

Nicotine Impairs the Anti-Contractile Function of Perivascular Adipose Tissue by Inhibiting the PPARγ–Adiponectin–AdipoR1 Axis
 , and
, and
Abstract
1. Introduction
2. Results
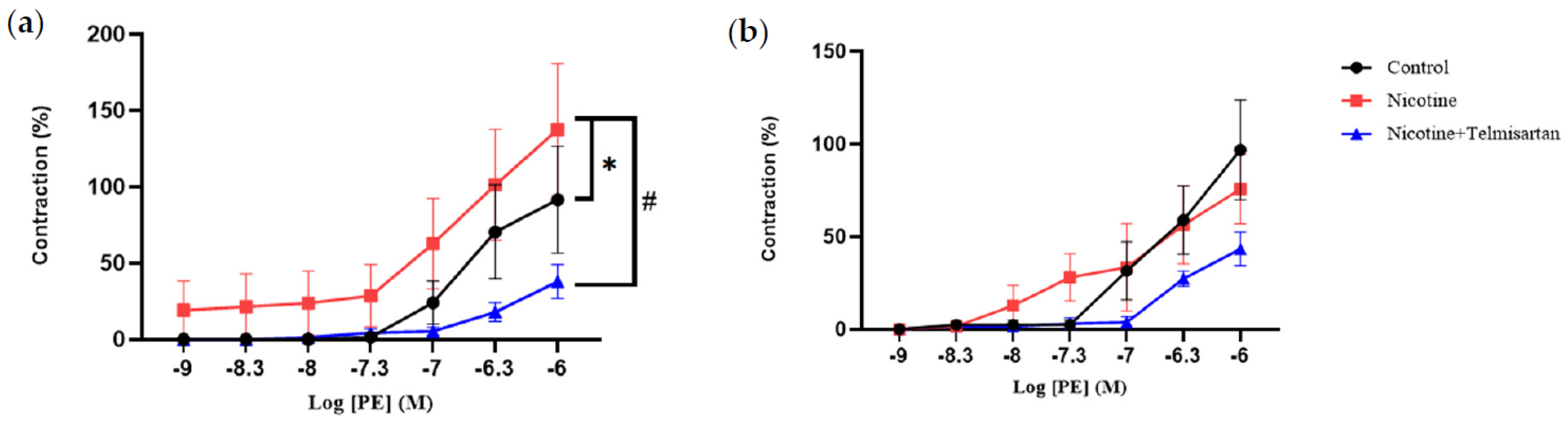
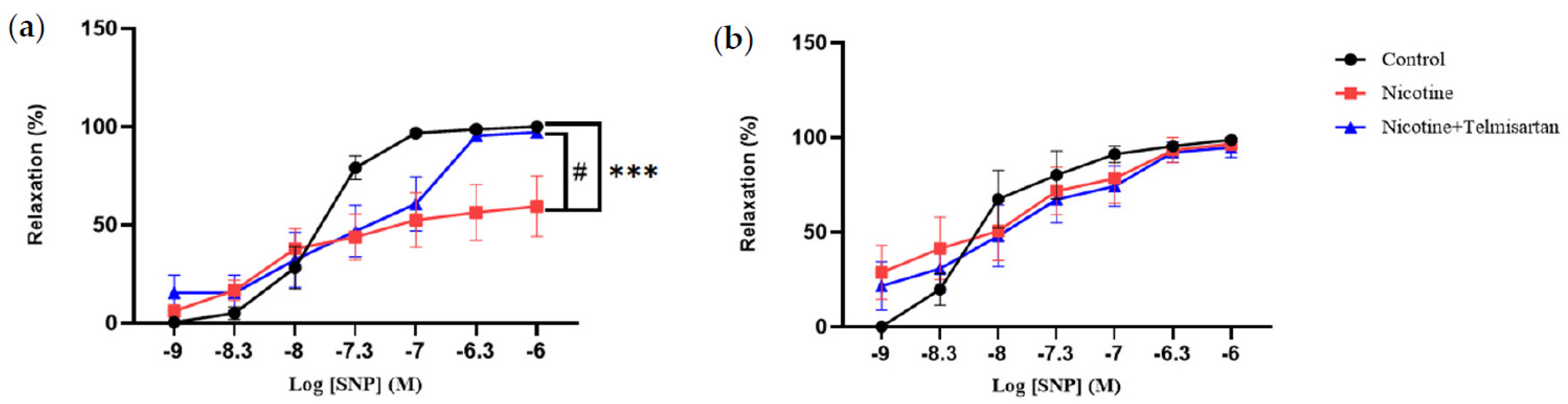
2.1. Effect of Nicotine on the Vasoreactive Function of PVAT in the Rat Aorta
2.1.1. Aortic Contraction to Phenylephrine (PE)
2.1.2. Aortic Relaxation to Sodium Nitroprusside (SNP)
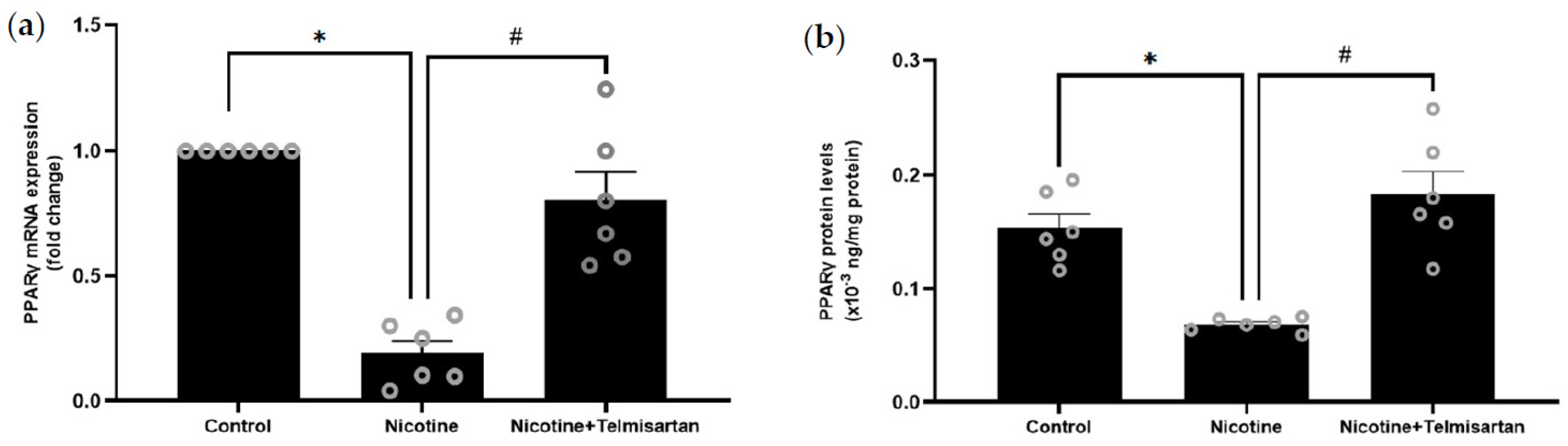
2.2. Effect of Nicotine on Gene and Protein Expression of PPARγ in PVAT
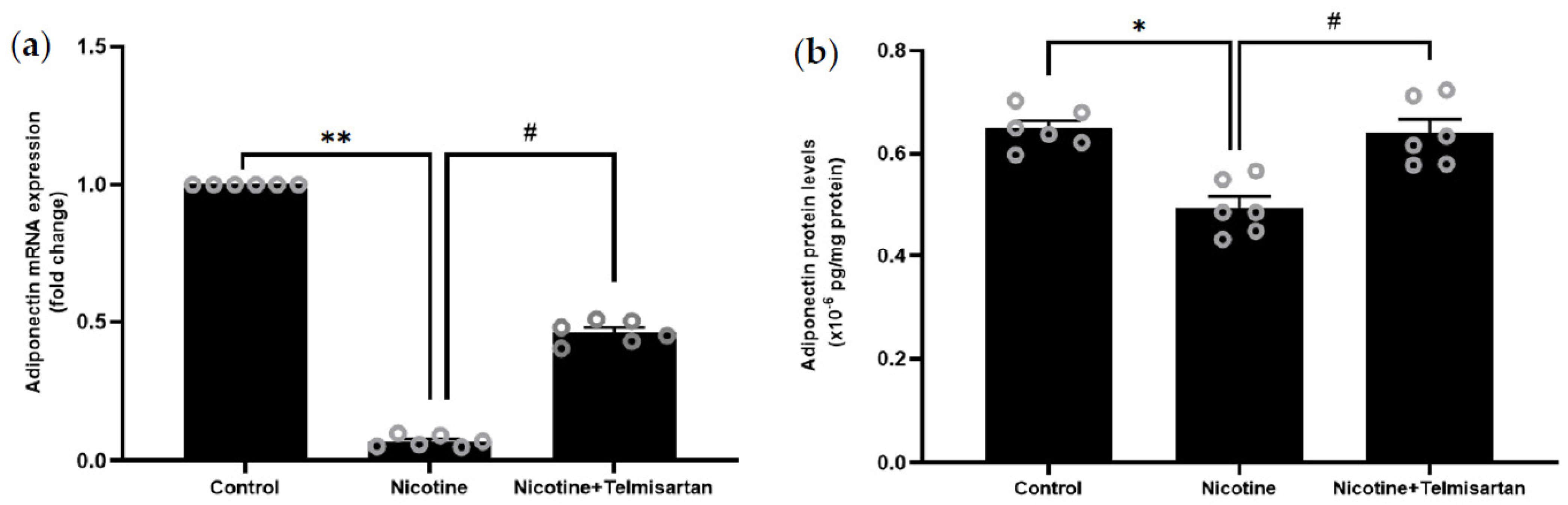
2.3. Effect of Nicotine on Gene and Protein Expression of Adiponectin in PVAT
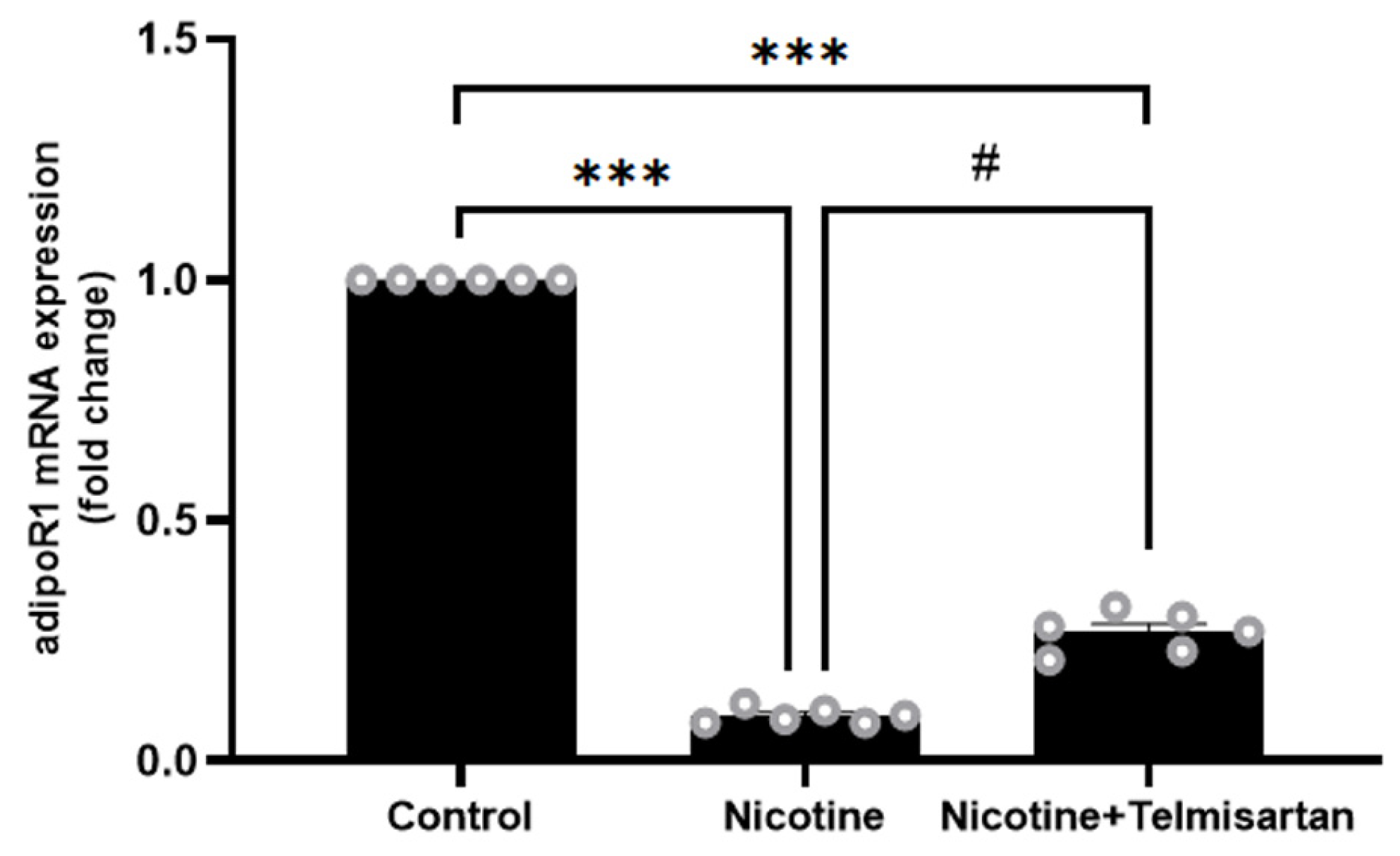
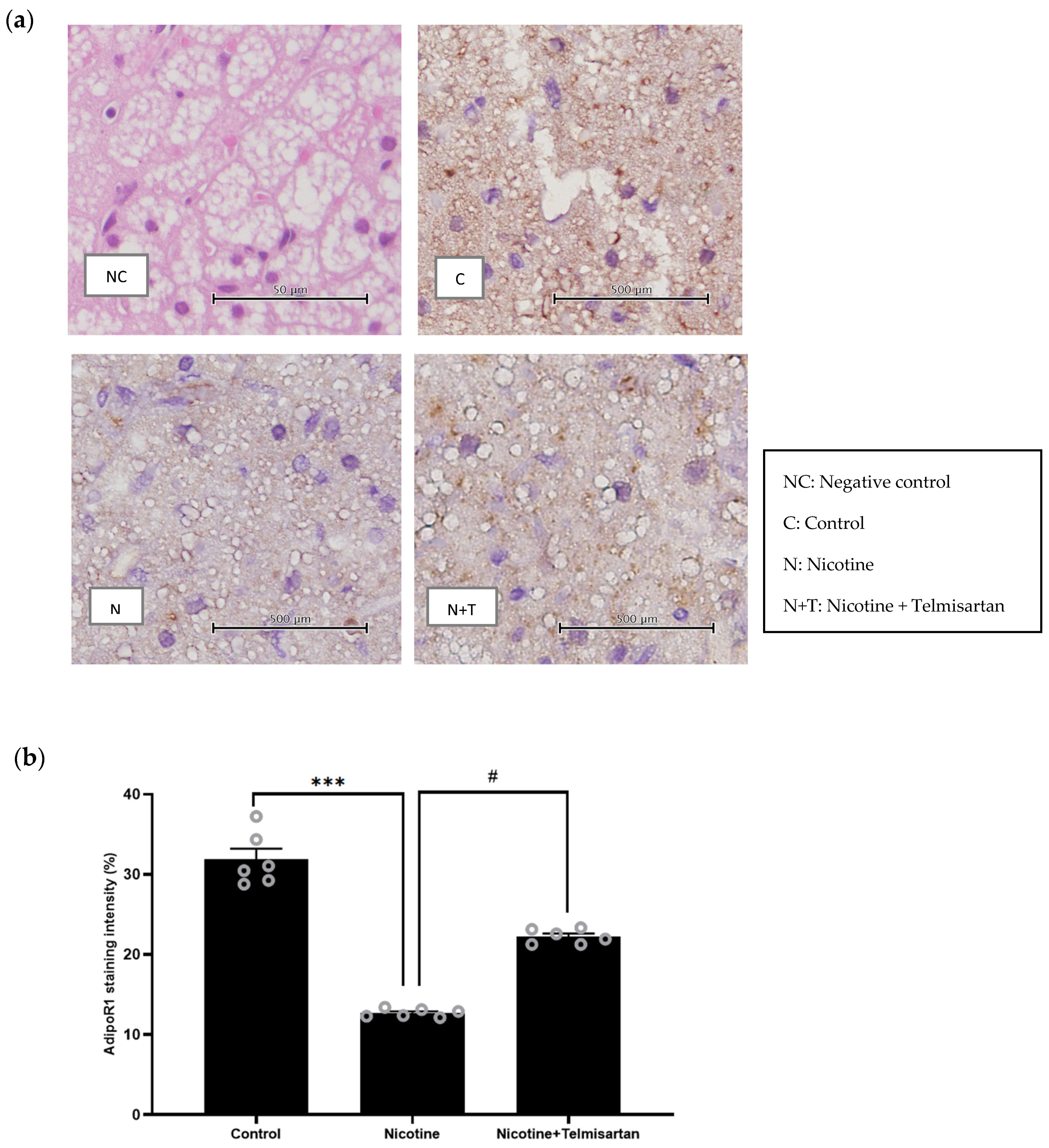
2.4. Effect of Nicotine on Gene Expression and Protein Distribution of AdipoR1 in PVAT
3. Discussion
4. Materials and Methods
4.1. Animal and Study Design
4.2. Wire Myography
4.3. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.4. Protein Extraction and Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Immunohistochemistry (IHC) Analysis
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Cardiovascular Diseases (CVDs); World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2020 Update: A Report From the American Heart Association. Circulation 2020, 141, e139–e596. [Google Scholar] [CrossRef] [PubMed]
- Amiri, P.; Mohammadzadeh-Naziri, K.; Abbasi, B.; Cheraghi, L.; Jalali-Farahani, S.; Momenan, A.A.; Amouzegar, A.; Hadaegh, F.; Azizi, F. Smoking Habits and Incidence of Cardiovascular Diseases in Men and Women: Findings of a 12 Year Follow up among an Urban Eastern-Mediterranean Population. BMC Public Health 2019, 19, 1042. [Google Scholar] [CrossRef] [PubMed]
- NIDA. Cigarettes and Other Tobacco Products DrugFacts; National Institute on Drug Abuse: North Bethesda, MA, USA, 2021. [Google Scholar]
- Valentine, G.; Sofuoglu, M. Cognitive Effects of Nicotine: Recent Progress. Curr. Neuropharmacol. 2017, 15, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Salleh, M.F.R.R.M.; Aminuddin, A.; Hamid, A.A.; Salamt, N.; Japar Sidik, F.Z.; Ugusman, A. Piper Sarmentosum Roxb. Attenuates Vascular Endothelial Dysfunction in Nicotine-Induced Rats. Front. Pharmacol. 2021, 12, 667102. [CrossRef] [PubMed]
- Zhang, P.; He, Y.; Wu, S.; Li, X.; Lin, X.; Gan, M.; Chen, L.; Zhao, Y.; Niu, L.; Zhang, S.; et al. Factors Associated with White Fat Browning: New Regulators of Lipid Metabolism. Int. J. Mol. Sci. 2022, 23, 7641. [Google Scholar] [CrossRef] [PubMed]
- Saely, C.H.; Geiger, K.; Drexel, H. Brown versus White Adipose Tissue: A Mini-Review. Gerontology 2012, 58, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Soltis, E.E.; Cassis, L.A. Influence of Perivascular Adipose Tissue on Rat Aortic Smooth Muscle Responsiveness. Clin. Exp. Hypertens. A 1991, 13, 277–296. [Google Scholar] [CrossRef]
- Grigoras, A.; Amalinei, C.; Balan, R.A.; Giusca, S.E.; Caruntu, I.D. Perivascular Adipose Tissue in Cardiovascular Diseases-an Update. Anatol. J. Cardiol. 2019, 22, 219–231. [Google Scholar] [CrossRef]
- Kim, H.W.; Shi, H.; Winkler, M.A.; Lee, R.; Weintraub, N.L. Perivascular Adipose Tissue and Vascular Perturbation/Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2569–2576. [Google Scholar] [CrossRef]
- Rami, A.Z.A.; Hamid, A.A.; Anuar, N.N.M.; Aminuddin, A.; Ugusman, A. Exploring the Relationship of Perivascular Adipose Tissue Inflammation and the Development of Vascular Pathologies. Mediat. Inflamm. 2022, 2022, 2734321. [Google Scholar] [CrossRef]
- Almabrouk, T.A.M.; Ugusman, A.B.; Katwan, O.J.; Salt, I.P.; Kennedy, S. Deletion of AMPKα1 Attenuates the Anticontractile Effect of Perivascular Adipose Tissue (PVAT) and Reduces Adiponectin Release. Br. J. Pharmacol. 2017, 174, 3398–3410. [Google Scholar] [CrossRef]
- Kagota, S.; Maruyama-Fumoto, K.; Iwata, S.; Shimari, M.; Koyanagi, S.; Shiokawa, Y.; McGuire, J.J.; Shinozuka, K. Perivascular Adipose Tissue-Enhanced Vasodilation in Metabolic Syndrome Rats by Apelin and N-Acetyl−l-Cysteine-Sensitive Factor(S). Int. J. Mol. Sci. 2018, 20, 106. [Google Scholar] [CrossRef]
- Rasmussen, M.S.; Lihn, A.S.; Pedersen, S.B.; Bruun, J.M.; Rasmussen, M.; Richelsen, B. Adiponectin Receptors in Human Adipose Tissue: Effects of Obesity, Weight Loss, and Fat Depots. Obesity 2006, 14, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, Y.; Fan, Y.; Tang, Z.; Wang, N. Overexpression of Adiponectin Receptors Potentiates the Antiinflammatory Action of Subeffective Dose of Globular Adiponectin in Vascular Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Man, A.W.C.; Zhou, Y.; Xia, N.; Li, H. Perivascular Adipose Tissue as a Target for Antioxidant Therapy for Cardiovascular Complications. Antioxidants 2020, 9, 574. [Google Scholar] [CrossRef] [PubMed]
- Komiyama, M.; Wada, H.; Yamakage, H.; Satoh-Asahara, N.; Sunagawa, Y.; Morimoto, T.; Ozaki, Y.; Shimatsu, A.; Takahashi, Y.; Hasegawa, K. Analysis of Changes on Adiponectin Levels and Abdominal Obesity after Smoking Cessation. PLoS ONE 2018, 13, e0201244. [Google Scholar] [CrossRef] [PubMed]
- Toffolo, M.C.F.; Gomes, A.D.S.; Van Keulen, H.V.; Louro, M.B.; Silva, F.M.C.E.; Luquetti, S.C.P.D.; de Aguiar, A.S.; Ferreira, A.P. Alteration of Inflammatory Adipokines after Four Months of Smoking Abstinence in Multidisciplinary Intervention Program. Nutr. Hosp. 2018, 35, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.N.; Yang, G.H.; Wang, Z.Q.; Liu, C.W.; Li, T.J.; Lai, Z.C.; Miao, S.Y.; Wang, L.F.; Liu, B. Role of Perivascular Adipose Tissue in Nicotine-Induced Endothelial Cell Inflammatory Responses. Mol. Med. Rep. 2016, 14, 5713–5718. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gao, Y.J.; Holloway, A.C.; Zeng, Z.H.; Lim, G.E.; Petrik, J.J.; Foster, W.G.; Lee, R.M.K.W. Prenatal Exposure to Nicotine Causes Postnatal Obesity and Altered Perivascular Adipose Tissue Function. Obes. Res. 2005, 13, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.J.; Holloway, A.C.; Su, L.Y.; Takemori, K.; Lu, C.; Lee, R.M.K.W. Effects of Fetal and Neonatal Exposure to Nicotine on Blood Pressure and Perivascular Adipose Tissue Function in Adult Life. Eur. J. Pharmacol. 2008, 590, 264–268. [Google Scholar] [CrossRef]
- Zhu, D.; Xu, L.; Wei, X.; Xia, B.; Gong, Y.; Li, Q.; Chen, X. PPARγ Enhanced Adiponectin Polymerization and Trafficking by Promoting RUVBL2 Expression during Adipogenic Differentiation. Gene 2021, 764, 145100. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Zhao, X.; Villacorta, L.; Rom, O.; Garcia-barrio, M.T.; Guo, Y.; Fan, Y.; Zhu, T.; Zhang, J.; Zeng, R.; et al. Brown Adipocyte Specific PPARγ Deletion Impairs PVAT Development and Enhances Atherosclerosis in Mice. Arter. Thromb. Vasc. Biol. 2018, 38, 1738–1747. [Google Scholar] [CrossRef]
- Akhrass, P.R.; McFarlane, S.I. Telmisartan and Cardioprotection. Vasc. Health Risk Manag. 2011, 7, 677–683. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Araki, K.; Masaki, T.; Katsuragi, I.; Tanaka, K.; Kakuma, T.; Yoshimatsu, H. Telmisartan Prevents Obesity and Increases the Expression of Uncoupling Protein 1 in Diet-Induced Obese Mice. Hypertension 2006, 48, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, Y.; Liu, Q.; Zhang, J.; Zhang, Z.; Wu, T.; Tang, Q.; Huang, C.; Li, R.; Zhou, J.; et al. Telmisartan Attenuates Obesity-Induced Insulin Resistance via Suppression of AMPK Mediated ER Stress. Biochem. Biophys. Res. Commun. 2020, 523, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Ayza, M.A.; Zewdie, K.A.; Tesfaye, B.A.; Tesfamariam, S.; Gebrekirstos; Berhe, D.F. Anti-Diabetic Effect of Telmisartan through Its Partial Pparγ-Agonistic Activity. Diabetes. Metab. Syndr. Obes. Targets Ther. 2020, 13, 3627–3635. [Google Scholar] [CrossRef]
- Bonaventura, D.; Lunardi, C.N.; Rodrigues, G.J.; Neto, M.A.; Bendhack, L.M. A Novel Mechanism of Vascular Relaxation Induced by Sodium Nitroprusside in the Isolated Rat Aorta. Nitric oxide Biol. Chem. 2008, 18, 287–295. [Google Scholar] [CrossRef]
- Ataabadi, E.A.; Golshiri, K.; Jüttner, A.; Krenning, G.; Danser, A.H.J.; Roks, A.J.M. Nitric Oxide-CGMP Signaling in Hypertension. Hypertension 2020, 76, 1055–1068. [Google Scholar] [CrossRef]
- Shen, J.; Yakel, J.L. Nicotinic Acetylcholine Receptor-Mediated Calcium Signaling in the Nervous System. Acta Pharmacol. Sin. 2009, 30, 673–680. [Google Scholar] [CrossRef]
- Oakes, J.M.; Fuchs, R.M.; Gardner, J.D.; Lazartigues, E.; Yue, X. Nicotine and the Renin-Angiotensin System. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R895–R906. [Google Scholar] [CrossRef]
- Chen, X.-P.; Qian, L.-R. The Vasodilatory Action of Telmisartan on Isolated Mesenteric Artery Rings from Rats. Iran. J. Basic Med. Sci. 2015, 18, 974–978. [Google Scholar] [PubMed]
- Scalera, F.; Martens-Lobenhoffer, J.; Bukowska, A.; Lendeckel, U.; Täger, M.; Bode-Böger, S.M. Effect of Telmisartan on Nitric Oxide–Asymmetrical Dimethylarginine System. Hypertension 2008, 51, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A.; Schwarz, E.J.; Dimaculangan, D.D.; Lazar, M.A. Peroxisome Proliferator-Activated Receptor (PPAR) Gamma: Adipose-Predominant Expression and Induction Early in Adipocyte Differentiation. Endocrinology 1994, 135, 798–800. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Futawaka, K.; Hayashi, M.; Murakami, K.; Mitsutani, M.; Hatai, M.; Watamoto, Y.; Yoshikawa, N.; Nakamura, K.; Tagami, T.; et al. Cigarette Smoke Extract Modulates Functions of Peroxisome Proliferator-Activated Receptors. Biol. Pharm. Bull. 2019, 42, 1628–1636. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Hou, G.; Li, E.; Wang, Q.; Kang, J. PPAR Gamma Agonists Regulate Tobacco Smoke-Induced Toll like Receptor 4 Expression in Alveolar Macrophages. Respir. Res. 2014, 15, 28. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Mao, S.; Chen, S.; Zhang, W.; Liu, C. PPARs-Orchestrated Metabolic Homeostasis in the Adipose Tissue. Int. J. Mol. Sci. 2021, 22, 8974. [Google Scholar] [CrossRef]
- Gastaldelli, A.; Sabatini, S.; Carli, F.; Gaggini, M.; Bril, F.; Belfort-DeAguiar, R.; Positano, V.; Barb, D.; Kadiyala, S.; Harrison, S.; et al. PPAR-γ-Induced Changes in Visceral Fat and Adiponectin Levels Are Associated with Improvement of Steatohepatitis in Patients with NASH. Liver Int. Off. J. Int. Assoc. Study Liver 2021, 41, 2659–2670. [Google Scholar] [CrossRef]
- Astapova, O.; Leff, T. Adiponectin and PPARγ: Cooperative and Interdependent Actions of Two Key Regulators of Metabolism. Vitam. Horm. 2012, 90, 143–162. [Google Scholar] [CrossRef]
- Long, Q.; Lei, T.; Feng, B.; Yin, C.; Jin, D.; Wu, Y.; Zhu, X.; Chen, X.; Gan, L.; Yang, Z. Peroxisome Proliferator-Activated Receptor-Gamma Increases Adiponectin Secretion via Transcriptional Repression of Endoplasmic Reticulum Chaperone Protein ERp44. Endocrinology 2010, 151, 3195–3203. [Google Scholar] [CrossRef]
- Shen, X.; Li, H.; Li, W.; Wu, X.; Sun, Z.; Ding, X. Telmisartan Ameliorates AdipoR1 and AdipoR2 Expression via PPAR-γ Activation in the Coronary Artery and VSMCs. Biomed. Pharmacother. 2017, 95, 129–136. [Google Scholar] [CrossRef]
- Sowka, A.; Dobrzyn, P. Role of Perivascular Adipose Tissue-Derived Adiponectin in Vascular Homeostasis. Cells 2021, 10, 1485. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Guo, Y.; Xu, L.; Hou, N.; Han, F.; Sun, X. Induction of Haemeoxygenase-1 Directly Improves Endothelial Function in Isolated Aortas from Obese Rats through the Ampk-Pi3k/Akt-Enos Pathway. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2015, 36, 1480–1490. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Gu, L.; Si, Y.; Yin, W.; Zhao, M.; Zhang, T.; Chen, M. Macrovascular Protecting Effects of Berberine through Anti-Inflammation and Intervention of BKCa in Type 2 Diabetes Mellitus Rats. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Sancho, M.; Kyle, B.D. The Large-Conductance, Calcium-Activated Potassium Channel: A Big Key Regulator of Cell Physiology. Front. Physiol. 2021, 12, 750615. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.S.; Sponton, A.C.S.; Delbin, M.A. Perivascular Adipose Tissue and Microvascular Endothelial Dysfunction in Obese Mice: Beneficial Effects of Aerobic Exercise in Adiponectin Receptor (AdipoR1) and PeNOS(Ser1177). Clin. Exp. Pharmacol. Physiol. 2021, 48, 1430–1440. [Google Scholar] [CrossRef] [PubMed]
- Edwards, T.L.; Velez Edwards, D.R.; Villegas, R.; Cohen, S.S.; Buchowski, M.S.; Fowke, J.H.; Schlundt, D.; Long, J.; Cai, Q.; Zheng, W.; et al. HTR1B, ADIPOR1, PPARGC1A, and CYP19A1 and Obesity in a Cohort of Caucasians and African Americans: An Evaluation of Gene-Environment Interactions and Candidate Genes. Am. J. Epidemiol. 2012, 175, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Fan, J.; Meng, Z.; Wang, R.; Liu, C.; Liu, J.; Liang, B.; Wang, J.; Xie, Y.; Zhao, J.; et al. Nicotine Aggravates Vascular Adiponectin Resistance via Ubiquitin-Mediated Adiponectin Receptor Degradation in Diabetic Apolipoprotein E Knockout Mouse. Cell Death Dis. 2021, 12, 508. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Palaniyandi, S.S. Tissue-Specific Role and Associated Downstream Signaling Pathways of Adiponectin. Cell Biosci. 2021, 11, 77. [Google Scholar] [CrossRef]
- Moon, H.K.; Kang, P.; Lee, H.S.; Min, S.S.; Seol, G.H. Effects of 1,8-Cineole on Hypertension Induced by Chronic Exposure to Nicotine in Rats. J. Pharm. Pharmacol. 2014, 66, 688–693. [Google Scholar] [CrossRef]
- Almabrouk, T.A.M.; White, A.D.; Ugusman, A.B.; Skiba, D.S.; Katwan, O.J.; Alganga, H.; Guzik, T.J.; Touyz, R.M.; Salt, I.P.; Kennedy, S. High Fat Diet Attenuates the Anticontractile Activity of Aortic PVAT via a Mechanism Involving AMPK and Reduced Adiponectin Secretion. Front. Physiol. 2018, 9, 51. [Google Scholar] [CrossRef]
- Sundar, U.M.; Ugusman, A.; Chua, H.K.; Latip, J.; Aminuddin, A. Piper Sarmentosum Promotes Endothelial Nitric Oxide Production by Reducing Asymmetric Dimethylarginine in Tumor Necrosis Factor-α-Induced Human Umbilical Vein Endothelial Cells. Front. Pharmacol. 2019, 10, 1033. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | NCBI Code |
|---|---|---|
| Glyceraldehyde 3-phosphate dehydrogenase | Forward: 5′-CTCTCTGCTCCTCCCTGTTC-3′ Reverse: 5′-GGTAACCAGGCGTCCGATAC-3′ | NM_017008.4 |
| Adiponectin | Forward: 5′-TCTGGGAGATCCTCCTGTTGA-3′ Reverse: 5′-CGAAGTTGGTGGGCCAGAAT-3′ | NM_144744.3 |
| PPARγ | Forward: 5′-CTGGCTCCAAGTGTATGGGG-3′ Reverse: 5′-TTTGATTCTCGGGGCTACGG-3′ | NM_013124.3 |
| AdipoR1 | Forward: 5′-CTCCCTCCTGGATAGGGTCG-3′, Reverse: 5′-CAGCTTGAGGAGAGGTTGGG-3′ | NM_207587.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd Rami, A.Z.; Aminuddin, A.; Hamid, A.A.; Mokhtar, M.H.; Ugusman, A. Nicotine Impairs the Anti-Contractile Function of Perivascular Adipose Tissue by Inhibiting the PPARγ–Adiponectin–AdipoR1 Axis. Int. J. Mol. Sci. 2023, 24, 15100. https://doi.org/10.3390/ijms242015100
Abd Rami AZ, Aminuddin A, Hamid AA, Mokhtar MH, Ugusman A. Nicotine Impairs the Anti-Contractile Function of Perivascular Adipose Tissue by Inhibiting the PPARγ–Adiponectin–AdipoR1 Axis. International Journal of Molecular Sciences. 2023; 24(20):15100. https://doi.org/10.3390/ijms242015100
Chicago/Turabian StyleAbd Rami, Afifah Zahirah, Amilia Aminuddin, Adila A. Hamid, Mohd Helmy Mokhtar, and Azizah Ugusman. 2023. "Nicotine Impairs the Anti-Contractile Function of Perivascular Adipose Tissue by Inhibiting the PPARγ–Adiponectin–AdipoR1 Axis" International Journal of Molecular Sciences 24, no. 20: 15100. https://doi.org/10.3390/ijms242015100
APA StyleAbd Rami, A. Z., Aminuddin, A., Hamid, A. A., Mokhtar, M. H., & Ugusman, A. (2023). Nicotine Impairs the Anti-Contractile Function of Perivascular Adipose Tissue by Inhibiting the PPARγ–Adiponectin–AdipoR1 Axis. International Journal of Molecular Sciences, 24(20), 15100. https://doi.org/10.3390/ijms242015100