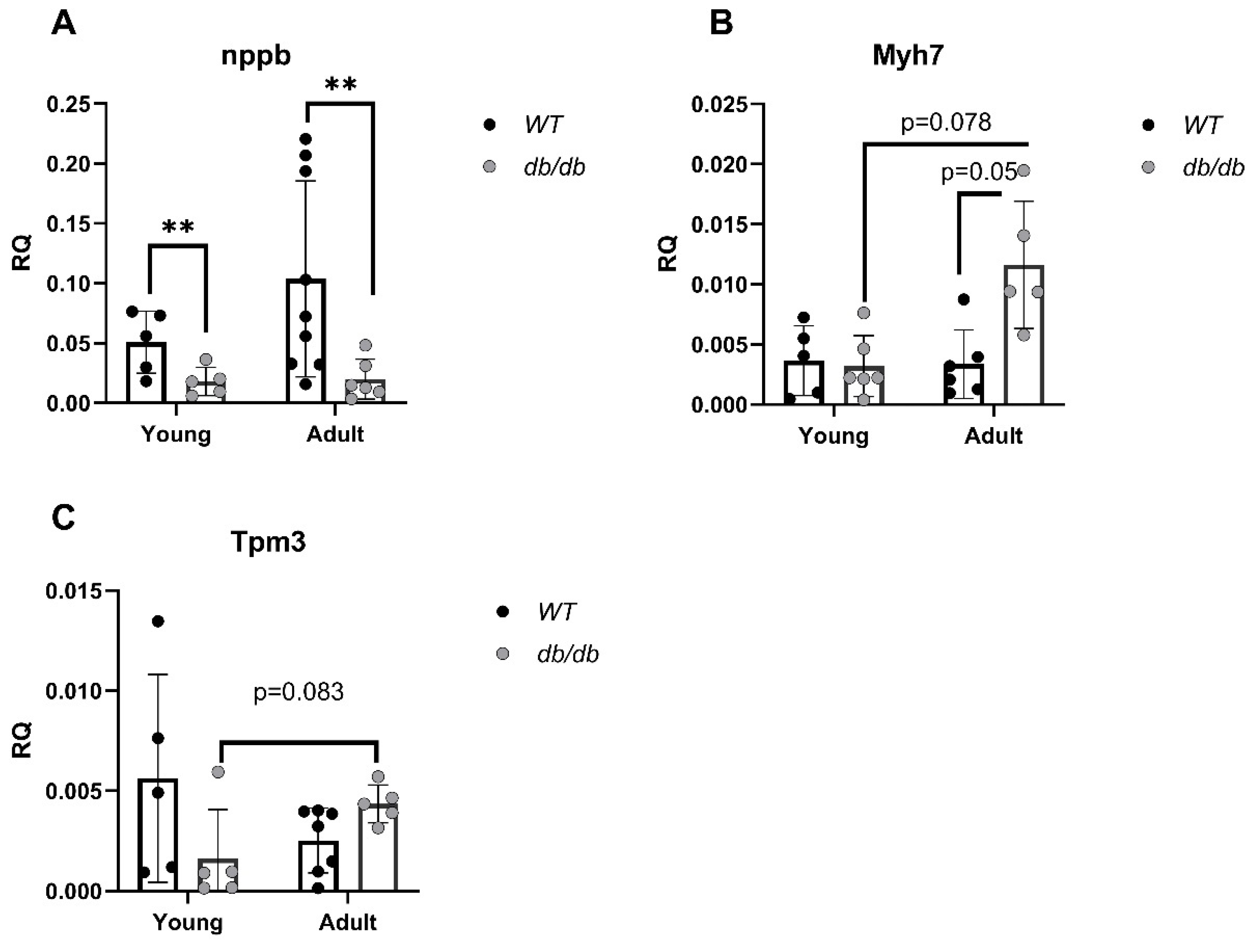
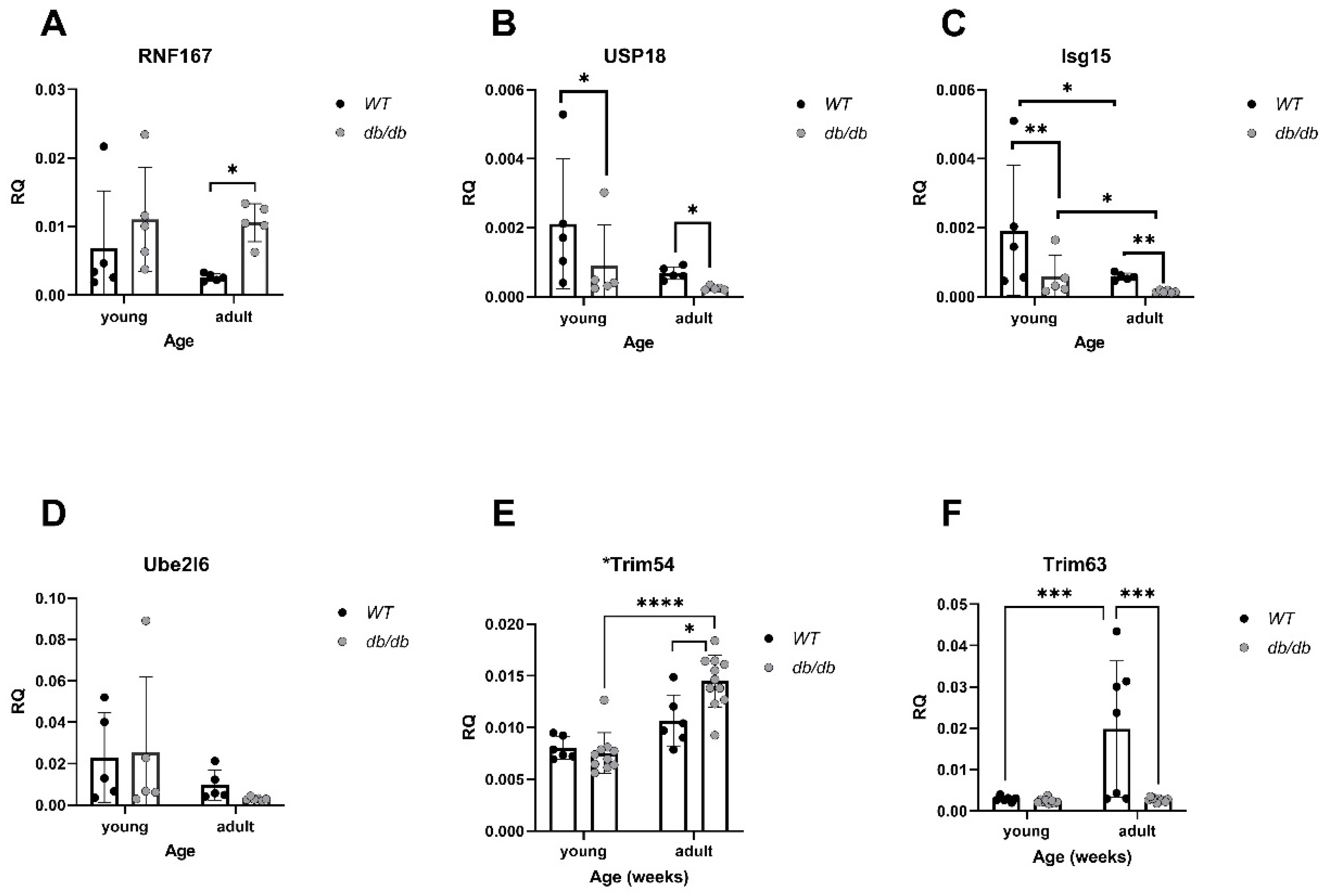
2.2. Cardiomyopathy- and UPS-Related Genes Are Differently Expressed in db/db Compared with WT Mice
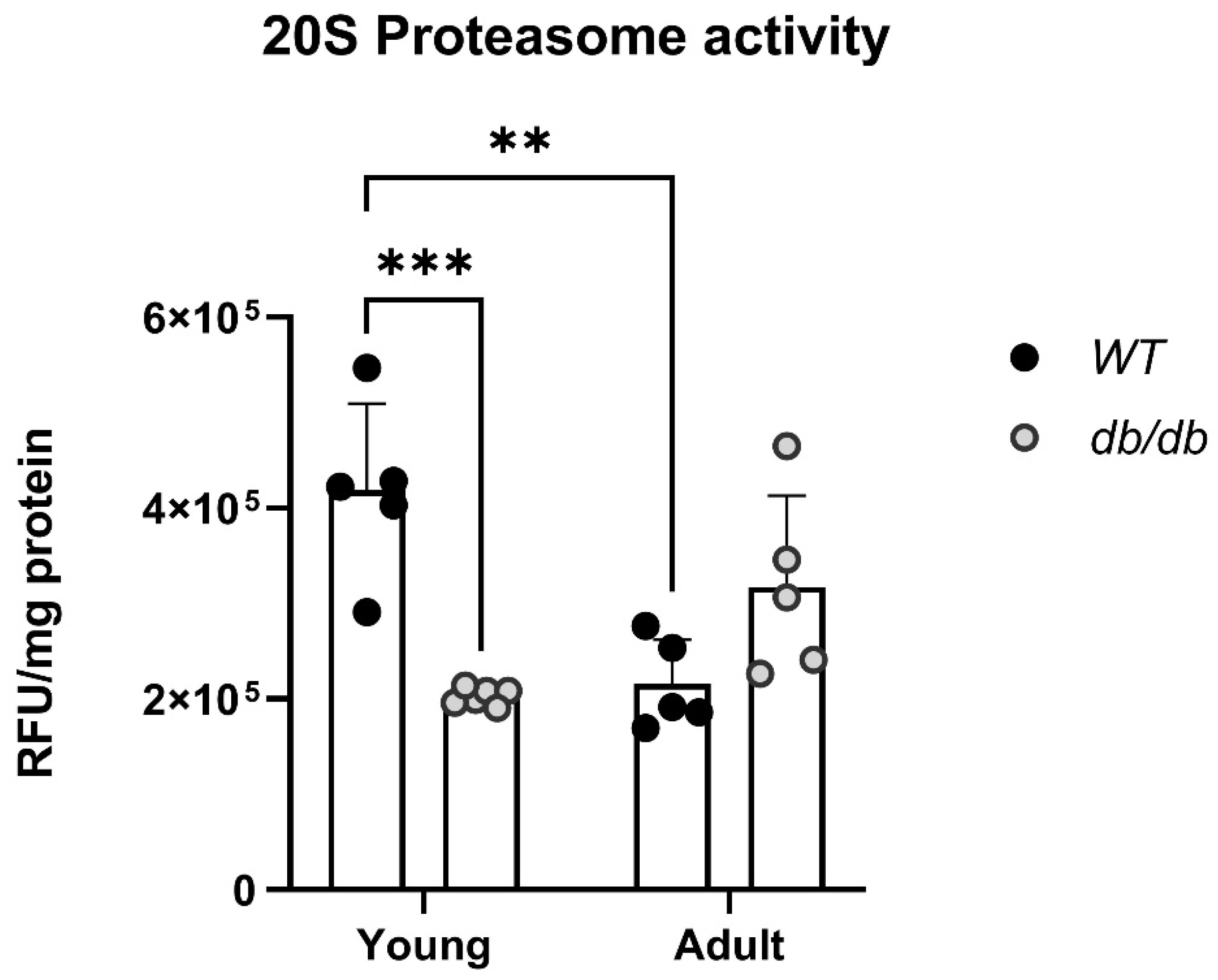
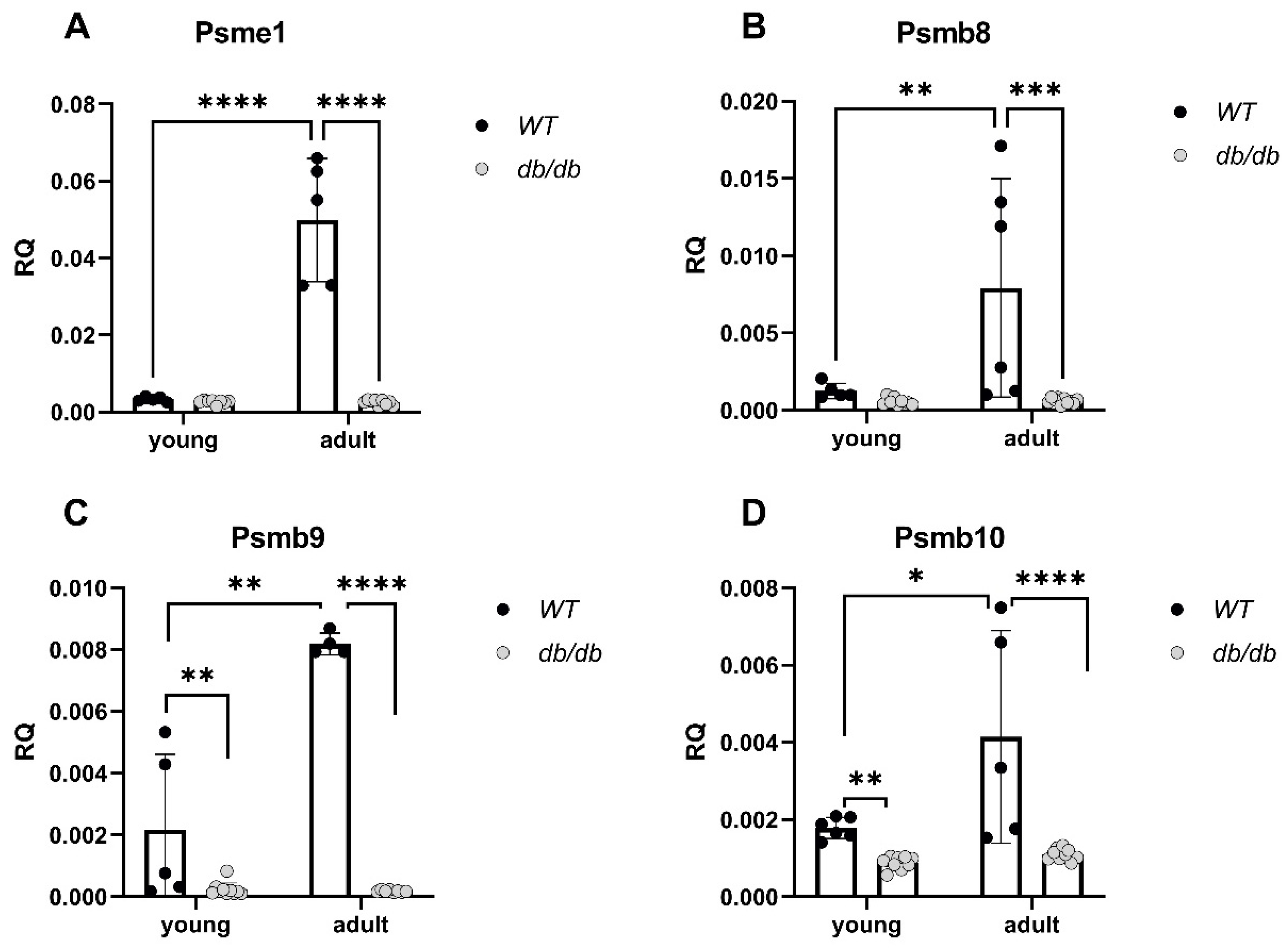
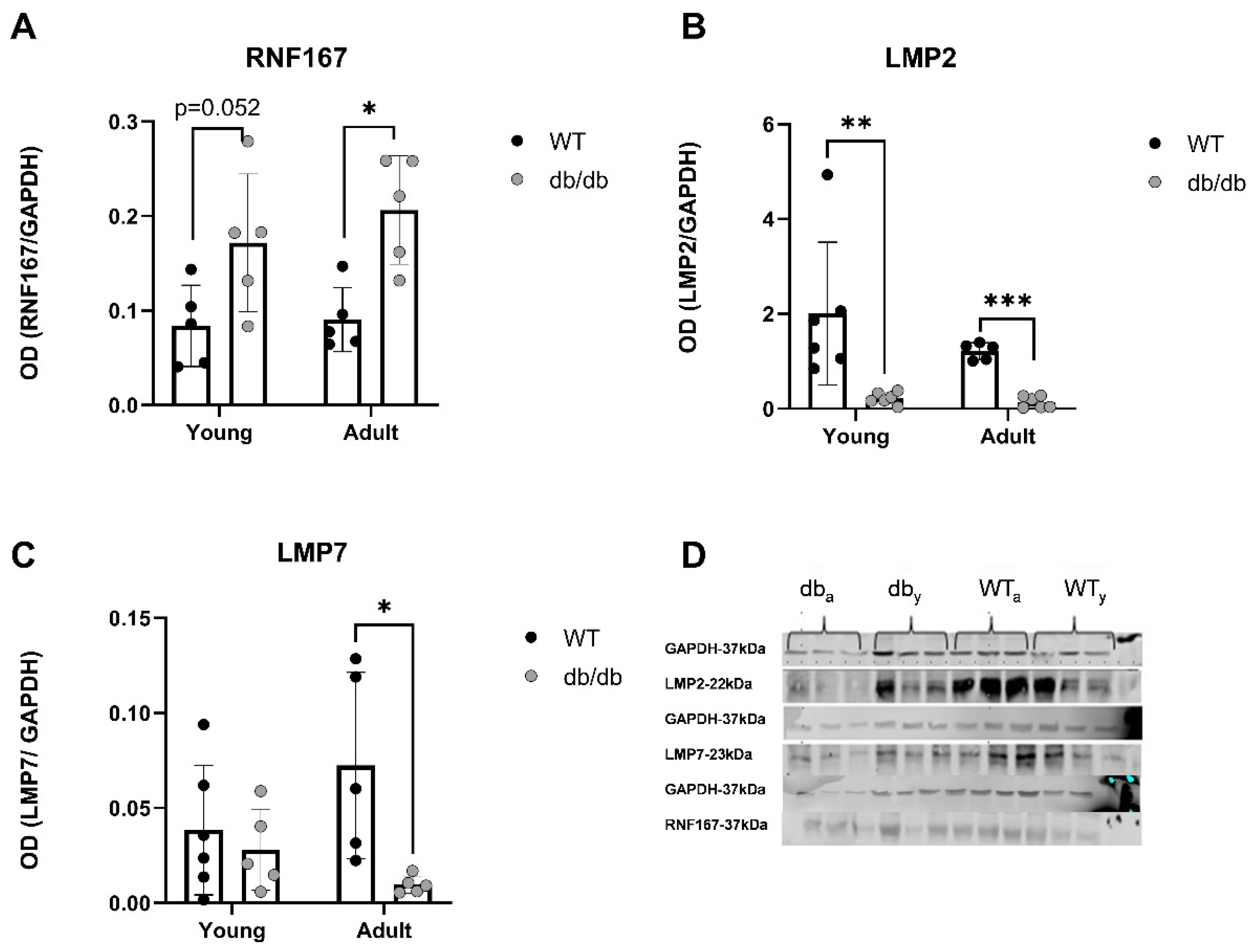
2.3. UPS-Related Protein Expression in db/db Mice
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Conflicts of Interest
References
- Deshpande, A.D.; Harris-Hayes, M.; Schootman, M. Epidemiology of diabetes and diabetes-related complications. Phys. Ther. 2008, 88, 1254–1264. [Google Scholar] [CrossRef]
- Kenny, H.C.; Abel, E.D. Heart Failure in Type 2 Diabetes Mellitus. Circ. Res. 2019, 124, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Tiderencel, K.A.; Hutcheon, D.A.; Ziegler, J. Probiotics for the treatment of type 2 diabetes: A review of randomized controlled trials. Diabetes Metab. Res. Rev. 2020, 36, e3213. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.D.; Wiviott, S.D.; Scirica, B.M.; Gurmu, Y.; Mosenzon, O.; Murphy, S.A.; Bhatt, D.L.; Leiter, L.A.; McGuire, D.K.; Wilding, J.P.; et al. Heart Failure Risk Stratification and Efficacy of Sodium-Glucose Cotransporter-2 Inhibitors in Patients with Type 2 Diabetes Mellitus. Circulation 2019, 140, 1569–1577. [Google Scholar] [CrossRef] [PubMed]
- Boudina, S.; Abel, E.D. Diabetic cardiomyopathy, causes and effects. Rev. Endocr. Metab. Disord. 2010, 11, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Pappachan, J.M.; Varughese, G.I.; Sriraman, R.; Arunagirinathan, G. Diabetic cardiomyopathy: Pathophysiology, diagnostic evaluation and management. World J. Diabetes 2013, 4, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Kawata, T.; Daimon, M.; Miyazaki, S.; Ichikawa, R.; Maruyama, M.; Chiang, S.J.; Ito, C.; Sato, F.; Watada, H.; Daida, H. Coronary microvascular function is independently associated with left ventricular filling pressure in patients with type 2 diabetes mellitus. Cardiovasc. Diabetol. 2015, 14, 98. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.; Bernis, C.; Vigneron, S.; Labbe, J.C.; Lorca, T. The anaphase-promoting complex: A key factor in the regulation of cell cycle. Oncogene 2005, 24, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A.; Orian, A.; Schwartz, A.L. Ubiquitin-mediated proteolysis: Biological regulation via destruction. Bioessays 2000, 22, 442–451. [Google Scholar] [CrossRef]
- Jesenberger, V.; Jentsch, S. Deadly encounter: Ubiquitin meets apoptosis. Nat. Rev. Mol. Cell Biol. 2002, 3, 112–121. [Google Scholar] [CrossRef]
- Ciechanover, A. Proteolysis: From the lysosome to ubiquitin and the proteasome. Nat. Rev. Mol. Cell Biol. 2005, 6, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Glickman, M.H.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, D.M.; Stoll, K.E.; Klevit, R.E. E2s: Structurally economical and functionally replete. Biochem. J. 2011, 433, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Parry, T.L.; Willis, M.S. Cardiac ubiquitin ligases: Their role in cardiac metabolism, autophagy, cardioprotection and therapeutic potential. Biochim. Biophys. Acta 2016, 1862, 2259–2269. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, J.; Wang, T.; Bai, J.; Zhang, Y.L.; Lin, Q.Y.; Li, J.M.; Zhao, Q.; Guo, S.B.; Li, H.H. Ablation of Immunoproteasome beta5i Subunit Suppresses Hypertensive Retinopathy by Blocking ATRAP Degradation in Mice. Mol. Ther. 2020, 28, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Benaroudj, N.; Tarcsa, E.; Cascio, P.; Goldberg, A.L. The unfolding of substrates and ubiquitin-independent protein degradation by proteasomes. Biochimie 2001, 83, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Bitzer, A.; Basler, M.; Krappmann, D.; Groettrup, M. Immunoproteasome subunit deficiency has no influence on the canonical pathway of NF-kappaB activation. Mol. Immunol. 2017, 83, 147–153. [Google Scholar] [CrossRef]
- Ostler, J.E.; Maurya, S.K.; Dials, J.; Roof, S.R.; Devor, S.T.; Ziolo, M.T.; Periasamy, M. Effects of insulin resistance on skeletal muscle growth and exercise capacity in type 2 diabetic mouse models. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E592–E605. [Google Scholar] [CrossRef]
- Kloß, A.; Meiners, S.; Ludwig, A.; Dahlmann, B. Multiple cardiac proteasome subtypes differ in their susceptibility to proteasome inhibitors. Cardiovasc. Res. 2010, 85, 367–375. [Google Scholar] [CrossRef]
- Greer, J.J.; Ware, D.P.; Lefer, D.J. Myocardial infarction and heart failure in the db/db diabetic mouse. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H146–H153. [Google Scholar] [CrossRef]
- Zhang, H.; Thoonen, R.; Yao, V.; Buys, E.S.; Popovich, J.; Su, Y.R.; Wang, T.J.; Scherrer-Crosbie, M. Regulation of B-type natriuretic peptide synthesis by insulin in obesity in male mice. Exp. Physiol. 2016, 101, 113–123. [Google Scholar] [CrossRef]
- Montag, J.; Syring, M.; Rose, J.; Weber, A.-L.; Ernstberger, P.; Mayer, A.-K.; Becker, E.; Keyser, B.; dos Remedios, C.; Perrot, A.; et al. Intrinsic MYH7 expression regulation contributes to tissue level allelic imbalance in hypertrophic cardiomyopathy. J. Muscle Res. Cell Motil. 2017, 38, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Predmore, J.M.; Wang, P.; Davis, F.; Bartolone, S.; Westfall, M.V.; Dyke, D.B.; Pagani, F.; Powell, S.R.; Day, S.M. Ubiquitin proteasome dysfunction in human hypertrophic and dilated cardiomyopathies. Circulation 2010, 121, 997–1004. [Google Scholar] [CrossRef]
- Lee, H.S.; Suh, J.Y.; Kang, B.-C.; Lee, E. Lipotoxicity dysregulates the immunoproteasome in podocytes and kidneys in type 2 diabetes. Am. J. Physiol. Renal Physiol. 2021, 320, F548–F558. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.R.; Samuel, S.M.; Wang, P.; Divald, A.; Thirunavukkarasu, M.; Koneru, S.; Wang, X.; Maulik, N. Upregulation of myocardial 11S-activated proteasome in experimental hyperglycemia. J. Mol. Cell. Cardiol. 2008, 44, 618–621. [Google Scholar] [CrossRef]
- Powell, S.R.; Herrmann, J.; Lerman, A.; Patterson, C.; Wang, X. The ubiquitin-proteasome system and cardiovascular disease. Prog. Mol. Biol. Transl. Sci. 2012, 109, 295–346. [Google Scholar] [PubMed]
- Shukla, S.K.; Rafiq, K. Proteasome biology and therapeutics in cardiac diseases. Transl. Res. 2019, 205, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Peris-Moreno, D.; Taillandier, D.; Polge, C. MuRF1/TRIM63, Master Regulator of Muscle Mass. Int. J. Mol. Sci. 2020, 21, 6663. [Google Scholar] [CrossRef] [PubMed]
- Portbury, A.L.; Ronnebaum, S.M.; Zungu, M.; Patterson, C.; Willis, M.S. Back to your heart: Ubiquitin proteasome system-regulated signal transduction. J. Mol. Cell. Cardiol. 2012, 52, 526–537. [Google Scholar] [CrossRef]
- Mattox, T.A.; Young, M.E.; Rubel, C.E.; Spaniel, C.; Rodríguez, J.E.; Grevengoed, T.J.; Gautel, M.; Xu, Z.; Anderson, E.J.; Willis, M.S. MuRF1 activity is present in cardiac mitochondria and regulates reactive oxygen species production in vivo. J. Bioenerg. Biomembr. 2014, 46, 173–187, Erratum in J. Bioenerg. Biomembr. 2015, 47, 281. [Google Scholar] [CrossRef]
- Rodríguez, J.E.; Schisler, J.C.; Patterson, C.; Willis, M.S. Seek and destroy: The ubiquitin-proteasome system in cardiac disease. Curr. Hypertens. Rep. 2009, 6, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.E.; Liao, J.Y.; He, J.; Schisler, J.C.; Newgard, C.B.; Drujan, D.; Glass, D.J.; Frederick, C.B.; Yoder, B.C.; Lalush, D.S.; et al. The ubiquitin ligase MuRF1 regulates PPARα activity in the heart by enhancing nuclear export via monoubiquitination. Mol. Cell. Endocrinol. 2015, 15, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Lam, Y.A.; Xu, W.; DeMartino, G.N.; Cohen, R.E. Editing of ubiquitin conjugates by an isopeptidase in the 26S proteasome. Nature 1997, 385, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.Y.; Wang, H.X.; Liu, X.H.; Guo, C.X.; Hua, Q.; Yu, X.H.; Li, N.; Yang, Y.Z.; Du, J.; Xia, Y.L.; et al. Circulating E3 ligases are novel and sensitive biomarkers for diagnosis of acute myocardial infarction. Clin. Sci. 2015, 128, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.V.; Narendradev, N.D.; Nambiar, R.P.; Kumar, R.; Srinivasula, S.M. Naturally occurring and tumor-associated variants of RNF167 promote lysosomal exocytosis and plasma membrane resealing. J. Cell Sci. 2020, 11, 133. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, J.R.; Yamazaki, Y.; Palmer, R.H. Tumour-associated mutations of PA-TM-RING ubiquitin ligases RNF167/RNF13 identify the PA domain as a determinant for endosomal localization. Biochem. J. 2014, 459, 27–36. [Google Scholar] [CrossRef]
- Lussier, M.P.; Herring, B.E.; Nasu-Nishimura, Y.; Neutzner, A.; Karbowski, M.; Youle, R.J.; Nicoll, R.A.; Roche, K.W. Ubiquitin ligase RNF167 regulates AMPA receptor-mediated synaptic transmission. Proc. Natl. Acad. Sci. USA 2012, 109, 19426–19431. [Google Scholar] [CrossRef] [PubMed]
- Govoruskina, N.; Jakovljevic, V.; Zivkovic, V.; Milosavljevic, I.; Jeremic, J.; Bradic, J.; Bolevich, S.; Omarov, I.A.; Djuric, D.; Radonjic, K.; et al. The Role of Cardiac N-Methyl-D-Aspartate Receptors in Heart Conditioning-Effects on Heart Function and Oxidative Stress. Biomolecules 2020, 10, 1065. [Google Scholar] [CrossRef]
- Barac, Y.D.; Emrich, F.; Krutzwakd-Josefson, E.; Schrepfer, S.; Sampaio, L.C.; Willerson, J.T.; Robbins, R.C.; Ciechanover, A.; Mohr, F.W.; Aravot, D.; et al. The ubiquitin-proteasome system: A potential therapeutic target for heart failure. J. Heart Lung Transplant. 2017, 36, 708–714. [Google Scholar] [CrossRef]
- Widagdo, J.; Guntupalli, S.; Jang, S.E.; Anggono, V. Regulation of AMPA Receptor Trafficking by Protein Ubiquitination. Front. Mol. Neurosci. 2017, 26, 347. [Google Scholar] [CrossRef]
- Gao, F.; Chen, D.; Si, J.; Hu, Q.; Qin, Z.; Fang, M.; Wang, G. The mitochondrial protein BNIP3L is the substrate of PARK2 and mediates mitophagy in PINK1/PARK2 pathway. Hum. Mol. Genet. 2015, 24, 2528–2538. [Google Scholar] [CrossRef] [PubMed]
- Uthman, L.; Baartscheer, A.; Schumacher, C.A.; Fiolet, J.W.; Kuschma, M.C.; Hollmann, M.W.; Coronel, R.; Weber, N.C.; Zuurbier, C.J. Direct Cardiac Actions of Sodium Glucose Cotransporter 2 Inhibitors Target Pathogenic Mechanisms Underlying Heart Failure in Diabetic Patients. Front. Physiol. 2018, 9, 1575. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Evans, E.; Waller-Evans, H. Lysosomal Ca2+ Homeostasis and Signaling in Health and Disease. Cold Spring Harb. Perspect. Biol. 2020, 12, a035311. [Google Scholar] [CrossRef] [PubMed]
- Bozi, L.H.M.; Takano, A.P.; Campos, J.C.; Rolim, N.; Dourado, P.M.; Voltarelli, V.A.; Wisløff, U.; Ferreira, J.C.; Barreto-Chaves, M.L.; Brum, P.C. Endoplasmic reticulum stress impairs cardiomyocyte contractility through JNK-dependent upregulation of BNIP3. Int. J. Cardiol. 2018, 272, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.; Zhao, Y.; Yao, T.; Yuan, A.; Xu, L.; Gao, L.; Ding, S.; Ding, H.; Pu, J.; He, B. Novel Protective Role for Ubiquitin-Specific Protease 18 in Pathological Cardiac Remodeling. Hypertension 2016, 68, 1160–1170. [Google Scholar] [CrossRef] [PubMed]
- Kasacka, I.; Piotrowska, Ż.; Niezgoda, M.; Lewandowska, A.; Łebkowski, W. Ageing-related changes in the levels of beta-catenin, CacyBP/SIP, galectin-3 and immunoproteasome subunit LMP7 in the heart of men. PLoS ONE 2020, 15, e0229462. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | db/db | |
|---|---|---|
| IVS (mm) | 0.8 ± 0.1 | 0.9 ± 0.1 |
| LVPW (mm) | 0.9 ± 0.1 | 0.9 ± 0.1 |
| LVEDD(mm) | 3.6 ± 0.7 | 3.9 ± 0.2 |
| LVESD (mm) | 2.9 ± 0.2 | 2.6 ± 0.3 * |
| FS (%) | 33 ± 14 | 34 ± 0.7 |
| Gene Primers | Catalog Number | Company |
|---|---|---|
| GAPDH | Mm99999915_g1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| PSME1 | Mm00650858_g1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| PSMB8 | Mm01278980_g1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| PSMB9 | Mm00479004_g1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| PSMB10 | Mm00479052_g1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| USP18 | Mm01188805_m1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| ISG15 | Mm01705338_s1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| RNF167 | Mm00550965_m1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| UBE2l6 | Mm00498295_m1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| Myh7 | Mm00600555_m1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| Nppb | Mm01255770_g1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| Tpm3 | Mm04336671_g1 | Applied Biosystems, Thermo Fisher scientific, Foster City, CA, USA |
| Primary Antibodies | Catalog Number | Company |
|---|---|---|
| GAPDH (1:1000) | sc-36502 | Santa Cruz Biotechnology, Dallas, TX, USA |
| RNF167 (1:100) | ab185099 | Abcam, Cambridge, UK |
| Lmp2 (1:100) | ab3328 | Abcam, Cambridge, UK |
| Lmp7 (1:100) | ab3329 | Abcam, Cambridge, UK |
| Secondary Antibodies | Cat Number | Company |
| Goat anti-rabbit IgG, IRDye®800RD (1:10,000) | 925-32213 | LI-COR Corporate, Lincoln, NE, USA |
| Donkey anti-mouse IgG, IRDye®680RD (1:10,000) | 925-98072 | LI-COR Corporate, Lincoln, NE, USA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nahum-Ankonina, O.; Kurtzwald-Josefson, E.; Ciechanover, A.; Waldman, M.; Shwartz-Rohaker, O.; Hochhauser, E.; Meyer, S.J.; Aravot, D.; Phillip, M.; Barac, Y.D. Ubiquitin Proteasome System Role in Diabetes-Induced Cardiomyopathy. Int. J. Mol. Sci. 2023, 24, 15376. https://doi.org/10.3390/ijms242015376
Nahum-Ankonina O, Kurtzwald-Josefson E, Ciechanover A, Waldman M, Shwartz-Rohaker O, Hochhauser E, Meyer SJ, Aravot D, Phillip M, Barac YD. Ubiquitin Proteasome System Role in Diabetes-Induced Cardiomyopathy. International Journal of Molecular Sciences. 2023; 24(20):15376. https://doi.org/10.3390/ijms242015376
Chicago/Turabian StyleNahum-Ankonina, Ortal, Efrat Kurtzwald-Josefson, Aaron Ciechanover, Maayan Waldman, Orna Shwartz-Rohaker, Edith Hochhauser, Sam J. Meyer, Dan Aravot, Moshe Phillip, and Yaron D. Barac. 2023. "Ubiquitin Proteasome System Role in Diabetes-Induced Cardiomyopathy" International Journal of Molecular Sciences 24, no. 20: 15376. https://doi.org/10.3390/ijms242015376
APA StyleNahum-Ankonina, O., Kurtzwald-Josefson, E., Ciechanover, A., Waldman, M., Shwartz-Rohaker, O., Hochhauser, E., Meyer, S. J., Aravot, D., Phillip, M., & Barac, Y. D. (2023). Ubiquitin Proteasome System Role in Diabetes-Induced Cardiomyopathy. International Journal of Molecular Sciences, 24(20), 15376. https://doi.org/10.3390/ijms242015376