
The Time-Resolved Salt Stress Response of Dunaliella tertiolecta—A Comprehensive System Biology Perspective

 , , , , and
, , , , and 
Abstract
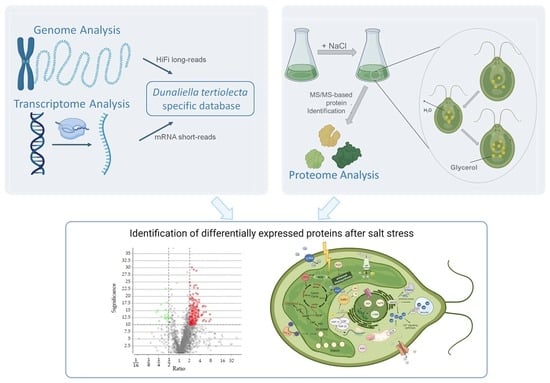
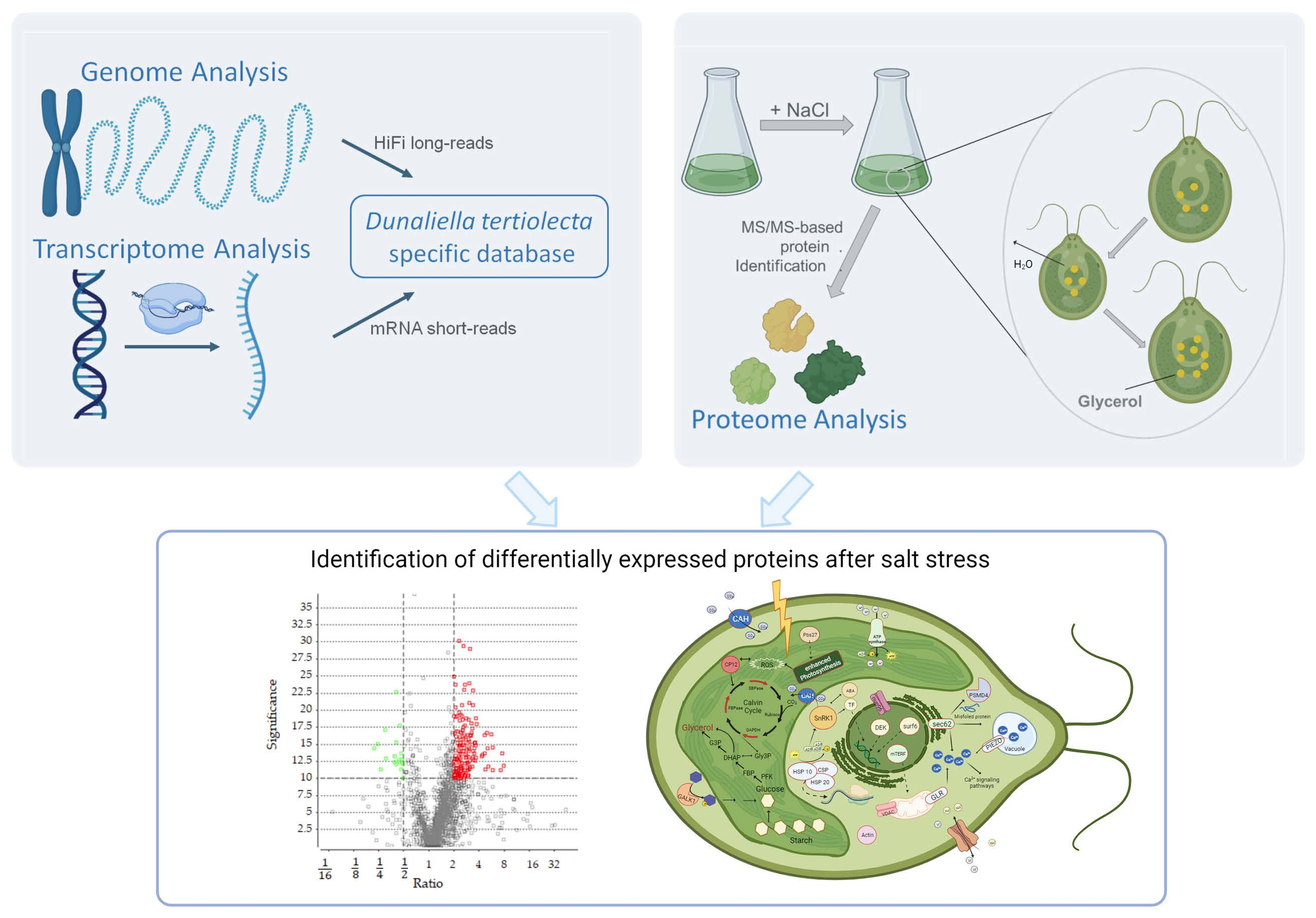
1. Introduction
2. Results and Discussion
2.1. Genome of D. tertiolecta and Functional Prediction of Protein Coding Sequences
2.2. Proteomic Changes after Salt Shock
2.3. Cluster of Orthologous Groups Analysis
2.4. Induced Expression of Proteins after 30 min Involved in Salt Stress
Identification of Phosphorylated Proteins under NaCl Stress
2.5. Differentially Expressed Proteins after 4 h of Salt Stress
2.6. Differentially Expressed Proteins Related to Long-Term Adaption
2.7. Changes in Key Enzymes of the Glycerol Pathway
2.8. Chaperons and Heat-Shock Proteins Affected by Salt Stress
2.9. Differently Expressed Transporters and Transmembrane Proteins
2.10. Further Adaption to Salt Stress and Comparison to D. salina
3. Materials and Methods
3.1. Algae Strains and Cultivation
3.2. Phylogenetic Characterization of Strains
3.3. High Molecular Weight DNA Extraction and Library Preparation
3.4. Total RNA Extraction and mRNA Sequencing
3.5. Genome Sequencing, Assembly, and Quality Assessment
3.6. Gene Prediction and Functional Analysis
3.7. Induction of Salt Stress
3.8. Proteomics
3.8.1. Protein Extraction
3.8.2. Protein Quantification and SDS-PAGE
3.8.3. In-Gel Digestion of Protein Samples and LC-MS/MS Analysis
3.8.4. Quantitative Analysis of Peptides and Proteins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iglina, T.; Iglin, P.; Pashchenko, D. Industrial CO2 Capture by Algae: A Review and Recent Advances. Sustainability 2022, 14, 3801. [Google Scholar] [CrossRef]
- Henley, W.J.; Cobbs, M.; Novoveská, L.; Buchheim, M.A. Phylogenetic analysis of Dunaliella (Chlorophyta) emphasizing new benthic and supralittoral isolates from Great Salt Lake. J. Phycol. 2018, 54, 483–493. [Google Scholar] [CrossRef]
- Hosseinzadeh Gharajeh, N.; Valizadeh, M.; Dorani, E.; Hejazi, M.A. Biochemical profiling of three indigenous Dunaliella isolates with main focus on fatty acid composition towards potential biotechnological application. Biotechnol. Rep. 2020, 26, e00479. [Google Scholar] [CrossRef] [PubMed]
- Preetha, K.; John, L.; Subin, C.S.; Vijayan, K.K. Phenotypic and genetic characterization of Dunaliella (Chlorophyta) from Indian salinas and their diversity. Aquat. Biosyst. 2012, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Jiang, J.-G. Osmotic responses of Dunaliella to the changes of salinity. J. Cell Physiol. 2009, 219, 251–258. [Google Scholar] [CrossRef]
- Oren, A. The ecology of Dunaliella in high-salt environments. J. Biol. Res. 2014, 21, 23. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, J.-G.; Wu, G.-H. Effects of salinity changes on the growth of Dunaliella salina and its isozyme activities of glycerol-3-phosphate dehydrogenase. J. Agric. Food Chem. 2009, 57, 6178–6182. [Google Scholar] [CrossRef]
- Shariati, M.; Reza, M. Microalgal Biotechnology and Bioenergy in Dunaliella. In Progress in Molecular and Environmental Bioengineering—From Analysis and Modeling to Technology Applications; Carpi, A., Ed.; InTech: London, UK, 2011; ISBN 978-953-307-268-5. [Google Scholar]
- Chow, Y.Y.S.; Goh, S.J.M.; Su, Z.; Ng, D.H.P.; Lim, C.Y.; Lim, N.Y.N.; Lin, H.; Fang, L.; Lee, Y.K. Continual production of glycerol from carbon dioxide by Dunaliella tertiolecta. Bioresour. Technol. 2013, 136, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Ben-Amotz, A.; Sussman, I.; Avron, M. Glycerol production by Dunaliella. In New Trends in Research and Utilization of Solar Energy through Biological Systems; Mislin, H., Bachofen, R., Eds.; Birkhäuser Basel: Basel, Switzerland, 1982; pp. 55–58. ISBN 978-3-0348-6307-0. [Google Scholar]
- Tan, H.W.; Abdul Aziz, A.R.; Aroua, M.K. Glycerol production and its applications as a raw material: A review. Renew. Sustain. Energy Rev. 2013, 27, 118–127. [Google Scholar] [CrossRef]
- Peijs, T.; Kirschbaum, R.; Lemstra, P.J. Chapter 5: A critical review of carbon fiber and related products from an industrial perspective. Adv. Ind. Eng. Polym. Res. 2022, 5, 90–106. [Google Scholar] [CrossRef]
- Arnold, U.; Brück, T.; de Palmenaer, A.; Kuse, K. Carbon Capture and Sustainable Utilization by Algal Polyacrylonitrile Fiber Production: Process Design, Techno-Economic Analysis, and Climate Related Aspects. Ind. Eng. Chem. Res. 2018, 57, 7922–7933. [Google Scholar] [CrossRef]
- Norzagaray-Valenzuela, C.D.; Germán-Báez, L.J.; Valdez-Flores, M.A.; Hernández-Verdugo, S.; Shelton, L.M.; Valdez-Ortiz, A. Establishment of an efficient genetic transformation method in Dunaliella tertiolecta mediated by Agrobacterium tumefaciens. J. Microbiol. Methods 2018, 150, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cong, Y.; Wang, Y.; Guo, Z.; Yue, J.; Xing, Z.; Gao, X.; Chai, X. Identification of Early Salinity Stress-Responsive Proteins in Dunaliella salina by isobaric tags for relative and absolute quantitation (iTRAQ)-Based Quantitative Proteomic Analysis. Int. J. Mol. Sci. 2019, 20, 599. [Google Scholar] [CrossRef]
- He, Q.; Lin, Y.; Tan, H.; Zhou, Y.; Wen, Y.; Gan, J.; Li, R.; Zhang, Q. Transcriptomic profiles of Dunaliella salina in response to hypersaline stress. BMC Genom. 2020, 21, 115. [Google Scholar] [CrossRef] [PubMed]
- Construction Industry Needs Whole Life Carbon Understanding to Hit Net Zero—New Report Shows. Available online: https://www.wbcsd.org/Programs/Cities-and-Mobility/Sustainable-Cities/Transforming-the-Built-Environment/Decarbonization/news/Construction-industry-needs-whole-life-carbon-understanding-to-hit-net-zero-new-report-shows (accessed on 23 December 2022).
- Mansour, M.M.F.; Hassan, F.A.S. How salt stress-responsive proteins regulate plant adaptation to saline conditions. Plant Mol. Biol. 2022, 108, 175–224. [Google Scholar] [CrossRef]
- Athar, H.-U.-R.; Zulfiqar, F.; Moosa, A.; Ashraf, M.; Zafar, Z.U.; Zhang, L.; Ahmed, N.; Kalaji, H.M.; Nafees, M.; Hossain, M.A.; et al. Salt stress proteins in plants: An overview. Front. Plant Sci. 2022, 13, 999058. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, J.; Wen, W.; Sun, J.; Shu, S.; Guo, S. Transcriptome and Proteome Analysis Identifies Salt Stress Response Genes in Bottle Gourd Rootstock-Grafted Watermelon Seedlings. Agronomy 2023, 13, 618. [Google Scholar] [CrossRef]
- Chen, H.; Lu, Y.; Jiang, J.-G. Comparative analysis on the key enzymes of the glycerol cycle metabolic pathway in Dunaliella salina under osmotic stresses. PLoS ONE 2012, 7, e37578. [Google Scholar] [CrossRef]
- Yao, L.; Tan, K.W.M.; Tan, T.W.; Lee, Y.K. Exploring the transcriptome of non-model oleaginous microalga Dunaliella tertiolecta through high-throughput sequencing and high performance computing. BMC Bioinform. 2017, 18, 122. [Google Scholar] [CrossRef]
- Rismani-Yazdi, H.; Haznedaroglu, B.Z.; Bibby, K.; Peccia, J. Transcriptome sequencing and annotation of the microalgae Dunaliella tertiolecta: Pathway description and gene discovery for production of next-generation biofuels. BMC Genom. 2011, 12, 148. [Google Scholar] [CrossRef]
- Harvey, P.J.; Ben-Amotz, A. Towards a sustainable Dunaliella salina microalgal biorefinery for 9-cis β-carotene production. Algal Res. 2020, 50, 102002. [Google Scholar] [CrossRef]
- Xu, Y.; Ibrahim, I.M.; Wosu, C.I.; Ben-Amotz, A.; Harvey, P.J. Potential of New Isolates of Dunaliella Salina for Natural β-Carotene Production. Biology 2018, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Bian, Y.; Zhao, Q.; Chen, S.; Mao, J.; Song, C.; Cheng, K.; Xiao, Z.; Zhang, C.; Ma, W.; et al. Salinity-Induced Palmella Formation Mechanism in Halotolerant Algae Dunaliella salina Revealed by Quantitative Proteomics and Phosphoproteomics. Front. Plant Sci. 2017, 8, 810. [Google Scholar] [CrossRef] [PubMed]
- Polle, J.E.; Calhoun, S.; McKie-Krisberg, Z.; Prochnik, S.; Neofotis, P.; Yim, W.C.; Hathwaik, L.T.; Jenkins, J.; Molina, H.; Bunkenborg, J.; et al. Genomic adaptations of the green alga Dunaliella salina to life under high salinity. Algal Res. 2020, 50, 101990. [Google Scholar] [CrossRef]
- Zhang, X.; Davenport, K.W.; Gu, W.; Daligault, H.E.; Munk, A.C.; Tashima, H.; Reitenga, K.; Green, L.D.; Han, C.S. Improving genome assemblies by sequencing PCR products with PacBio. Biotechniques 2012, 53, 61–62. [Google Scholar] [CrossRef]
- Berg, J.M.; Tymoczko, J.L.; Gatto, G.J., Jr.; Stryer, L.; Biochemie, S. Stryer Biochemie, 7th ed.; Auflage; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 978-3-8274-2989-6. [Google Scholar]
- Seifikalhor, M.; Aliniaeifard, S.; Shomali, A.; Azad, N.; Hassani, B.; Lastochkina, O.; Li, T. Calcium signaling and salt tolerance are diversely entwined in plants. Plant Signal. Behav. 2019, 14, 1665455. [Google Scholar] [CrossRef]
- Stanke, M.; Schöffmann, O.; Morgenstern, B.; Waack, S. Gene prediction in eukaryotes with a generalized hidden Markov model that uses hints from external sources. BMC Bioinform. 2006, 7, 62. [Google Scholar] [CrossRef]
- Hoff, K.J.; Lange, S.; Lomsadze, A.; Borodovsky, M.; Stanke, M. BRAKER1: Unsupervised RNA-Seq-Based Genome Annotation with GeneMark-ET and AUGUSTUS. Bioinformatics 2016, 32, 767–769. [Google Scholar] [CrossRef]
- Hoff, K.J.; Lomsadze, A.; Borodovsky, M.; Stanke, M. Whole-Genome Annotation with BRAKER. Methods Mol. Biol. 2019, 1962, 65–95. [Google Scholar] [CrossRef]
- Brůna, T.; Hoff, K.J.; Lomsadze, A.; Stanke, M.; Borodovsky, M. BRAKER2: Automatic eukaryotic genome annotation with GeneMark-EP+ and AUGUSTUS supported by a protein database. NAR Genom. Bioinform. 2021, 3, lqaa108. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Barnett, D.W.; Garrison, E.K.; Quinlan, A.R.; Strömberg, M.P.; Marth, G.T. BamTools: A C++ API and toolkit for analyzing and managing BAM files. Bioinformatics 2011, 27, 1691–1692. [Google Scholar] [CrossRef] [PubMed]
- Lomsadze, A.; Burns, P.D.; Borodovsky, M. Integration of mapped RNA-Seq reads into automatic training of eukaryotic gene finding algorithm. Nucleic Acids Res. 2014, 42, e119. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Stanke, M.; Diekhans, M.; Baertsch, R.; Haussler, D. Using native and syntenically mapped cDNA alignments to improve de novo gene finding. Bioinformatics 2008, 24, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Han, B.; Wang, T.; Chen, S.; Li, H.; Zhang, Y.; Dai, S. Mechanisms of plant salt response: Insights from proteomics. J. Proteome Res. 2012, 11, 49–67. [Google Scholar] [CrossRef]
- Cheng, Y.; Chen, G.; Hao, D.; Lu, H.; Shi, M.; Mao, Y.; Huang, X.; Zhang, Z.; Xue, L. Salt-induced root protein profile changes in seedlings of maize inbred lines with differing salt tolerances. Chilean J. Agric. Res. 2014, 74, 468–476. [Google Scholar] [CrossRef]
- Son, S.; Park, S.R. Plant translational reprogramming for stress resilience. Front. Plant Sci. 2023, 14, 1151587. [Google Scholar] [CrossRef]
- He, J.; Zhuang, Y.; Li, C.; Sun, X.; Zhao, S.; Ma, C.; Lin, H.; Zhou, H. SIMP1 modulates salt tolerance by elevating ERAD efficiency through UMP1A-mediated proteasome maturation in plants. New Phytol. 2021, 232, 625–641. [Google Scholar] [CrossRef]
- Läuchli, A.; Lüttge, U. (Eds.) Salinity: Environment—Plants—Molecules; Springer: Dordrecht, The Netherlands, 2002; ISBN 978-0-306-48155-0. [Google Scholar]
- Thoden, J.B.; Holden, H.M. Molecular structure of galactokinase. J. Biol. Chem. 2003, 278, 33305–33311. [Google Scholar] [CrossRef]
- Slepak, T.; Tang, M.; Addo, F.; Lai, K. Intracellular galactose-1-phosphate accumulation leads to environmental stress response in yeast model. Mol. Genet. Metab. 2005, 86, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Jha, B. Isolation and characterization of extracellular polymeric substances from micro-algae Dunaliellasalina under salt stress. Bioresour. Technol. 2009, 100, 3382–3386. [Google Scholar] [CrossRef] [PubMed]
- Jamsheer, K.M.; Kumar, M.; Srivastava, V. SNF1-related protein kinase 1: The many-faced signaling hub regulating developmental plasticity in plants. J. Exp. Bot. 2021, 72, 6042–6065. [Google Scholar] [CrossRef]
- Ramon, M.; Dang, T.V.T.; Broeckx, T.; Hulsmans, S.; Crepin, N.; Sheen, J.; Rolland, F. Default Activation and Nuclear Translocation of the Plant Cellular Energy Sensor SnRK1 Regulate Metabolic Stress Responses and Development. Plant Cell 2019, 31, 1614–1632. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, L.M.; Tandoc, K.; Topisirovic, I.; Furic, L. Cross-talk between protein synthesis, energy metabolism and autophagy in cancer. Curr. Opin. Genet. Dev. 2018, 48, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Carianopol, C.S.; Chan, A.L.; Dong, S.; Provart, N.J.; Lumba, S.; Gazzarrini, S. An abscisic acid-responsive protein interaction network for sucrose non-fermenting related kinase1 in abiotic stress response. Commun. Biol. 2020, 3, 145. [Google Scholar] [CrossRef]
- Cowan, A.K.; Rose, P.D. Abscisic Acid Metabolism in Salt-Stressed Cells of Dunaliella salina: Possible Interrelationship with beta-Carotene Accumulation. Plant Physiol. 1991, 97, 798–803. [Google Scholar] [CrossRef]
- Körner, S.; Pick, T.; Bochen, F.; Wemmert, S.; Körbel, C.; Menger, M.D.; Cavalié, A.; Kühn, J.-P.; Schick, B.; Linxweiler, M. Antagonizing Sec62 function in intracellular Ca2+ homeostasis represents a novel therapeutic strategy for head and neck cancer. Front. Physiol. 2022, 13, 880004. [Google Scholar] [CrossRef]
- Yang, M.; Luo, S.; Wang, X.; Li, C.; Yang, J.; Zhu, X.; Xiao, L.; Sun, L. ER-Phagy: A New Regulator of ER Homeostasis. Front. Cell Dev. Biol. 2021, 9, 684526. [Google Scholar] [CrossRef]
- Hu, S.; Ye, H.; Cui, Y.; Jiang, L. AtSec62 is critical for plant development and is involved in ER-phagy in Arabidopsis thaliana. J. Integr. Plant Biol. 2020, 62, 181–200. [Google Scholar] [CrossRef] [PubMed]
- Hammani, K.; Cook, W.B.; Barkan, A. RNA binding and RNA remodeling activities of the half-a-tetratricopeptide (HAT) protein HCF107 underlie its effects on gene expression. Proc. Natl. Acad. Sci. USA 2012, 109, 5651–5656. [Google Scholar] [CrossRef] [PubMed]
- Preker, P.J.; Keller, W. The HAT helix, a repetitive motif implicated in RNA processing. Trends Biochem. Sci. 1998, 23, 15–16. [Google Scholar] [CrossRef]
- Nie, Y.; Li, Y.; Liu, M.; Ma, B.; Sui, X.; Chen, J.; Yu, Y.; Dong, C.-H. The nucleoporin NUP160 and NUP96 regulate nucleocytoplasmic export of mRNAs and participate in ethylene signaling and response in Arabidopsis. Plant Cell Rep. 2023, 42, 549–559. [Google Scholar] [CrossRef]
- Shumkova, G.A.; Popova, L.G.; Balnokin, Y.V. Export of Na+ from cells of the halotolerant microalga Dunaliella maritima: Na+/H+ antiporter or primary Na+-pump? Biochemistry 2000, 65, 917–923. [Google Scholar]
- Zulawski, M.; Braginets, R.; Schulze, W.X. PhosPhAt goes kinases—searchable protein kinase target information in the plant phosphorylation site database PhosPhAt. Nucleic Acids Res. 2013, 41, D1176–D1184. [Google Scholar] [CrossRef]
- Kamiyama, Y.; Katagiri, S.; Umezawa, T. Growth Promotion or Osmotic Stress Response: How SNF1-Related Protein Kinase 2 (SnRK2) Kinases Are Activated and Manage Intracellular Signaling in Plants. Plants 2021, 10, 1443. [Google Scholar] [CrossRef]
- Ardito, F.; Giuliani, M.; Perrone, D.; Troiano, G.; Lo Muzio, L. The crucial role of protein phosphorylation in cell signaling and its use as targeted therapy (Review). Int. J. Mol. Med. 2017, 40, 271–280. [Google Scholar] [CrossRef]
- Katz, A.; Pick, U.; Avron, M. Characterization and reconstitution of the Na+/H+ antiporter from the plasma membrane of the halotolerant alga Dunaliella. Biochim. Biophys. Acta 1989, 983, 9–14. [Google Scholar] [CrossRef]
- Zhou, J.; Zhao, L.; Wu, Y.; Zhang, X.; Cheng, S.; Wei, F.; Zhang, Y.; Zhu, H.; Zhou, Y.; Feng, Z.; et al. A DEK domain-containing protein GhDEK2D mediated Gossypium hirsutum enhanced resistance to Verticillium dahliae. Plant Signal. Behav. 2022, 17, 2024738. [Google Scholar] [CrossRef]
- Sawatsubashi, S.; Murata, T.; Lim, J.; Fujiki, R.; Ito, S.; Suzuki, E.; Tanabe, M.; Zhao, Y.; Kimura, S.; Fujiyama, S.; et al. A histone chaperone, DEK, transcriptionally coactivates a nuclear receptor. Genes Dev. 2010, 24, 159–170. [Google Scholar] [CrossRef] [PubMed]
- DIELEN, A.-S.; BADAOUI, S.; CANDRESSE, T.; GERMAN-RETANA, S. The ubiquitin/26S proteasome system in plant–pathogen interactions: A never-ending hide-and-seek game. Mol. Plant Pathol. 2009, 11, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L. Role of the Ubiquitin Proteasome System in Plant Response to Abiotic Stress. Int. Rev. Cell Mol. Biol. 2019, 343, 65–110. [Google Scholar] [CrossRef]
- Kurepa, J.; Wang, S.; Li, Y.; Smalle, J. Proteasome regulation, plant growth and stress tolerance. Plant Signal. Behav. 2009, 4, 924–927. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-W.; Li, X.; Thompson, D.; Wooding, K.; Chang, T.; Tang, Z.; Yu, H.; Thomas, P.J.; DeMartino, G.N. ATP Binding and ATP Hydrolysis Play Distinct Roles in the Function of 26S Proteasome. Mol. Cell 2006, 24, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wang, H.; Liu, Z.; Pang, Z.; Zhang, C.; Zhao, M.; Ning, B.; Song, B.; Liu, S.; He, Z.; et al. The 26S Proteasome Regulatory Subunit GmPSMD Promotes Resistance to Phytophthora sojae in Soybean. Front. Plant Sci. 2021, 12, 513388. [Google Scholar] [CrossRef]
- Tanaka, K. The proteasome: Overview of structure and functions. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 12–36. [Google Scholar] [CrossRef]
- VerPlank, J.; Goldberg, A. Regulating Protein Breakdown Through Proteasome Phosphorylation. Biochem. J. 2017, 474, 3355–3371. [Google Scholar] [CrossRef]
- Güler, G.; Köçkar, F.; Aydoğan Türkoğlu, S. The Bioinformatic and Expression Analysis of PSMD4 Gene. Hacet. J. Biol. Chem. 2018, 3, 329–336. [Google Scholar] [CrossRef]
- La Angeles de Torre-Ruiz, M.; Torres, J.; Arino, J.; Herrero, E. Sit4 is required for proper modulation of the biological functions mediated by Pkc1 and the cell integrity pathway in Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 33468–33476. [Google Scholar] [CrossRef]
- Han, Q.; Pan, C.; Wang, Y.; Zhao, L.; Wang, Y.; Sang, J. PP2A-Like Protein Phosphatase (Sit4) Regulatory Subunits, Sap155 and Sap190, Regulate Candida albicans’ Cell Growth, Morphogenesis, and Virulence. Front. Microbiol. 2019, 10, 2943. [Google Scholar] [CrossRef] [PubMed]
- Sutton, A.; Immanuel, D.; Arndt, K.T. The SIT4 protein phosphatase functions in late G1 for progression into S phase. Mol. Cell Biol. 1991, 11, 2133–2148. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Davenport, R. Glutamate receptors in plants. Ann. Bot. 2002, 90, 549–557. [Google Scholar] [CrossRef] [PubMed]
- De Bortoli, S.; Teardo, E.; Szabò, I.; Morosinotto, T.; Alboresi, A. Evolutionary insight into the ionotropic glutamate receptor superfamily of photosynthetic organisms. Biophys. Chem. 2016, 218, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Liu, N.; Tang, S.; Qin, T.; Huang, J. Roles of Glutamate Receptor-Like Channels (GLRs) in Plant Growth and Response to Environmental Stimuli. Plants 2022, 11, 3450. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.-M.; Sun, Y.-Y.; Ye, X.-Y.; Li, Z.-G. Signaling Role of Glutamate in Plants. Front. Plant Sci. 2020, 10, 1743. [Google Scholar] [CrossRef]
- Knight, H. Calcium signaling during abiotic stress in plants. Int. Rev. Cytol. 2000, 195, 269–324. [Google Scholar] [CrossRef]
- Issa, A.A. The Role of Calcium in the Stress Response of the Halotolerant Green Alga Dunaliella bardawi. Available online: https://www.researchgate.net/profile/Ahmed-Issa-10/publication/259484655_Issa_AA_1996_The_role_of_calcium_in_the_stress_response_of_the_halotolerant_green_alga_Dunaliella_bardawil_Ben_Amotz_et_Avron_Phyton_36_295-302/links/0a85e52f5f95bf3610000000/Issa-AA-1996-The-role-of-calcium-in-the-stress-response-of-the-halotolerant-green-alga-Dunaliella-bardawil-Ben-Amotz-et-Avron-Phyton-36-295-302.pdf (accessed on 20 September 2023).
- Xu, T.; Niu, J.; Jiang, Z. Sensing Mechanisms: Calcium Signaling Mediated Abiotic Stress in Plants. Front. Plant Sci. 2022, 13, 925863. [Google Scholar] [CrossRef]
- Batistič, O.; Kudla, J. Analysis of calcium signaling pathways in plants. Biochim. Biophys. Acta 2012, 1820, 1283–1293. [Google Scholar] [CrossRef]
- Yip Delormel, T.; Boudsocq, M. Properties and functions of calcium-dependent protein kinases and their relatives in Arabidopsis thaliana. New Phytol. 2019, 224, 585–604. [Google Scholar] [CrossRef]
- Mehlmer, N.; Wurzinger, B.; Stael, S.; Hofmann-Rodrigues, D.; Csaszar, E.; Pfister, B.; Bayer, R.; Teige, M. The Ca2+-dependent protein kinase CPK3 is required for MAPK-independent salt-stress acclimation in Arabidopsis. Plant J. 2010, 63, 484–498. [Google Scholar] [CrossRef] [PubMed]
- López-Calcagno, P.E.; Howard, T.P.; Raines, C.A. The CP12 protein family: A thioredoxin-mediated metabolic switch? Front. Plant Sci. 2014, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Tamoi, M.; Shigeoka, S. CP12 Is Involved in Protection against High Light Intensity by Suppressing the ROS Generation in Synechococcus elongatus PCC7942. Plants 2021, 10, 1275. [Google Scholar] [CrossRef] [PubMed]
- Elena López-Calcagno, P.; Omar Abuzaid, A.; Lawson, T.; Anne Raines, C. Arabidopsis CP12 mutants have reduced levels of phosphoribulokinase and impaired function of the Calvin-Benson cycle. J. Exp. Bot. 2017, 68, 2285–2298. [Google Scholar] [CrossRef] [PubMed]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Oxidative damage and antioxidative system in algae. Toxicol. Rep. 2019, 6, 1309–1313. [Google Scholar] [CrossRef]
- DiMario, R.J.; Clayton, H.; Mukherjee, A.; Ludwig, M.; Moroney, J.V. Plant Carbonic Anhydrases: Structures, Locations, Evolution, and Physiological Roles. Mol. Plant 2017, 10, 30–46. [Google Scholar] [CrossRef]
- Liska, A.J.; Shevchenko, A.; Pick, U.; Katz, A. Enhanced photosynthesis and redox energy production contribute to salinity tolerance in Dunaliella as revealed by homology-based proteomics. Plant Physiol. 2004, 136, 2806–2817. [Google Scholar] [CrossRef]
- Zheng, Z.; Gao, S.; He, Y.; Li, Z.; Li, Y.; Cai, X.; Gu, W.; Wang, G. The enhancement of the oxidative pentose phosphate pathway maybe involved in resolving imbalance between photosystem I and II in Dunaliella salina. Algal Res. 2017, 26, 402–408. [Google Scholar] [CrossRef]
- Huang, G.; Xiao, Y.; Pi, X.; Zhao, L.; Zhu, Q.; Wang, W.; Kuang, T.; Han, G.; Sui, S.-F.; Shen, J.-R. Structural insights into a dimeric Psb27-photosystem II complex from a cyanobacterium Thermosynechococcus vulcanus. Proc. Natl. Acad. Sci. USA 2021, 118, e2018053118. [Google Scholar] [CrossRef]
- Nowaczyk, M.M.; Hebeler, R.; Schlodder, E.; Meyer, H.E.; Warscheid, B.; Rögner, M. Psb27, a Cyanobacterial Lipoprotein, Is Involved in the Repair Cycle of Photosystem II. Plant Cell 2006, 18, 3121–3131. [Google Scholar] [CrossRef]
- Roy, U.K.; Nielsen, B.V.; Milledge, J.J. Antioxidant Production in Dunaliella. Appl. Sci. 2021, 11, 3959. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as Sources of Carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Harvey, P.J. Carotenoid Production by Dunaliella salina under Red Light. Antioxidants 2019, 8, 123. [Google Scholar] [CrossRef] [PubMed]
- Jahnke, L.S.; White, A.L. Long-term hyposaline and hypersaline stresses produce distinct antioxidant responses in the marine alga Dunaliella tertiolecta. J. Plant Physiol. 2003, 160, 1193–1202. [Google Scholar] [CrossRef]
- Wang, W.-R.; Liang, J.-H.; Wang, G.-F.; Sun, M.-X.; Peng, F.-T.; Xiao, Y.-S. Overexpression of PpSnRK1α in tomato enhanced salt tolerance by regulating ABA signaling pathway and reactive oxygen metabolism. BMC Plant Biol. 2020, 20, 128. [Google Scholar] [CrossRef]
- Murphy, M.P.; Bayir, H.; Belousov, V.; Chang, C.J.; Davies, K.J.A.; Davies, M.J.; Dick, T.P.; Finkel, T.; Forman, H.J.; Janssen-Heininger, Y.; et al. Guidelines for measuring reactive oxygen species and oxidative damage in cells and in vivo. Nat. Metab. 2022, 4, 651–662. [Google Scholar] [CrossRef]
- Magoulas, C.; Zatsepina, O.V.; Jordan, P.W.; Jordan, E.G.; Fried, M. The SURF-6 protein is a component of the nucleolar matrix and has a high binding capacity for nucleic acids in vitro. Eur. J. Cell Biol. 1998, 75, 174–183. [Google Scholar] [CrossRef]
- Moraleva, A.; Magoulas, C.; Polzikov, M.; Hacot, S.; Mertani, H.C.; Diaz, J.-J.; Zatsepina, O. Involvement of the specific nucleolar protein SURF6 in regulation of proliferation and ribosome biogenesis in mouse NIH/3T3 fibroblasts. Cell Cycle 2017, 16, 1979–1991. [Google Scholar] [CrossRef]
- Hudson, R.C.; Daniel, R.M. L-glutamate dehydrogenases: Distribution, properties and mechanism. Comp. Biochem. Physiol. B 1993, 106, 767–792. [Google Scholar] [CrossRef]
- Tercé-Laforgue, T.; Clément, G.; Marchi, L.; Restivo, F.M.; Lea, P.J.; Hirel, B. Resolving the Role of Plant NAD-Glutamate Dehydrogenase: III. Overexpressing Individually or Simultaneously the Two Enzyme Subunits under Salt Stress Induces Changes in the Leaf Metabolic Profile and Increases Plant Biomass Production. Plant Cell Physiol. 2015, 56, 1918–1929. [Google Scholar] [CrossRef]
- Li, C.; Zhao, T.; Ren, L.; Cui, D.; Zhan, Y.; Chang, Y. Characterization of a novel glutamate dehydrogenase gene and its response to heat stress in the sea urchin Strongylocentrotus intermedius. Aquac. Rep. 2023, 28, 101446. [Google Scholar] [CrossRef]
- Wu, N.; Yang, M.; Gaur, U.; Xu, H.; Yao, Y.; Li, D. Alpha-Ketoglutarate: Physiological Functions and Applications. Biomol. Ther. 2016, 24, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Plaitakis, A.; Kalef-Ezra, E.; Kotzamani, D.; Zaganas, I.; Spanaki, C. The Glutamate Dehydrogenase Pathway and Its Roles in Cell and Tissue Biology in Health and Disease. Biology 2017, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Omar, T.A.; Zewain, S.K.; Ghonaim, M.M.; Refaat, K.A.; Abou-Elela, D.H. Role of engulfment and cell motility 1 (ELMO1) gene polymorphism in development of diabetic kidney disease. Egypt. J. Med. Hum. Genet. 2021, 22, 49. [Google Scholar] [CrossRef]
- Sato, Y.; Sato, A.; Mizuno, S.; Hirota, J.-N.; Fujima, S.; Ishii, C.; Sano, Y.; Furuichi, T. Comparative gene expression analysis of the engulfment and cell motility (ELMO) protein family in the mouse brain. Gene Expr. Patterns 2019, 34, 119070. [Google Scholar] [CrossRef]
- Tunc-Ozdemir, M.; Miller, G.; Song, L.; Kim, J.; Sodek, A.; Koussevitzky, S.; Misra, A.N.; Mittler, R.; Shintani, D. Thiamin confers enhanced tolerance to oxidative stress in Arabidopsis. Plant Physiol. 2009, 151, 421–432. [Google Scholar] [CrossRef]
- Keil, L.; Breitsameter, J.; Rieger, B.; Garbe, D.; Brueck, T. Optimization of the glycerol production from Dunaliella tertiolecta and Dunaliella isolates. bioRxiv 2023. bioRxiv:18.558238. [Google Scholar]
- Hostettler, C.; Kölling, K.; Santelia, D.; Streb, S.; Kötting, O.; Zeeman, S.C. Analysis of starch metabolism in chloroplasts. Methods Mol. Biol. 2011, 775, 387–410. [Google Scholar] [CrossRef]
- Xu, Y.; Ibrahim, I.M.; Harvey, P.J. The influence of photoperiod and light intensity on the growth and photosynthesis of Dunaliella salina (chlorophyta) CCAP 19/30. Plant Physiol. Biochem. 2016, 106, 305–315. [Google Scholar] [CrossRef]
- Goyal, A. Osmoregulation in Dunaliella, Part II: Photosynthesis and starch contribute carbon for glycerol synthesis during a salt stress in Dunaliella tertiolecta. Plant Physiol. Biochem. 2007, 45, 705–710. [Google Scholar] [CrossRef]
- Yang, W.; Cao, Y.; Sun, X.; Huang, F.; He, Q.; Qiao, D.; Bai, L. Isolation of a FAD-GPDH gene encoding a mitochondrial FAD-dependent glycerol-3-phosphate dehydrogenase from Dunaliella salina. J. Basic. Microbiol. 2007, 47, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Lan, Y.; Cao, X.; Yao, H.; Qiao, D.; Xu, H.; Cao, Y. Characterization and diverse evolution patterns of glycerol-3-phosphate dehydrogenase family genes in Dunaliella salina. Gene 2019, 710, 161–169. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Toh, J.D.; Ero, R.; Qiao, Z.; Kumar, V.; Serra, A.; Tan, J.; Sze, S.K.; Gao, Y.-G. The unusual di-domain structure of Dunaliella salina glycerol-3-phosphate dehydrogenase enables direct conversion of dihydroxyacetone phosphate to glycerol. Plant J. 2020, 102, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; He, L.-H.; Yu, T.-Y. Molecular clone and expression of a NAD+-dependent glycerol-3-phosphate dehydrogenase isozyme gene from the halotolerant alga Dunaliella salina. PLoS ONE 2013, 8, e62287. [Google Scholar] [CrossRef]
- Shen, W.; Wei, Y.; Dauk, M.; Zheng, Z.; Zou, J. Identification of a mitochondrial glycerol-3-phosphate dehydrogenase from Arabidopsis thaliana: Evidence for a mitochondrial glycerol-3-phosphate shuttle in plants. FEBS Lett. 2003, 536, 92–96. [Google Scholar] [CrossRef]
- Mráček, T.; Drahota, Z.; Houštěk, J. The function and the role of the mitochondrial glycerol-3-phosphate dehydrogenase in mammalian tissues. Biochim. Biophys. Acta 2013, 1827, 401–410. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, X.; Zhang, Z.; Pan, W.; Li, S.; Xing, Y.; Xin, W.; Zhang, Z.; Hu, Z.; Liu, C.; et al. GmGPDH12, a mitochondrial FAD-GPDH from soybean, increases salt and osmotic stress resistance by modulating redox state and respiration. Crop J. 2021, 9, 79–94. [Google Scholar] [CrossRef]
- Tomasko, D.A.; Edwards, J.K.; Meyer, H.A.; Lawrence, J.M. Variation of glyceraldehyde-3-phosphate dehydrogenase activity in response to reduced salinity in the red alga Gracilaria verrucosa (Hudson) papenfuss (Gigartinales, rhodophyta). Comp. Biochem. Physiol. B 1987, 87, 843–845. [Google Scholar] [CrossRef]
- Xia, B.-B.; Wang, S.-H.; Duan, J.-B.; Bai, L.-H. The relationship of glycerol and glycolysis metabolism patway under hyperosmotic stress in Dunaliella salina. Open Life Sci. 2014, 9, 901–908. [Google Scholar] [CrossRef]
- Goloubinoff, P.; Sassi, A.S.; Fauvet, B.; Barducci, A.; de Los Rios, P. Chaperones convert the energy from ATP into the nonequilibrium stabilization of native proteins. Nat. Chem. Biol. 2018, 14, 388–395. [Google Scholar] [CrossRef]
- Nirody, J.A.; Budin, I.; Rangamani, P. ATP synthase: Evolution, energetics, and membrane interactions. J. Gen. Physiol. 2020, 152, e201912475. [Google Scholar] [CrossRef] [PubMed]
- Jacob, P.; Hirt, H.; Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Fu, X. Chaperone function and mechanism of small heat-shock proteins. Acta Biochim. Biophys. Sin. 2014, 46, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Romero, P.; Gabrielli, A.; Sampedro, R.; Perea-García, A.; Puig, S.; Lafuente, M.T. Identification and molecular characterization of the high-affinity copper transporters family in Solanum lycopersicum. Int. J. Biol. Macromol. 2021, 192, 600–610. [Google Scholar] [CrossRef]
- Rocha, G.S.; Parrish, C.C.; Espíndola, E.L. Effects of copper on photosynthetic and physiological parameters of a freshwater microalga (Chlorophyceae). Algal Res. 2021, 54, 102223. [Google Scholar] [CrossRef]
- Mousavi, S.A.R.; Dubin, A.E.; Zeng, W.-Z.; Coombs, A.M.; Do, K.; Ghadiri, D.A.; Keenan, W.T.; Ge, C.; Zhao, Y.; Patapoutian, A. PIEZO ion channel is required for root mechanotransduction in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2021, 118, e2102188118. [Google Scholar] [CrossRef]
- Hoffmann, T.; Schütz, A.; Brosius, M.; Völker, A.; Völker, U.; Bremer, E. High-Salinity-Induced Iron Limitation in Bacillus subtilis. J. Bacteriol. 2002, 184, 718–727. [Google Scholar] [CrossRef]
- Fisher, M.; Gokhman, I.; Pick, U.; Zamir, A. A structurally novel transferrin-like protein accumulates in the plasma membrane of the unicellular green alga Dunaliella salina grown in high salinities. J. Biol. Chem. 1997, 272, 1565–1570. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, H.; Cai, C.; Lin, Z.; Zhen, Z.; Chu, J.; Guo, K. Histone acetyltransferase GCN5-mediated lysine acetylation modulates salt stress aadaption of Trichoderma. Appl. Microbiol. Biotechnol. 2022, 106, 3033–3049. [Google Scholar] [CrossRef]
- Ben Amor, F.; Elleuch, F.; Ben Hlima, H.; Garnier, M.; Saint-Jean, B.; Barkallah, M.; Pichon, C.; Abdelkafi, S.; Fendri, I. Proteomic Analysis of the Chlorophyta Dunaliella New Strain AL-1 Revealed Global Changes of Metabolism during High Carotenoid Production. Mar. Drugs 2017, 15, 293. [Google Scholar] [CrossRef]
- Livanos, P.; Galatis, B.; Apostolakos, P. The interplay between ROS and tubulin cytoskeleton in plants. Plant Signal. Behav. 2014, 9, e28069. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A.; Siva, C.J. The taxonomy of the genus Dunaliella (Chlorophyta, Dunaliellales) with emphasis on the marine and halophilic species. J. Appl. Phycol. 2007, 19, 567–590. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Shetty, P.; Gitau, M.M.; Maróti, G. Salinity Stress Responses and Adaptation Mechanisms in Eukaryotic Green Microalgae. Cells 2019, 8, 1657. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, L.J.; Brown, A.D. The salt relations of marine and halophilic species of the unicellular green alga, Dunaliella. The role of glycerol as a compatible solute. Arch. Mikrobiol. 1974, 96, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, D.J.; Hipkins, M.F.; Webber, A.N.; Baker, N.R.; Boney, A.D. The effect of ionic stress on photosynthesis in Dunaliella tertiolecta: Chlorophyll fluorescence kinetics and spectral characteristics. Planta 1985, 163, 250–256. [Google Scholar] [CrossRef]
- Highfield, A.; Ward, A.; Pipe, R.; Schroeder, D.C. Molecular and phylogenetic analysis reveals new diversity of Dunaliella salina from hypersaline environments. J. Mar. Biol. Ass. 2021, 101, 27–37. [Google Scholar] [CrossRef]
- Dehghani, J.; Atazadeh, E.; Omidi, Y.; Movafeghi, A. The use of 18S ribosomal DNA, ITS and rbcL molecular markers to study the genus Dunaliella (Dunaliellaceae) in Iranian samples: A phylogenetic approach. Oceanol. Hydrobiol. Stud. 2019, 49, 88–98. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Borowitzka, L.J. Micro-Algal Biotechnology; Cambridge University Press: New York, NY, USA, 1988; ISBN 0521323495. [Google Scholar]
- Woortman, D.V.; Fuchs, T.; Striegel, L.; Fuchs, M.; Weber, N.; Brück, T.B.; Rychlik, M. Microalgae a Superior Source of Folates: Quantification of Folates in Halophile Microalgae by Stable Isotope Dilution Assay. Front. Bioeng. Biotechnol. 2019, 7, 481. [Google Scholar] [CrossRef]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef]
- Kamil, I.; Talha, G.; Fadime, K.; Elif, C. Molecular identification of different actinomycetes isolated from East Black Sea region plateau soil by 16S rDNA gene sequencing. Afr. J. Microbiol. Res. 2014, 8, 878–887. [Google Scholar] [CrossRef]
- Healey, A.; Furtado, A.; Cooper, T.; Henry, R.J. Protocol: A simple method for extracting next-generation sequencing quality genomic DNA from recalcitrant plant species. Plant Methods 2014, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Inglis, P.W.; Pappas, M.d.C.R.; Resende, L.V.; Grattapaglia, D. Fast and inexpensive protocols for consistent extraction of high quality DNA and RNA from challenging plant and fungal samples for high-throughput SNP genotyping and sequencing applications. PLoS ONE 2018, 13, e0206085. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.O.; Bendich, A.J. Extraction of total cellular DNA from plants, algae and fungi. In Plant Molecular Biology Manual; Gelvin, S.B., Schilperoort, R.A., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 183–190. ISBN 978-94-011-7654-5. [Google Scholar]
- Ritz, M.; Ahmad, N.; Brueck, T.; Mehlmer, N. Comparative Genome-Wide Analysis of Two Caryopteris x Clandonensis Cultivars: Insights on the Biosynthesis of Volatile Terpenoids. Plants 2023, 12, 632. [Google Scholar] [CrossRef]
- GitHub—PacificBiosciences/pbipa: Improved Phased Assembler. Available online: https://github.com/PacificBiosciences/pbipa (accessed on 24 May 2023).
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Nachtigall, P.G.; Kashiwabara, A.Y.; Durham, A.M. CodAn: Predictive models for precise identification of coding regions in eukaryotic transcripts. Brief. Bioinform. 2021, 22, bbaa045. [Google Scholar] [CrossRef]
- Buchfink, B.; Reuter, K.; Drost, H.-G. Sensitive protein alignments at tree-of-life scale using DIAMOND. Nat. Methods 2021, 18, 366–368. [Google Scholar] [CrossRef]
- BlastKOALA: Automatic KO Assignment and KEGG Mapping Service. Available online: https://www.kegg.jp/blastkoala/ (accessed on 24 May 2023).
- Mercator4: Protein Function Mapping. Available online: https://www.plabipd.de/mercator_main.html (accessed on 24 May 2023).
- eggNOG-mapper: Genome-Wide Functunal Annotation. Available online: http://eggnog-mapper.embl.de/ (accessed on 18 September 2023).
- CateGOrizer: Previously Known as “GO Terms Classifications Counter”. Available online: https://www.animalgenome.org/tools/catego/ (accessed on 24 May 2023).
- Kruger, N.J. The Bradford method for protein quantitation. Methods Mol. Biol. 1994, 32, 9–15. [Google Scholar] [CrossRef]
- Fuchs, T.; Melcher, F.; Rerop, Z.S.; Lorenzen, J.; Shaigani, P.; Awad, D.; Haack, M.; Prem, S.A.; Masri, M.; Mehlmer, N.; et al. Identifying carbohydrate-active enzymes of Cutaneotrichosporon oleaginosus using systems biology. Microb. Cell Fact. 2021, 20, 205. [Google Scholar] [CrossRef]
- Engelhart-Straub, S.; Cavelius, P.; Hölzl, F.; Haack, M.; Awad, D.; Brueck, T.; Mehlmer, N. Effects of Light on Growth and Metabolism of Rhodococcus erythropolis. Microorganisms 2022, 10, 1680. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.H.; Qiao, R.; Xin, L.; Chen, X.; Liu, C.; Zhang, X.; Shan, B.; Ghodsi, A.; Li, M. Deep learning enables de novo peptide sequencing from data-independent-acquisition mass spectrometry. Nat. Methods 2019, 16, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.H.; Rahman, M.Z.; He, L.; Xin, L.; Shan, B.; Li, M. Complete De Novo Assembly of Monoclonal Antibody Sequences. Sci. Rep. 2016, 6, 31730. [Google Scholar] [CrossRef]
- Tran, N.H.; Zhang, X.; Xin, L.; Shan, B.; Li, M. De novo peptide sequencing by deep learning. Proc. Natl. Acad. Sci. USA 2017, 114, 8247–8252. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | 30 min | 4 h | 24 h |
|---|---|---|---|
| galactokinase | 11.46 | 3.30 | - |
| ADP-ribosylation Crystallin J1. Source PGD | 8.01 | - | - |
| HAT (Half-A-TPR) repeats | 7.85 | 3.60 | - |
| Snf1-related protein kinase 1 (SnRK1) | 5.56 | - | - |
| nuclear protein 96 | 5.53 | - | - |
| belongs to the actin family | 4.93 | - | - |
| translocation protein Sec62 | 4.88 | - | - |
| rieske-like [2Fe-2S] domain | 4.67 | - | - |
| photosystem II Pbs27 | - | 11.05 | - |
| sulfhydryl oxidase ALR/ERV | - | 9.21 | - |
| mitochondrial Transcription Termination Factors (MTERF) superfamily | - | 7.43 | - |
| thiamin pyrophosphokinase, vitamin B1 binding domain | - | 6.40 | - |
| EF hand family protein | - | 4.61 | - |
| adaptin C-terminal domain | - | 4.45 | - |
| CP12 | 2.50 | 4.25 | 2.48 |
| thioredoxin-like domain | - | 4.03 | 2.44 |
| ELMO/CED-12 family | - | - | 31.89 |
| glutamate dehydrogenase | - | - | 23.02 |
| 7,8-dihydroneopterin aldolase/epimerase/oxygenase | - | - | 12.17 |
| surfeit locus protein 6 | 3.63 | - | 12.09 |
| ketohexokinase activity | - | - | 9.45 |
| biotin and thiamin Synthesis-associated domain | - | - | 8.97 |
| large subunit ribosomal protein L6 | - | - | 6.51 |
| large subunit ribosomal protein L11e | - | - | 6.17 |
| Enzyme | 30 min | 4 h | 24 h |
|---|---|---|---|
| protein-l-isoaspartate O-methyltransferase | 0.24 | - | - |
| ribosomal protein L7 L12 | 0.24 | 0.31 | - |
| RNA-binding protein with multiple splicing | 0.32 | - | 0.29 |
| belongs to the DEAD box helicase family | 0.42 | - | - |
| acyl carrier protein-like protein | 0.44 | - | 0.25 |
| AIG1 family | - | 0.05 | - |
| glutathione S-transferase | - | 0.13 | |
| phosphomannomutase | - | 0.15 | 0.47 |
| serine threonine-protein kinase | - | 0.15 | - |
| enoyl-(Acyl carrier protein) reductase | - | 0.16 | - |
| chloroplast envelope transporter | - | - | 0.11 |
| Isy1-like splicing family | - | - | 0.12 |
| Sugar (and other) transporter | - | - | 0.20 |
| photosystem ii reaction center w | - | - | 0.31 |
| sigma 54 modulation protein/S30EA ribosomal protein | - | - | 0.31 |
| Enzyme | 30 min | 4 h | 24 h |
|---|---|---|---|
| chlorophyll A-B binding protein | 2.66 | 2.31 | 0.45 |
| major light-harvesting chlorophyll a/b protein 3 | 3.3 | - | 2.02 |
| photosystem II biogenesis protein Psp29 | 1.67 | - | - |
| photosystem II subunit 28 | 2.21 | - | - |
| photosystem I assembly | 2.60 | 1.63 | - |
| the polypeptide of the oxygen-evolving complex of photosystem II | 2.27 | - | - |
| photosystem I light-harvesting chlorophyll-a/b protein 2 | - | 1.75 | - |
| plastocyanin/azurin family-domain-containing protein | - | 3.10 | 2.80 |
| photosystem II Pbs27 | - | 11.05 | - |
| photosystem ii reaction center w | - | - | 0.31 |
| Enzyme | 30 min | 4 h | 24 h |
|---|---|---|---|
| small molecular HSP 10 | 2.43 | 2.66 | 2.54 |
| HSP20-like chaperone | 3.04 | - | - |
| cold-shock protein domain | 2.05 | - | - |
| HSP 70C | - | 0.42 | - |
| heat-shock chaperonin-binding motif | - | 1.94 | 1.79 |
| activator of Hsp90 ATPase | - | 2.49 | |
| zinc chaperone CobW-like | - | 5.24 | - |
| HSP20-like chaperone | - | - | 2.24 |
| DNAJ-like protein/HSP 40 | - | - | 2.88 |
| Enzyme | 30 min | 4 h | 24 h |
|---|---|---|---|
| voltage-dependent anion channel protein 2 | 2.42 | - | - |
| Ctr copper transporter family | - | 2.34 | 2.27 |
| potassium:proton antiporter activity | - | 3.64 | - |
| ZIP Zinc transporter | - | 2.36 | - |
| polycystin cation channel | - | 2.09 | - |
| PIEZO non-specific cation channel | - | 3.03 | - |
| chloroplast envelope transporter | - | - | 0.11 |
| major facilitator superfamily domain-containing protein | - | - | 0.20 |
| Enzyme | 30 min | 4 h | 24 h | Function |
|---|---|---|---|---|
| glutamate receptor (GLR) | P | - | - | Ligand-binding and ion channel activity: Involved in Ca2+ signaling processes |
| sodium hydrogen exchanger | P | - | - | Intracellular regulation of pH and Na+ concentration |
| 26S proteasome non-ATPase regulatory subunit 4 (PSMD4) | P | - | - | Protein degradation |
| galactokinase | 11.46 | - | - | Phosphorylation of galactose to galactose-1-phosphate |
| Snf1-like protein kinase (SnRK1) | 5.56 | - | - | Plant equivalent of the heterotrimeric AMPK/Snf. Mediates the crosstalk between ABA and salt stress signaling |
| PIEZO | - | 3.03 | - | Non-selective cation channel. Mechanical stress-induced Ca2+ signaling |
| calcium/calmodulin-dependent protein kinases | 3.42 | 2.38, 4.61 | 0.37 | Serine/threonine kinase activated by increasing Ca2+ concentration |
| CP12 | 2.50 | 4.25 | 2.48 | Protecting cells against oxidative stress and suppressing the production of ROS. Control of electron flow from Photosystem I to NADPH |
| gamma carbonic anhydrase 3 | 1.75 | - | - | CO2 uptake |
| carbonic anhydrase | - | - | 1.87 | CO2 uptake |
| fructose-bisphosphatase | 2.34 | - | 2.93 | Calvin–Benson cycle |
| sedoheptulose-bisphosphatase | 3.66 | - | - | |
| glyceraldehyde 3-phosphate | 2.78 | - | 1.85 | |
| ribulose 1,5-bisphosphate carboxylase/oxygenase (Rubisco) | - | - | 2.95 | |
| phosphofructokinase (PFK) | - | - | 2.26 | Conversion of glucose to fructose-1, 6-bisphosphate |
| engulfment and cell motility (ELMO) protein | - | - | 31.89 | Regulates cytoskeletal reorganization, cell apoptosis, and cell migration |
| glycerol-3-phosphate dehydrogenase (G3pdh) with di-domain | - | - | - | Direct conversion of DHAP to glycerol |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keil, L.; Mehlmer, N.; Cavelius, P.; Garbe, D.; Haack, M.; Ritz, M.; Awad, D.; Brück, T. The Time-Resolved Salt Stress Response of Dunaliella tertiolecta—A Comprehensive System Biology Perspective. Int. J. Mol. Sci. 2023, 24, 15374. https://doi.org/10.3390/ijms242015374
Keil L, Mehlmer N, Cavelius P, Garbe D, Haack M, Ritz M, Awad D, Brück T. The Time-Resolved Salt Stress Response of Dunaliella tertiolecta—A Comprehensive System Biology Perspective. International Journal of Molecular Sciences. 2023; 24(20):15374. https://doi.org/10.3390/ijms242015374
Chicago/Turabian StyleKeil, Linda, Norbert Mehlmer, Philipp Cavelius, Daniel Garbe, Martina Haack, Manfred Ritz, Dania Awad, and Thomas Brück. 2023. "The Time-Resolved Salt Stress Response of Dunaliella tertiolecta—A Comprehensive System Biology Perspective" International Journal of Molecular Sciences 24, no. 20: 15374. https://doi.org/10.3390/ijms242015374
APA StyleKeil, L., Mehlmer, N., Cavelius, P., Garbe, D., Haack, M., Ritz, M., Awad, D., & Brück, T. (2023). The Time-Resolved Salt Stress Response of Dunaliella tertiolecta—A Comprehensive System Biology Perspective. International Journal of Molecular Sciences, 24(20), 15374. https://doi.org/10.3390/ijms242015374