
The Inhibiting Effects of High-Dose Biochar Application on Soil Microbial Metagenomics and Rice (Oryza sativa L.) Production


Abstract
:1. Introduction
2. Results
2.1. Effects of High-Dose Biochar Application on Soil Properties
2.2. Effects of High-Dose Biochar Application on Rice Yield and Yield Components
2.3. Effects of High-Dose Biochar Application on Structure and Composition of Soil Microorganisms
2.3.1. Quality Evaluation of Metagenomic Sequences
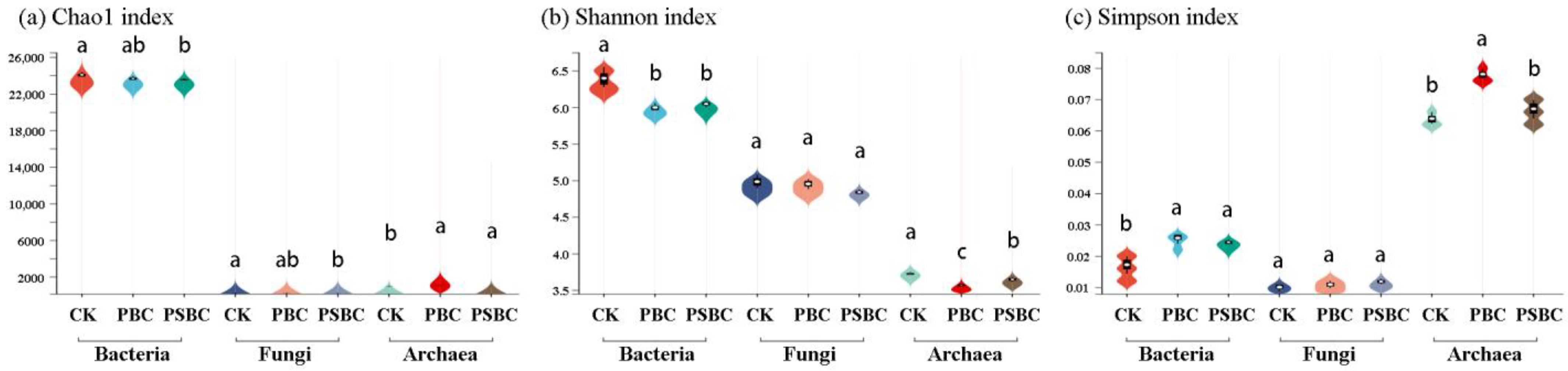
2.3.2. α Diversity Analysis of Soil Microorganisms
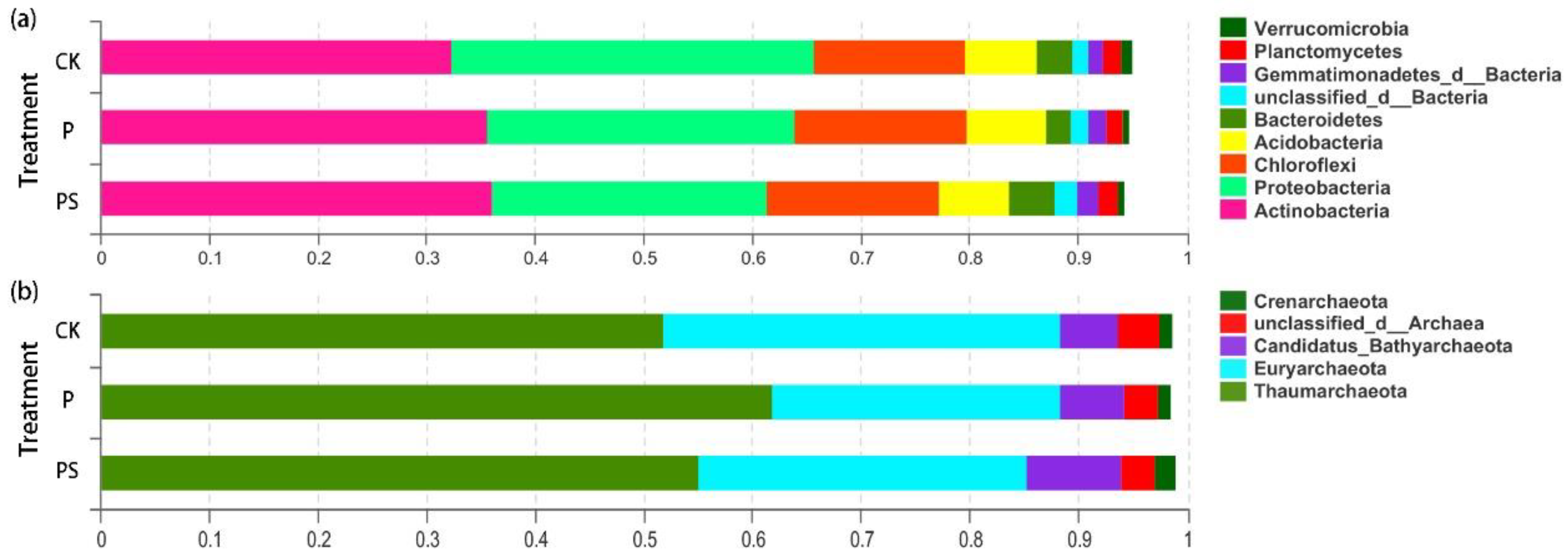
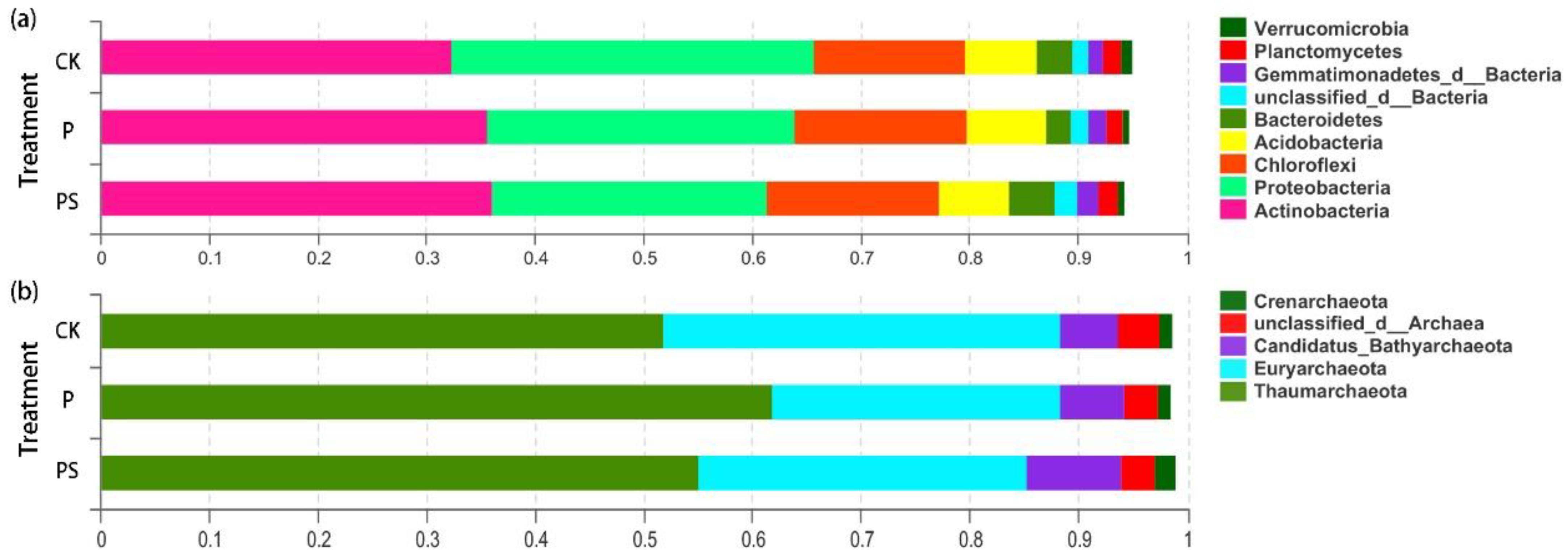
2.3.3. Differences in Structure and Composition of Soil Microorganisms
2.3.4. β Diversity Analysis of Soil Microorganisms
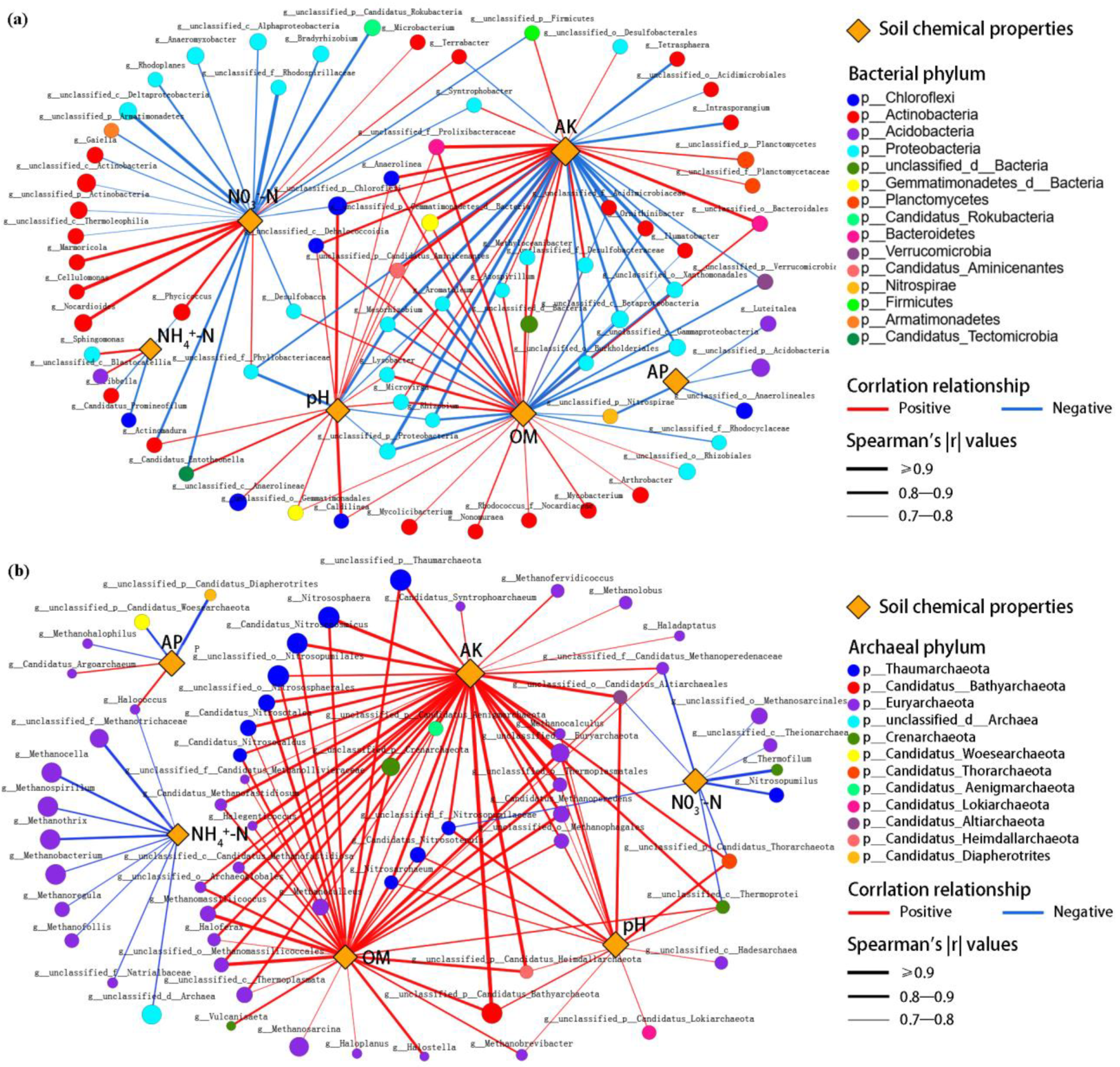
2.4. Relationships between Soil Microorganisms, Environmental Factors, and Rice Yield
2.5. Effect of High-Dose Biochar Application on Soil Microbial Metabolism
3. Discussion
3.1. Effects of High-Dose Biochar Application on Soil Fungal Communities
3.2. Effects of High-Dose Biochar Application on Soil Bacterial and Archaeal Communities
3.3. Interaction between High-Dose Biochar with Rice Production and Soil Microorganisms
3.4. Interaction between High-Dose Biochar Application and Microbial Metabolic Functions
3.5. Possible Inhibiting Mechanism of High-Dose Biochar Application on Rice Production
3.6. Applicability and Limitations
4. Materials and Methods
4.1. Experimental Site, Materials, and Treatments
4.2. Plant Sampling and Yield Measurement
4.3. Soil Sampling and Determination
4.4. Soil Metagenomic DNA Extraction, Sequencing, and Data Analysis
4.5. Statistical Analyses and Bioinformatics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ibrahim, M.M.; Tong, C.X.; Hu, K.; Zhou, B.Q.; Xing, S.H.; Mao, Y.L. Biochar-fertilizer interaction modifies N-sorption, enzyme activities and microbial functional abundance regulating nitrogen retention in rhizosphere soil. Sci. Total Environ. 2020, 739, 140065. [Google Scholar] [CrossRef]
- Fallah, N.; Yang, Z.Q.; Tayyab, M.; Zhang, C.F.; Abubakar, A.Y.; Lin, Z.L.; Pang, Z.Q.; Allison, A.; Zhang, H. Depth-dependent influence of biochar application on the abundance and community structure of diazotrophic under sugarcane growth. PLoS ONE 2021, 16, e0253970. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Y.; Jing, Y.M.; Chen, C.R.; Xiang, Y.Z.; Rashti, M.R.; Li, Y.T.; Deng, Q.; Zhang, R.D. Effects of biochar application on soil nitrogen transformation, microbial functional genes, enzyme activity, and plant nitrogen uptake: A meta-analysis of field studies. GCB Bioenergy 2021, 13, 1859–1873. [Google Scholar] [CrossRef]
- Hossain, M.Z.; Bahar, M.M.; Sarkar, B.; Donne, S.W.; Ok, Y.S.; Palansooriya, K.N.; Kirkham, M.B.; Chowdhury, S.; Bolan, N. Biochar and its importance on nutrient dynamics in soil and plant. Biochar 2020, 2, 379–420. [Google Scholar] [CrossRef]
- Zhou, F.; Geng, Z.C.; Xu, C.Y.; Liu, L.L.; Zhang, J.C.; Li, Q.Q.; Chen, S.L.; Wang, M.; Wang, H.L. Effect of biochar dosage on soil microbial biomass and carbon source metabolic activity. J. Plant Nutr. Fertil. 2019, 25, 1277–1289. [Google Scholar] [CrossRef]
- Feng, Y.F.; He, H.Y.; Xue, L.H.; Liu, Y.; Sun, H.J.; Guo, Z.; Wang, Y.M.; Zheng, X.B. The inhibiting effects of biochar-derived organic materials on rice production. J. Environ. Manag. 2021, 293, 112909. [Google Scholar] [CrossRef]
- Ghodake, G.S.; Shinde, S.K.; Kadam, A.A.; Saratale, R.G.; Saratale, G.D.; Kumar, M.; Palem, R.R.; AL-Shwaiman, H.A.; Elgorban, A.M.; Syed, A.; et al. Review on biomass feedstocks, pyrolysis mechanism and physicochemical properties of biochar: State-ofthe-art framework to speed up vision of circular bioeconomy. J. Clean. Prod. 2021, 297, 126645. [Google Scholar] [CrossRef]
- Ge, S.B.; Foong, S.Y.; Ma, N.L.; Liew, R.K.; Mahari, W.A.W.; Xia, C.L.; Yek, P.N.Y.; Peng, W.X.; Nam, W.L.; Lim, X.Y. Vacuum pyrolysis incorporating microwave heating and base mixture modification: An integrated approach to transform biowaste into eco-friendly bioenergy products. Renew. Sustain. Energy Rev. 2020, 127, 109871. [Google Scholar] [CrossRef]
- Wang, J.L.; Wang, S.Z. Preparation, modification and environmental application of biochar: A review. J. Clean. Prod. 2019, 227, 1002–1022. [Google Scholar] [CrossRef]
- Zhang, Q.F.; Zhang, D.H.; Xu, H.; Lu, W.Y.; Ren, X.J.; Cai, H.; Lei, H.W.; Huo, E.G.; Zhao, Y.F.; Qian, M. Biochar filled high-density polyethylene composites with excellent properties: Towards maximizing the utilization of agricultural wastes. Ind. Crop. Prod. 2020, 146, 112185. [Google Scholar] [CrossRef]
- He, M.J.; Xiong, X.N.; Wang, L.; Hou, D.Y.; Bolan, N.S.; Ok, Y.S.; Rinklebe, J.; Tsang, D.C.W. A critical review on performance indicators for evaluating soil biota and soil health of biochar-amended soils. J. Hazard. Mater. 2021, 414, 125378. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.X.; Chapman, S.J.; Li, Y.Y.; Yao, H.Y. Straw amendment to paddy soil stimulates denitrification but biochar amendment promotes anaerobic ammonia oxidation. J. Soils Sediments 2017, 17, 2428–2437. [Google Scholar] [CrossRef]
- Liu, J.Y.; Wang, Z.X.; Li, H.Y.; Hu, C.W.; Raymer, P.; Huang, Q.G. Effect of solid state fermentation of peanut shell on its dye adsorption performance. Bioresour. Technol. 2018, 249, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.B.; Feng, H.L.; Xu, C.S.; Xu, Q.; Fu, B.; Azwar, E.; Wei, Y.W.; Lam, S.S.; Liu, G.S. Exogenous application and interaction of biochar with environmental factors for improving functional diversity of rhizosphere’s microbial community and health. Chemosphere 2022, 294, 133710. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.W.; Gao, B.; Yao, Y.; Inyang, M.; Zhang, M.; Zimmerman, A.R.; Ro, K.S. Hydrogen peroxide modification enhances the ability of biochar (hydrochar) produced from hydrothermal carbonization of peanut hull to remove aqueous heavy metals: Batch and column tests. Chem. Eng. J. 2012, 200, 673–680. [Google Scholar] [CrossRef]
- Miltner, A.; Bombach, P.; Schmidtbrücken, B.; Kastner, M. SOM genesis: Microbial biomass as a significant source. Biogeochemistry 2012, 111, 41–55. [Google Scholar] [CrossRef]
- Viktória, F.; Éva, U.; Emese, V.; Mónika, M. Influence of red mud on soil microbial communities: Application and comprehensive evaluation of the Biolog EcoPlate approach as a tool in soil microbiological studies. Sci. Total Environ. 2017, 595, 903–911. [Google Scholar] [CrossRef]
- Insam, H.; Hutchinson, T.C.; Reber, H.H. Effects of heavy metal stress on the metabolic quotient of the soil microflora. Soil Biol. Biochem. 1995, 28, 691–694. [Google Scholar] [CrossRef]
- Feng, Y.Y.; Feng, Y.F.; Liu, Q.; Chen, S.; Hou, P.F.; Poinern, G.; Jiang, Z.T.; Fawcett, D.; Xue, L.H.; Lam, S.S.; et al. How does biochar aging affect NH3 volatilization and GHGs emissions from agricultural soils? Environ. Pollut. 2022, 294, 118598. [Google Scholar] [CrossRef]
- Natasha, N.; Shahid, M.; Khalid, S.; Bibi, I.; Naeem, M.A.; Niazi, N.K.; Tack, F.M.G.; Ippolito, J.A.; Rinklebe, J. Influence of biochar on trace element uptake, toxicity and detoxification in plants and associated health risks: A critical review. Crit. Rev. Env. Sci. Tec. 2021, 51, 2803–2843. [Google Scholar] [CrossRef]
- Berg, I.A.; Kockelkorn, D.; Buckel, W.; Fuchs, G. A 3-hydroxypropionate/4hydroxybutyrate autotrophic carbon dioxide assimilation pathway in archaea. Science 2007, 318, 1782–1786. [Google Scholar] [CrossRef]
- Geisseler, D.; Linquist, B.A.; Lazicki, P.A. Effect of fertilization on soil microorganisms in paddy rice systems—A meta-analysis. Soil Biol. Biochem. 2017, 115, 452–460. [Google Scholar] [CrossRef]
- Baker, B.J.; Anda, V.D.; Seitz, K.W.; Dombrowski, N.; Santoro, A.E.; Lloyd, K.G. Diversity, ecology and evolution of Archaea. Nat. Microbiol. 2020, 5, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Chelsea, C.J.; Dove, N.C.; Beman, J.M.; Stephen, C.H.; Emma, L.A. Meta-analysis reveals ammonia-oxidizing bacteria respond more strongly to nitrogen addition than ammonia-oxidizing archaea. Soil Biol. Biochem. 2016, 99, 158–166. [Google Scholar] [CrossRef]
- Williams, A.; Börjesson, G.; Hedlund, K. The effects of 55 years of different inorganic fertilizer regimes on soil properties and microbial community composition. Soil Biol. Biochem. 2013, 67, 41–46. [Google Scholar] [CrossRef]
- Carson, C.M.; Zeglinm, L.H. Long-term fire management history affects N-fertilization sensitivity, but not seasonality, of grassland soil microbial communities. Soil Biol. Biochem. 2018, 121, 231–239. [Google Scholar] [CrossRef]
- Pei, Y.X.; Yu, Z.S.; Ji, J.; Khan, A.; Li, X. Microbial community structure and function indicate the severity of chromium contamination of the Yellow River. Front. Microbiol. 2018, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Zong, H.Y.; Zheng, H.; Liu, G.C.; Chen, L.; Xing, B.S. Reduced nitrification and abundance of ammonia-oxidizing bacteria in acidic soil amended with biochar. Chemosphere 2015, 138, 576–583. [Google Scholar] [CrossRef]
- Manuel, D.B.; Giaramida, L.; Reich, P.B.; Khachane, A.N.; Hamonts, K.; Edwards, C.; Lawton, L.A.; Singh, B.K. Lack of functional redundancy in the relationship between microbial diversity and ecosystem functioning. J. Ecol. 2016, 104, 936–946. [Google Scholar] [CrossRef]
- He, Z.R.; Huang, X.T.; Fan, Y.; Yang, M.; Zhou, E.X. Metatranscriptomic analysis reveals rich mycoviral diversity in three major fungal pathogens of rice. Int. J. Mol. Sci. 2022, 23, 9192. [Google Scholar] [CrossRef]
- He, M.; Su, J.; Xu, Y.P.; Chen, J.H.; Chern, M.; Lei, M.L. Discovery of broad-spectrum fungicides that block septin-dependent infection processes of pathogenic fungi. Nat. Microbiol. 2020, 5, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wang, J.L.; Wang, C.; Wang, Q. The 19-years inorganic fertilization increased bacterial diversity and altered bacterial community composition and potential functions in a paddy soil. Appl. Soil Ecol. 2019, 144, 60–67. [Google Scholar] [CrossRef]
- Zhou, J.; Guan, D.W.; Zhou, B.K.; Zhao, B.S.; Ma, M.C.; Qin, J.; Jiang, X.; Chen, S.F.; Cao, F.M.; Shen, D.L.; et al. Influence of 34-years of fertilization on bacterial communities in an intensively cultivated black soil in northeast China. Soil Biol. Biochem. 2015, 90, 42–51. [Google Scholar] [CrossRef]
- Wang, H.; Boutton, T.W.; Xu, W.H.; Hu, G.Q.; Jiang, P.; Bai, E. Quality of fresh organic matter affects priming of soil organic matter and substrate utilization patterns of microbes. Sci. Rep. 2015, 5, 10102. [Google Scholar] [CrossRef] [PubMed]
- Aller, J.Y.; Kemp, P.F. Are archaea inherently less diverse than bacteria in the same environments? FEMS Microbiol. Ecol. 2008, 65, 74–87. [Google Scholar] [CrossRef]
- Ma, Q.; Qian, Y.S.; Yu, Q.Q.; Cao, Y.F.; Tao, R.R.; Zhu, M.; Ding, J.F.; Li, C.Y.; Guo, W.S.; Zhu, X.K. Controlled-release nitrogen fertilizer application mitigated N losses and modified microbial community while improving wheat yield and N use efficiency. Agr. Ecosyst. Environ. 2023, 349, 108445. [Google Scholar] [CrossRef]
- Liu, G.Q.; Wu, X.W.; Li, D.Y.; Jiang, L.G.; Zhuang, L. Long-term low dissolved oxygen operation decreases N2O emissions in the activated dludge process. Environ. Sci. Technol. 2021, 55, 6975–6983. [Google Scholar] [CrossRef]
- Van, D.B.E.M.; Marissa, B.; Gijs, K.J.; Robbert, K.; Van, L.M.C.M. DNRA and denitrification coexist over a broad range of acetate/N-NO3- ratios, in a chemostat enrichment culture. Front. Microbiol. 2016, 7, 1842. [Google Scholar] [CrossRef]
- Kraft, B.; Tegetmeyer, H.E.; Sharma, R.; Klotz, M.G.; Ferdelman, T.G.; Hettich, R.L.; Geelhoed, J.S.; Strous, M. The environmental controls that govern the end product of bacterial nitrate respiration. Science 2014, 345, 676–679. [Google Scholar] [CrossRef]
- Wang, S.Y.; Liu, C.L.; Wang, X.X.; Yuan, D.D.; Zhu, G.B. Dissimilatory nitrate reduction to ammonium (DNRA) in traditional municipal wastewater treatment plants in China: Widespread but low contribution. Water Res. 2020, 179, 115877. [Google Scholar] [CrossRef]
- Giblin, A.E.; Tobias, C.; Song, B.; Weston, N.; Banta, G.T.; Rivera-monroy, V.H. The importance of dissimilatory nitrate reduction to ammonium (DNRA) in the nitrogen cycle of coastal ecosystems. Oceanography 2013, 26, 124–131. [Google Scholar] [CrossRef]
- Wei, Z.; Senbayram, M.; Zhao, X.; Li, C.; Jin, K.; Wu, M.; Rahman, M.M.; Shan, J.; Yan, X.Y. Biochar amendment alters the partitioning of nitrate reduction by significantly enhancing DNRA in a paddy field. Biochar 2022, 4, 44. [Google Scholar] [CrossRef]
- Shi, D.L.; Wang, X.L.; Liu, A.K.; Hou, Z.F.; Liang, G.T. Response of microbial biomass carbon and nitrogen and rice quality in a yellow soil paddy field to biochar combined with nitrogen fertilizer. Huan Jing Ke Xue 2021, 42, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.C.; Liu, S.; Wang, Z.; Zhang, X.X.; Li, M.; Wu, B. Metagenomic profiles and antibiotic resistance genes in gut microbiota of mice exposed to arsenic and iron. Chemosphere 2014, 112, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.Y.; Shi, Y.Y.; Wang, R.; Feng, Y.; Wang, L.S.; Zhang, H.S.; Shi, X.Y.; Jing, G.Q.; Deng, P.; Song, T.Z. The transcription factor OsMYBc and an E3 ligase regulate expression of a K+ transporter during salt stress. Plant Physiol. 2022, 1, 843–859. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, Q.; Hu, W.; Qin, J.M.; Zheng, Y.R.; Wang, J.F.; Wang, Q.Q.; Xu, Y.X.; Guo, G.M.; Hu, S.; et al. Effects of plastic mulch film residues on soil-microbe-plant systems under different soil pH conditions. Chemosphere 2021, 267, 128901. [Google Scholar] [CrossRef]
- Wang, Q.Q.; Huang, Q.; Wang, J.X.; Muhammad, A.K.; Guo, G.; Liu, Y.; Hu, S.; Jin, F.M.; Wang, J.F.; Yu, Y.B. Dissolved organic carbon drives nutrient cycling via microbial community in paddy soil. Chemosphere 2021, 285, 131472. [Google Scholar] [CrossRef]
- Elrys, A.S.; Wang, J.; Metwally, M.A.S.; Cheng, Y.; Zhang, J.B.; Cai, Z.C.; Chang, S.X.; Muller, C. Global gross nitrification rates are dominantly driven by soil carbon-to-nitrogen stoichiometry and total nitrogen. Glob. Chang. Biol. 2021, 27, 6512–6524. [Google Scholar] [CrossRef]
- Chen, H.; Yuan, J.H.; Chen, G.L.; Zhao, X.; Wang, S.Q.; Wang, D.J.; Wang, L.; Wang, Y.J.; Wang, Y. Long-term biochar addition significantly decreases rice rhizosphere available phosphorus and its release risk to the environment. Biochar 2022, 4, 54. [Google Scholar] [CrossRef]
- Liao, J.Y.; Hu, A.; Zhao, Z.W.; Liu, X.R.; Jiang, C.; Zhang, Z.H. Biochar with large specific surface area recruits N2O-reducing microbes and mitigate N2O emission. Soil Biol. Biochem. 2021, 156, 108212. [Google Scholar] [CrossRef]
- Pang, Z.Q.; Huang, J.W.; Fallah, N.; Lin, W.X.; Yuan, Z.N.; Hu, C.H. Combining N rertilization with biochar affects root-shoot growth, rhizosphere soil properties and bacterial communities under sugarcane monocropping. Ind. Crop. Prod. 2022, 182, 114899. [Google Scholar] [CrossRef]
- Nan, Q.; Wang, C.; Wang, H.; Yi, Q.Q.; Liang, B.Q.; Xu, J.; Wu, W.X. Biochar drives microbially-mediated rice production by increasing soil carbon. J. Hazard. Mater. 2020, 387, 121680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yu, Z.W.; Zhang, Y.L.; Shi, Y. Impacts of fertilization optimization on soil nitrogen cycling and wheat nitrogen utilization under water-saving irrigation. Front. Plant Sci. 2022, 13, 878424. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Zhou, J.Z.; Deng, Y.; Shen, L.; Wen, C.Q.; Yan, Q.Y.; Ning, D.L.; Qin, Y.J.; Xue, K.; Wu, L.Y.; He, Z.L.; et al. Temperature mediates continental-scale diversity of microbes in forest soils. Nat. Commun. 2016, 7, 12083. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Yan, W.; Shangguan, Z. Impact of long-term N additions upon coupling between soil microbial community structure and activity, and nutrient-use efficiencies. Soil Biol. Biochem. 2015, 91, 151–159. [Google Scholar] [CrossRef]
- Schmidt, R.; Mitchell, J.; Scow, K. Cover cropping and no-till increase diversity and symbiotroph: Saprotroph ratios of soil fungal communities. Soil Biol. Biochem. 2019, 129, 99–109. [Google Scholar] [CrossRef]
- Wu, L.Y.; Wen, C.Q.; Qin, Y.H.; Yin, H.Q.; Tu, Q.C.; Joy, D.V.N.; Yuan, T.; Yuan, M.T.; Deng, Y.; Zhou, J.Z. Phasing amplicon sequencing on illumina miseq for robust environmental microbial community analysis. BMC Microbiol. 2015, 15, 125. [Google Scholar] [CrossRef]
- Mueller, R.C.; Paula, F.S.; Mirza, B.S.; Rodrigues, J.L.M.; Nüsslein, K.; Bohannan, B.J.M. Links between plant and fungal communities across a deforestation chronosequence in the amazon rainforest. ISME J. 2014, 8, 1548–1550. [Google Scholar] [CrossRef]
- Bolger, A.M.; Marc, L.; Bjoern, U. Trimmomatic: A flexible trimmer for illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, D.H.; Liu, C.M.; Luo, R.B.; Kunihiko, S.; Tak-Wah, L. Megahit: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de brujn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef]
- Fu, L.M.; Niu, B.F.; Zhu, Z.W.; Wu, S.T.; Li, W.Z. Cd-hit: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Luo, R.B.; Liu, B.H.; Xie, Y.L.; Li, Z.Y.; Huang, W.H.; Yuan, J.Y.; He, G.Z.; Chen, Y.X.; Pan, Q.; Liu, Y.J.; et al. Erratum: Soapdenovo2: An empirically improved memory-efficient short-read de novo assembler. GigaScience 2015, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Jia, S.Y.; Zhang, X.X.; Tong, Z.; Cheng, S.P.; Li, A.M. Metagenomic insights into chlorination effects on microbial antibiotic resistance in drinking water. Water Res. 2013, 47, 111–120. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Buchfink, B.; Reuter, K.; Drost, H.G. Sensitive protein alignments at tree-of-life scale using DIAMOND. Nat. Methods 2021, 18, 366–368. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, 277–280. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, Y.M.; Huang, H.Y.; Mou, L.C.; Ru, J.L.; Zhao, J.H.; Xiao, S. Long-term and high-concentration heavy-metal contamination strongly influences the microbiome and functional genes in yellow river sediments. Sci. Total Environ. 2018, 637–638, 1400–1412. [Google Scholar] [CrossRef]
- Lawson, C.E.; Wu, S.; Bhattacharjee, A.S.; Hamilton, J.J.; McMahon, K.D.; Goel, R.; Noguera, D.R. Metabolic network analysis reveals microbial community interactions in anammox granules. Nat. Commun. 2017, 8, 15416. [Google Scholar] [CrossRef]
- Li, P.F.; Liu, J.; Jiang, C.Y.; Wu, M.; Liu, M.; Li, Z.P. Distinct successions of common and rare bacteria in soil under humic acid amendment—A microcosm study. Front. Microbiol. 2019, 10, 2271. [Google Scholar] [CrossRef]
- Stegen, J.C.; Lin, X.J.; Konopka, A.E.; Fredrickson, J.K. Stochastic and deterministic assembly processes in subsurface microbial communities. ISME J. 2012, 6, 1653–1664. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.N.; Burton, O.T.; Wise, P.; Zhang, Y.Q.; Hobson, S.A.; Lloret, M.G.; Chehoud, C.; Kuczynski, J.; DeSantis, T.; Warrington, J.; et al. A microbiota signature associated with experimental food allergy promotes allergic sensitization and anaphylaxis. J. Allergy Clin. Immun. 2013, 131, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.H.; Li, S.F.; Yang, L.; Huang, P.; Li, W.J.; Wang, S.Y.; Zhao, G.P.; Zhang, M.H.; Pang, X.Y.; Yan, Z.; et al. Structural modulation of gut microbiota in life-long calorie-restricted mice. Nat. Commun. 2013, 4, 2163. [Google Scholar] [CrossRef] [PubMed]
- Goethem, M.W.V.; Pierneef, R.; Bezuidt, O.K.I.; Peer, Y.V.D.; Cowan, D.A.; Makhalanyane, T.P. A reservoir of ‘historical’ antibiotic resistance genes in remote pristine antarctic soils. Microbiome 2018, 6, 40. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | CK | PBC | PSBC |
|---|---|---|---|
| pH | 6.93 ± 0.01 b | 6.97 ± 0.02 a | 6.98 ± 0.03 a |
| NO3−–N (mg kg−1) | 9.83 ± 0.15 a | 7.60 ± 0.10 c | 8.97 ± 0.15 b |
| NH4+–N (mg kg−1) | 10.77 ± 0.40 b | 13.67 ± 0.54 a | 10.24 ± 0.42 b |
| AP (mg kg−1) | 30.16 ± 1.65 b | 28.52 ± 1.35 b | 32.86 ± 0.85 a |
| AK (mg kg−1) | 27.00 ± 0.56 c | 38.16 ± 1.92 b | 50.82 ± 1.96 a |
| OM (g kg−1) | 40.75 ± 0.46 c | 49.77 ± 0.79 b | 72.97 ± 1.17 a |
| Parameters | Topsoil | Biochar | |
|---|---|---|---|
| PBC | PSBC | ||
| pH | 6.38 | 10.87 | 11.69 |
| NO3−–N (mg kg−1) | 5.1 | 3.42 | 3.64 |
| NH4+–N (mg kg−1) | 43 | 3.93 | 3.72 |
| AP (mg kg−1) | 63.2 | / | / |
| AK (mg kg−1) | 118.37 | / | / |
| OM (g kg−1) | 8.64 | / | / |
| Specific surface area (m2 g−1) | / | 166.70 | 128.53 |
| Productivity (%) | / | 38.77 | 41.37 |
| Ashes (%) | / | 21.64 | 23.85 |
| Treatment | CK | PBC | PSBC |
|---|---|---|---|
| Spike (pot−1) | 53.00 ± 1.00 a | 45.67 ± 2.52 b | 49.00 ± 3.61 ab |
| Grain (spike−1) | 82.00 ± 3.61 a | 85.33 ± 2.08 a | 87.33 ± 2.08 a |
| Thousand-grain weight (g) | 26.17 ± 0.43 a | 25.42 ± 0.19 b | 25.88 ± 0.24 ab |
| Percentage of setting (%) | 0.903 ± 0.021 a | 0.817 ± 0.015 b | 0.900 ± 0.010 a |
| Yield (g pot−1) | 102.66 ± 2.96 a | 81.06 ± 8.01 b | 99.76 ± 9.22 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, N.; Yu, Q.; Song, L.; Sheng, H. The Inhibiting Effects of High-Dose Biochar Application on Soil Microbial Metagenomics and Rice (Oryza sativa L.) Production. Int. J. Mol. Sci. 2023, 24, 15043. https://doi.org/10.3390/ijms242015043
Zhu N, Yu Q, Song L, Sheng H. The Inhibiting Effects of High-Dose Biochar Application on Soil Microbial Metagenomics and Rice (Oryza sativa L.) Production. International Journal of Molecular Sciences. 2023; 24(20):15043. https://doi.org/10.3390/ijms242015043
Chicago/Turabian StyleZhu, Nanyan, Qiaoqiao Yu, Lingqi Song, and Haijun Sheng. 2023. "The Inhibiting Effects of High-Dose Biochar Application on Soil Microbial Metagenomics and Rice (Oryza sativa L.) Production" International Journal of Molecular Sciences 24, no. 20: 15043. https://doi.org/10.3390/ijms242015043
APA StyleZhu, N., Yu, Q., Song, L., & Sheng, H. (2023). The Inhibiting Effects of High-Dose Biochar Application on Soil Microbial Metagenomics and Rice (Oryza sativa L.) Production. International Journal of Molecular Sciences, 24(20), 15043. https://doi.org/10.3390/ijms242015043