
The Phospholipase A2 Superfamily: Structure, Isozymes, Catalysis, Physiologic and Pathologic Roles



Abstract
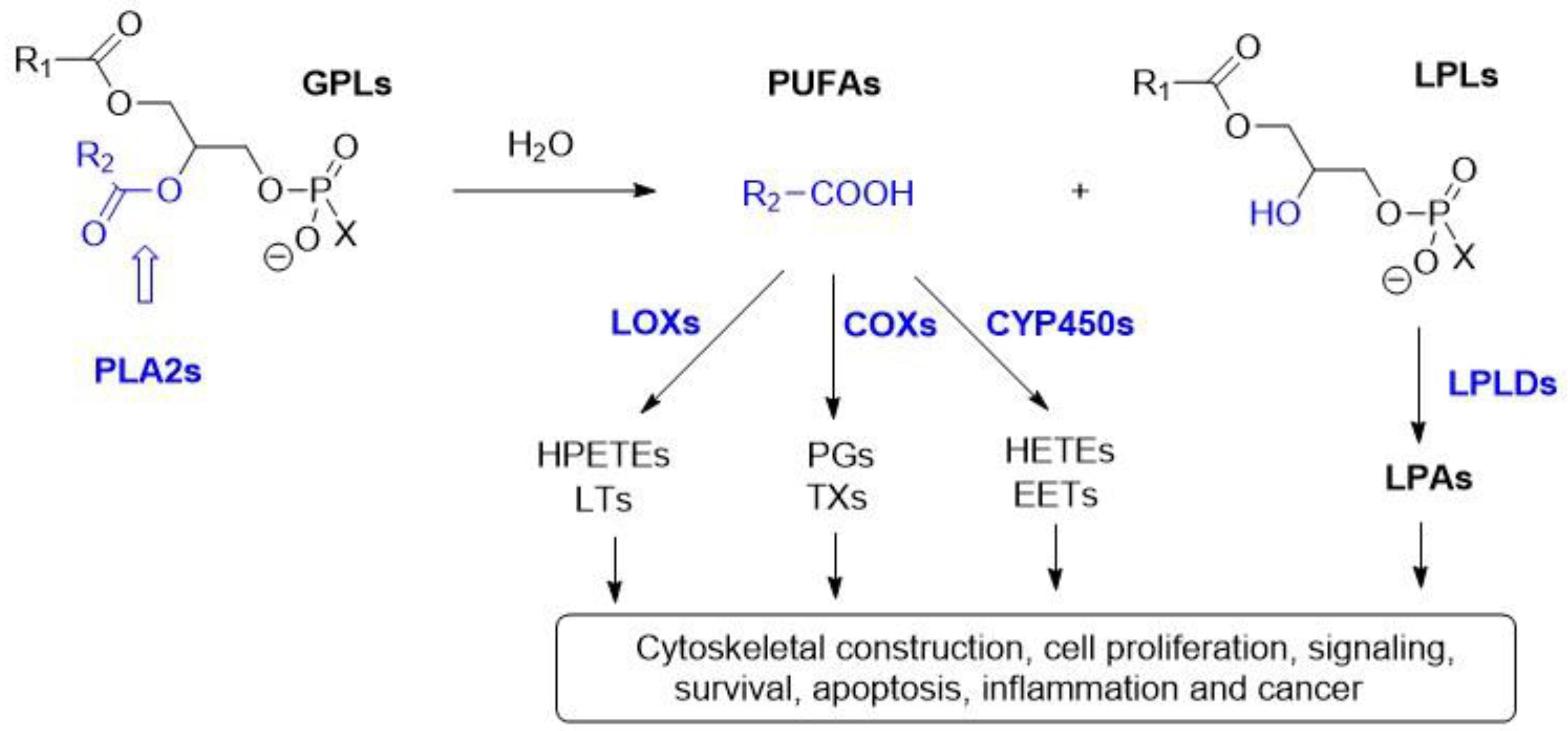
1. Introduction
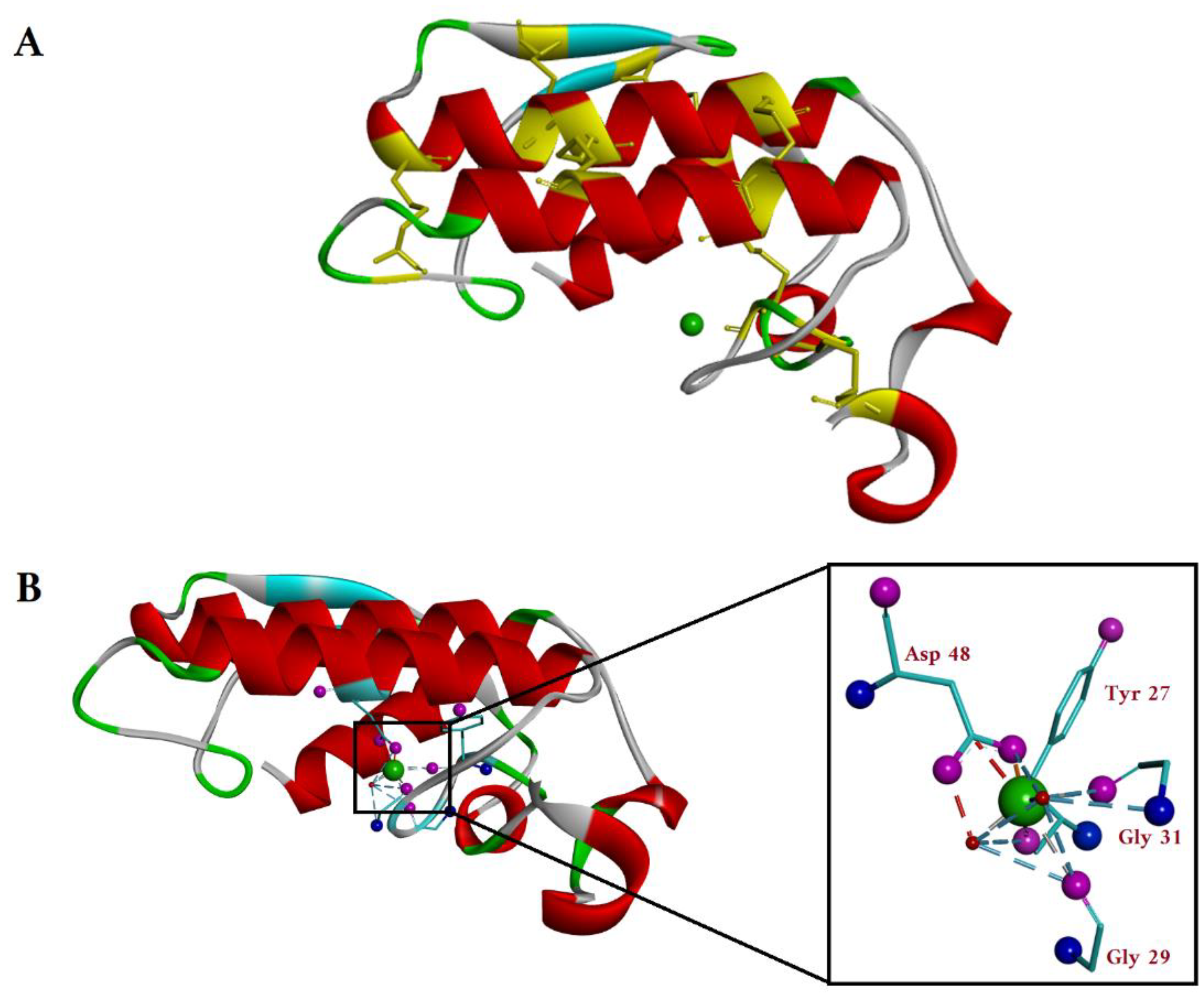
2. Secreted PLA2 (sPLA2)
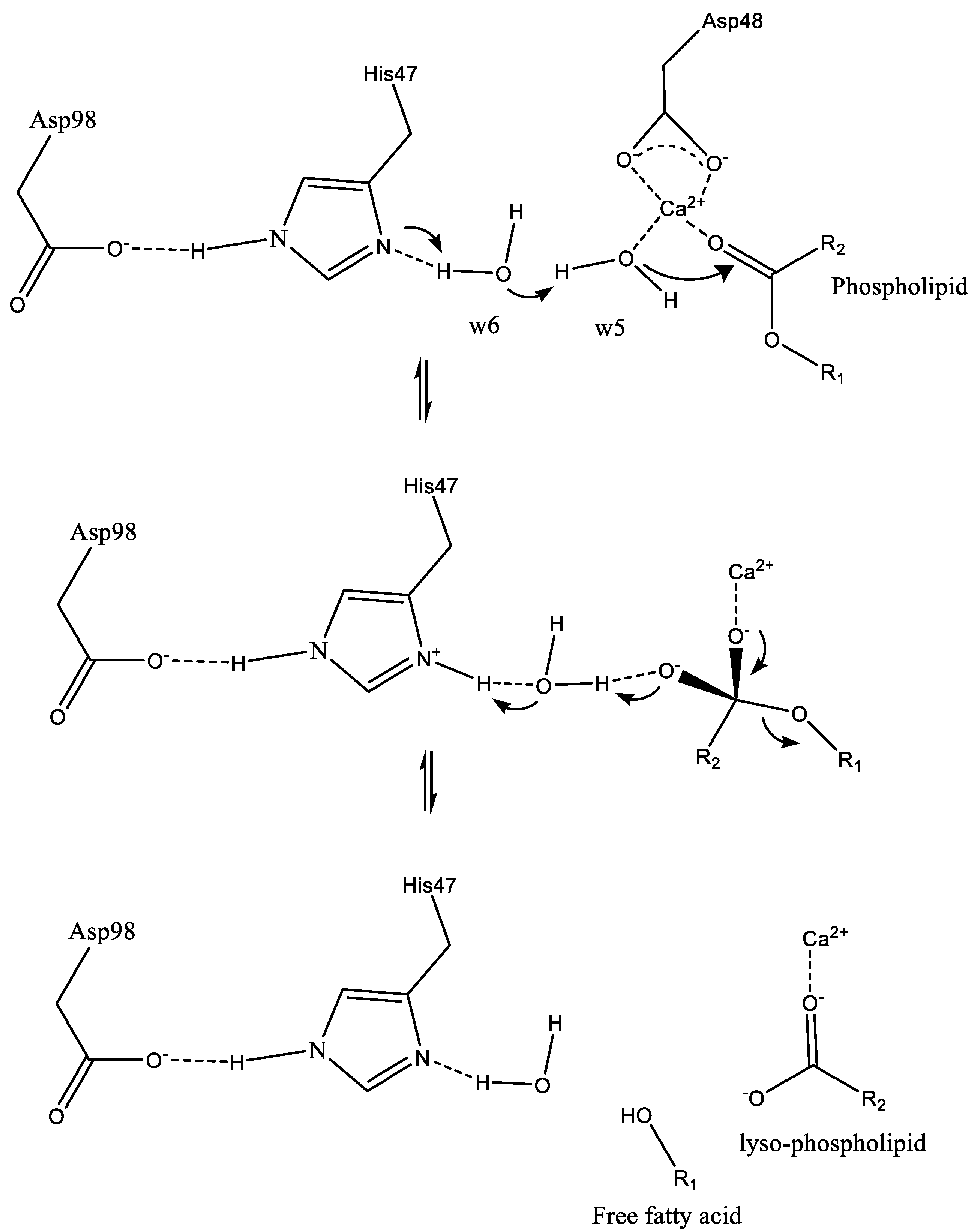
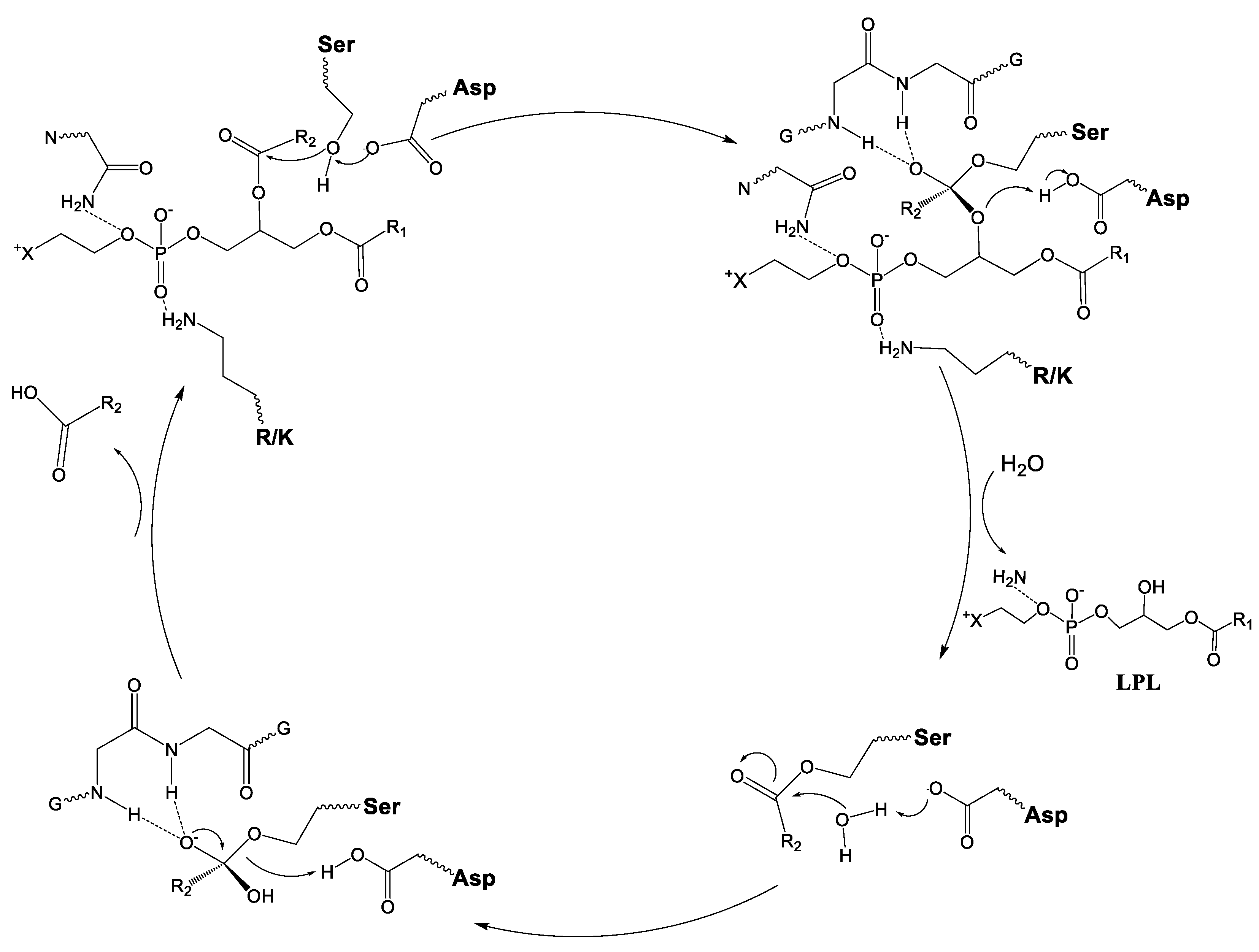
2.1. sPLA2 Catalytic Mechanism
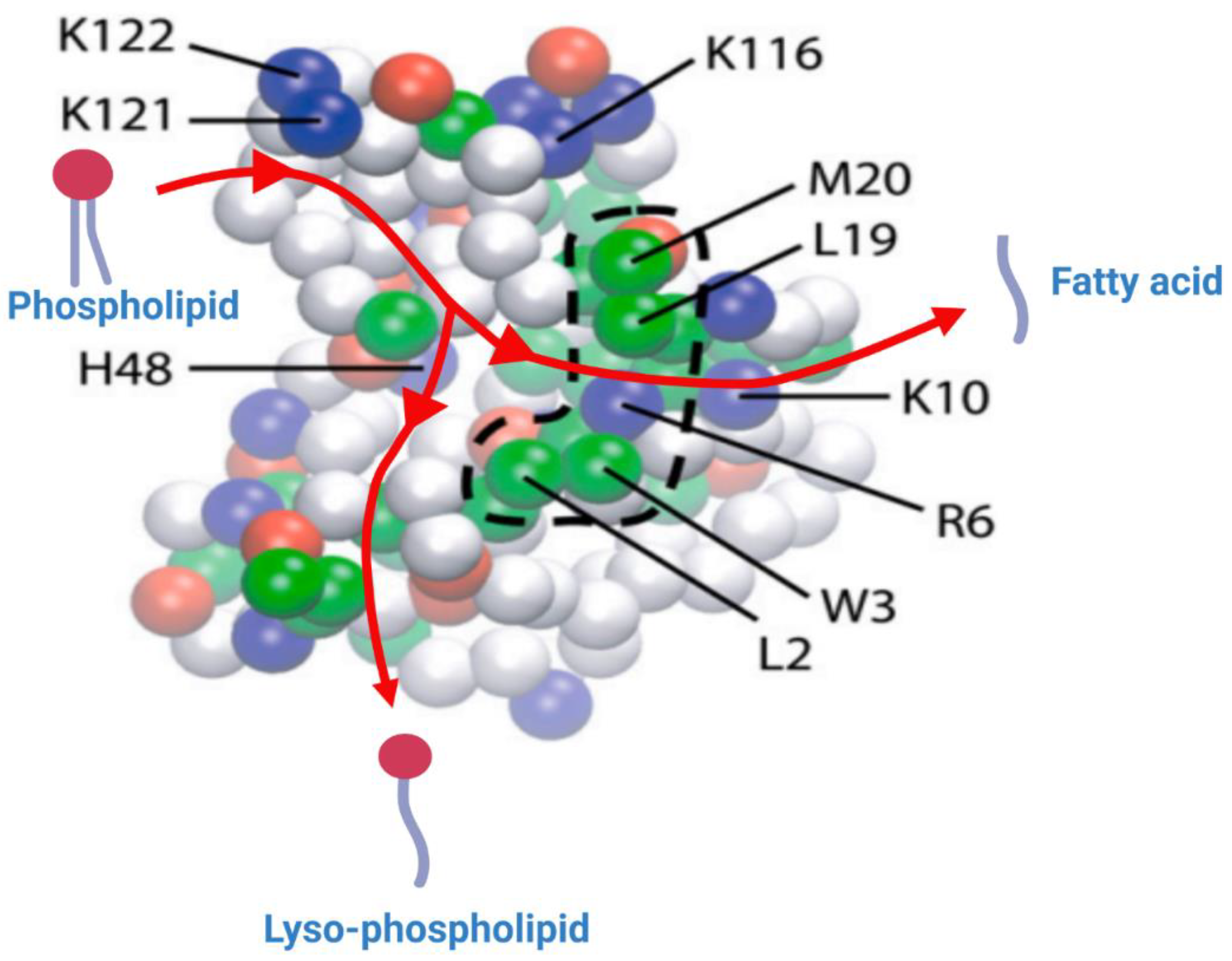
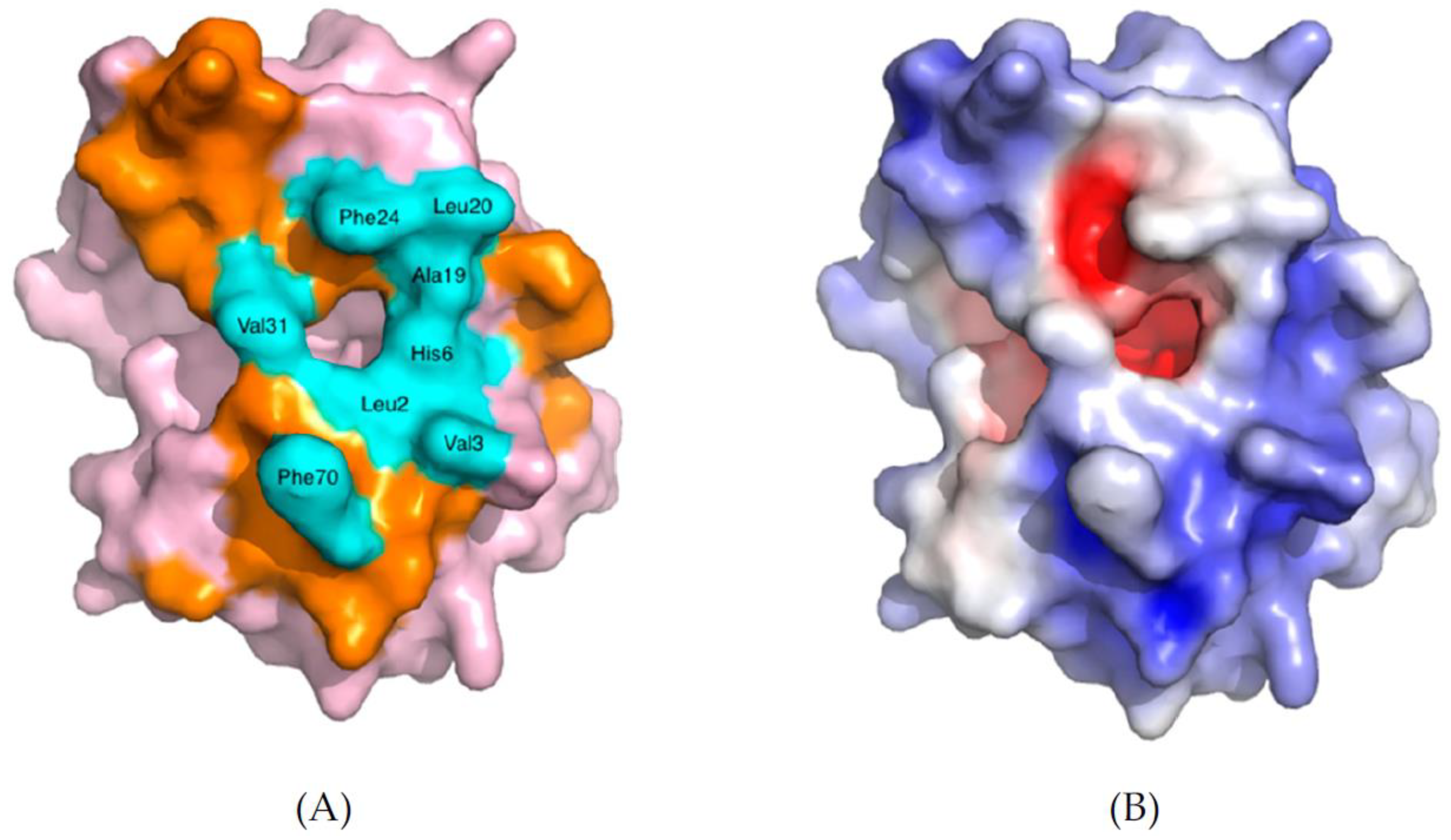
2.2. Interaction of sPLA2 with Phospholipid Substrate
2.3. Interaction of sPLA2 with Phospholipid Bilayers: Coarse-Grained Molecular Dynamic (CG-MD) and Atomistic Molecular Dynamic (AT-MD) Simulation Results
2.4. sPLA2 Substrate Binding and Interfacial Kinetics
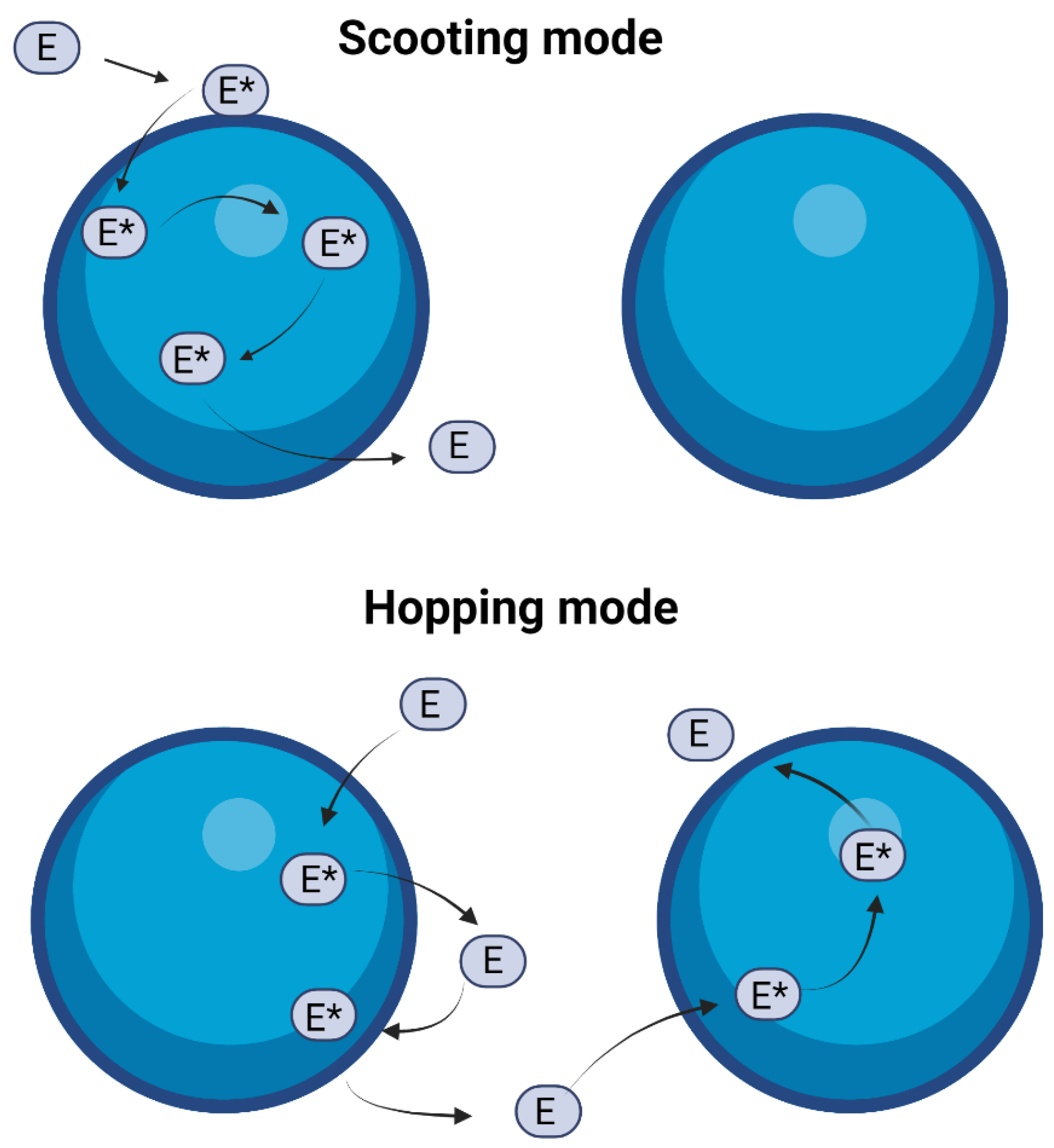
2.5. Scooting vs. Hopping Mode of Action of sPLA2
3. Cytosolic PLA2 (cPLA2)
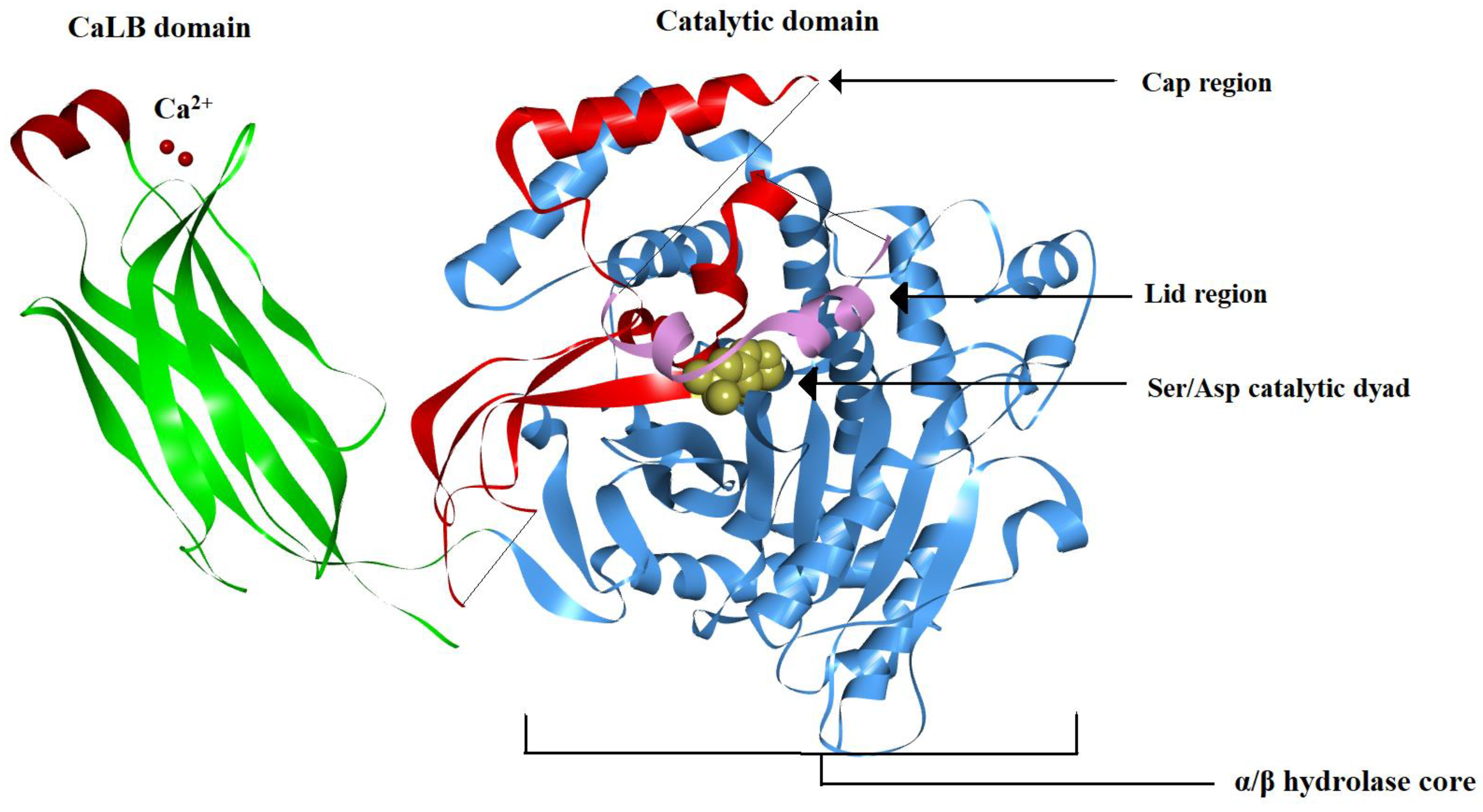
3.1. cPLA2 Crystal Structure
3.2. cPLA2 Activation Mechanism
4. Ca2+ Independent PLA2 (iPLA2)
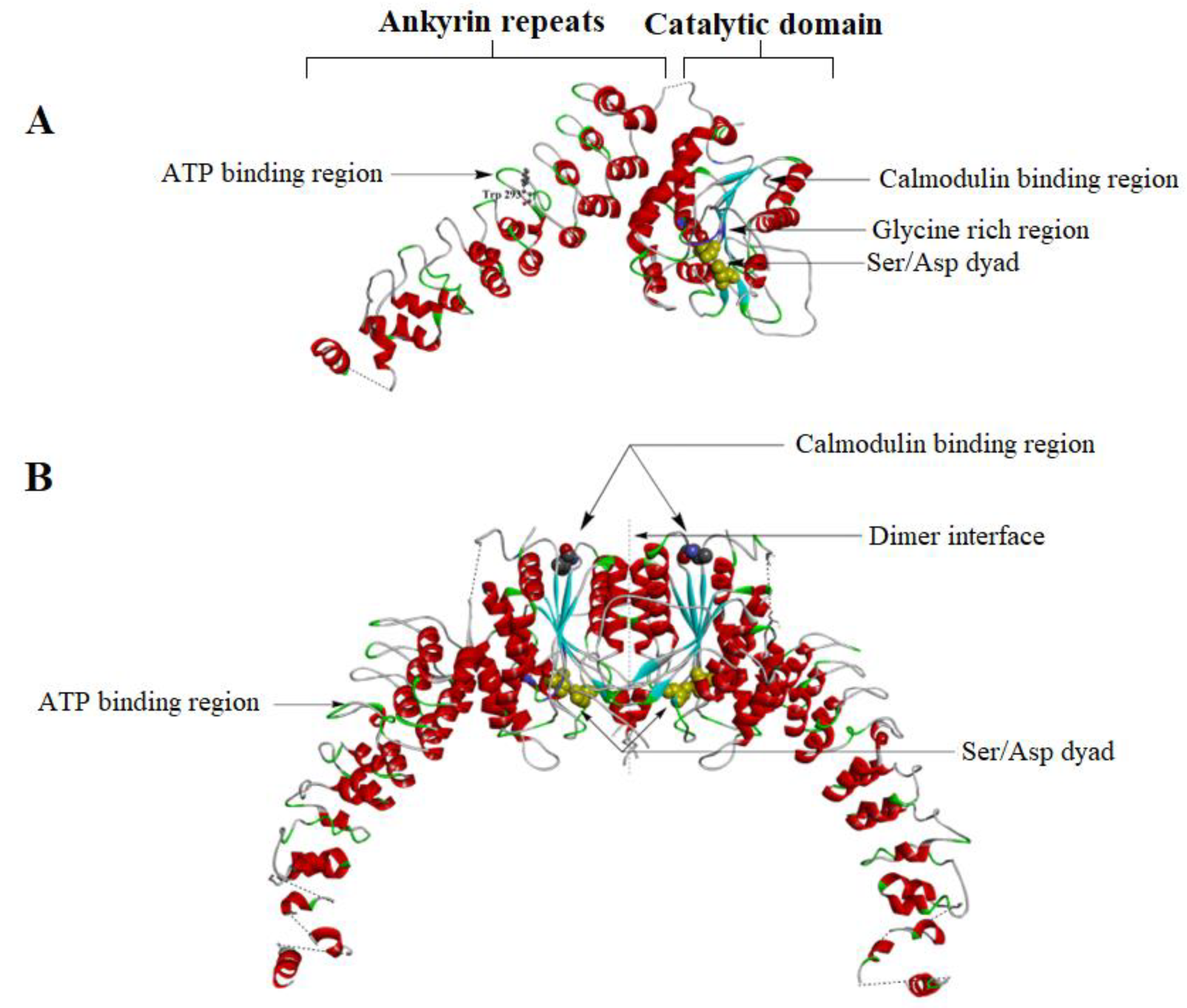
4.1. iPLA2 Crystal Structure
4.2. iPLA2 Regulatory Mechanisms
4.3. Mechanism for Catalysis for cPLA2 and iPLA2
5. Comparison of Membrane Surface Interaction within Four Major Classes of PLA2
6. Physiologic and Pathologic Role of PLA2s
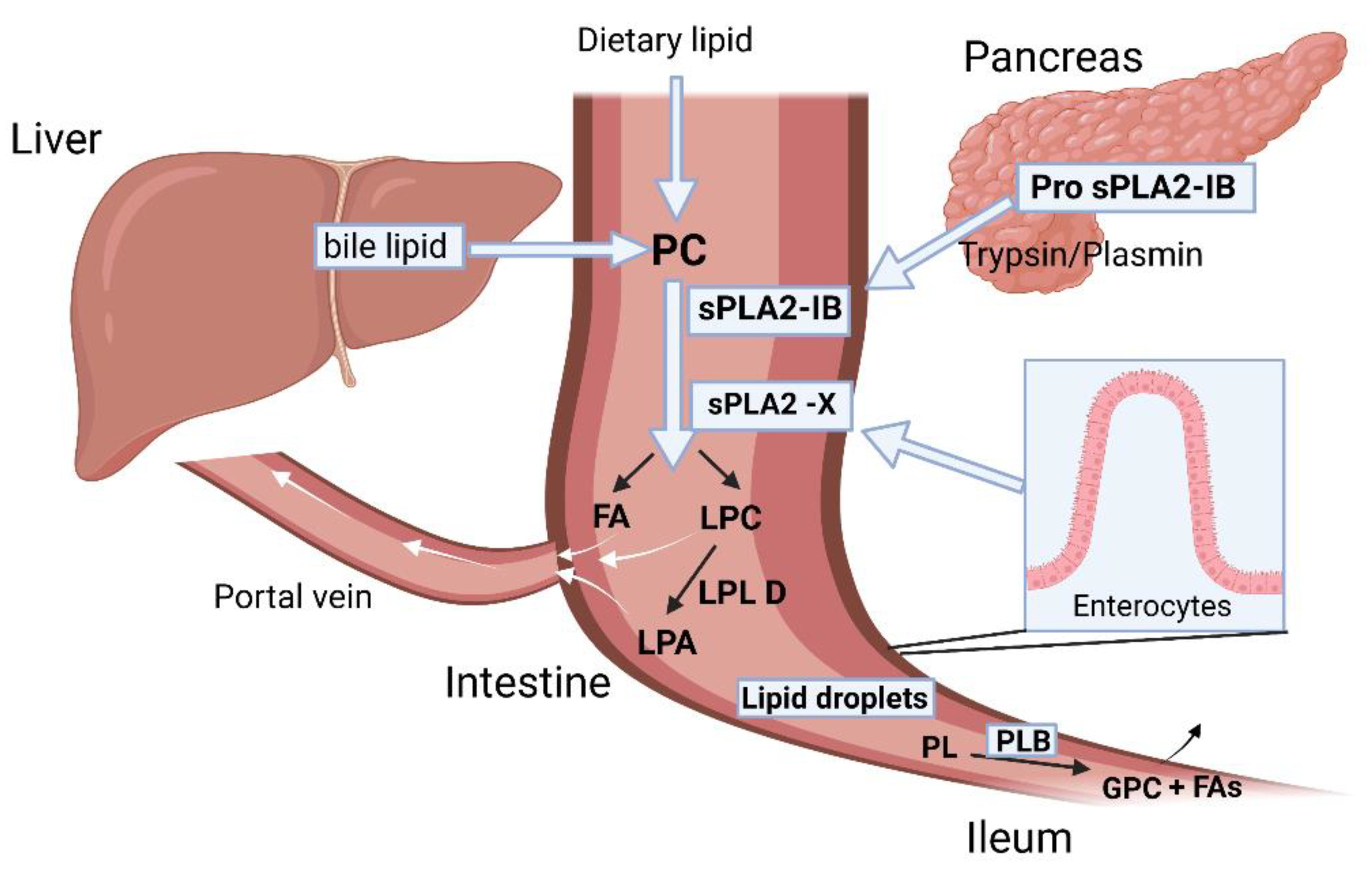
6.1. sPLA2 Role in Physiology and Pathology
6.2. Role of sPLA2 in Cancer
6.3. Physiologic and Pathologic Role of cPLA2
6.4. Role of cPLA2 in Cancer
6.5. Physiologic and Pathologic Role of iPLA2s
Role of iPLA2 in Myocardial Ischemia
6.6. Role of iPLA2 in Cancer
6.7. Physiologic and Pathologic role of Lipoprotein-Associated PLA2 (Lp-PLA2)
6.8. Physiologic and Pathologic Role of Lysosomal PLA2 (LPLA2)
6.9. Physiologic and Pathologic Role of Adipose Specific PLA2 (Ad-PLA2)
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murakami, M.; Kudo, I. Phospholipase A2. J. Biochem. 2002, 131, 285–292. [Google Scholar] [CrossRef]
- Cathcart, M.-C.; Lysaght, J.; Pidgeon, G.P. Eicosanoid Signalling Pathways in the Development and Progression of Colorectal Cancer: Novel Approaches for Prevention/Intervention. Cancer Metastasis Rev. 2011, 30, 363–385. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Chang, Y.; Fan, J.; Ji, W.; Su, C. Phospholipase A2 Superfamily in Cancer. Cancer Lett. 2021, 497, 165–177. [Google Scholar] [CrossRef]
- Moolenaar, W.H.; van Meeteren, L.A.; Giepmans, B.N.G. The Ins and Outs of Lysophosphatidic Acid Signaling. BioEssays 2004, 26, 870–881. [Google Scholar] [CrossRef]
- Lam, L.; Ilies, M.A. Evaluation of the Impact of Esterases and Lipases from the Circulatory System against Substrates of Different Lipophilicity. Int. J. Mol. Sci. 2022, 23, 1262. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A.; Cao, J.; Hsu, Y.-H.; Magrioti, V.; Kokotos, G. Phospholipase A Enzymes: Physical Structure, Biological Function, Disease Implication, Chemical Inhibition, and Therapeutic Intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [PubMed]
- Six, D.A.; Dennis, E.A. The Expanding Superfamily of Phospholipase A2 Enzymes: Classification and Characterization. Biochim. Et Biophys. Acta BBA-Mol. Cell Biol. Lipids 2000, 1488, 1–19. [Google Scholar] [CrossRef]
- Abbott, M.J.; Tang, T.; Sul, H.S. The Role of Phospholipase A2-Derived Mediators in Obesity. Drug Discov. Today Dis. Mech. 2010, 7, e213–e218. [Google Scholar] [CrossRef]
- Burke, J.E.; Dennis, E.A. Phospholipase A2 Structure/Function, Mechanism, and Signaling. J. Lipid Res. 2009, 50, S237–S242. [Google Scholar] [CrossRef]
- Heinrikson, R.L.; Krueger, E.T.; Keim, P.S. Amino Acid Sequence of Phospholipase A2-Alpha from the Venom of Crotalus Adamanteus. A New Classification of Phospholipases A2 Based upon Structural Determinants. J. Biol. Chem. 1977, 252, 4913–4921. [Google Scholar] [CrossRef]
- Murakami, M.; Sato, H.; Miki, Y.; Yamamoto, K.; Taketomi, Y. A New Era of Secreted Phospholipase A2. J. Lipid Res. 2015, 56, 1248–1261. [Google Scholar] [CrossRef]
- Masuda, S.; Yamamoto, K.; Hirabayashi, T.; Ishikawa, Y.; Ishii, T.; Kudo, I.; Murakami, M. Human Group III Secreted Phospholipase A2 Promotes Neuronal Outgrowth and Survival. Biochem. J. 2008, 409, 429–438. [Google Scholar] [CrossRef]
- Murakami, M.; Miki, Y.; Sato, H.; Murase, R.; Taketomi, Y.; Yamamoto, K. Group IID, IIE, IIF and III Secreted Phospholipase A2s. Biochim. Et Biophys. Acta BBA-Mol. Cell Biol. Lipids 2019, 1864, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Balsinde, J.; Winstead, M.V.; Dennis, E.A. Phospholipase A(2) Regulation of Arachidonic Acid Mobilization. FEBS Lett. 2002, 531, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Turk, J. The Molecular Biology of the Group VIA Ca2+-Independent Phospholipase A2. In Progress in Nucleic Acid Research and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2001; Volume 67, pp. 1–33. ISBN 978-0-12-540067-1. [Google Scholar]
- Talmud, P.J.; Holmes, M.V. Deciphering the Causal Role of SPLA2s and Lp-PLA2 in Coronary Heart Disease. Arter. Thromb Vasc Biol. 2015, 35, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Shayman, J.A.; Kelly, R.; Kollmeyer, J.; He, Y.; Abe, A. Group XV Phospholipase A2, a Lysosomal Phospholipase A2. Prog. Lipid Res. 2011, 50, 1–13. [Google Scholar] [CrossRef]
- Pang, X.-Y.; Cao, J.; Addington, L.; Lovell, S.; Battaile, K.P.; Zhang, N.; Rao, J.L.U.M.; Dennis, E.A.; Moise, A.R. Structure/Function Relationships of Adipose Phospholipase A2 Containing a Cys-His-His Catalytic Triad. J. Biol. Chem. 2012, 287, 35260–35274. [Google Scholar] [CrossRef]
- Scott, D.L.; White, S.P.; Otwinowski, Z.; Yuan, W.; Gelb, M.H.; Sigler, P.B. Interfacial Catalysis: The Mechanism of Phospholipase A2. Science 1990, 250, 1541–1546. [Google Scholar] [CrossRef]
- Berg, O.G.; Gelb, M.H.; Tsai, M.-D.; Jain, M.K. Interfacial Enzymology: The Secreted Phospholipase A -Paradigm. Chem. Rev. 2001, 101, 2613–2654. [Google Scholar] [CrossRef]
- Dennis, E.A. Diversity of Group Types, Regulation, and Function of Phospholipase A2. J. Biol. Chem. 1994, 269, 13057–13060. [Google Scholar] [CrossRef]
- Wee, C.L.; Balali-Mood, K.; Gavaghan, D.; Sansom, M.S.P. The Interaction of Phospholipase A2 with a Phospholipid Bilayer: Coarse-Grained Molecular Dynamics Simulations. Biophys. J. 2008, 95, 1649–1657. [Google Scholar] [CrossRef] [PubMed]
- Lambeau, G.; Gelb, M.H. Biochemistry and Physiology of Mammalian Secreted Phospholipases A2. Annu. Rev. Biochem. 2008, 77, 495–520. [Google Scholar] [CrossRef] [PubMed]
- Winget, J.M.; Pan, Y.H.; Bahnson, B.J. The Interfacial Binding Surface of Phospholipase A2s. Biochim. Et Biophys. Acta BBA-Mol. Cell Biol. Lipids 2006, 1761, 1260–1269. [Google Scholar] [CrossRef]
- Jain, M.K.; Berg, O.G. Coupling of the I-Face and the Active Site of Phospholipase A2 for Interfacial Activation. Curr. Opin. Chem. Biol. 2006, 10, 473–479. [Google Scholar] [CrossRef]
- Kim, R.R.; Chen, Z.; Mann, T.J.; Bastard, K.; Scott, K.F.; Church, W.B. Structural and Functional Aspects of Targeting the Secreted Human Group IIA Phospholipase A2. Molecules 2020, 25, 4459. [Google Scholar] [CrossRef]
- Ramirez, F.; Jain, M.K. Phospholipase A2 at the Bilayer Interface. Proteins Struct. Funct. Bioinform. 1991, 9, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Tucker, D.E.; Burchett, S.A.; Leslie, C.C. Properties of the Group IV Phospholipase A2 Family. Prog. Lipid Res. 2006, 45, 487–510. [Google Scholar] [CrossRef]
- Kramer, R.M.; Checani, G.C.; Deykin, A.; Pritzker, C.R.; Deykin, D. Solubilization and Properties of Ca2+-Dependent Human Platelet Phospholipase A2. Biochim. Et Biophys. Acta BBA-Lipids Lipid Metab. 1986, 878, 394–403. [Google Scholar] [CrossRef]
- Alonso, F.; Henson, P.M.; Leslie, C.C. A Cytosolic Phospholipase in Human Neutrophils That Hydrolyzes Arachidonoyl-Containing Phosphatidylcholine. Biochim. Et Biophys. Acta BBA-Lipids Lipid Metab. 1986, 878, 273–280. [Google Scholar] [CrossRef]
- Dessen, A.; Tang, J.; Schmidt, H.; Stahl, M.; Clark, J.D.; Seehra, J.; Somers, W.S. Crystal Structure of Human Cytosolic Phospholipase A2 Reveals a Novel Topology and Catalytic Mechanism. Cell 1999, 97, 349–360. [Google Scholar] [CrossRef]
- Burke, J.E.; Hsu, Y.-H.; Deems, R.A.; Li, S.; Woods, V.L.; Dennis, E.A. A Phospholipid Substrate Molecule Residing in the Membrane Surface Mediates Opening of the Lid Region in Group IVA Cytosolic Phospholipase A2. J. Biol. Chem. 2008, 283, 31227–31236. [Google Scholar] [CrossRef] [PubMed]
- Dessen, A.; Somers, W.S.; Stahl, M.L.; Seehra, J.S. Crystal Structure of CPLA2 and Methods of Identifying Agonists and Antagonists Using Same. U.S. Patent US 6801860, 5 October 2004. [Google Scholar]
- Hsu, Y.-H.; Burke, J.E.; Stephens, D.L.; Deems, R.A.; Li, S.; Asmus, K.M.; Woods, V.L.; Dennis, E.A. Calcium Binding Rigidifies the C2 Domain and the Intradomain Interaction of GIVA Phospholipase A2 as Revealed by Hydrogen/Deuterium Exchange Mass Spectrometry. J. Biol. Chem. 2008, 283, 9820–9827. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Rafter, J.D.; Kim, K.P.; Gygi, S.P.; Cho, W. Mechanism of Group IVA Cytosolic Phospholipase A2 Activation by Phosphorylation. J. Biol. Chem. 2003, 278, 41431–41442. [Google Scholar] [CrossRef] [PubMed]
- Casas, J.; Balsinde, J.; Balboa, M.A. Phosphorylation of CPLA2α at Ser505 Is Necessary for Its Translocation to PtdInsP2-Enriched Membranes. Molecules 2022, 27, 2347. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-L.; Wartmann, M.; Lin, A.Y.; Knopf, J.L.; Seth, A.; Davis, R.J. CPLA2 Is Phosphorylated and Activated by MAP Kinase. Cell 1993, 72, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Das, S.; Murthy, K.S. Erk1/2- and P38 MAP Kinase-Dependent Phosphorylation and Activation of CPLA by M3 and M2 Receptors. Am. J. Physiol.-Gastrointest. Liver Physiol. 2003, 284, G472–G480. [Google Scholar] [CrossRef]
- Nahas, N.; Waterman, W.H.; Sha’Afi, R.I. Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF) Promotes Phosphorylation and an Increase in the Activity of Cytosolic Phospholipase A2 in Human Neutrophils. Biochem. J. 1996, 313, 503–508. [Google Scholar] [CrossRef]
- Palavicini, J.P.; Wang, C.; Chen, L.; Hosang, K.; Wang, J.; Tomiyama, T.; Mori, H.; Han, X. Oligomeric Amyloid-Beta Induces MAPK-Mediated Activation of Brain Cytosolic and Calcium-Independent Phospholipase A2 in a Spatial-Specific Manner. Acta Neuropathol. Commun. 2017, 5, 56. [Google Scholar] [CrossRef]
- Xu, J.; Weng, Y.-I.; Simonyi, A.; Krugh, B.W.; Liao, Z.; Weisman, G.A.; Sun, G.Y.; Simoni, A. Role of PKC and MAPK in Cytosolic PLA2 Phosphorylation and Arachadonic Acid Release in Primary Murine Astrocytes. J. Neurochem. 2002, 83, 259–270. [Google Scholar] [CrossRef]
- Xing, M.; Firestein, B.L.; Shen, G.H.; Insel, P.A. Dual Role of Protein Kinase C in the Regulation of CPLA2-Mediated Arachidonic Acid Release by P2U Receptors in MDCK-D1 Cells: Involvement of MAP Kinase-Dependent and -Independent Pathways. J. Clin. Investig. 1997, 99, 805–814. [Google Scholar] [CrossRef]
- Hoeferlin, L.A.; Wijesinghe, D.S.; Chalfant, C.E. The Role of Ceramide-1-Phosphate in Biological Functions. In Handbook of Experimental Pharmacology; Springer: Vienna, Austria, 2013; pp. 153–166. [Google Scholar] [CrossRef]
- Pettus, B.J.; Bielawska, A.; Subramanian, P.; Wijesinghe, D.S.; Maceyka, M.; Leslie, C.C.; Evans, J.H.; Freiberg, J.; Roddy, P.; Hannun, Y.A.; et al. Ceramide 1-Phosphate Is a Direct Activator of Cytosolic Phospholipase A2. J. Biol. Chem. 2004, 279, 11320–11326. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Hirabayashi, T.; Shimizu, M.; Murayama, T. Ceramide-1-Phosphate Activates Cytosolic Phospholipase A2alpha Directly and by PKC Pathway. Biochem. Pharm. 2006, 71, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Mencarelli, C.; Martinez–Martinez, P. Ceramide Function in the Brain: When a Slight Tilt Is Enough. Cell. Mol. Life Sci. 2013, 70, 181–203. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. The Inositol Trisphosphate/Calcium Signaling Pathway in Health and Disease. Physiol. Rev. 2016, 96, 1261–1296. [Google Scholar] [CrossRef]
- Katan, M.; Cockcroft, S. Phosphatidylinositol(4,5)Bisphosphate: Diverse Functions at the Plasma Membrane. Essays Biochem. 2020, 64, 513–531. [Google Scholar] [CrossRef]
- Das, S.; Cho, W. Roles of Catalytic Domain Residues in Interfacial Binding and Activation of Group IV Cytosolic Phospholipase A2*. J. Biol. Chem. 2002, 277, 23838–23846. [Google Scholar] [CrossRef]
- Mosior, M.; Six, D.A.; Dennis, E.A. Group IV Cytosolic Phospholipase A2 Binds with High Affinity and Specificity to Phosphatidylinositol 4,5-Bisphosphate Resulting in Dramatic Increases in Activity. J. Biol. Chem. 1998, 273, 2184–2191. [Google Scholar] [CrossRef]
- Casas, J.; Gijón, M.A.; Vigo, A.G.; Crespo, M.S.; Balsinde, J.; Balboa, M.A. Phosphatidylinositol 4,5-Bisphosphate Anchors Cytosolic Group IVA Phospholipase A2 to Perinuclear Membranes and Decreases Its Calcium Requirement for Translocation in Live Cells. MBoC 2006, 17, 155–162. [Google Scholar] [CrossRef]
- Ackermann, E.J.; Kempner, E.S.; Dennis, E.A. Ca(2+)-Independent Cytosolic Phospholipase A2 from Macrophage-like P388D1 Cells. Isolation and Characterization. J. Biol. Chem. 1994, 269, 9227–9233. [Google Scholar] [CrossRef]
- Ramanadham, S.; Ali, T.; Ashley, J.W.; Bone, R.N.; Hancock, W.D.; Lei, X. Calcium-Independent Phospholipases A2 and Their Roles in Biological Processes and Diseases. J. Lipid Res. 2015, 56, 1643–1668. [Google Scholar] [CrossRef]
- Malley, K.R.; Koroleva, O.; Miller, I.; Sanishvili, R.; Jenkins, C.M.; Gross, R.W.; Korolev, S. The Structure of IPLA2β Reveals Dimeric Active Sites and Suggests Mechanisms of Regulation and Localization. Nat. Commun. 2018, 9, 765. [Google Scholar] [CrossRef]
- Hsu, Y.-H.; Burke, J.E.; Li, S.; Woods, V.L.; Dennis, E.A. Localizing the Membrane Binding Region of Group VIA Ca2+-Independent Phospholipase A2 Using Peptide Amide Hydrogen/Deuterium Exchange Mass Spectrometry. J. Biol. Chem. 2009, 284, 23652–23661. [Google Scholar] [CrossRef] [PubMed]
- Balsinde, J.; Pérez, R.; Balboa, M.A. Calcium-Independent Phospholipase A2 and Apoptosis. Biochim. Biophys. Acta 2006, 1761, 1344–1350. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.M.; Wolf, M.J.; Mancuso, D.J.; Gross, R.W. Identification of the Calmodulin-Binding Domain of Recombinant Calcium-Independent Phospholipase A2β. J. Biol. Chem. 2001, 276, 7129–7135. [Google Scholar] [CrossRef]
- Mouchlis, V.D.; Bucher, D.; McCammon, J.A.; Dennis, E.A. Membranes Serve as Allosteric Activators of Phospholipase A2, Enabling It to Extract, Bind, and Hydrolyze Phospholipid Substrates. Proc. Natl. Acad. Sci. USA 2015, 112, E516–E525. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Burke, J.E.; Dennis, E.A. Using Hydrogen/Deuterium Exchange Mass Spectrometry to Define the Specific Interactions of the Phospholipase A2 Superfamily with Lipid Substrates, Inhibitors, and Membranes. J. Biol. Chem. 2013, 288, 1806–1813. [Google Scholar] [CrossRef]
- Burke, J.E.; Karbarz, M.J.; Deems, R.A.; Li, S.; Woods, V.L.; Dennis, E.A. Interaction of Group IA Phospholipase A2 with Metal Ions and Phospholipid Vesicles Probed with Deuterium Exchange Mass Spectrometry. Biochemistry 2008, 47, 6451–6459. [Google Scholar] [CrossRef]
- Cao, J.; Hsu, Y.-H.; Li, S.; Woods, V.L.; Dennis, E.A. Lipoprotein-Associated Phospholipase A2 Interacts with Phospholipid Vesicles Via a Surface-Disposed Hydrophobic α-Helix. Biochemistry 2011, 50, 5314–5321. [Google Scholar] [CrossRef]
- Murakami, M.; Sato, H.; Taketomi, Y. Updating Phospholipase A2 Biology. Biomolecules 2020, 10, 1457. [Google Scholar] [CrossRef]
- Murakami, M.; Taketomi, Y.; Girard, C.; Yamamoto, K.; Lambeau, G. Emerging Roles of Secreted Phospholipase A2 Enzymes: Lessons from Transgenic and Knockout Mice. Biochimie 2010, 92, 561–582. [Google Scholar] [CrossRef]
- Hui, D.Y. Intestinal Phospholipid and Lysophospholipid Metabolism in Cardiometabolic Disease. Curr. Opin. Lipidol. 2016, 27, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.-W.; Qu, J.; Black, D.D.; Tso, P. Regulation of Intestinal Lipid Metabolism: Current Concepts and Relevance to Disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.Y. Group 1B Phospholipase A2 in Metabolic and Inflammatory Disease Modulation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 784–788. [Google Scholar] [CrossRef] [PubMed]
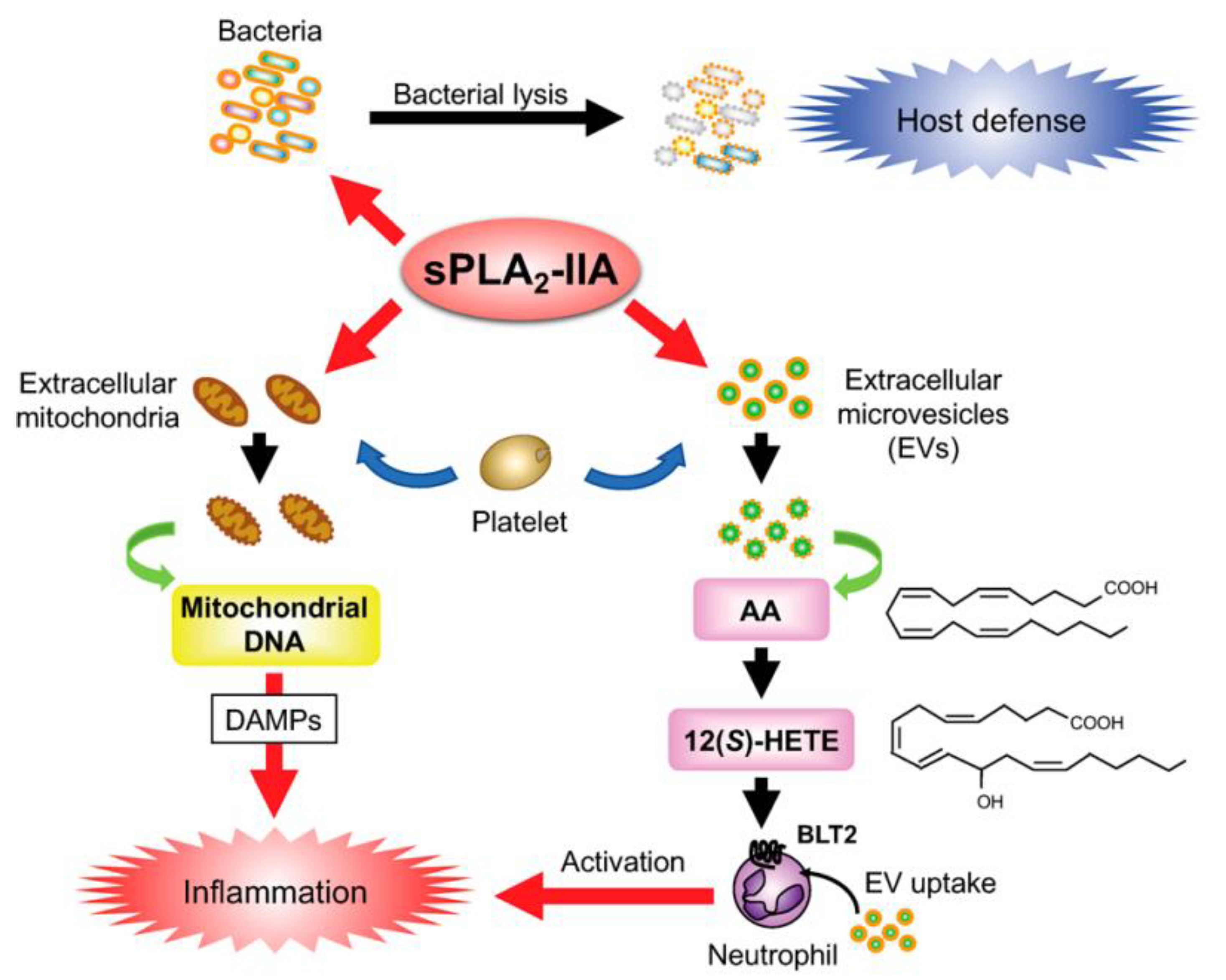
- Dore, E.; Boilard, E. Roles of Secreted Phospholipase A2 Group IIA in Inflammation and Host Defense. Biochim. Et Biophys. Acta BBA-Mol. Cell Biol. Lipids 2019, 1864, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Nevalainen, T.; Peuravuori, H. Secretion of Group 2 Phospholipase A2 by Lacrimal Glands. Investig. Ophthalmol. Vis. Sci. 1994, 35, 417–421. [Google Scholar]
- Grönroos, J.O.; Laine, V.J.O.; Nevalainen, T.J. Bactericidal Group IIA Phospholipase A2 in Serum of Patients with Bacterial Infections. J. Infect. Dis. 2002, 185, 1767–1772. [Google Scholar] [CrossRef]
- Murakami, M.; Kambe, T.; Shimbara, S.; Yamamoto, S.; Kuwata, H.; Kudo, I. Functional Association of Type IIA Secretory Phospholipase A2 with the Glycosylphosphatidylinositol-Anchored Heparan Sulfate Proteoglycan in the Cyclooxygenase-2-Mediated Delayed Prostanoid-Biosynthetic Pathway. J. Biol. Chem. 1999, 274, 29927–29936. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Koduri, R.S.; Enomoto, A.; Shimbara, S.; Seki, M.; Yoshihara, K.; Singer, A.; Valentin, E.; Ghomashchi, F.; Lambeau, G.; et al. Distinct Arachidonate-Releasing Functions of Mammalian Secreted Phospholipase A2s in Human Embryonic Kidney 293 and Rat Mastocytoma RBL-2H3 Cells through Heparan Sulfate Shuttling and External Plasma Membrane Mechanisms. J. Biol. Chem. 2001, 276, 10083–10096. [Google Scholar] [CrossRef]
- Mattsson, N.; Magnussen, C.G.; Rönnemaa, T.; Mallat, Z.; Benessiano, J.; Jula, A.; Taittonen, L.; Kähönen, M.; Juonala, M.; Viikari, J.S.A.; et al. Metabolic Syndrome and Carotid Intima-Media Thickness in Young Adults: Roles of Apolipoprotein B, Apolipoprotein A-I, C-Reactive Protein, and Secretory Phospholipase A2: The Cardiovascular Risk in Young Finns Study. ATVB 2010, 30, 1861–1866. [Google Scholar] [CrossRef]
- Ivandic, B.; Castellani, L.W.; Wang, X.-P.; Qiao, J.-H.; Mehrabian, M.; Navab, M.; Fogelman, A.M.; Grass, D.S.; Swanson, M.E.; de Beer, M.C.; et al. Role of Group II Secretory Phospholipase A2 in Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 1284–1290. [Google Scholar] [CrossRef]
- Avoranta, T.; Sundström, J.; Korkeila, E.; Syrjänen, K.; Pyrhönen, S.; Laine, J. The Expression and Distribution of Group IIA Phospholipase A2 in Human Colorectal Tumours. Virchows Arch. 2010, 457, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.-I.; Yamashita, J.-I.; Ogawa, M. Overexpression of Group II Phospholipase A2 in Human Breast Cancer Tissues Is Closely Associated with Their Malignant Potency. Br. J. Cancer 1994, 69, 1166–1170. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Zhao, X.; Wang, J.; Liu, C.; Sun, Y.; Cai, H.; Liu, J. Plasma Phospholipase A2 Activity May Serve as a Novel Diagnostic Biomarker for the Diagnosis of Breast Cancer. Oncol. Lett. 2018, 15, 5236–5242. [Google Scholar] [CrossRef]
- Yu, J.A.; Li, H.; Meng, X.; Fullerton, D.A.; Nemenoff, R.A.; Mitchell, J.D.; Weyant, M.J. Group IIa Secretory Phospholipase Expression Correlates with Group IIa Secretory Phospholipase Inhibition–Mediated Cell Death in K-Ras Mutant Lung Cancer Cells. J. Thorac. Cardiovasc. Surg. 2012, 144. [Google Scholar] [CrossRef]
- Cai, Q.; Zhao, Z.; Antalis, C.; Yan, L.; Del Priore, G.; Hamed, A.H.; Stehman, F.B.; Schilder, J.M.; Xu, Y. Elevated and Secreted Phospholipase A2 Activities as New Potential Therapeutic Targets in Human Epithelial Ovarian Cancer. FASEB J. 2012, 26, 3306–3320. [Google Scholar] [CrossRef] [PubMed]
- Gorovetz, M.; Baekelandt, M.; Berner, A.; Trope’, C.G.; Davidson, B.; Reich, R. The Clinical Role of Phospholipase A2 Isoforms in Advanced-Stage Ovarian Carcinoma. Gynecol. Oncol. 2006, 103, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Graff, J.R.; Konicek, B.W.; Deddens, J.A.; Chedid, M.; Hurst, B.M.; Colligan, B.; Neubauer, B.L.; Carter, H.W.; Carter, J.H. Expression of Group IIa Secretory Phospholipase A2 Increases with Prostate Tumor Grade. Clin. Cancer Res. 2001, 7, 3857–3861. [Google Scholar] [PubMed]
- Xing, X.-F.; Li, H.; Zhong, X.-Y.; Zhang, L.-H.; Wang, X.-H.; Liu, Y.-Q.; Jia, S.-Q.; Shi, T.; Niu, Z.-J.; Peng, Y.; et al. Phospholipase A2 Group IIA Expression Correlates with Prolonged Survival in Gastric Cancer. Histopathology 2011, 59, 198–206. [Google Scholar] [CrossRef]
- Brglez, V.; Lambeau, G.; Petan, T. Secreted Phospholipases A2 in Cancer: Diverse Mechanisms of Action. Biochimie 2014, 107, 114–123. [Google Scholar] [CrossRef]
- Mannello, F.; Qin, W.; Zhu, W.; Fabbri, L.; Tonti, G.A.; Sauter, E.R. Nipple Aspirate Fluids from Women with Breast Cancer Contain Increased Levels of Group IIa Secretory Phospholipase A2. Breast Cancer Res. Treat. 2008, 111, 209–218. [Google Scholar] [CrossRef]
- Yamashita, S.; Ogawa, M.; Sakamoto, K.; Abe, T.; Arakawa, H.; Yamashita, J. Elevation of Serum Group II Phospholipase A2 Levels in Patients with Advanced Cancer. Clin. Chim. Acta 1994, 228, 91–99. [Google Scholar] [CrossRef]
- Brglez, V.; Pucer, A.; Pungerčar, J.; Lambeau, G.; Petan, T. Secreted Phospholipases A2 Are Differentially Expressed and Epigenetically Silenced in Human Breast Cancer Cells. Biochem. Biophys. Res. Commun. 2014, 445, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Leslie, C.C. Cytosolic Phospholipase A2: Physiological Function and Role in Disease. J. Lipid Res. 2015, 56, 1386–1402. [Google Scholar] [CrossRef] [PubMed]
- Wenk, G.L. Neuropathologic Changes in Alzheimer’s Disease. J. Clin. Psychiatry 2003, 64 (Suppl. 9), 7–10. [Google Scholar]
- Desbène, C.; Malaplate-Armand, C.; Youssef, I.; Garcia, P.; Stenger, C.; Sauvée, M.; Fischer, N.; Rimet, D.; Koziel, V.; Escanyé, M.-C.; et al. Critical Role of CPLA2 in Aβ Oligomer-Induced Neurodegeneration and Memory Deficit. Neurobiol. Aging 2012, 33, 1123.e17–1123.e29. [Google Scholar] [CrossRef]
- Anfuso, C.D.; Assero, G.; Lupo, G.; Nicotra, A.; Cannavò, G.; Strosznajder, R.P.; Rapisarda, P.; Pluta, R.; Alberghina, M. Amyloid Beta(1-42) and Its Beta(25-35) Fragment Induce Activation and Membrane Translocation of Cytosolic Phospholipase A2 in Bovine Retina Capillary Pericytes. Biochim. Biophys. Acta 2004, 1686, 125–138. [Google Scholar] [CrossRef]
- Sun, G.Y.; Geng, X.; Teng, T.; Yang, B.; Appenteng, M.K.; Greenlief, C.M.; Lee, J.C. Dynamic Role of Phospholipases A2 in Health and Diseases in the Central Nervous System. Cells 2021, 10, 2963. [Google Scholar] [CrossRef] [PubMed]
- Kaltschmidt, B.; Uherek, M.; Volk, B.; Baeuerle, P.A.; Kaltschmidt, C. Transcription Factor NF-ΚB Is Activated in Primary Neurons by Amyloid β Peptides and in Neurons Surrounding Early Plaques from Patients with Alzheimer Disease. Proc. Natl. Acad. Sci. USA 1997, 94, 2642–2647. [Google Scholar] [CrossRef] [PubMed]
- Teng, T.; Dong, L.; Ridgley, D.M.; Ghura, S.; Tobin, M.K.; Sun, G.Y.; LaDu, M.J.; Lee, J.C. Cytosolic Phospholipase A2 Facilitates Oligomeric Amyloid-β Peptide Association with Microglia via Regulation of Membrane-Cytoskeleton Connectivity. Mol. Neurobiol. 2019, 56, 3222–3234. [Google Scholar] [CrossRef]
- Sanchez-Mejia, R.O.; Newman, J.W.; Toh, S.; Yu, G.-Q.; Zhou, Y.; Halabisky, B.; Cissé, M.; Scearce-Levie, K.; Cheng, I.H.; Gan, L.; et al. Phospholipase A2 Reduction Ameliorates Cognitive Deficits in a Mouse Model of Alzheimer’s Disease. Nat. Neurosci. 2008, 11, 1311–1318. [Google Scholar] [CrossRef]
- Shah, K.; Lahiri, D.K. Cdk5 Activity in the Brain—Multiple Paths of Regulation. J. Cell Sci. 2014, 127, 2391–2400. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, J.R.; Chan, E.S.; Poore, C.P.; Pareek, T.K.; Cheong, W.F.; Shui, G.; Tang, N.; Low, C.-M.; Wenk, M.R.; Kesavapany, S. Cdk5/P25-Induced Cytosolic PLA2-Mediated Lysophosphatidylcholine Production Regulates Neuroinflammation and Triggers Neurodegeneration. J. Neurosci. 2012, 32, 1020–1034. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Bowen, W.C.; Li, G.; Demetris, A.J.; Michalopoulos, G.K.; Wu, T. Cytosolic Phospholipase A2α and Peroxisome Proliferator-Activated Receptor γ Signaling Pathway Counteracts Transforming Growth Factor β–Mediated Inhibition of Primary and Transformed Hepatocyte Growth. Hepatology 2010, 52, 644–655. [Google Scholar] [CrossRef]
- Yarla, N.S.; Bishayee, A.; Vadlakonda, L.; Chintala, R.; Duddukuri, G.; Pallu, R.; Kaladhar, D.S.V.G.K. Phospholipase A2 Isoforms as Novel Targets for Prevention and Treatment of Inflammatory and Oncologic Diseases. Curr. Drug Targets 2016, 17, 1940–1962. [Google Scholar] [CrossRef] [PubMed]
- Quach, N.D.; Arnold, R.D.; Cummings, B.S. Secretory Phospholipase A2 Enzymes as Pharmacological Targets for Treatment of Disease. Biochem. Pharmacol. 2014, 90, 338–348. [Google Scholar] [CrossRef]
- Chen, L.; Fu, H.; Luo, Y.; Chen, L.; Cheng, R.; Zhang, N.; Guo, H. CPLA2α Mediates TGF-β-Induced Epithelial–Mesenchymal Transition in Breast Cancer through PI3k/Akt Signaling. Cell Death Dis. 2017, 8, e2728. [Google Scholar] [CrossRef]
- Chiorazzo, M.G.; Tunset, H.M.; Popov, A.V.; Johansen, B.; Moestue, S.; Delikatny, E.J. Detection and Differentiation of Breast Cancer Sub-Types Using a CPLA2α Activatable Fluorophore. Sci. Rep. 2019, 9, 6122. [Google Scholar] [CrossRef]
- Wen, Z.-H.; Su, Y.-C.; Lai, P.-L.; Zhang, Y.; Xu, Y.-F.; Zhao, A.; Yao, G.-Y.; Jia, C.-H.; Lin, J.; Xu, S.; et al. Critical Role of Arachidonic Acid-Activated MTOR Signaling in Breast Carcinogenesis and Angiogenesis. Oncogene 2013, 32, 160–170. [Google Scholar] [CrossRef]
- Balsinde, J.; Balboa, M.A. Cellular Regulation and Proposed Biological Functions of Group VIA Calcium-Independent Phospholipase A2 in Activated Cells. Cell. Signal. 2005, 17, 1052–1062. [Google Scholar] [CrossRef]
- Lio, Y.-C.; Dennis, E. Interfacial Activation, Lysophospholipase and Transacylase Activity of Group VI Ca2+-Independent Phospholipase A2. Biochim. Et Biophys. Acta BBA-Lipids Lipid Metab. 1998, 1392, 320–332. [Google Scholar] [CrossRef]
- Jin, T.; Lin, J.; Gong, Y.; Bi, X.; Hu, S.; Lv, Q.; Chen, J.; Li, X.; Chen, J.; Zhang, W.; et al. IPLA2β Contributes to ER Stress-Induced Apoptosis during Myocardial Ischemia/Reperfusion Injury. Cells 2021, 10, 1446. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.H.; Mancuso, D.J.; Sims, H.F.; Liu, X.; Nguyen, A.L.; Yang, K.; Guan, S.; Dilthey, B.G.; Jenkins, C.M.; Weinheimer, C.J.; et al. Cardiac Myocyte-Specific Knock-out of Calcium-Independent Phospholipase A2γ (IPLA2γ) Decreases Oxidized Fatty Acids during Ischemia/Reperfusion and Reduces Infarct Size. J. Biol. Chem. 2016, 291, 19687–19700. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.D.; Gottlieb, R.A. Inhibition of Mitochondrial Calcium-Independent Phospholipase A2 (IPLA2) Attenuates Mitochondrial Phospholipid Loss and Is Cardioprotective. Biochem. J. 2002, 362, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wilkins, W.P.; Hu, W.; Murthy, K.S.; Chen, J.; Lee, Z.; Barbour, S.E.; Fang, X. Expression and Growth-Promoting Effect of Group VIA Calcium Independent Phospholipase A2 in Ovarian Cancer. Cancer Res. 2006, 66, 224. [Google Scholar]
- Zhao, X.; Wang, D.; Zhao, Z.; Xiao, Y.; Sengupta, S.; Xiao, Y.; Zhang, R.; Lauber, K.; Wesselborg, S.; Feng, L.; et al. Caspase-3-Dependent Activation of Calcium-Independent Phospholipase A2 Enhances Cell Migration in Non-Apoptotic Ovarian Cancer Cells. J. Biol. Chem. 2006, 281, 29357–29368. [Google Scholar] [CrossRef]
- Song, Y.; Wilkins, P.; Hu, W.; Murthy, K.S.; Chen, J.; Lee, Z.; Oyesanya, R.; Wu, J.; Barbour, S.E.; Fang, X. Inhibition of Calcium-Independent Phospholipase A2 Suppresses Proliferation and Tumorigenicity of Ovarian Carcinoma Cells. Biochem. J. 2007, 406, 427–436. [Google Scholar] [CrossRef]
- Kispert, S.; Schwartz, T.; McHowat, J. Cigarette Smoke Regulates Calcium-Independent Phospholipase A2 Metabolic Pathways in Breast Cancer. Am. J. Pathol. 2017, 187, 1855–1866. [Google Scholar] [CrossRef]
- Calderon, L.E.; Liu, S.; Arnold, N.; Breakall, B.; Rollins, J.; Ndinguri, M. Bromoenol Lactone Attenuates Nicotine-Induced Breast Cancer Cell Proliferation and Migration. PLoS ONE 2015, 10, e0143277. [Google Scholar] [CrossRef]
- McHowat, J.; Gullickson, G.; Hoover, R.G.; Sharma, J.; Turk, J.; Kornbluth, J. Platelet-Activating Factor and Metastasis: Calcium-Independent Phospholipase A2 β Deficiency Protects against Breast Cancer Metastasis to the Lung. Am. J. Physiol.-Cell Physiol. 2011, 300, C825–C832. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wang, K.; Shen, J. Lipoprotein-Associated Phospholipase A2: The Story Continues. Med. Res. Rev. 2020, 40, 79–134. [Google Scholar] [CrossRef]
- Rosenson, R.S.; Stafforini, D.M. Modulation of Oxidative Stress, Inflammation, and Atherosclerosis by Lipoprotein-Associated Phospholipase A2. J. Lipid Res. 2012, 53, 1767–1782. [Google Scholar] [CrossRef]
- Braun, L.T.; Davidson, M.H. Lp-PLA2: A New Target for Statin Therapy. Curr. Atheroscler. Rep. 2010, 12, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Mallat, Z.; Lambeau, G.; Tedgui, A. Lipoprotein-Associated and Secreted Phospholipases A2 in Cardiovascular Disease: Roles as Biological Effectors and Biomarkers. Circulation 2010, 122, 2183–2200. [Google Scholar] [CrossRef] [PubMed]
- Kono, N.; Inoue, T.; Yoshida, Y.; Sato, H.; Matsusue, T.; Itabe, H.; Niki, E.; Aoki, J.; Arai, H. Protection against Oxidative Stress-Induced Hepatic Injury by Intracellular Type II Platelet-Activating Factor Acetylhydrolase by Metabolism of Oxidized Phospholipids in Vivo. J. Biol. Chem. 2008, 283, 1628–1636. [Google Scholar] [CrossRef]
- Matsuzawa, A.; Hattori, K.; Aoki, J.; Arai, H.; Inoue, K. Protection against Oxidative Stress-Induced Cell Death by Intracellular Platelet-Activating Factor-Acetylhydrolase II. J. Biol. Chem. 1997, 272, 32315–32320. [Google Scholar] [CrossRef] [PubMed]
- Abe, A.; Hiraoka, M.; Wild, S.; Wilcoxen, S.E.; Paine, R.; Shayman, J.A. Lysosomal Phospholipase A2 Is Selectively Expressed in Alveolar Macrophages. J. Biol. Chem. 2004, 279, 42605–42611. [Google Scholar] [CrossRef]
- Hiraoka, M.; Abe, A.; Lu, Y.; Yang, K.; Han, X.; Gross, R.W.; Shayman, J.A. Lysosomal Phospholipase A2 and Phospholipidosis. Mol. Cell. Biol. 2006, 26, 6139–6148. [Google Scholar] [CrossRef]
- Hiraoka, M.; Abe, A.; Inatomi, S.; Sawada, K.; Ohguro, H. Augmentation of Lysosomal Phospholipase A2 Activity in the Anterior Chamber in Glaucoma. Curr. Eye Res. 2016, 41, 683–688. [Google Scholar] [CrossRef]
- Hiraoka, M.; Abe, A.; Lennikov, A.; Kitaichi, N.; Ishida, S.; Ohguro, H. Increase of Lysosomal Phospholipase A2 in Aqueous Humor by Uveitis. Exp. Eye Res. 2014, 118, 13–19. [Google Scholar] [CrossRef]
- Baheti, U.; Siddique, S.S.; Foster, C.S. Cataract Surgery in Patients with History of Uveitis. Saudi J. Ophthalmol. 2012, 26, 55–60. [Google Scholar] [CrossRef]
- Sers, C.; Husmann, K.; Nazarenko, I.; Zhumabayeva, B.; Adhikari, P. The Class II Tumour Suppressor Gene H-REV107-1 Is a Target of Interferon-Regulatory Factor-1 and Is Involved in IFNgamma-Induced Cell Death in Human Ovarian Carcinoma Cells. Oncogene 2002, 21, 2829–2839. [Google Scholar] [CrossRef] [PubMed]
- Parmer, C.; De Sousa-Coelho, A.L.; Cheng, H.S.; Daher, G.; Burkart, A.; Dreyfuss, J.M.; Pan, H.; Prenner, J.C.; Keilson, J.M.; Pande, R.; et al. Skeletal Muscle Expression of Adipose-Specific Phospholipase in Peripheral Artery Disease. Vasc. Med. 2020, 25, 401–410. [Google Scholar] [CrossRef] [PubMed]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Group | Subgroup | Source | Molecular Weight (kDa) | Catalytic Residues | Substrate Specificity | Secreting Cells | Ref. |
|---|---|---|---|---|---|---|---|---|
| sPLA2 | I | A | Cobras/kraits | 13–15 | His/Asp Ca2+ is required for substrate binding and catalysis | More active against zwitterionic PC | [6] | |
| I | B | Human/porcine pancreas | 13–15 | Pancreatic acinar cells | [9] | |||
| II | A | Rattlesnake, human synovial fluid | 13–15 | High affinity for anionic phospholipids such as PE, PG, PS | Synovial fluid, tears, leukocytes, platelets, macrophages, monocytes, T-cells, neutrophils | [6,9] | ||
| II | B | Gaboon viper | 13–15 | [7] | ||||
| II | C | Rat/murine testis | 15 | [7] | ||||
| II | D, E, F | Human/murine pancreas/brain/spleen/heart/uterus/embryo/testis | 14–17 | G-IID in lymph tissue DC; G-IIE in hypertrophic adipocytes. These are also expressed in macrophages and smooth muscle cells | [9] | |||
| III | Lizard/bee | 15–18 | Sperm membrane phospholipid remodeling | CNS, peripheral, neural fibers, mast cells, epididymal epithelium | [9,10] | |||
| V | Human/murine macrophage/heart/lung | 14 | High affinity for cell membrane PCs | Bronchial epithelial cells, macrophage, neutrophils, eosinophils, cardiomyocytes, hypertrophic adipocytes | [9,11] | |||
| IX | Snail venom | [7] | ||||||
| X | Human spleen/thymus/leukocyte | 14 | High activity toward PCs | Bronchoalveolar lavage fluid of asthmatic patients, airway epithelium, macrophages, smooth muscle cells, adrenal glands, pancreatic β cells, gut epithelium, sperm acrosome, hair follicles | [9] | |||
| XI | A, B | Green rice shoots | 13 | [12] | ||||
| XII | A, B | Human/murine | 19 | Hepatocytes | [9] | |||
| XII | Parvovirus | <10 | [12] | |||||
| XIV | Symbiotic fungus/Streptomyces | 13–19 | [12] | |||||
| cPLA2 | IV | A, B, C, D, E, F | Human macrophage/pancreas/liver/brain/heart/skeletal muscle Murine placenta/heart/thyroid/stomach/testis/liver cells, neuronal cells | 60–144 | Ser/Asp Ca2+ is required for membrane binding and activation | High specificity for PC, PE containing arachidonic acid at sn-2 position | Intracellularly localized in endoplasmic reticulum, Golgi complexes in epithelial cells | [6,13] |
| iPLA2 | VI | A, B, C, D, E, F | Human/murine | 84–90 | Ser/Asp Ca2+ is not required for catalysis | Phospholipase. Lysophospholipase activity on PL and lyso-PL, transacylase, acyl-CoA thioesterase activity | Group VI-A localizes on mitochondrial membrane and group VI-B on mitochondria and peroxisomes; group VI-C is expressed in neurons and localized on ER and Golgi body; groups VI-D, VI-E and VI-F are localized in adipose tissues | [6,13] |
| Lp-PLA2 (Lipoprotein- associated PLA2) | VII | A, B | Human, porcine, murine, bovine | 40–45 | Ser/His/Asp Ca2+ is not required for catalysis | Groups VII and VIII hydrolyze the acetyl group from sn-2 position of PAF and generate lyso-PAF and acetate. Group VII-A prefers PC with a shorter chain at sn-2 position | Group VII-A is secreted extracellularly by macrophages, monocyte, mast cells and T-cells. These are also associated with LDL and HDL in plasma; group VII-B is intracellularly expressed in epithelial cells of the kidney, intestine and hepatocytes. They are also overexpressed in neurons; group VIII is intracellularly expressed in brain cells | [6,14] |
| VIII | A, B | Human | 26–40 | |||||
| LPLA2 (Lysosomal PLA2) | XV | Human, murine, bovine | 45 | Ser/His/Asp Ca2+ is not required for catalysis | LPLA2 is specific towards PC and PE. They hydrolyze the acyl chain at both sn-1 and sn-2 positions of PC and PE with a preference for sn-2 position | Highly expressed in alveolar macrophages; localized in lysosomes | [15] | |
| Ad-PLA2 (Adipose tissue specific PLA2) | XVI | Human, mouse | 18 | Cys/His/His Ca2+ is not required for catalysis | Ubiquitously expressed throughout the body. Highly expressed in adipose tissue. | [6,16] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, S.A.; Ilies, M.A. The Phospholipase A2 Superfamily: Structure, Isozymes, Catalysis, Physiologic and Pathologic Roles. Int. J. Mol. Sci. 2023, 24, 1353. https://doi.org/10.3390/ijms24021353
Khan SA, Ilies MA. The Phospholipase A2 Superfamily: Structure, Isozymes, Catalysis, Physiologic and Pathologic Roles. International Journal of Molecular Sciences. 2023; 24(2):1353. https://doi.org/10.3390/ijms24021353
Chicago/Turabian StyleKhan, Shibbir Ahmed, and Marc A. Ilies. 2023. "The Phospholipase A2 Superfamily: Structure, Isozymes, Catalysis, Physiologic and Pathologic Roles" International Journal of Molecular Sciences 24, no. 2: 1353. https://doi.org/10.3390/ijms24021353
APA StyleKhan, S. A., & Ilies, M. A. (2023). The Phospholipase A2 Superfamily: Structure, Isozymes, Catalysis, Physiologic and Pathologic Roles. International Journal of Molecular Sciences, 24(2), 1353. https://doi.org/10.3390/ijms24021353