
BMP-9 Improves the Osteogenic Differentiation Ability over BMP-2 through p53 Signaling In Vitro in Human Periosteum-Derived Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
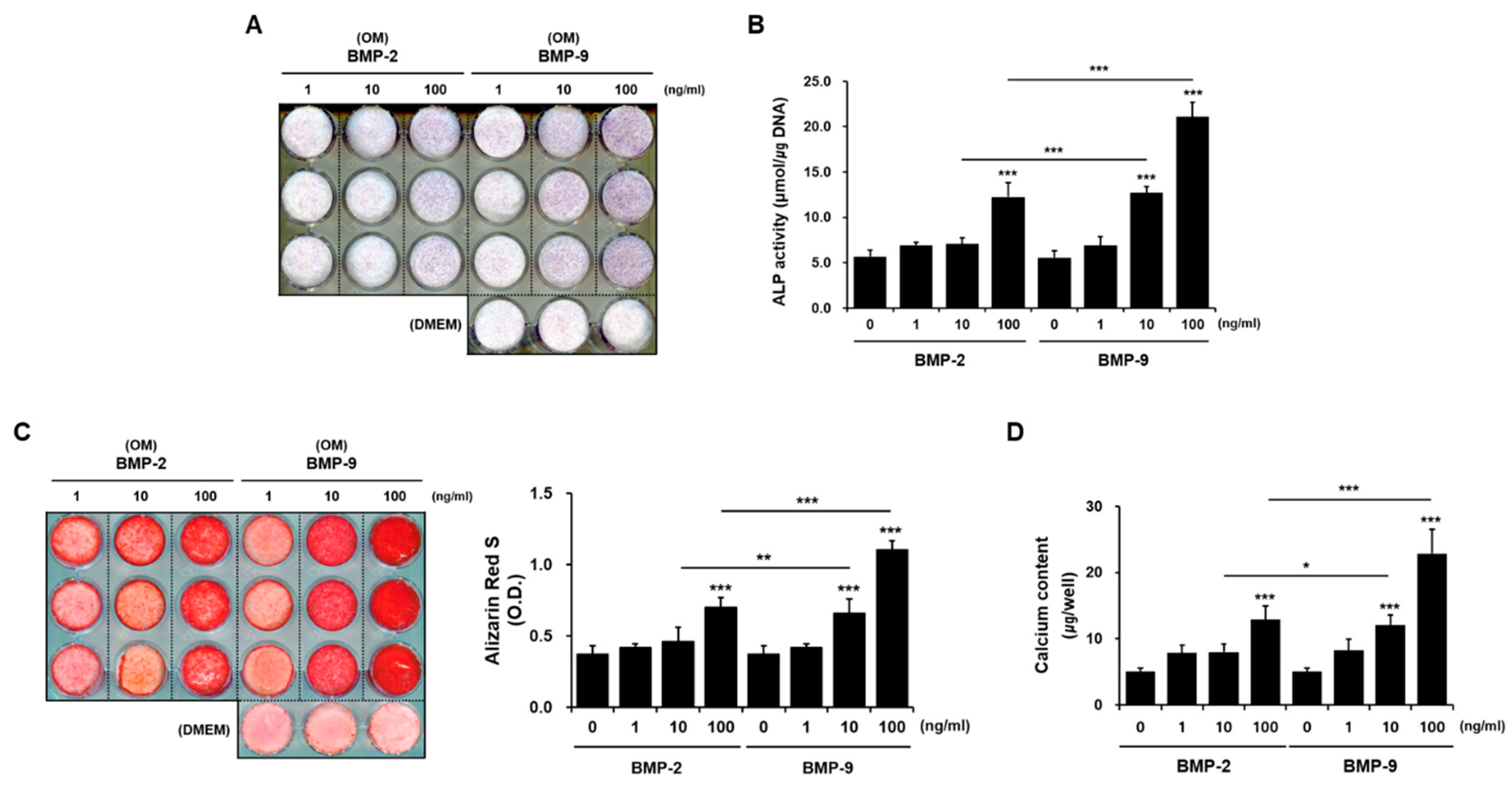
2.1. Osteogenic Potential of hPDCs Induced by BMP-9 In Vitro
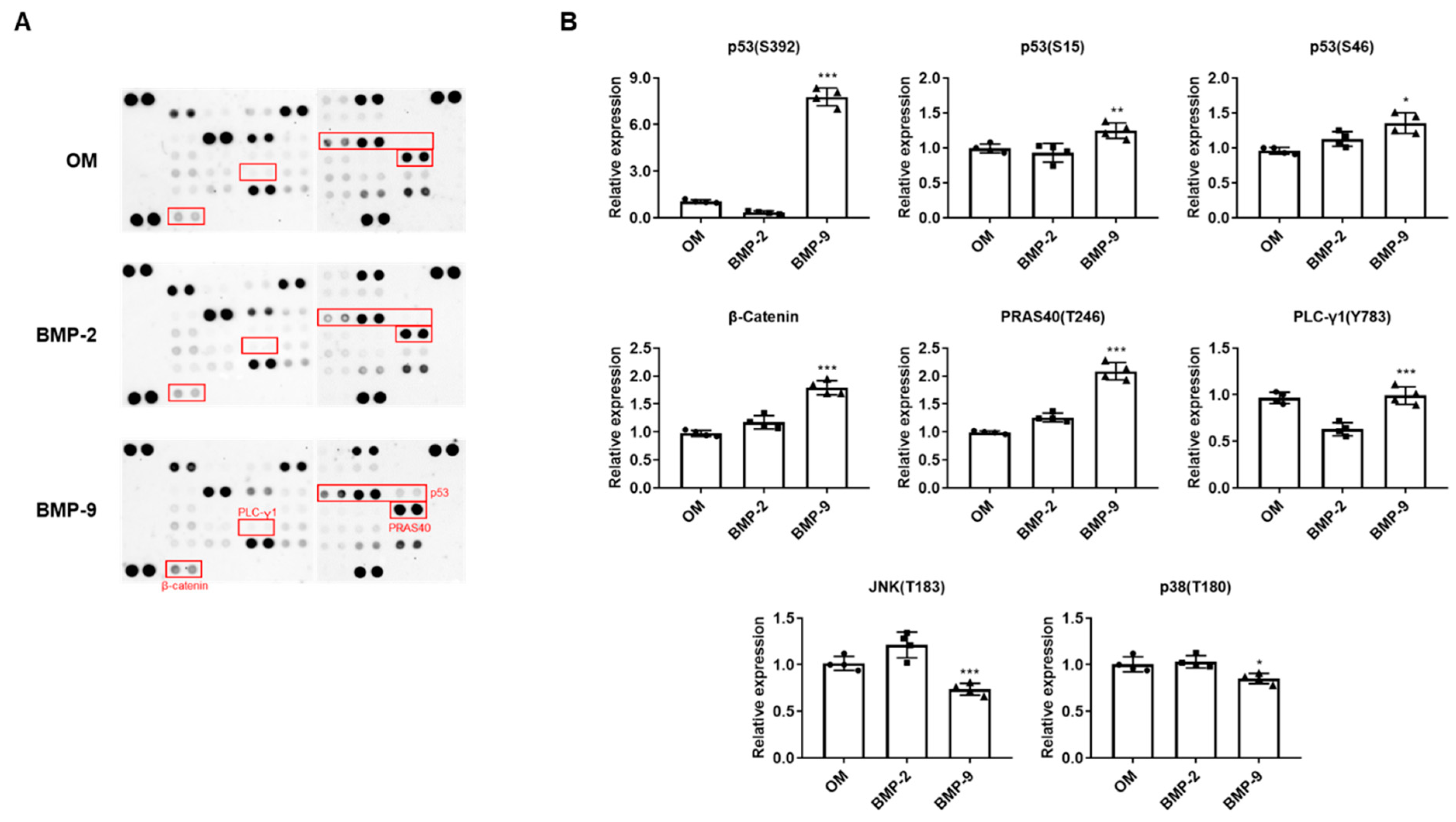
2.2. Comparison of Intracellular Signaling Pathway Differences in BMP-Induced hPDCs
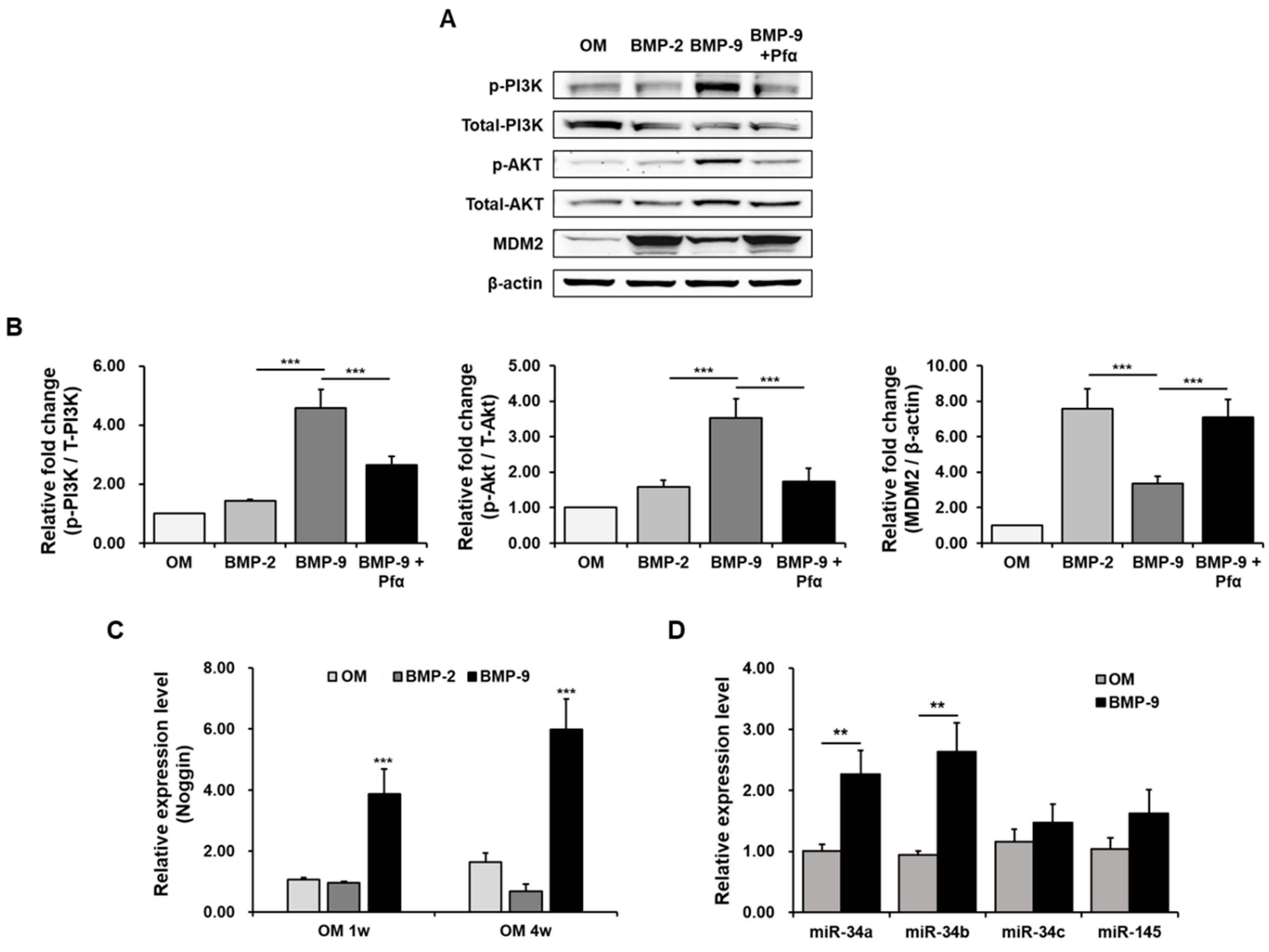
2.3. Reduction in Osteogenic Differentiation by Pifithrin-α
2.4. Mechanism of Action of BMP-9 and p53
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Culture and Osteogenic Differentiation of the hPDCs
4.3. Evaluation of Osteogenic Phenotype
4.4. Phosphorylation Array
4.5. Western Blot Analysis
4.6. Reverse Transcription and Quantitative RT-PCR (qRT-PCR) Using TaqMan Assays
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bharadwaz, A.; Jayasuri, A.C.y. Osteogenic differentiation cues of the bone morphogenetic protein-9 (BMP-9) and its recent advances in bone tissue regeneration. Mater. Sci. Eng. C 2021, 120, 111748. [Google Scholar] [CrossRef]
- Luo, J.; Sun, M.H.; Kang, Q.; Peng, Y.; Jiang, W.; Luu, H.H.; Luo, Q.; Park, J.Y.; Li, Y.; Haydon, R.C.; et al. Gene therapy for bone regeneration. Curr. Gene Ther. 2005, 5, 167–179. [Google Scholar] [CrossRef]
- Yang, J.; Shi, P.; Tu, M.; Wang, Y.; Liu, M.; Fan, F.; Du, M. Bone morphogenetic proteins: Relationship between molecular structure and their osteogenic activity. Food Sci. Hum. Wellness 2014, 3, 127–135. [Google Scholar] [CrossRef]
- Bleuming, S.A.; He, X.C.; Kodach, L.L.; Hardwick, J.C.; Koopman, F.A.; Ten Kate, F.J.; van Deventer, S.J.; Hommes, D.W.; Peppelenbosch, M.P.; Offerhaus, G.J.; et al. Bone morphogenetic protein signaling suppresses tumorigenesis at gastric epithelial transition zones in mice. Cancer Res. 2007, 67, 8149–8155. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.G.; del Castillo, L.M.; Ochoa-Bolton, E.; Yamauchi, K.; Smogorzewski, J.; Butler, S.J. BMPs direct sensory interneuron identity in the developing spinal cord using signal- specific not morphogenic activities. eLife 2017, 6, e30647. [Google Scholar] [CrossRef] [PubMed]
- Urist, M.R.; Strates Basil, S. Bone Morphogenetic Protein. J. Dent. Res. 1971, 50, 1392–1406. [Google Scholar] [CrossRef]
- Reddi, A.H.; Huggins, C. Biochemical sequences in the transformation of normal fibroblasts in adolescent rats. Proc. Natl. Acad. Sci. USA 1972, 69, 1601–1605. [Google Scholar] [CrossRef] [PubMed]
- Sampath, T.K.; Reddi, A.H. Dissociative extraction and reconstitution of extracellular matrix components involved in local bone differentiation. Proc. Natl. Acad. Sci. USA 1981, 78, 7599–7603. [Google Scholar] [CrossRef] [PubMed]
- Sampath, T.K.; Muthukumaran, N.; Reddi, A.H. Isolation of osteogenin, an extracellular matrix-associated, bone-inductive protein, by heparin affinity chromatography. Proc. Natl. Acad. Sci. USA 1987, 84, 7109–7113. [Google Scholar] [CrossRef]
- Ong, K.L.; Villarraga, M.L.; Lau, E.; Carreon, L.Y.; Kurtz, S.M.; Glassman, S.D. Off-label use of bone morphogenetic proteins in the United States using administrative data. Spine 2010, 35, 1794–1800. [Google Scholar] [CrossRef]
- Even, J.; Eskander, M.; Kang, J. Bone morphogenetic protein in spine surgery: Current and future uses. J. Am. Acad. Orthop. Surg. 2012, 20, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.A.; Zhao, Q.; Baker, K.A.; Naik, C.; Chen, C.; Pukac, L.; Singh, M.; Tsareva, T.; Parice, Y.; Mahoney, A.; et al. Crystal structure of BMP-9 and functional interactions with pro-region and receptors. J. Biol. Chem. 2005, 280, 25111–25118. [Google Scholar] [CrossRef] [PubMed]
- Luther, G.; Wagner, E.R.; Zhu, G.; Kang, Q.; Luo, Q.; Lamplot, J.; Bi, Y.; Luo, X.; Luo, J.; Teven, C.; et al. BMP-9 Induced Osteogenic Differentiation of Mesenchymal Stem Cells: Molecular Mechanism and Therapeutic Potential. Curr. Gene Ther. 2011, 11, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Castonguay, R.; Werner, E.D.; Matthews, R.G.; Presman, E.; Mulivor, A.W.; Solban, N.; Sako, D.; Pearsall, R.S.; Underwood, K.W.; Seehra, J.; et al. Soluble endoglin specifically binds bone morphogenetic proteins 9 and 10 via its orphan domain, inhibits blood vessel formation, and suppresses tumor growth. J. Biol. Chem. 2011, 286, 30034–30046. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.Y.; Golzio, C.; Gatza, C.E.; Sharma, A.; Katsanis, N.; Blobe, G.C. Endoglin regulates PI3-kinase/Akt trafficking and signaling to alter endothelial capillary stability during angiogenesis. Mol. Biol. Cell 2012, 23, 2412–2423. [Google Scholar] [CrossRef]
- Saito, T.; Bokhove, M.; Croci, R.; Zamora-Caballero, S.; Han, L.; Letarte, M.; de Sanctis, D.; Jovine, L. Structural basis of the human endoglin-BMP9 interaction: Insights into BMP signaling and HHT1. Cell Rep. 2017, 19, 1917–1928. [Google Scholar] [CrossRef]
- Bergeron, E.; Senta, H.; Mailloux, A.; Park, H.; Lord, E.; Faucheux, N. Murine preosteoblast differentiation induced by a peptide derived from bone morphogenetic proteins-9. Tissue Eng. Part A 2009, 15, 3341–3349. [Google Scholar] [CrossRef]
- Kang, Q.; Sun, M.H.; Cheng, H.; Peng, Y.; Montag, A.G.; Deyrup, A.T.; Jiang, W.; Luu, H.H.; Luo, J.; Szatkowski, J.P.; et al. Characterization of the distinct orthotopic bone-forming activity of 14 BMPs using recombinant adenovirus-mediated gene delivery. Gene Ther. 2004, 11, 1312–1320. [Google Scholar] [CrossRef]
- Eiraku, N.; Chiba, N.; Nakamura, T.; Amir, M.S.; Seong, C.-H.; Ohnishi, T.; Kusuyama, J.; Noguchi, K.; Matsuguchi, T. BMP9 directly induces rapid GSK3-b phosphorylation in a Wnt-independent manner through class I PI3K-Akt axis in osteoblasts. FASEB J. 2019, 33, 12124–12134. [Google Scholar] [CrossRef]
- Artigas, N.; Gámez, B.; Cubillos-Rojas, M.; Diego, C.S.-D.; Valer, J.A.; Pons, G.; Rosa, J.L.; Ventura, F. p53 inhibits SP7/Osterix activity in the transcriptional program of osteoblast differentiation. Cell Death Differ. 2017, 24, 2022–2031. [Google Scholar] [CrossRef]
- Shah, S.; Pendleton, E.; Couture, O.; Broachwalla, M.; Kusper, T.; Alt, L.A.C.; Fay, M.J.; Chandar, N. P53 regulation of osteoblast differentiation is mediated through specific microRNAs. Biochem. Biophys. Rep. 2021, 25, 100920. [Google Scholar] [CrossRef] [PubMed]
- Lengner, C.J.; Steinman, H.A.; Gagnon, J.; Smith, T.W.; Henderson, J.E.; Kream, B.E.; Stein, G.S.; Lian, J.B.; Jones, S.N. Osteoblast differentiation and skeletal development are regulated by Mdm2–p53 signaling. J. Cell Biol. 2006, 172, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Cell Death in Chondrocytes, Osteoblasts, and Osteocytes. Int. J. Mol. Sci. 2016, 17, 2045. [Google Scholar] [CrossRef]
- Cheng, H.; Jiang, W.; Phillips, F.M.; Haydon, R.C.; Peng, Y.; Zhou, L.; Luu, H.H.; An, N.; Breyer, B.; Vanichakarn, P.; et al. Osteogenic activity of the fourteen types of human bone morphogenetic proteins (BMPs). J. Bone Joint Surg. Am. 2003, 85, 1544–1552. [Google Scholar] [CrossRef]
- Kang, Q.; Song, W.X.; Luo, Q.; Tang, N.; Luo, J.; Luo, X.; Chen, J.; Bi, Y.; He, B.C.; Park, J.K.; et al. A comprehensive analysis of the dual roles of BMPs in regulating adipogenic and osteogenic differentiation of mesenchymal progenitor cells. Stem Cells Dev. 2009, 18, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Park, J.-H.; Kim, M.J.; Lee, J.H.; Oh, S.H.; Byun, J.-H. The effects of VEGF-centered biomimetic delivery of growth factors on bone regeneration. Biomater. Sci. 2021, 9, 3675–3691. [Google Scholar] [CrossRef]
- Chen, G.; Deng, C.; Li, Y.-P. TGF-β and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef]
- Vousden, K.H.; Lu, X. Live or let die: The cell’s response to p53. Nat. Rev. Cancer 2002, 2, 594–604. [Google Scholar] [CrossRef]
- Oren, M. Decision making by p53: Life. death and cancer. Cell Death Differ. 2003, 10, 431–442. [Google Scholar] [CrossRef]
- McGonnell, I.M.; Grigoriadis, A.E.; Lam, E.W.-F.; Price, J.S.; Sunters, A. A specific role for phosphoinositide 3-kinase and AKT in osteoblasts. Front. Endocrinol. 2012, 20, 88. [Google Scholar] [CrossRef]
- Zhou, J.; Li, Z.; Huang, Y.; Ju, W.; Wang, D.; Zhu, X.; He, X. MicroRNA-26a targets the mdm2/p53 loop directly in response to liver regeneration. Int. J. Mol. Med. 2019, 44, 1505–1514. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yan, R.; Li, B.; Liu, J.; Liu, X.; Song, W.; Zhu, C. Silencing CCNG1 protects MPC-5 cells from high glucose-induced proliferation-inhibition and apoptosis-promotion via MDM2/p53 signaling pathway. Int. Urol. Nephrol. 2020, 52, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Guo, H.; Kuang, P.; Cui, H.; Deng, H.; Liu, H.; Lu, Y.; Wei, Q.; Chen, L.; Fang, J.; et al. Sodium Fluoride Arrests Renal G2/M Phase Cell-Cycle Progression by Activating ATM-Chk2-P53/Cdc25C Signaling Pathway in Mice. Cell Physiol. Biochem. 2018, 51, 2421–2433. [Google Scholar] [CrossRef]
- Mei, X.-L.; Zhong, S. Long noncoding RNA LINC00520 prevents the progression of cutaneous squamous cell carcinoma through the inactivation of the PI3K/Akt signaling pathway by downregulating EGFR. Chin. Med. J. 2019, 132, 454–465. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, Y.; Yu, Q. The PI3K/Akt signaling pathway exerts effects on the implantation of mouse embryos by regulating the expression of RhoA. Int. J. Mol. Med. 2014, 33, 1089–1096. [Google Scholar] [CrossRef]
- Meng, X.; Xia, C.; Yeb, Q.; Nie, X. tert-Butyl-p-benzoquinone induces autophagy by inhibiting the Akt/mTOR signaling pathway in RAW 264.7 cells. Food Funct. 2020, 11, 4193–4201. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Lieberman, J.; Lal, A. Desperately seeking microRNA targets. Nat. Struct. Mol. Biol. 2010, 17, 1169–1174. [Google Scholar] [CrossRef]
- Sera, S.R.; Zur Nieden, N.I. microRNA Regulation of skeletal development. Curr. Osteoporos. Rep. 2017, 15, 353–366. [Google Scholar] [CrossRef]
- Corney, D.C.; Flesken-Nikitin, A.; Godwin, A.K.; Wang, W.; Nikitin, A.Y. MicroRNA-34b and MicroRNA-34c Are Targets of p53 and Cooperate in Control of Cell Proliferation and Adhesion-Independent Growth. Cancer Res. 2007, 18, 8433–8438. [Google Scholar] [CrossRef]
- Hensley, A.P.; McAlinden, A. The role of microRNAs in bone development. Bone 2020, 143, 115760. [Google Scholar] [CrossRef]
- Suh, S.O.; Chen, Y.; Zaman, M.S.; Hirata, H.; Yamamura, S.; Shahryari, V.; Liu, J.; Tabatabai, Z.L.; Kakar, S.; Deng, G.; et al. MicroRNA-145 is regulated by DNA methylation and p53 gene mutation in prostate cancer. Carcinogenesis 2011, 5, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, G.; Lee, J.; Lee, Y.; Kim, J.-H. Secretome of stem cells: Roles of extracellular vesicles in diseases, stemness, differentiation, and reprogramming. Tissue Eng. Regen. Med. 2021, 19, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-H.; Koh, E.-B.; Seo, Y.-J.; Oh, H.-S.; Won, J.-Y.; Hwang, S.-C.; Byun, J.-H. Tiron Has Negative Effects on Osteogenic Differentiation via Mitochondrial Dysfunction in Human Periosteum-Derived Cells. Int. J. Mol. Sci. 2022, 23, 14040. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-H.; Park, B.-W.; Kang, Y.-H.; Byun, S.-H.; Hwang, S.-C.; Kim, D.R.; Woo, D.K.; Byun, J.-H. Lin28a enhances in vitro osteoblastic differentiation of human periosteum-derived cells. Cell Biochem. Funct. 2017, 35, 497–509. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-H.; Koh, E.-B.; Seo, Y.-J.; Oh, H.-S.; Byun, J.-H. BMP-9 Improves the Osteogenic Differentiation Ability over BMP-2 through p53 Signaling In Vitro in Human Periosteum-Derived Cells. Int. J. Mol. Sci. 2023, 24, 15252. https://doi.org/10.3390/ijms242015252
Park J-H, Koh E-B, Seo Y-J, Oh H-S, Byun J-H. BMP-9 Improves the Osteogenic Differentiation Ability over BMP-2 through p53 Signaling In Vitro in Human Periosteum-Derived Cells. International Journal of Molecular Sciences. 2023; 24(20):15252. https://doi.org/10.3390/ijms242015252
Chicago/Turabian StylePark, Jin-Ho, Eun-Byeol Koh, Young-Jin Seo, Hye-Seong Oh, and June-Ho Byun. 2023. "BMP-9 Improves the Osteogenic Differentiation Ability over BMP-2 through p53 Signaling In Vitro in Human Periosteum-Derived Cells" International Journal of Molecular Sciences 24, no. 20: 15252. https://doi.org/10.3390/ijms242015252
APA StylePark, J.-H., Koh, E.-B., Seo, Y.-J., Oh, H.-S., & Byun, J.-H. (2023). BMP-9 Improves the Osteogenic Differentiation Ability over BMP-2 through p53 Signaling In Vitro in Human Periosteum-Derived Cells. International Journal of Molecular Sciences, 24(20), 15252. https://doi.org/10.3390/ijms242015252