

Identification of SALL4 Expressing Islet-1+ Cardiovascular Progenitor Cell Clones


Abstract
1. Introduction
2. Results
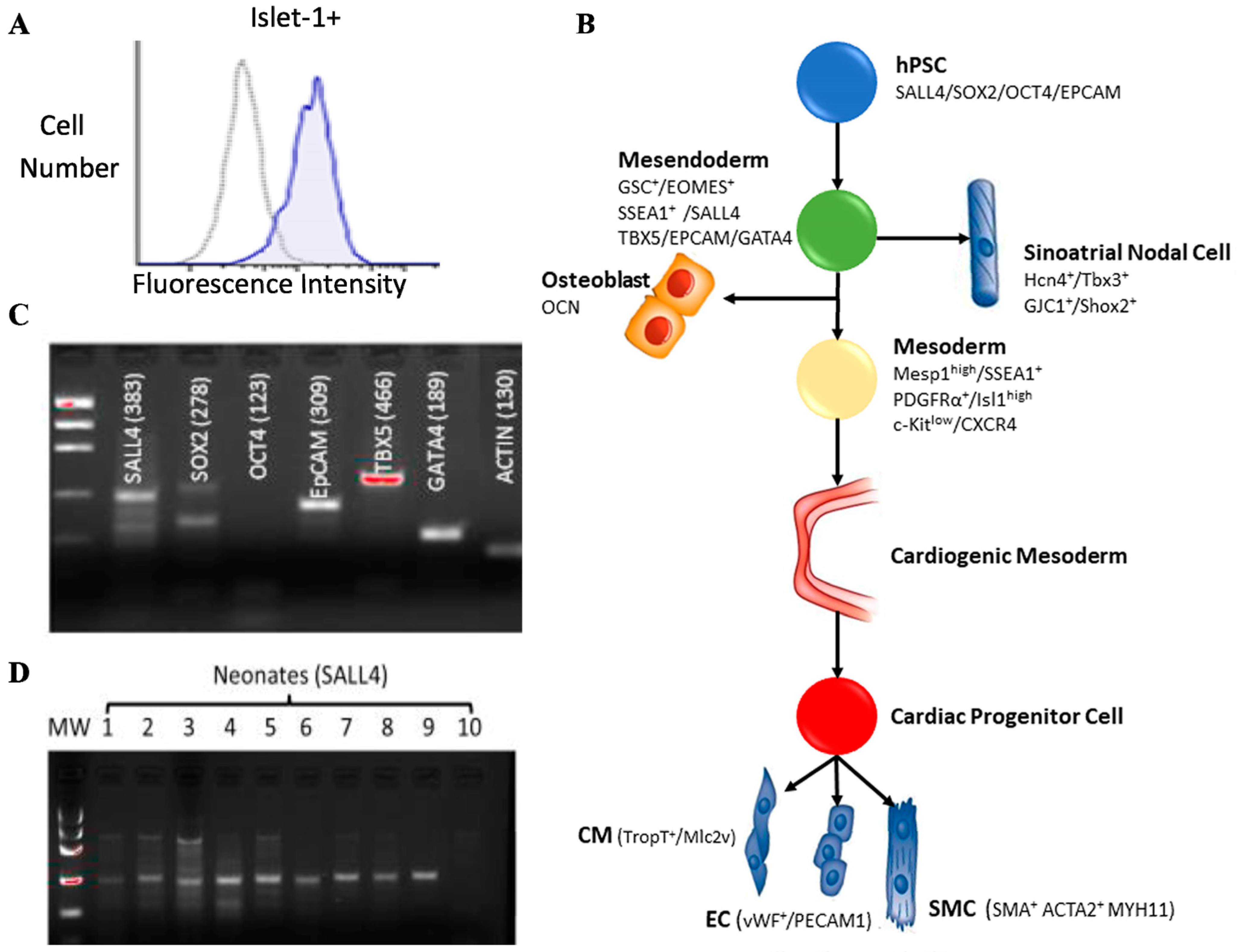
2.1. Islet-1+ Cardiovascular Progenitor Cell Clones Express Early-Stage Markers
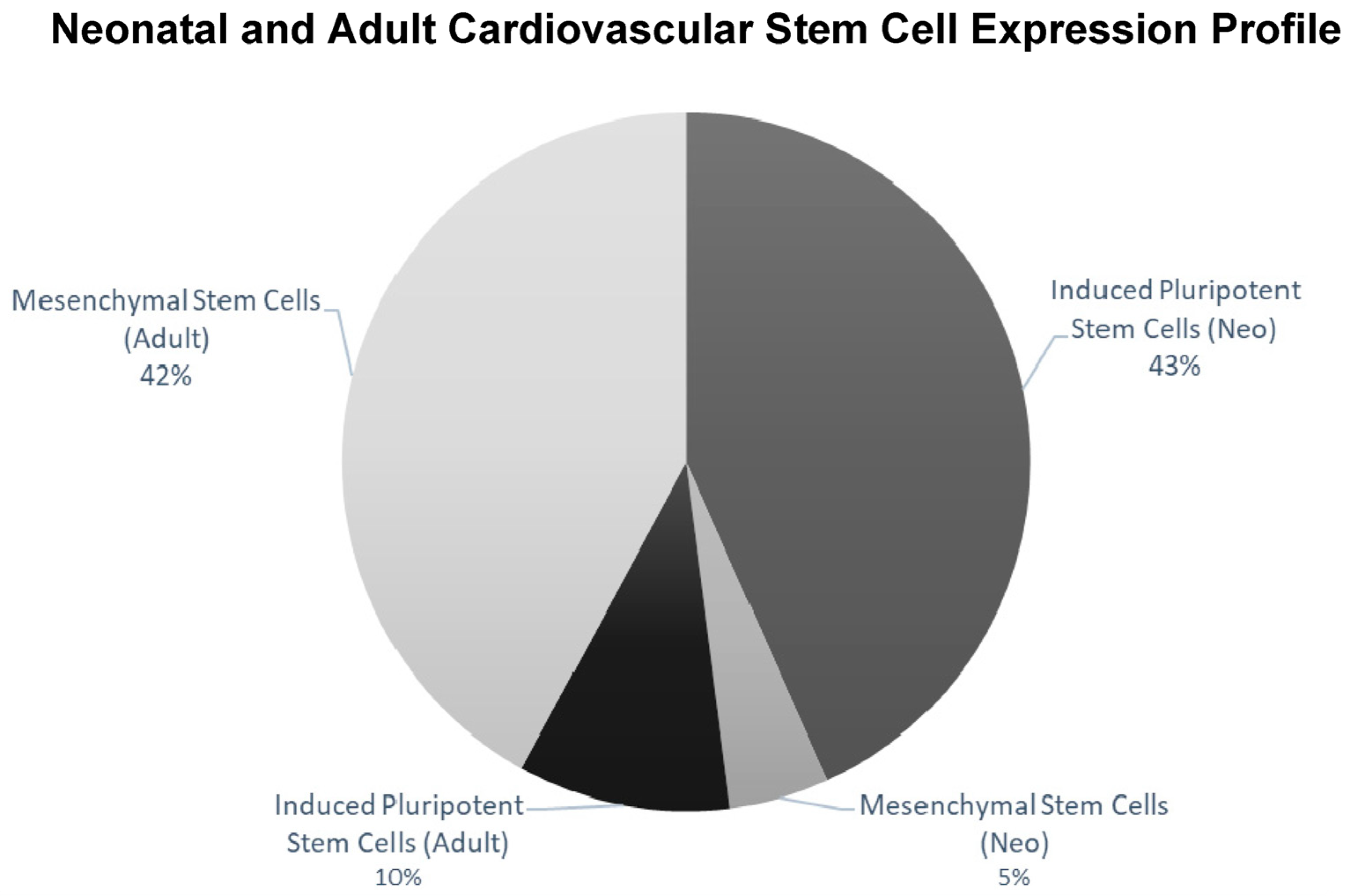
2.2. Islet-1+ Neonatal CPC Clones Express Characteristics of Enhanced Stemness
2.3. Transcriptomic Profiling Reveals SALL4 Interacts with Pluripotency Factors
2.4. Transcriptomic Profiling Elucidates Pathway and Stage-Specific Differences between Neonatal CPC Clones
2.5. Select Islet-1+ CPC Clones Express TFAP2C
3. Discussion
4. Methods
4.1. Ethics Statement and Maintenance of Cardiac Progenitor Cells
4.2. Cardiac Progenitor Cell Expansion
4.3. Purification of Total RNA
4.4. RNA Sequencing
4.5. Transcriptomic Analysis
4.6. RT-PCR
4.7. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CPC | Cardiac Progenitor Cell |
| Isl-1 | Islet-1 |
| SALL4 | Spalt-like transcription factor 4 |
| SOX2 | SRY-Box Transcription Factor 2 |
| EpCAM | Epithelial Cellular Adhesion Molecule |
| TBX5 | T-box transcription factor 5 |
| TFAP2C | Transcription Factor AP-2 Gamma |
| hESC | Human Embryonic Stem Cell |
| iPSC | Induced Pluripotent Stem Cell |
| RT-PCR | Reverse Transcription Polymerase Chain Reaction |
| IPA | Ingenuity Pathway Analysis |
| FHF | First Heart Field |
| SHF | Second Heart Field |
References
- Cai, C.-L.; Liang, X.; Shi, Y.; Chu, P.-H.; Pfaff, S.L.; Chen, J.; Evans, S. Isl1 Identifies a Cardiac Progenitor Population that Proliferates Prior to Differentiation and Contributes a Majority of Cells to the Heart. Dev. Cell 2003, 5, 877–889. [Google Scholar] [CrossRef]
- Andersen, P.; Tampakakis, E.; Jimenez, D.V.; Kannan, S.; Miyamoto, M.; Shin, H.K.; Saberi, A.; Murphy, S.; Sulistio, E.; Chelko, S.P.; et al. Precardiac organoids form two heart fields via Bmp/Wnt signaling. Nat. Commun. 2018, 9, 3140. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Zhou, B.; Pu, W.T. Reassessment of Isl1 and Nkx2-5 cardiac fate maps using a Gata4-based reporter of Cre activity. Dev. Biol. 2008, 323, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Tyser, R.C.V.; Mahammadov, E.; Nakanoh, S.; Vallier, L.; Scialdone, A.; Srinivas, S. Single-cell transcriptomic characterization of a gastrulating human embryo. Nature 2021, 600, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Muhr, J.; Ackerman, K.M. Embryology, Gastrulation. 2021 Apr 15. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Pastor, W.A.; Liu, W.; Chen, D.; Ho, J.; Kim, R.; Hunt, T.J.; Lukianchikov, A.; Liu, X.; Polo, J.M.; Jacobsen, S.E.; et al. TFAP2C regulates transcription in human naive pluripotency by opening enhancers. Nature 2018, 20, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Menasché, P.; Vanneaux, V.; Hagège, A.; Bel, A.; Cholley, B.; Cacciapuoti, I.; Parouchev, A.; Benhamouda, N.; Tachdjian, G.; Tosca, L.; et al. Human embryonic stem cell-derived cardiac progenitors for severe heart failure treatment: First clinical case report. Eur. Heart J. 2015, 36, 2011–2017. [Google Scholar] [CrossRef]
- Yang, L.; Cai, C.-L.; Lin, L.; Qyang, Y.; Chung, C.; Monteiro, R.; Mummery, C.; Fishman, G.; Cogen, A.; Evans, S. Isl1Cre reveals a common Bmp pathway in heart and limb development. Development 2006, 133, 1575–1585. [Google Scholar] [CrossRef]
- Akiyama, R.; Kawakami, H.; Wong, J.; Oishi, I.; Nishinakamura, R.; Kawakami, Y. Sall4-Gli3 system in early limb progenitors is essential for the development of limb skeletal elements. Proc. Natl. Acad. Sci. USA 2015, 112, 5075–5080. [Google Scholar] [CrossRef]
- Belair, D.G.; Lu, G.; Waller, L.E.; Gustin, J.A.; Collins, N.D.; Kolaja, K.L. Thalidomide Inhibits Human iPSC Mesendoderm Differentiation by Modulating CRBN-dependent Degradation of SALL4. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Stavish, D.; Böiers, C.; Price, C.; Frith, T.J.R.; Halliwell, J.; Saldaña-Guerrero, I.; Wray, J.; Brown, J.; Carr, J.; James, C.; et al. Generation and trapping of a mesoderm biased state of human pluripotency. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Fisher, J.B.; Pulakanti, K.; Rao, S.; Duncan, S.A. GATA6 is essential for endoderm formation from human pluripotent stem cells. Biol. Open 2017, 6, 1084–1095. [Google Scholar] [CrossRef] [PubMed]
- Mononen, M.M.; Leung, C.Y.; Xu, J.; Chien, K.R. Trajectory mapping of human embryonic stem cell cardiogenesis reveals lineage branch points and an ISL1 progenitor-derived cardiac fibroblast lineage. STEM CELLS 2020, 38, 1267–1278. [Google Scholar] [CrossRef] [PubMed]
- Tanimura, N.; Saito, M.; Ebisuya, M.; Nishida, E.; Ishikawa, F. Stemness-related Factor Sall4 Interacts with Transcription Factors Oct-3/4 and Sox2 and Occupies Oct-Sox Elements in Mouse Embryonic Stem Cells. J. Biol. Chem. 2013, 288, 5027–5038. [Google Scholar] [CrossRef] [PubMed]
- Tahara, N.; Kawakami, H.; Chen, K.Q.; Anderson, A.; Peterson, M.Y.; Gong, W.; Shah, P.; Hayashi, S.; Nishinakamura, R.; Nakagawa, Y.; et al. Sall4 regulates neuromesodermal progenitors and their descendants during body elongation in mouse embryos. Development 2019, 146, 177659. [Google Scholar] [CrossRef]
- Yang, J. SALL4 as a transcriptional and epigenetic regulator in normal and leukemic hematopoiesis. Biomark. Res. 2018, 6, 1–9. [Google Scholar] [CrossRef]
- Koshiba-Takeuchi, K.; Takeuchi, J.K.; Arruda, E.P.; Kathiriya, I.S.; Mo, R.; Hui, C.-C.; Srivastava, D.; Bruneau, B.G. Cooperative and antagonistic interactions between Sall4 and Tbx5 pattern the mouse limb and heart. Nat. Genet. 2005, 38, 175–183. [Google Scholar] [CrossRef]
- Abboud, N.; Moore-Morris, T.; Hiriart, E.; Yang, H.; Bezerra, H.; Gualazzi, M.-G.; Stefanovic, S.; Guénantin, A.-C.; Evans, S.M.; Pucéat, M. A cohesin–OCT4 complex mediates Sox enhancers to prime an early embryonic lineage. Nat. Commun. 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Churko, J.M.; Garg, P.; Treutlein, B.; Venkatasubramanian, M.; Wu, H.; Lee, J.; Wessells, Q.N.; Chen, S.-Y.; Chen, W.-Y.; Chetal, K.; et al. Defining human cardiac transcription factor hierarchies using integrated single-cell heterogeneity analysis. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Valcourt, J.R.; Huang, R.E.; Kundu, S.; Venkatasubramanian, D.; Kingston, R.E.; Ramanathan, S. Modulating mesendoderm competence during human germ layer differentiation. Cell Rep. 2021, 37, 109990. [Google Scholar] [CrossRef]
- Yeo, J.-C.; Ng, H.H. The transcriptional regulation of pluripotency. Cell Res. 2012, 23, 20–32. [Google Scholar] [CrossRef]
- Young, W. Future of Muse Cells. Adv. Exp. Med. Biol. 2018, 1103, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, T.I.; Appleby, N.; Tsay, E.; Martinez, J.J.; Bailey, L.; Hasaniya, N.; Kearns-Jonker, M. Human Neonatal Cardiovascular Progenitors: Unlocking the Secret to Regenerative Ability. PLoS ONE 2013, 8, e77464. [Google Scholar] [CrossRef] [PubMed]
- Pinto, J.P.; Kalathur, R.K.R.; Oliveira, D.; Barata, T.; Machado, R.; Machado, S.; Pacheco-Leyva, I.; Duarte, G.I.D.S.; Futschik, M.E. StemChecker: A web-based tool to discover and explore stemness signatures in gene sets. Nucleic Acids Res. 2015, 43, W72–W77. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence | Reverse Sequence |
|---|---|---|
| SALL4 | CACAAGTGTCGGAGCAGTCT | CCGTCCGTACCTAACAGAGC |
| SOX2 | AACCAGCGCATGGACAGTTA | GACTTGACCACCGAACCCAT |
| EpCAM | TGGGGAACAACTGGATCTGG | CCCACGCACACACATTTGTAA |
| TBX5 | CTCAGTCCCCCGGAACAAC | CACGTACCTCCCAGCTCAAG |
| Tfap2c | TGGTTGGTTTTTGTGTCCGC | TTGCTTCGTGCCTACCCTTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteon, A.; Hughes, L.; Camberos, V.; Kearns-Jonker, M. Identification of SALL4 Expressing Islet-1+ Cardiovascular Progenitor Cell Clones. Int. J. Mol. Sci. 2023, 24, 1780. https://doi.org/10.3390/ijms24021780
Monteon A, Hughes L, Camberos V, Kearns-Jonker M. Identification of SALL4 Expressing Islet-1+ Cardiovascular Progenitor Cell Clones. International Journal of Molecular Sciences. 2023; 24(2):1780. https://doi.org/10.3390/ijms24021780
Chicago/Turabian StyleMonteon, Andrea, Lorelei Hughes, Victor Camberos, and Mary Kearns-Jonker. 2023. "Identification of SALL4 Expressing Islet-1+ Cardiovascular Progenitor Cell Clones" International Journal of Molecular Sciences 24, no. 2: 1780. https://doi.org/10.3390/ijms24021780
APA StyleMonteon, A., Hughes, L., Camberos, V., & Kearns-Jonker, M. (2023). Identification of SALL4 Expressing Islet-1+ Cardiovascular Progenitor Cell Clones. International Journal of Molecular Sciences, 24(2), 1780. https://doi.org/10.3390/ijms24021780