
Dopamine D4 Receptor Agonist Drastically Increases Delta Activity in the Thalamic Nucleus Reuniens: Potential Role in Communication between Prefrontal Cortex and Hippocampus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Delta Oscillations during Waking
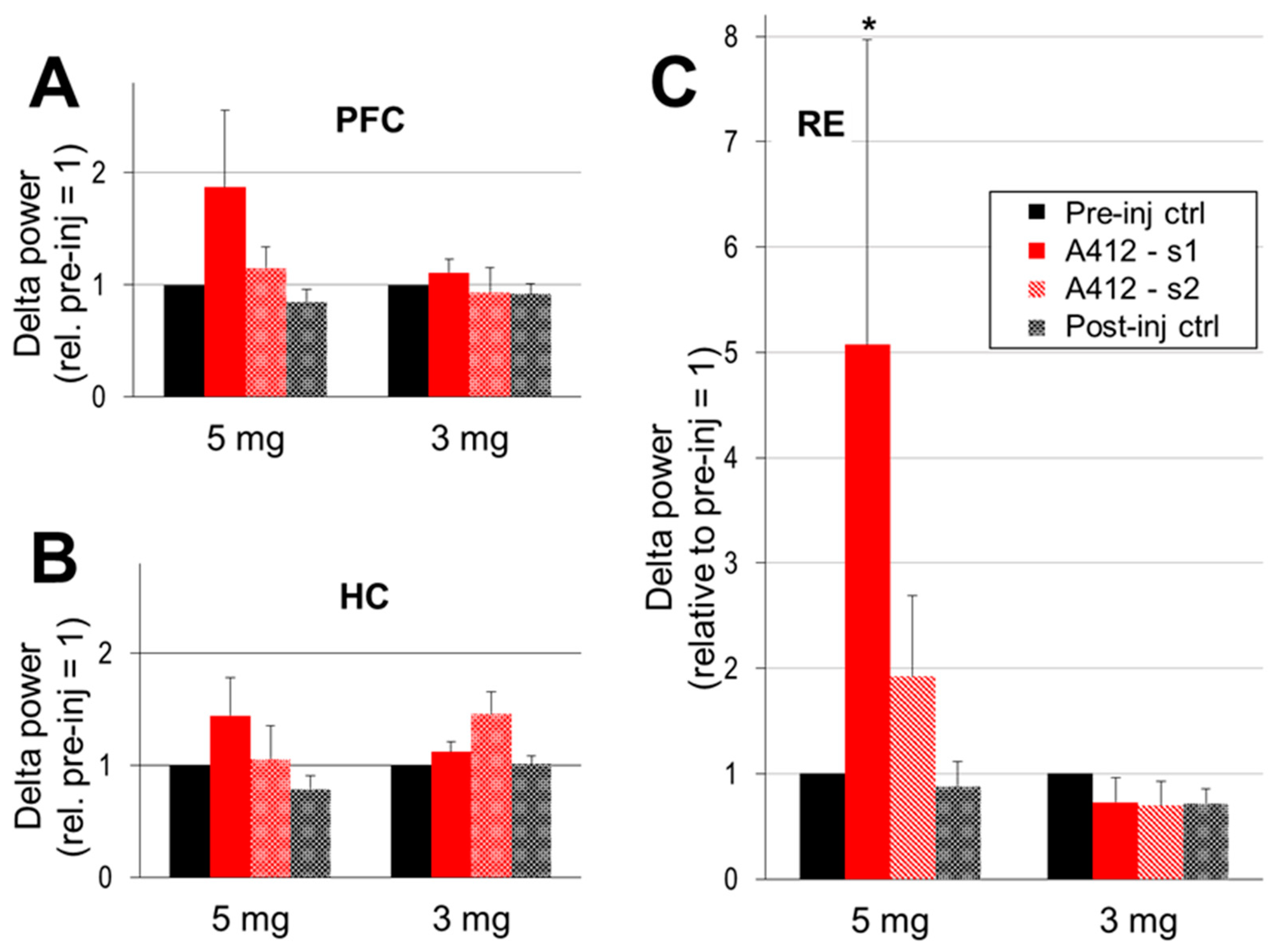
2.2. The Effect of Dopamine D4 Receptor Activation on Delta Activity in the PFC, HC, and RE

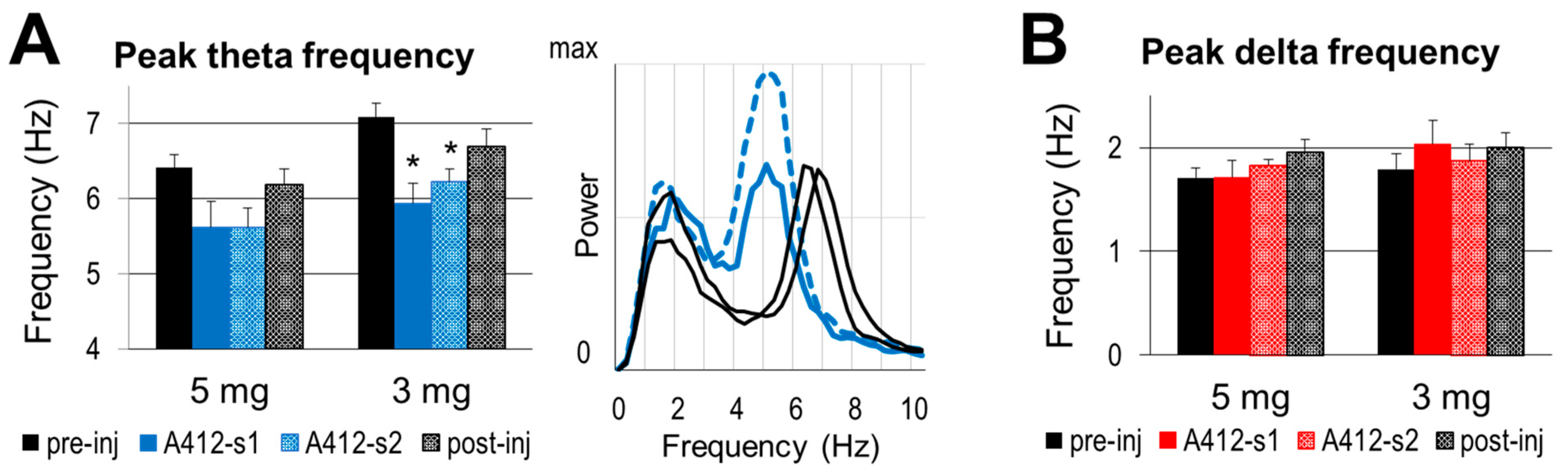
2.3. Effect of Dopamine D4 Receptor Activation on Theta Activity


3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Surgery
4.4. The Identification of Electrode Position
4.5. Data Recording
4.6. Data and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lanre-Amos, T.; Kocsis, B. Hippocampal oscillations in the rodent model of schizophrenia induced by amygdala GABA receptor blockade. Front. Psychiatry 2010, 1, 132. [Google Scholar] [CrossRef]
- Pittman-Polletta, B.; Hu, K.; Kocsis, B. Subunit-specific NMDAR antagonism dissociates schizophrenia subtype-relevant oscillopathies associated with frontal hypofunction and hippocampal hyperfunction. Sci. Rep. 2018, 8, 11588. [Google Scholar] [CrossRef]
- Pittman-Polletta, B.R.; Kocsis, B.; Vijayan, S.; Whittington, M.A.; Kopell, N.J. Brain rhythms connect impaired inhibition to altered cognition in schizophrenia. Biol. Psychiatry 2015, 77, 1020–1030. [Google Scholar] [CrossRef][Green Version]
- Brown, J.; Grayson, B.; Neill, J.C.; Harte, M.; Wall, M.J.; Ngomba, R.T. Oscillatory Deficits in the Sub-Chronic PCP Rat Model for Schizophrenia Are Reversed by mGlu5 Receptor-Positive Allosteric Modulators VU0409551 and VU0360172. Cells 2023, 12, 919. [Google Scholar] [CrossRef]
- Cordon, I.; Nicolas, M.J.; Arrieta, S.; Lopetegui, E.; Lopez-Azcarate, J.; Alegre, M.; Artieda, J.; Valencia, M. Coupling in the cortico-basal ganglia circuit is aberrant in the ketamine model of schizophrenia. Eur. Neuropsychopharmacol. 2015, 25, 1375–1387. [Google Scholar] [CrossRef] [PubMed]
- Goda, S.A.; Olszewski, M.; Piasecka, J.; Rejniak, K.; Whittington, M.A.; Kasicki, S.; Hunt, M.J. Aberrant high frequency oscillations recorded in the rat nucleus accumbens in the methylazoxymethanol acetate neurodevelopmental model of schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2015, 61, 44–51. [Google Scholar] [CrossRef]
- Jenkins, B.W.; Buckhalter, S.; Perreault, M.L.; Khokhar, J.Y. Cannabis Vapor Exposure Alters Neural Circuit Oscillatory Activity in a Neurodevelopmental Model of Schizophrenia: Exploring the Differential Impact of Cannabis Constituents. Schizophr. Bull. Open 2022, 3, sgab052. [Google Scholar] [CrossRef]
- Kaefer, K.; Malagon-Vina, H.; Dickerson, D.D.; O’Neill, J.; Trossbach, S.V.; Korth, C.; Csicsvari, J. Disrupted-in-schizophrenia 1 overexpression disrupts hippocampal coding and oscillatory synchronization. Hippocampus 2019, 29, 802–816. [Google Scholar] [CrossRef] [PubMed]
- Rosen, A.M.; Spellman, T.; Gordon, J.A. Electrophysiological endophenotypes in rodent models of schizophrenia and psychosis. Biol. Psychiatry 2015, 77, 1041–1049. [Google Scholar] [CrossRef]
- Young, A.M.; Stubbendorff, C.; Valencia, M.; Gerdjikov, T.V. Disruption of medial prefrontal synchrony in the subchronic phencyclidine model of schizophrenia in rats. Neuroscience 2015, 287, 157–163. [Google Scholar] [CrossRef]
- Hajos, M.; Hoffmann, W.E.; Kocsis, B. Activation of cannabinoid-1 receptors disrupts sensory gating and neuronal oscillation: Relevance to schizophrenia. Biol. Psychiatry 2008, 63, 1075–1083. [Google Scholar] [CrossRef]
- Kittelberger, K.; Hur, E.E.; Sazegar, S.; Keshavan, V.; Kocsis, B. Comparison of the effects of acute and chronic administration of ketamine on hippocampal oscillations: Relevance for the NMDA receptor hypofunction model of schizophrenia. Brain Struct. Funct. 2012, 217, 395–409. [Google Scholar] [CrossRef]
- Sorman, E.; Wang, D.; Hajos, M.; Kocsis, B. Control of hippocampal theta rhythm by serotonin: Role of 5-HT2c receptors. Neuropharmacology 2011, 61, 489–494. [Google Scholar] [CrossRef]
- Kocsis, B.; Lee, P.; Deth, R. Enhancement of gamma activity after selective activation of dopamine D4 receptors in freely moving rats and in a neurodevelopmental model of schizophrenia. Brain Struct. Funct. 2014, 219, 2173–2180. [Google Scholar] [CrossRef]
- Li, S.; Topchiy, I.; Kocsis, B. The effect of atropine administered in the medial septum or hippocampus on high- and low-frequency theta rhythms in the hippocampus of urethane anesthetized rats. Synapse 2007, 61, 412–419. [Google Scholar] [CrossRef]
- Ly, S.; Pishdari, B.; Lok, L.L.; Hajos, M.; Kocsis, B. Activation of 5-HT6 receptors modulates sleep-wake activity and hippocampal theta oscillation. ACS Chem. Neurosci. 2013, 4, 191–199. [Google Scholar] [CrossRef]
- Andersson, R.; Johnston, A.; Fisahn, A. Dopamine D4 receptor activation increases hippocampal gamma oscillations by enhancing synchronization of fast-spiking interneurons. PLoS ONE 2012, 7, e40906. [Google Scholar] [CrossRef]
- Andersson, R.H.; Johnston, A.; Herman, P.A.; Winzer-Serhan, U.H.; Karavanova, I.; Vullhorst, D.; Fisahn, A.; Buonanno, A. Neuregulin and dopamine modulation of hippocampal gamma oscillations is dependent on dopamine D4 receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 13118–13123. [Google Scholar] [CrossRef]
- Ebstein, R.P.; Nemanov, L.; Klotz, I.; Gritsenko, I.; Belmaker, R.H. Additional evidence for an association between the dopamine D4 receptor (D4DR) exon III repeat polymorphism and the human personality trait of Novelty Seeking. Mol. Psychiatry 1997, 2, 472–477. [Google Scholar] [CrossRef]
- Munafo, M.R.; Yalcin, B.; Willis-Owen, S.A.; Flint, J. Association of the dopamine D4 receptor (DRD4) gene and approach-related personality traits: Meta-analysis and new data. Biol. Psychiatry 2008, 63, 197–206. [Google Scholar] [CrossRef]
- LaHoste, G.J.; Swanson, J.M.; Wigal, S.B.; Glabe, C.; Wigal, T.; King, N.; Kennedy, J.L. Dopamine D4 receptor gene polymorphism is associated with attention deficit hyperactivity disorder. Mol. Psychiatry 1996, 1, 121–124. [Google Scholar]
- Ferre, S.; Belcher, A.M.; Bonaventura, J.; Quiroz, C.; Sanchez-Soto, M.; Casado-Anguera, V.; Cai, N.S.; Moreno, E.; Boateng, C.A.; Keck, T.M.; et al. Functional and pharmacological role of the dopamine D(4) receptor and its polymorphic variants. Front. Endocrinol. 2022, 13, 1014678. [Google Scholar] [CrossRef]
- Asghari, V.; Sanyal, S.; Buchwaldt, S.; Paterson, A.; Jovanovic, V.; Van Tol, H.H. Modulation of intracellular cyclic AMP levels by different human dopamine D4 receptor variants. J. Neurochem. 1995, 65, 1157–1165. [Google Scholar] [CrossRef]
- Hodgson, N.W.; Waly, M.I.; Trivedi, M.S.; Power-Charnitsky, V.A.; Deth, R.C. Methylation-related metabolic effects of D4 dopamine receptor expression and activation. Transl. Psychiatry 2019, 9, 295. [Google Scholar] [CrossRef]
- Sharma, A.; Kramer, M.L.; Wick, P.F.; Liu, D.; Chari, S.; Shim, S.; Tan, W.; Ouellette, D.; Nagata, M.; DuRand, C.J.; et al. D4 dopamine receptor-mediated phospholipid methylation and its implications for mental illnesses such as schizophrenia. Mol. Psychiatry 1999, 4, 235–246. [Google Scholar] [CrossRef][Green Version]
- Deth, R.C. Autism: A redox/methylation disorder. Glob. Adv. Health Med. 2013, 2, 68–73. [Google Scholar] [CrossRef]
- Reynolds, G.P.; Zhang, Z.J.; Beasley, C.L. Neurochemical correlates of cortical GABAergic deficits in schizophrenia: Selective losses of calcium binding protein immunoreactivity. Brain Res. Bull. 2001, 55, 579–584. [Google Scholar] [CrossRef]
- Sohal, V.S.; Zhang, F.; Yizhar, O.; Deisseroth, K. Parvalbumin neurons and gamma rhythms enhance cortical circuit performance. Nature 2009, 459, 698–702. [Google Scholar] [CrossRef]
- Kuznetsova, A.Y.; Deth, R.C. A model for modulation of neuronal synchronization by D4 dopamine receptor-mediated phospholipid methylation. J. Comput. Neurosci. 2008, 24, 314–329. [Google Scholar] [CrossRef]
- Thorn, C.W.; Kafetzopoulos, V.; Kocsis, B. Differential Effect of Dopamine D4 Receptor Activation on Low-Frequency Oscillations in the Prefrontal Cortex and Hippocampus May Bias the Bidirectional Prefrontal-Hippocampal Coupling. Int. J. Mol. Sci. 2022, 23, 11705. [Google Scholar] [CrossRef]
- Nandi, B.; Swiatek, P.; Kocsis, B.; Ding, M. Inferring the direction of rhythmic neural transmission via inter-regional phase-amplitude coupling (ir-PAC). Sci. Rep. 2019, 9, 6933. [Google Scholar] [CrossRef]
- Cardoso-Cruz, H.; Paiva, P.; Monteiro, C.; Galhardo, V. Bidirectional optogenetic modulation of prefrontal-hippocampal connectivity in pain-related working memory deficits. Sci. Rep. 2019, 9, 10980. [Google Scholar] [CrossRef]
- Helfrich, R.F.; Lendner, J.D.; Mander, B.A.; Guillen, H.; Paff, M.; Mnatsakanyan, L.; Vadera, S.; Walker, M.P.; Lin, J.J.; Knight, R.T. Bidirectional prefrontal-hippocampal dynamics organize information transfer during sleep in humans. Nat. Commun. 2019, 10, 3572. [Google Scholar] [CrossRef]
- Jayachandran, M.; Viena, T.D.; Garcia, A.; Veliz, A.V.; Leyva, S.; Roldan, V.; Vertes, R.P.; Allen, T.A. Nucleus reuniens transiently synchronizes memory networks at beta frequencies. Nat. Commun. 2023, 14, 4326. [Google Scholar] [CrossRef]
- Goto, Y.; Grace, A.A. Dopamine-dependent interactions between limbic and prefrontal cortical plasticity in the nucleus accumbens: Disruption by cocaine sensitization. Neuron 2005, 47, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Cassel, J.C.; Pereira de Vasconcelos, A.; Loureiro, M.; Cholvin, T.; Dalrymple-Alford, J.C.; Vertes, R.P. The reuniens and rhomboid nuclei: Neuroanatomy, electrophysiological characteristics and behavioral implications. Prog. Neurobiol. 2013, 111, 34–52. [Google Scholar] [CrossRef]
- Vertes, R.P. Major diencephalic inputs to the hippocampus: Supramammillary nucleus and nucleus reuniens. Circuitry and function. Prog. Brain Res. 2015, 219, 121–144. [Google Scholar]
- Dolleman-van der Weel, M.J.; Griffin, A.L.; Ito, H.T.; Shapiro, M.L.; Witter, M.P.; Vertes, R.P.; Allen, T.A. The nucleus reuniens of the thalamus sits at the nexus of a hippocampus and medial prefrontal cortex circuit enabling memory and behavior. Learn. Mem. 2019, 26, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Schlecht, M.; Jayachandran, M.; Rasch, G.E.; Allen, T.A. Dual projecting cells linking thalamic and cortical communication routes between the medial prefrontal cortex and hippocampus. Neurobiol. Learn. Mem. 2022, 188, 107586. [Google Scholar] [CrossRef]
- Vertes, R.P.; Hoover, W.B.; Szigeti-Buck, K.; Leranth, C. Nucleus reuniens of the midline thalamus: Link between the medial prefrontal cortex and the hippocampus. Brain Res. Bull. 2007, 71, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, E.C.; Grace, A.A. Prefrontal cortex modulates firing pattern in the nucleus reuniens of the midline thalamus via distinct corticothalamic pathways. Eur. J. Neurosci. 2018, 48, 3255–3272. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, E.C.; Grace, A.A. The Nucleus Reuniens of the Midline Thalamus Gates Prefrontal-Hippocampal Modulation of Ventral Tegmental Area Dopamine Neuron Activity. J. Neurosci. 2016, 36, 8977–8984. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y. Theta frequency prefrontal-hippocampal driving relationship during free exploration in mice. Neuroscience 2015, 300, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Oberlander, V.C.; Xu, X.; Chini, M.; Hanganu-Opatz, I.L. Developmental dysfunction of prefrontal-hippocampal networks in mouse models of mental illness. Eur. J. Neurosci. 2019, 50, 3072–3084. [Google Scholar] [CrossRef]
- Kafetzopoulos, V.; Kokras, N.; Sotiropoulos, I.; Oliveira, J.F.; Leite-Almeida, H.; Vasalou, A.; Sardinha, V.M.; Papadopoulou-Daifoti, Z.; Almeida, O.F.X.; Antoniou, K.; et al. The nucleus reuniens: A key node in the neurocircuitry of stress and depression. Mol. Psychiatry 2018, 23, 579–586. [Google Scholar] [CrossRef]
- Hunt, M.J.; Kopell, N.J.; Traub, R.D.; Whittington, M.A. Aberrant Network Activity in Schizophrenia. Trends Neurosci. 2017, 40, 371–382. [Google Scholar] [CrossRef]
- Roy, A.; Svensson, F.P.; Mazeh, A.; Kocsis, B. Prefrontal-hippocampal coupling by theta rhythm and by 2–5 Hz oscillation in the delta band: The role of the nucleus reuniens of the thalamus. Brain Struct. Funct. 2017, 222, 2819–2830. [Google Scholar] [CrossRef]
- Jung, F.; Witte, V.; Yanovsky, Y.; Klumpp, M.; Brankack, J.; Tort, A.B.L.; Draguhn, A. Differential modulation of parietal cortex activity by respiration and theta oscillations. J. Neurophysiol. 2022, 127, 801–817. [Google Scholar] [CrossRef]
- Rajji, T.K.; Zomorrodi, R.; Barr, M.S.; Blumberger, D.M.; Mulsant, B.H.; Daskalakis, Z.J. Ordering Information in Working Memory and Modulation of Gamma by Theta Oscillations in Humans. Cereb. Cortex 2017, 27, 1482–1490. [Google Scholar] [CrossRef]
- Takahashi, M.; Nishida, H.; Redish, A.D.; Lauwereyns, J. Theta phase shift in spike timing and modulation of gamma oscillation: A dynamic code for spatial alternation during fixation in rat hippocampal area CA1. J. Neurophysiol. 2014, 111, 1601–1614. [Google Scholar] [CrossRef]
- Dejean, C.; Arbuthnott, G.; Wickens, J.R.; Le Moine, C.; Boraud, T.; Hyland, B.I. Power fluctuations in beta and gamma frequencies in rat globus pallidus: Association with specific phases of slow oscillations and differential modulation by dopamine D1 and D2 receptors. J. Neurosci. 2011, 31, 6098–6107. [Google Scholar] [CrossRef]
- Fujisawa, S.; Buzsaki, G. A 4 Hz oscillation adaptively synchronizes prefrontal, VTA, and hippocampal activities. Neuron 2011, 72, 153–165. [Google Scholar] [CrossRef]
- Duan, A.R.; Varela, C.; Zhang, Y.; Shen, Y.; Xiong, L.; Wilson, M.A.; Lisman, J. Delta frequency optogenetic stimulation of the thalamic nucleus reuniens is sufficient to produce working memory deficits: Relevance to schizophrenia. Biol. Psychiatry 2015, 77, 1098–1107. [Google Scholar] [CrossRef] [PubMed]
- Pinault, D. N-methyl d-aspartate receptor antagonists ketamine and MK-801 induce wake-related aberrant gamma oscillations in the rat neocortex. Biol. Psychiatry 2008, 63, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Kocsis, B.; Brown, R.E.; McCarley, R.W.; Hajos, M. Impact of ketamine on neuronal network dynamics: Translational modeling of schizophrenia-relevant deficits. CNS Neurosci. Ther. 2013, 19, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Ahnaou, A.; Huysmans, H.; Van de Casteele, T.; Drinkenburg, W. Cortical high gamma network oscillations and connectivity: A translational index for antipsychotics to normalize aberrant neurophysiological activity. Transl. Psychiatry 2017, 7, 1285. [Google Scholar] [CrossRef]
- Kocsis, B. Differential role of NR2A and NR2B subunits in N-methyl-D-aspartate receptor antagonist-induced aberrant cortical gamma oscillations. Biol. Psychiatry 2012, 71, 987–995. [Google Scholar] [CrossRef]
- Wood, J.; Kim, Y.; Moghaddam, B. Disruption of prefrontal cortex large scale neuronal activity by different classes of psychotomimetic drugs. J. Neurosci. 2012, 32, 3022–3031. [Google Scholar] [CrossRef]
- Bernaerts, P.; Tirelli, E. Facilitatory effect of the dopamine D4 receptor agonist PD168,077 on memory consolidation of an inhibitory avoidance learned response in C57BL/6J mice. Behav. Brain Res. 2003, 142, 41–52. [Google Scholar] [CrossRef]
- Browman, K.E.; Curzon, P.; Pan, J.B.; Molesky, A.L.; Komater, V.A.; Decker, M.W.; Brioni, J.D.; Moreland, R.B.; Fox, G.B. A-412997, a selective dopamine D4 agonist, improves cognitive performance in rats. Pharmacol. Biochem. Behav. 2005, 82, 148–155. [Google Scholar] [CrossRef]
- Woolley, M.L.; Waters, K.A.; Reavill, C.; Bull, S.; Lacroix, L.P.; Martyn, A.J.; Hutcheson, D.M.; Valerio, E.; Bate, S.; Jones, D.N.; et al. Selective dopamine D4 receptor agonist (A-412997) improves cognitive performance and stimulates motor activity without influencing reward-related behaviour in rat. Behav. Pharmacol. 2008, 19, 765–776. [Google Scholar] [CrossRef]
- Shu, I.W.; Granholm, E.L.; Singh, F. Targeting Frontal Gamma Activity with Neurofeedback to Improve Working Memory in Schizophrenia. Curr. Top. Behav. Neurosci. 2023, 63, 153–172. [Google Scholar] [PubMed]
- White, R.S.; Siegel, S.J. Cellular and circuit models of increased resting-state network gamma activity in schizophrenia. Neuroscience 2016, 321, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, C.S.; Frund, I.; Lenz, D. Human gamma-band activity: A review on cognitive and behavioral correlates and network models. Neurosci. Biobehav. Rev. 2010, 34, 981–992. [Google Scholar] [CrossRef]
- Kaiser, J.; Lutzenberger, W. Human gamma-band activity: A window to cognitive processing. Neuroreport 2005, 16, 207–211. [Google Scholar] [CrossRef]
- Rieder, M.K.; Rahm, B.; Williams, J.D.; Kaiser, J. Human gamma-band activity and behavior. Int. J. Psychophysiol. 2011, 79, 39–48. [Google Scholar] [CrossRef]
- Ferbinteanu, J.; Kennedy, P.J.; Shapiro, M.L. Episodic memory—From brain to mind. Hippocampus 2006, 16, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Preston, A.R.; Eichenbaum, H. Interplay of hippocampus and prefrontal cortex in memory. Curr. Biol. 2013, 23, R764–R773. [Google Scholar] [CrossRef]
- Zhu, X.; Cabungcal, J.H.; Cuenod, M.; Uliana, D.L.; Do, K.Q.; Grace, A.A. Thalamic reticular nucleus impairments and abnormal prefrontal control of dopamine system in a developmental model of schizophrenia: Prevention by N-acetylcysteine. Mol. Psychiatry 2021, 26, 7679–7689. [Google Scholar] [CrossRef]
- Zhao, Y.; Liang, X.; Zhu, F.; Wen, Y.; Xu, J.; Yang, J.; Ding, M.; Cheng, B.; Ma, M.; Zhang, L.; et al. A large-scale integrative analysis of GWAS and common meQTLs across whole life course identifies genes, pathways and tissue/cell types for three major psychiatric disorders. Neurosci. Biobehav. Rev. 2018, 95, 347–352. [Google Scholar] [CrossRef]
- Steullet, P.; Cabungcal, J.H.; Monin, A.; Dwir, D.; O′Donnell, P.; Cuenod, M.; Do, K.Q. Redox dysregulation, neuroinflammation, and NMDA receptor hypofunction: A “central hub” in schizophrenia pathophysiology? Schizophr. Res. 2016, 176, 41–51. [Google Scholar] [CrossRef]
- Steullet, P.; Cabungcal, J.H.; Coyle, J.; Didriksen, M.; Gill, K.; Grace, A.A.; Hensch, T.K.; LaMantia, A.S.; Lindemann, L.; Maynard, T.M.; et al. Oxidative stress-driven parvalbumin interneuron impairment as a common mechanism in models of schizophrenia. Mol. Psychiatry 2017, 22, 936–943. [Google Scholar] [CrossRef]
- Cuenod, M.; Steullet, P.; Cabungcal, J.H.; Dwir, D.; Khadimallah, I.; Klauser, P.; Conus, P.; Do, K.Q. Caught in vicious circles: A perspective on dynamic feed-forward loops driving oxidative stress in schizophrenia. Mol. Psychiatry 2022, 27, 1886–1897. [Google Scholar] [CrossRef]
- Hardingham, G.E.; Do, K.Q. Linking early-life NMDAR hypofunction and oxidative stress in schizophrenia pathogenesis. Nat. Rev. Neurosci. 2016, 17, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Marchal-Victorion, S.; Bronzetti, E.; Parini, A.; Amenta, F.; Tayebati, S.K. Dopamine D4 receptor expression in rat kidney: Evidence for pre- and postjunctional localization. J. Histochem. Cytochem. 2002, 50, 1091–1096. [Google Scholar] [CrossRef] [PubMed]
- Amenta, F.; Ricci, A.; Tayebati, S.K.; Zaccheo, D. The peripheral dopaminergic system: Morphological analysis, functional and clinical applications. Ital. J. Anat. Embryol. 2002, 107, 145–167. [Google Scholar] [PubMed]
- Ricci, A.; Bronzetti, E.; Fedele, F.; Ferrante, F.; Zaccheo, D.; Amenta, F. Pharmacological characterization and autoradiographic localization of a putative dopamine D4 receptor in the heart. J. Auton. Pharmacol. 1998, 18, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Wedzony, K.; Chocyk, A.; Mackowiak, M.; Fijal, K.; Czyrak, A. Cortical localization of dopamine D4 receptors in the rat brain--immunocytochemical study. J. Physiol. Pharmacol. 2000, 51, 205–221. [Google Scholar]
- Defagot, M.C.; Malchiodi, E.L.; Villar, M.J.; Antonelli, M.C. Distribution of D4 dopamine receptor in rat brain with sequence-specific antibodies. Brain Res. Mol. Brain Res. 1997, 45, 1–12. [Google Scholar] [CrossRef]
- Mrzljak, L.; Bergson, C.; Pappy, M.; Huff, R.; Levenson, R.; Goldman-Rakic, P.S. Localization of dopamine D4 receptors in GABAergic neurons of the primate brain. Nature 1996, 381, 245–248. [Google Scholar] [CrossRef]
- Mofleh, R.; Kocsis, B. Respiratory coupling between prefrontal cortex and hippocampus of rats anaesthetized with urethane in theta and non-theta states. Eur. J. Neurosci. 2021, 54, 5507–5517. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.R.G.; Ward-Flanagan, R.; Dickson, C.T. Long-term stability of physiological signals within fluctuations of brain state under urethane anesthesia. PLoS ONE 2021, 16, e0258939. [Google Scholar] [CrossRef] [PubMed]
- Buzsaki, G.; Leung, L.W.; Vanderwolf, C.H. Cellular bases of hippocampal EEG in the behaving rat. Brain Res. 1983, 287, 139–171. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuang, J.; Kafetzopoulos, V.; Deth, R.; Kocsis, B. Dopamine D4 Receptor Agonist Drastically Increases Delta Activity in the Thalamic Nucleus Reuniens: Potential Role in Communication between Prefrontal Cortex and Hippocampus. Int. J. Mol. Sci. 2023, 24, 15289. https://doi.org/10.3390/ijms242015289
Kuang J, Kafetzopoulos V, Deth R, Kocsis B. Dopamine D4 Receptor Agonist Drastically Increases Delta Activity in the Thalamic Nucleus Reuniens: Potential Role in Communication between Prefrontal Cortex and Hippocampus. International Journal of Molecular Sciences. 2023; 24(20):15289. https://doi.org/10.3390/ijms242015289
Chicago/Turabian StyleKuang, J., V. Kafetzopoulos, Richard Deth, and B. Kocsis. 2023. "Dopamine D4 Receptor Agonist Drastically Increases Delta Activity in the Thalamic Nucleus Reuniens: Potential Role in Communication between Prefrontal Cortex and Hippocampus" International Journal of Molecular Sciences 24, no. 20: 15289. https://doi.org/10.3390/ijms242015289
APA StyleKuang, J., Kafetzopoulos, V., Deth, R., & Kocsis, B. (2023). Dopamine D4 Receptor Agonist Drastically Increases Delta Activity in the Thalamic Nucleus Reuniens: Potential Role in Communication between Prefrontal Cortex and Hippocampus. International Journal of Molecular Sciences, 24(20), 15289. https://doi.org/10.3390/ijms242015289