
Ectopic Expression of Sugarcane ScAMT1.1 Has the Potential to Improve Ammonium Assimilation and Grain Yield in Transgenic Rice under Low Nitrogen Stress

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
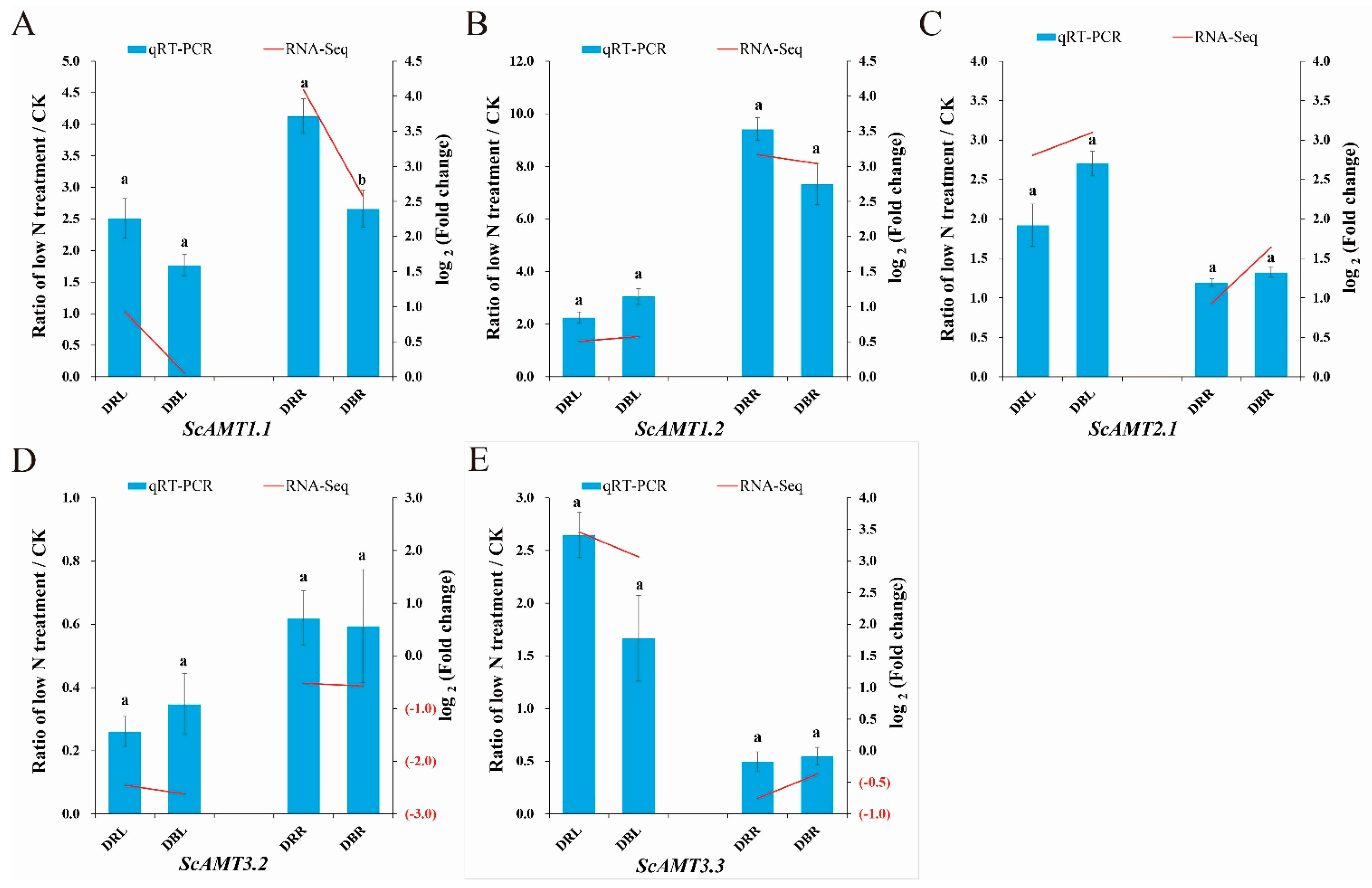
2.1. The ScAMT1.1 Gene Responded Positively to Low N Stress in Sugarcane
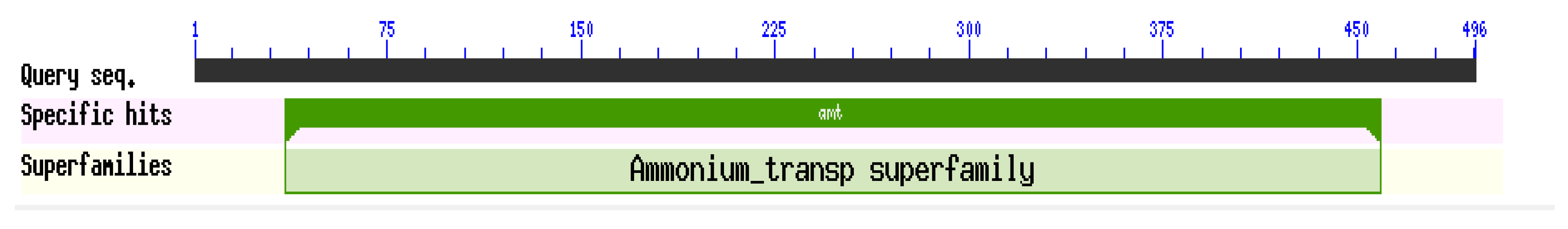
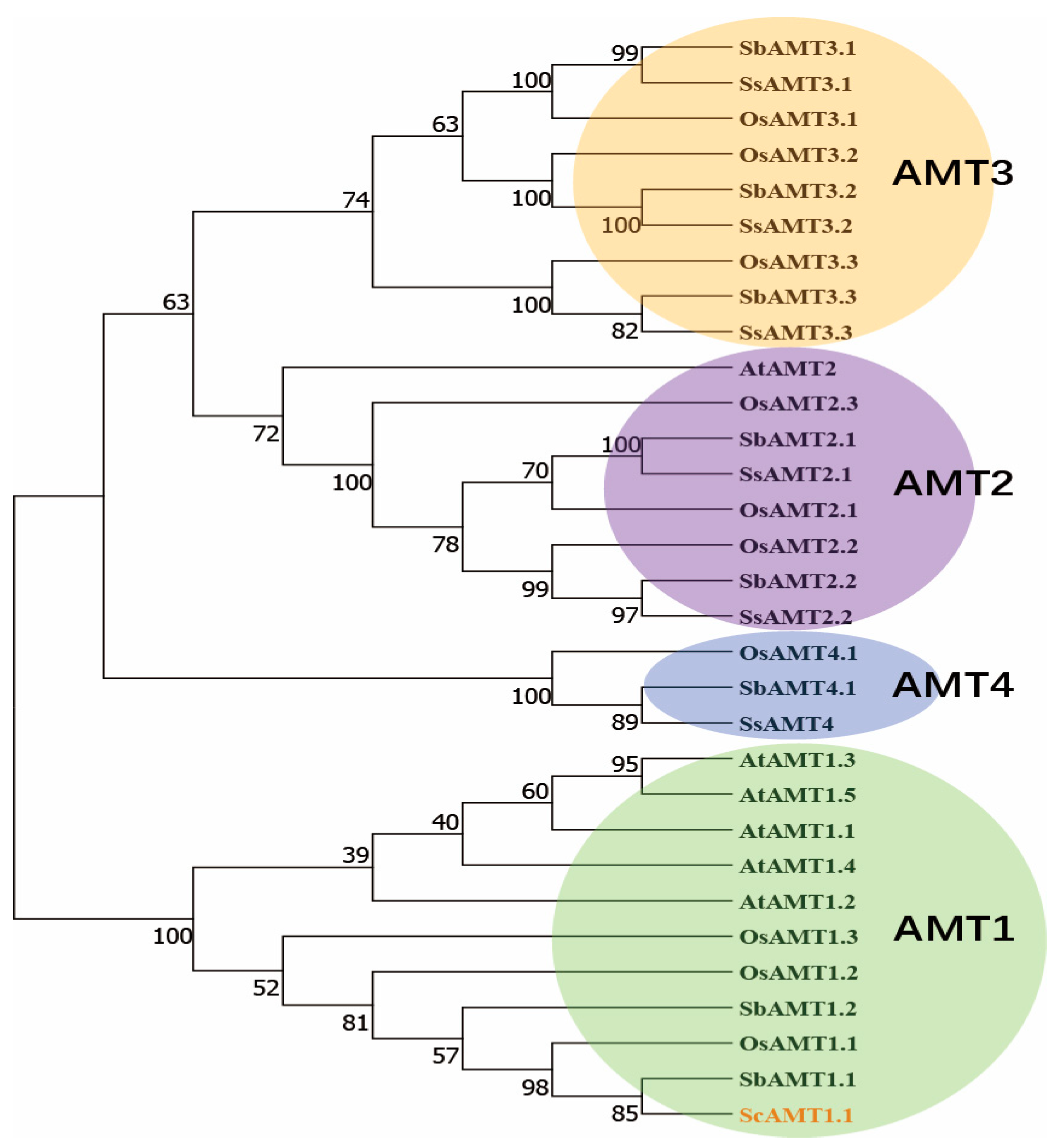
2.2. Gene Cloning and Bioinformatics Analysis of ScAMT1.1 Gene
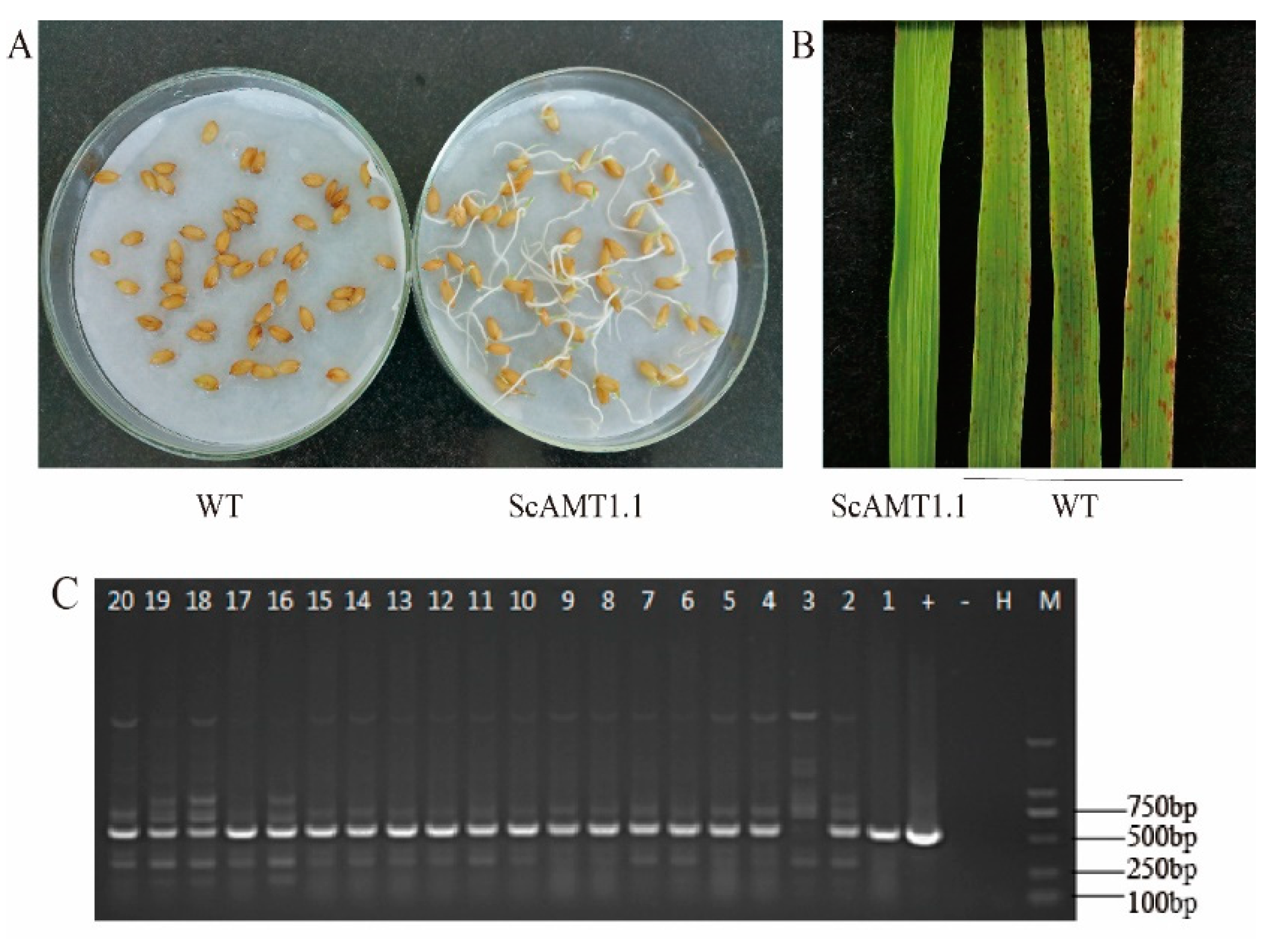
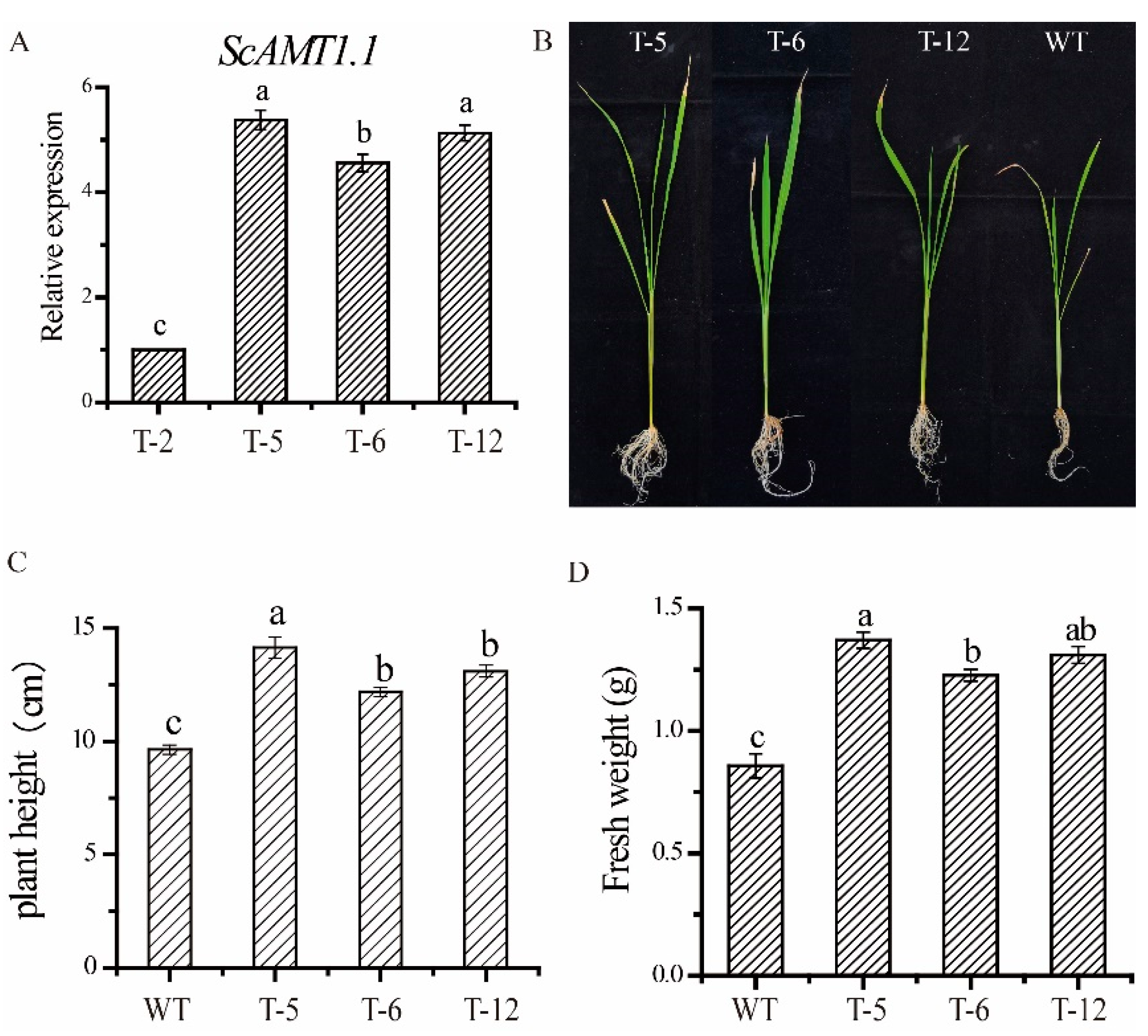
2.3. Identification of the ScAMT1.1 Overexpressed Rice
2.4. Phenotypic Index Analysis Shows Overexpressing ScAMT1.1 Enhances Plants’ Height and Fresh Weight in the Transgenic Rice under Low N Stress
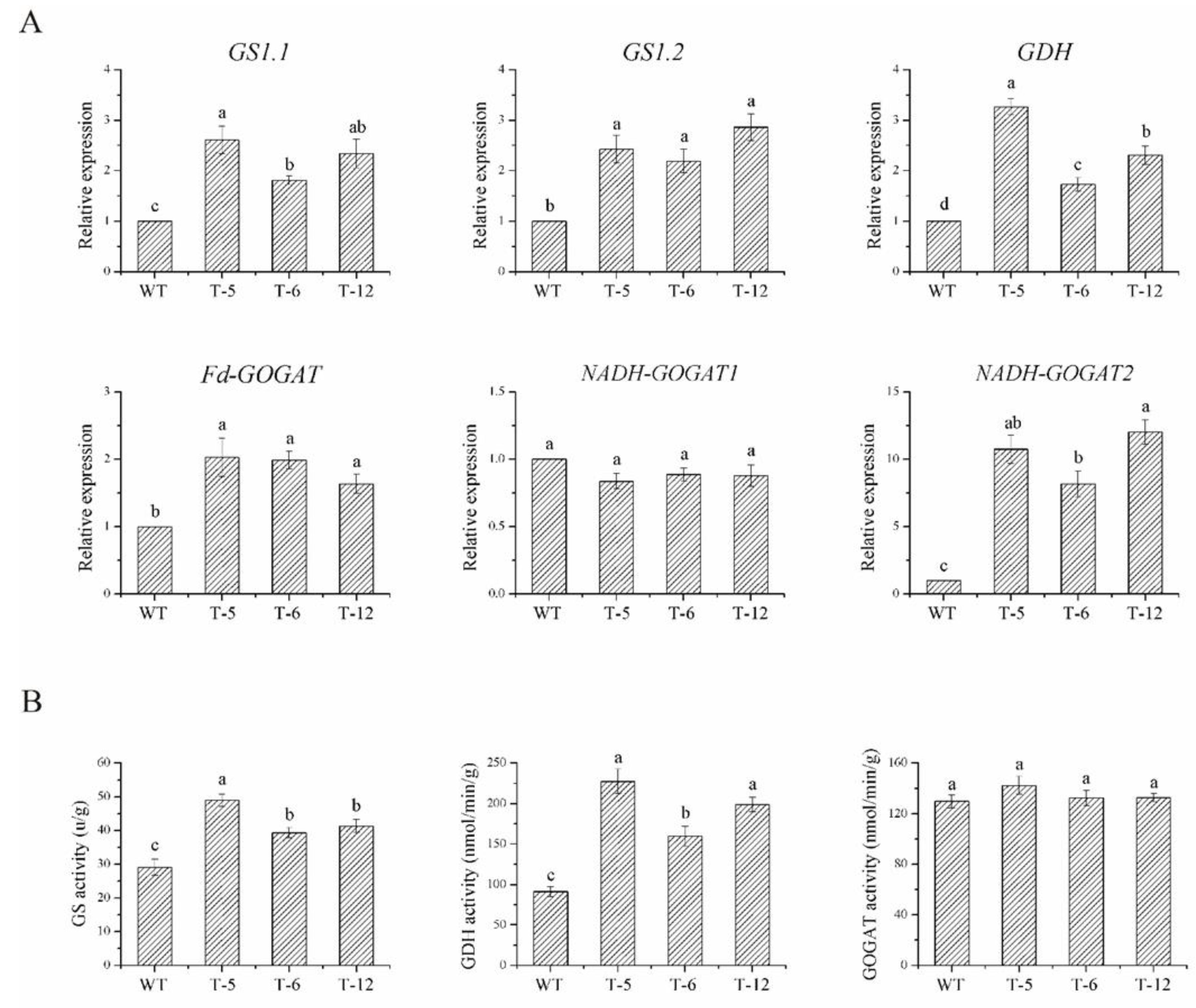
2.5. The Four Ammonium Assimilation Related Enzymes Were More Active in the Transgenic Lines Than That in the Wild-Type Plants
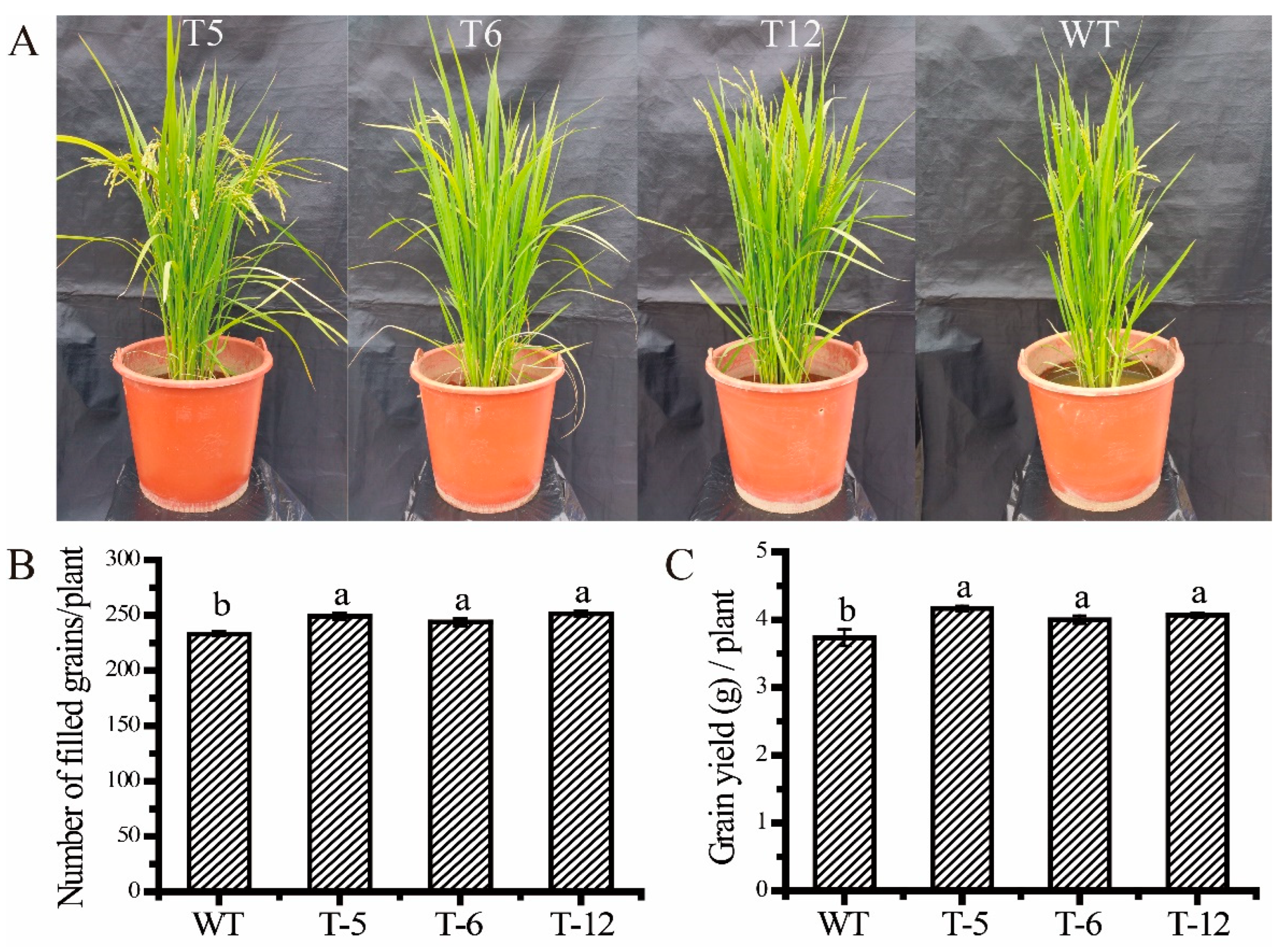
2.6. Overexpressed ScAMT1.1 Enhanced Grain Number and Grain Yield of the Transgenic Plants in the Pot Experiment under Low N Condition
3. Discussion
4. Materials and Methods
4.1. Materials and Treatments
4.2. RNA-Seq Data Analysis and qRT-PCR Experiment
4.3. Gene Cloning and Bioinformatics Analysis of Sugarcane ScAMT1.1 Gene
4.4. Generation of the Transgenic Rice Overexpressed ScAMT1.1
4.5. Determination of Enzyme Activities Involved in Ammonium Assimilation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sonoda, Y.; Ikeda, A.; Saiki, S.; von Wiren, N.; Yamaya, T.; Yamaguchi, J. Distinct expression and function of three ammonium transporter genes (OsAMT1;1-1;3). Plant Cell Physiol. 2003, 44, 726–734. [Google Scholar] [CrossRef]
- Franco, H.C.J.; Otto, R.; Faroni, C.E.F.; Vitti, A.C.; de Oliveira, E.C.A.; Trivelin, P.C.O. Nitrogen in sugarcane derived from fertiliser under Brazilian field conditions. Field Crop. Res. 2011, 121, 29–41. [Google Scholar] [CrossRef]
- Kingston, G.; Anink, M.C.; Allen, D. Acquisition of nitrogen by ratoon crops of sugarcane as influenced by waterlogging and split applications. Proc. Conf. Aust. Soc. Sugar Cane Technol. 2008, 30, 202–211. [Google Scholar]
- Meyer, J.H.; Schumann, A.W.; Wood, R.A.; Nixon, D.; Van den Berg, M. Recent advances to improve nitrogen use efficiency of sugarcane in the South African sugar industry. Proc. Int. Soc. Sugar Cane Technol. 2007, 26, 238–246. [Google Scholar]
- Robinson, N.; Brackin, R.; Vinall, K.; Soper, F.; Holst, J.; Gamage, H.; Paungfoo-Lonhienne, C.; Rennenberg, H.; Lakshmanan, P.; Schmidt, S. Nitrate paradigm does not hold up for sugarcane. PLoS ONE 2011, 6, e19045. [Google Scholar] [CrossRef]
- Hirel, B.; Bertin, P.; Quilleré, I.; Bourdoncle, W.; Attagnant, C.; Dellay, C.; Gouy, A.; Cadiou, S.; Retailliau, C.; Falque, M.; et al. Towards a better understanding of the genetic and physiological basis for nitrogen use efficiency in maize. Plant Physiol. 2001, 125, 1258–1270. [Google Scholar] [CrossRef]
- Wu, Z.L.; Gao, X.N.; Zhang, N.N.; Feng, X.M.; Huang, Y.H.; Zeng, Q.Y.; Wu, J.Y.; Zhang, J.S.; Qi, Y.W. Genome-wide identification and transcriptional analysis of ammonium transporters in Saccharum. Genomics 2021, 113, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J.; Sukrapanna, S.S.; Warner, R.L. Root respiration associated with ammonium and nitrate absorption and assimilation by barley. Plant Physiol. 1992, 99, 1294–1301. [Google Scholar] [CrossRef]
- Ranathunge, K.; El-Kereamy, A.; Gidda, S.; Bi, Y.M.; Rothstein, S.J. AMT1;1 transgenic rice plants with enhanced NH4+ permeability show superior growth and higher yield under optimal and suboptimal NH4+ conditions. J. Exp. Bot. 2014, 65, 965–979. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate. Trends. Plant Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Siddiqi, M.Y.; Glass, A.D.M. Kinetics of NH4+ influx in spruce. Plant Physiol. 1996, 110, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Rennenberg, H.; Dannenmann, M.; Gessler, A.; Kreuzwieser, J.; Simon, J.; Papen, H. Nitrogen balance in forest soils: Nutritional limitation of plants under climate change stresses. Plant Biol. 2009, 11, 4–23. [Google Scholar] [CrossRef] [PubMed]
- Koegel, S.; Lahmidi, N.A.; Arnould, C.; Chatagnier, O.; Walder, F.; Ineichen, K.; Boller, T.; Wipf, D.; Wiemken, A.; Courty, P. The family of ammonium transporters (AMT) in Sorghum bicolor: Two AMT members are induced locally, but not systemically in roots colonized by arbuscular mycorrhizal fungi. New Phytol. 2013, 198, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Ninnemann, O.; Jauniaux, J.C.; Frommer, W.B. Identification of a high affinity NH4+ transporter from plants. EMBO J. 1994, 13, 3464–3471. [Google Scholar] [CrossRef] [PubMed]
- Giehl, R.F.H.; Laginha, A.M.; Duan, F.; Rentsch, D.; Yuan, L.; von Wire’n, N. A critical role of AMT2;1 in root-to-shoot translocation of ammonium in Arabidopsis. Mol. Plant. 2017, 10, 1449–1460. [Google Scholar] [CrossRef]
- Li, C.; Tang, Z.; Wei, J.; Qu, H.Y.; Xie, Y.J.; Xu, G.H. The OsAMT1.1 gene functions in ammonium uptake and ammonium–potassium homeostasis over low and high ammonium concentration ranges. J. Genet. Genom. 2016, 43, 639–649. [Google Scholar] [CrossRef]
- Esmaeilzadeh-Salestani, K.; Samandari-Bahraseman, M.R.; Tohidfar, M.; Khaleghdoust, B.; Keres, I.; Mottus, A.; Loit, E. Expression of AMT1.1 and AMT2.1 is stimulated by mineral nitrogen and reproductive growth stage in barley under field conditions. J. Plant Nutr. 2022, 1–13. [Google Scholar] [CrossRef]
- Engineer, C.B.; Kranz, R.G. Reciprocal leaf and root expression of AtAMT1.1 and root architectural changes in response to nitrogen starvation. Plant Physiol. 2007, 143, 236–250. [Google Scholar] [CrossRef]
- Kaiser, B.N.; Rawat, S.R.; Siddiqi, M.Y.; Masle, J.; Glass, A.D.M. Functional analysis of an Arabidopsis T-DNA “knockout” of the high-affinity NH4+ transporter AtAMT1;1. Plant Physiol. 2002, 130, 1263–1275. [Google Scholar] [CrossRef]
- Kumar, A.; Silim, S.N.; Okamoto, M.; Siddiqi, M.Y.; Glass, A.D.M. Differential expression of three members of the AMT1 gene family encoding putative high-affinity NH4+ transporters in roots of Oryza sativa subspecies indica. Plant Cell Environ. 2003, 26, 907–914. [Google Scholar] [CrossRef]
- Kumar, A.; Kaiser, B.N.; Siddiqi, M.Y.; Glass, A.D.M. Functional characterization of OsAMT1.1 overexpression lines of rice. Funct. Plant Biol. 2006, 33, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Hoque, M.S.; Masle, J.; Udvardi, M.K.; Ryan, P.R.; Upadhyaya, N.M. Over-expression of the rice OsAMT1.1 gene increases ammonium uptake and content, but impairs growth and development of plants under high ammonium nutrition. Funct. Plant Biol. 2006, 33, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Y.; Gao, S.W.; Su, Y.C.; Lin, Z.L.; Guo, J.L.; Li, M.J.; Wang, Z.T.; Que, Y.X.; Xu, L.P. Transcripts and low nitrogen tolerance: Regulatory and metabolic pathways in sugarcane under low nitrogen stress. Environ. Exp. Bot. 2019, 163, 97–111. [Google Scholar] [CrossRef]
- Ludewig, U. Ion transport versus gas conduction: Function of AMT/Rh-type proteins. Transfus. Clin. Biol. 2006, 13, 111–116. [Google Scholar] [CrossRef]
- Marini, A.M.; Vissers, S.; Urrestarazu, A.; Andre, B. Cloning and expression of the MEP1 gene encoding an ammonium transporter in Saccharomyces cerevisiae. EMBO J. 1994, 13, 3456–3463. [Google Scholar] [CrossRef]
- Hao, D.L.; Zhou, J.Y.; Yang, S.Y.; Qi, W.; Yang, K.J.; Su, Y.H. Function and regulation of ammonium transporters in plants. Int. J. Mol. Sci. 2020, 21, 3557. [Google Scholar] [CrossRef] [PubMed]
- Daryanto, S.; Wang, L.X.; Gilhooly, W.P.; Jacinthe, P.A. Nitrogen preference across generations under changing ammonium nitrate ratios. J. Plant Ecol. 2019, 12, 235–244. [Google Scholar] [CrossRef]
- Lanquar, V.; Loqué, D.; Hörmann, F.; Yuan, L.X.; Bohner, A.; Engelsberger, W.R.; Lalonde, S.; Schulze, W.X.; von Wirén, N.; Frommer, W.B. Feedback inhibition of ammonium uptake by a phospho-dependent allosteric mechanism in Arabidopsis. Plant Cell. 2009, 21, 3610–3622. [Google Scholar] [CrossRef]
- Tabuchi, M.; Abiko, T.; Yamaya, T. Assimilation of ammonium ions and reutilization of nitrogen in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 2319–2327. [Google Scholar] [CrossRef]
- Ling, H.; Wu, Q.B.; Guo, J.L.; Xu, L.P.; Que, Y.X. Comprehensive selection of reference genes for gene expression normalization in sugarcane by real time quantitative RT-PCR. PLoS One 2014, 9, e97469. [Google Scholar] [CrossRef]
- Shin, S.Y.; Jeong, J.S.; Lim, J.Y.; Kim, T.; Park, J.H.; Kim, J.K.; Shin, C. Transcriptomic analyses of rice (Oryza sativa) genes and non-coding RNAs under nitrogen starvation using multiple omics technologies. BMC Genom. 2018, 19, 532. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.L.; Ding, G.D.; Li, L.; Cai, H.M.; Ye, X.S.; Zou, J.; Xu, F.S. Identification and characterization of improved nitrogen efficiency in interspecific hybridized new-type Brassica napus. Ann. Bot. 2014, 114, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Groat, R.G.; Vance, C.P. Root nodule enzymes of ammonia assimilation in alfalfa (Medicago sativa L.) developmental patterns and response to applied nitrogen. Plant Physiol. 1981, 67, 1198–1203. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, S.; Yang, Y.; Guo, J.; Zhang, X.; Feng, M.; Su, Y.; Que, Y.; Xu, L. Ectopic Expression of Sugarcane ScAMT1.1 Has the Potential to Improve Ammonium Assimilation and Grain Yield in Transgenic Rice under Low Nitrogen Stress. Int. J. Mol. Sci. 2023, 24, 1595. https://doi.org/10.3390/ijms24021595
Gao S, Yang Y, Guo J, Zhang X, Feng M, Su Y, Que Y, Xu L. Ectopic Expression of Sugarcane ScAMT1.1 Has the Potential to Improve Ammonium Assimilation and Grain Yield in Transgenic Rice under Low Nitrogen Stress. International Journal of Molecular Sciences. 2023; 24(2):1595. https://doi.org/10.3390/ijms24021595
Chicago/Turabian StyleGao, Shiwu, Yingying Yang, Jinlong Guo, Xu Zhang, Minxie Feng, Yachun Su, Youxiong Que, and Liping Xu. 2023. "Ectopic Expression of Sugarcane ScAMT1.1 Has the Potential to Improve Ammonium Assimilation and Grain Yield in Transgenic Rice under Low Nitrogen Stress" International Journal of Molecular Sciences 24, no. 2: 1595. https://doi.org/10.3390/ijms24021595
APA StyleGao, S., Yang, Y., Guo, J., Zhang, X., Feng, M., Su, Y., Que, Y., & Xu, L. (2023). Ectopic Expression of Sugarcane ScAMT1.1 Has the Potential to Improve Ammonium Assimilation and Grain Yield in Transgenic Rice under Low Nitrogen Stress. International Journal of Molecular Sciences, 24(2), 1595. https://doi.org/10.3390/ijms24021595