
Specific Binding of the α-Component of the Lantibiotic Lichenicidin to the Peptidoglycan Precursor Lipid II Predetermines Its Antimicrobial Activity

 ,
,  , ,
, ,  ,
,  , ,
, ,  ,
,  and
and
Abstract
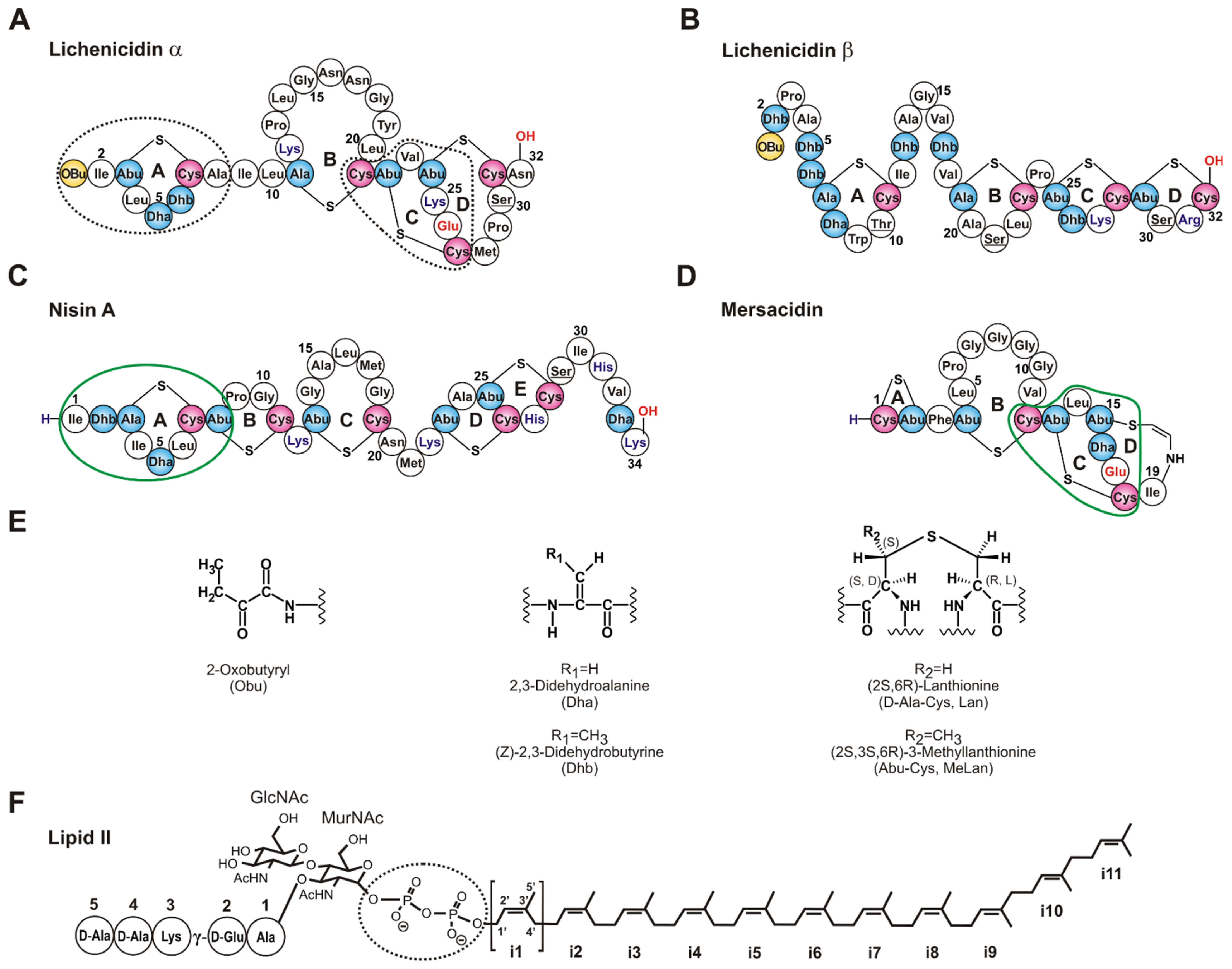
1. Introduction

2. Results
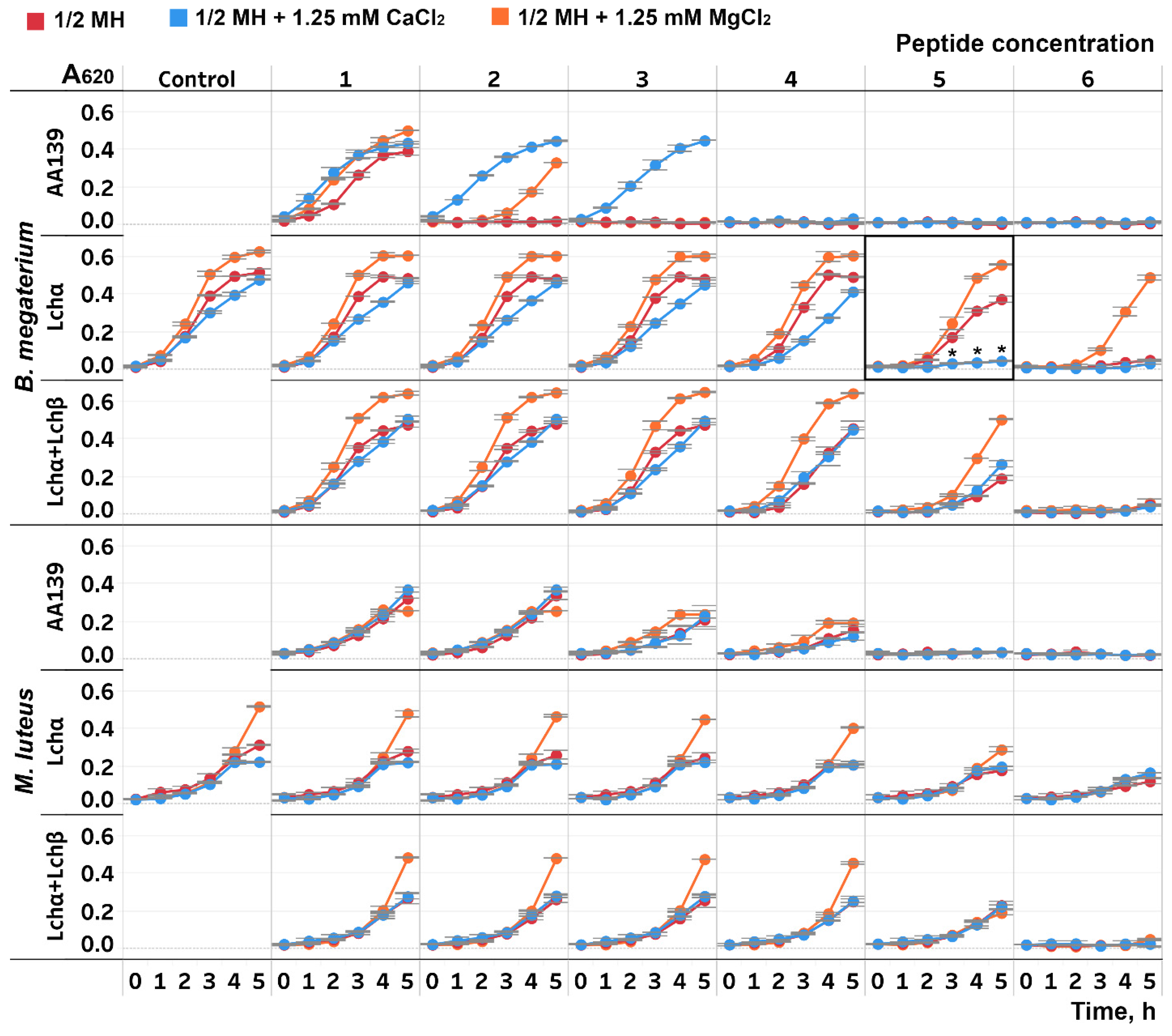
2.1. The Impact of Divalent Cations on the Antibacterial Activity of Lchα
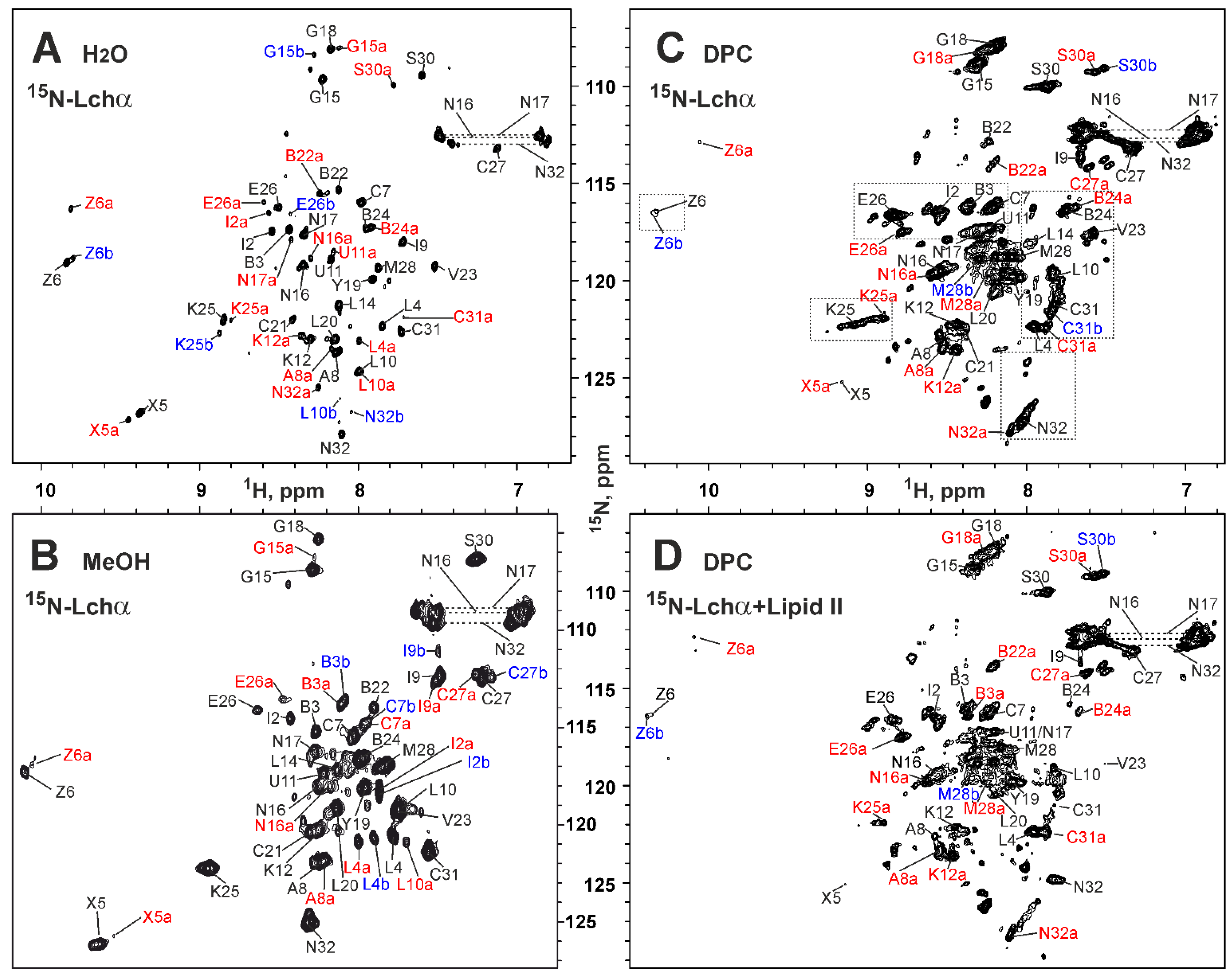
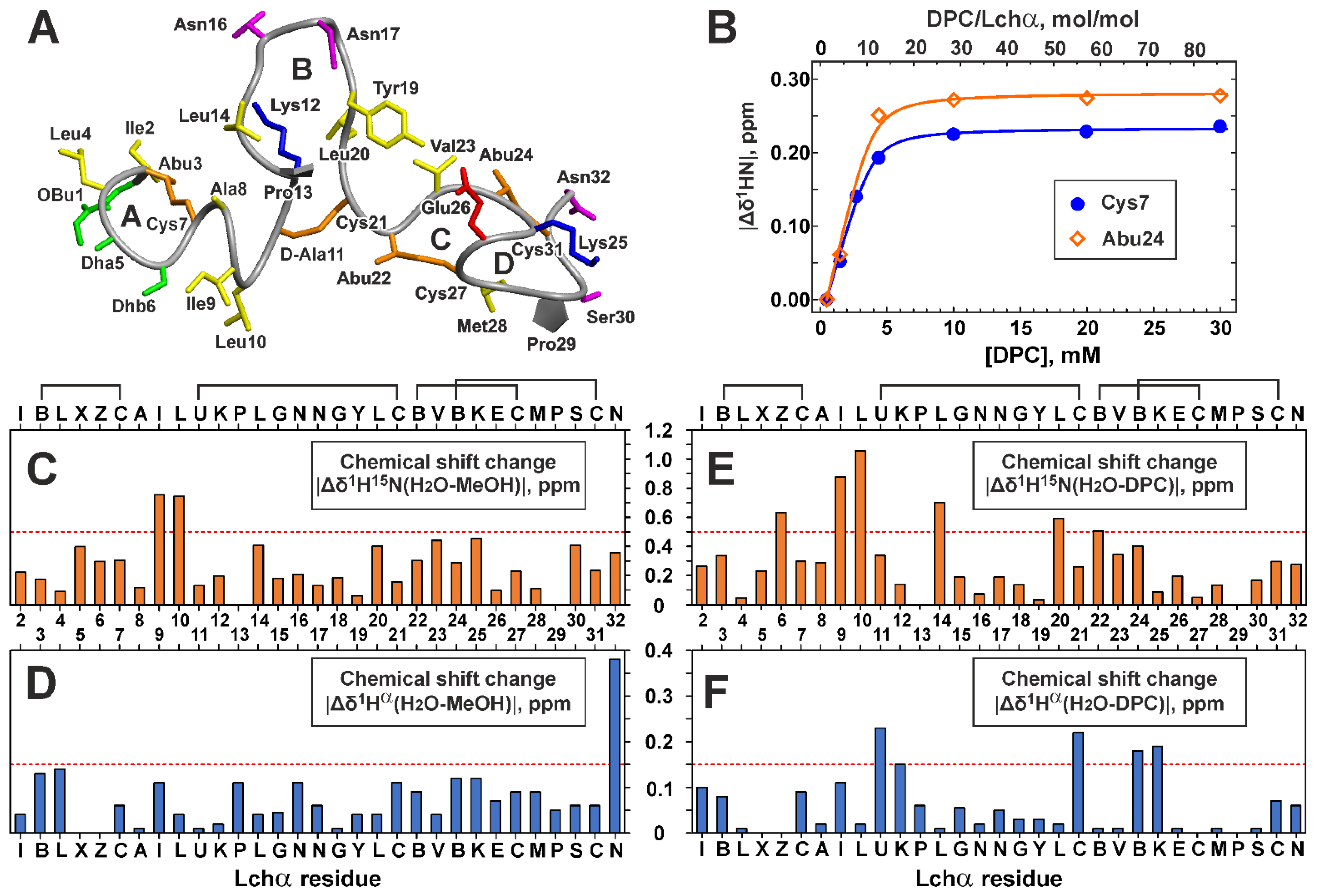
2.2. NMR Study of Lchα in Water and Methanol
2.3. NMR Study of Lchα in DPC Micelles
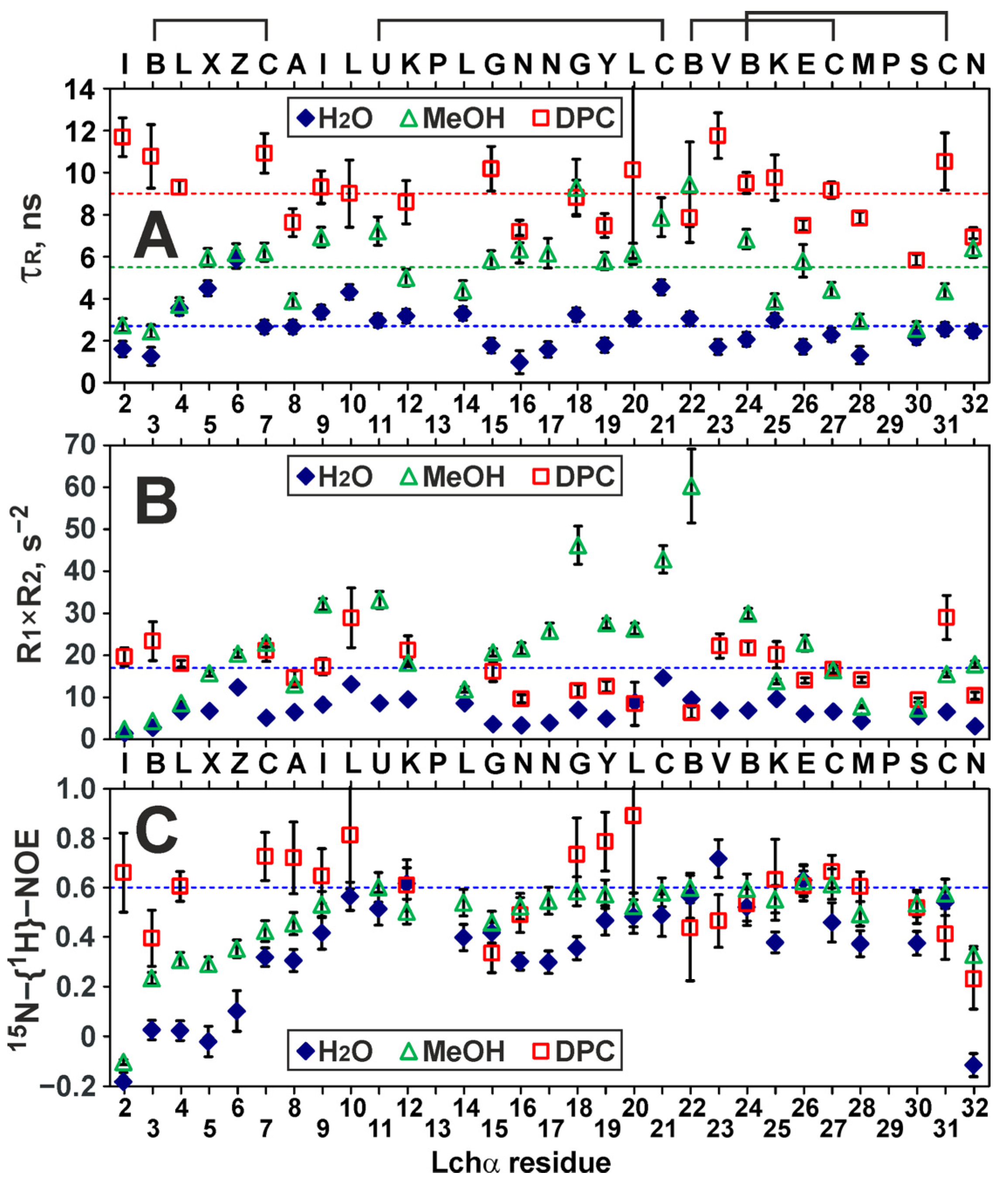
2.4. Backbone Dynamics of Lchα in Water, Methanol, and DPC Micelles
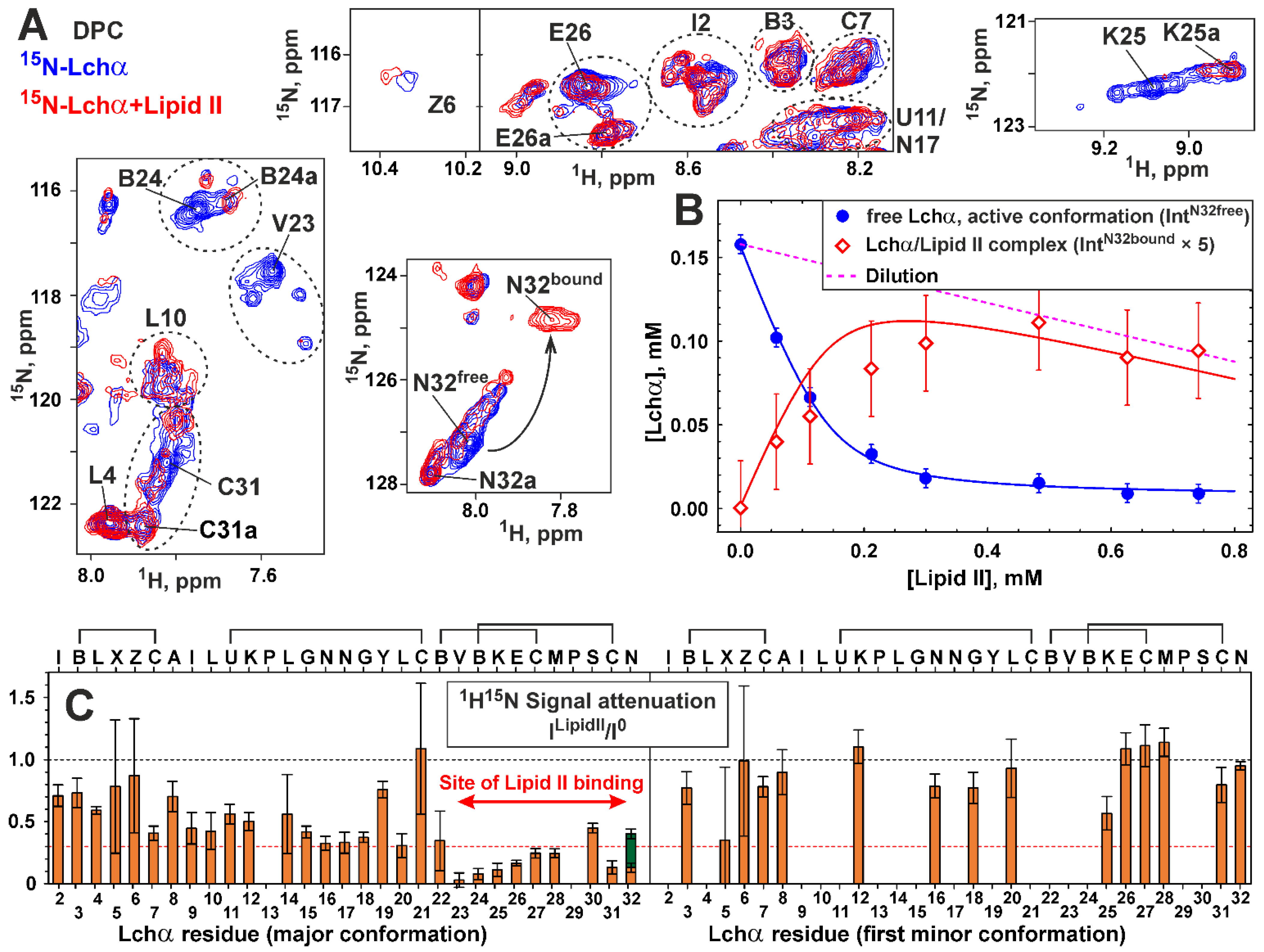
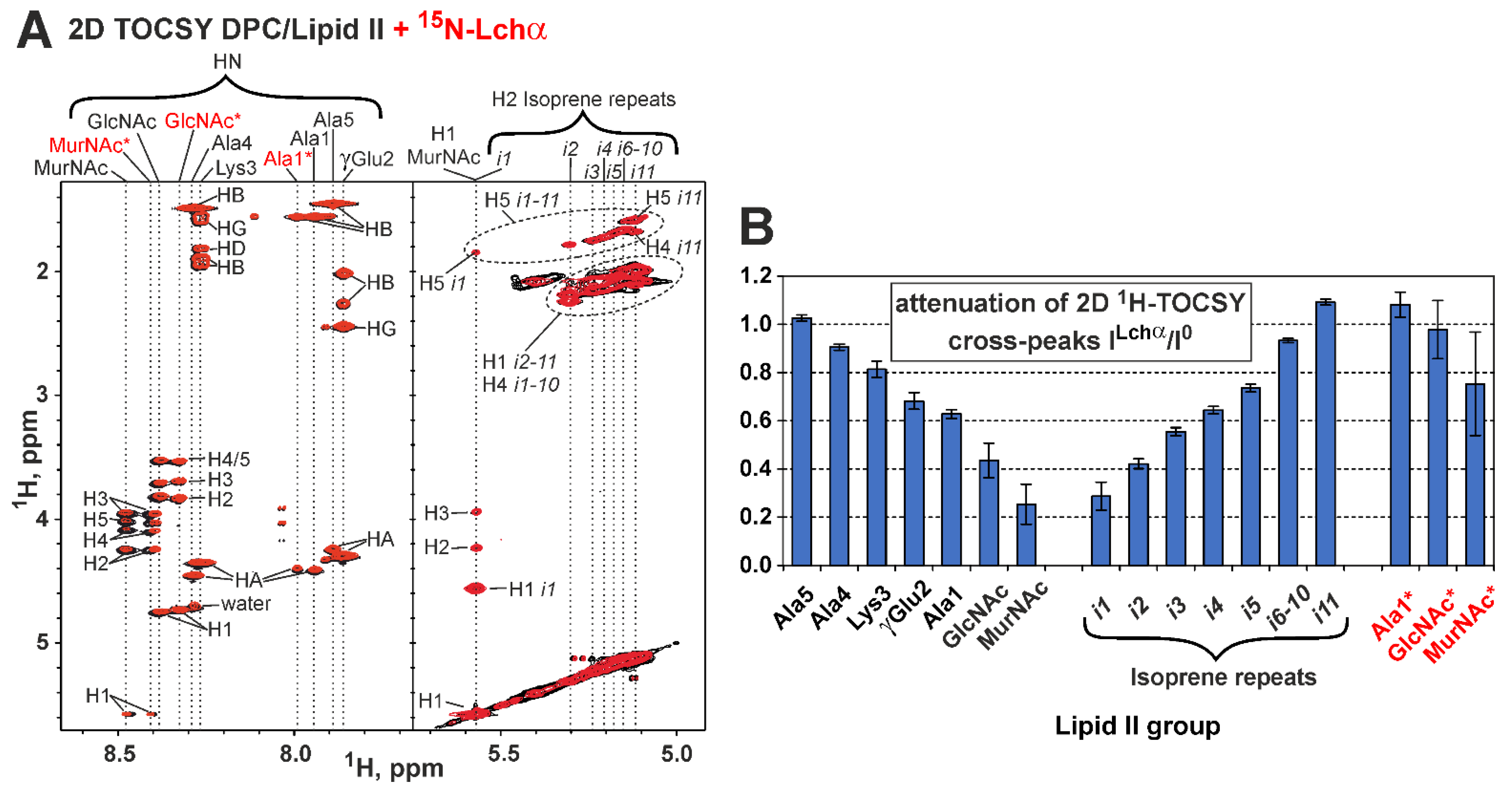
2.5. NMR Investigation of Lchα/Lipid II Interaction in DPC Micelles
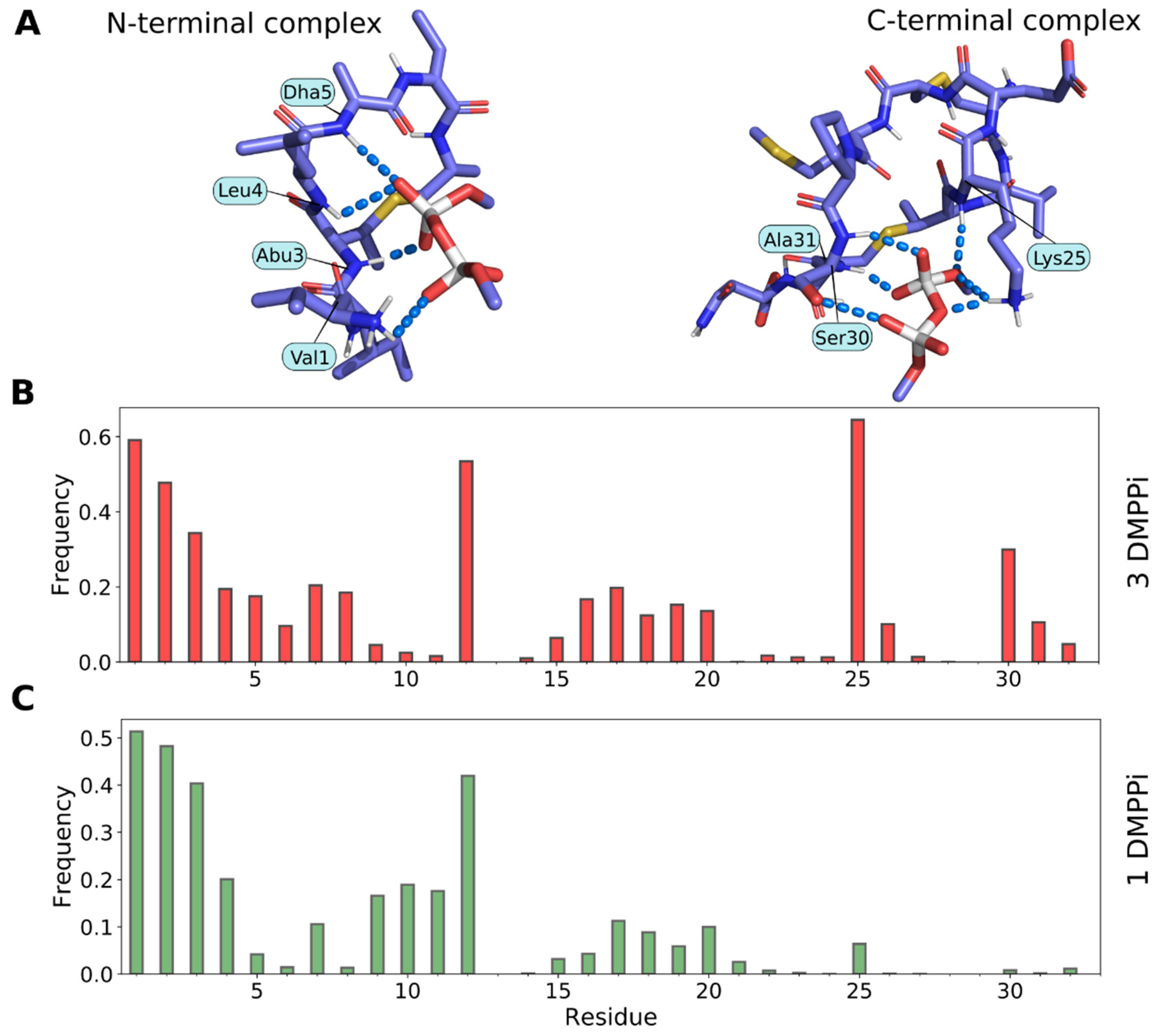
2.6. Molecular Modeling of Pyrophosphate Recognition by Lchα
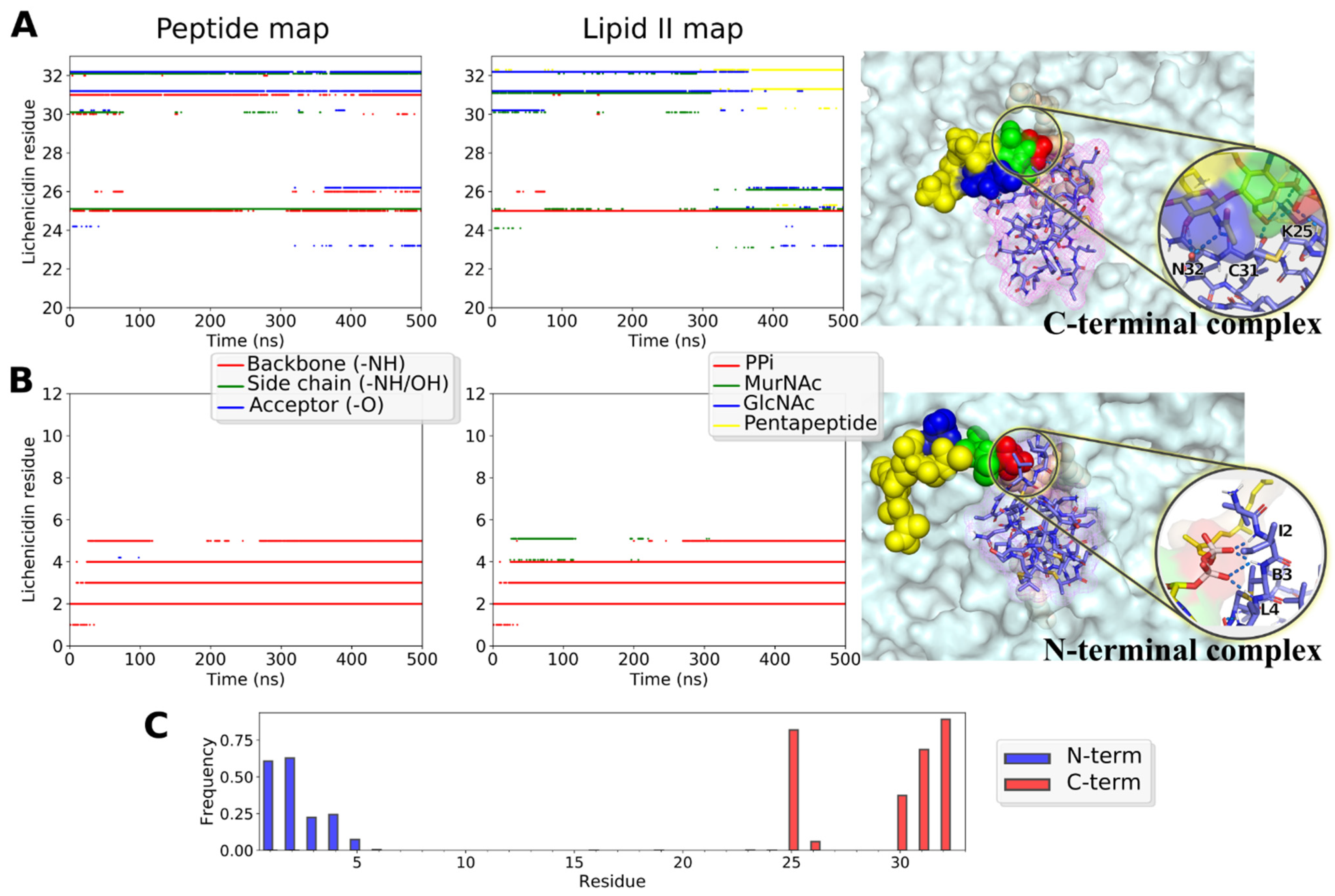
2.7. Predicted Structure of Lchα/Lipid II Complexes in the Model Bacterial Membrane
3. Discussion
4. Materials and Methods
4.1. Lipid II Preparation
4.2. Antimicrobial Peptide Preparations
4.3. Antibacterial Activity Assay
4.4. NMR Experiments and Data Analysis
4.5. Molecular Modeling
- Full-length Lchα in the presence of DMPPi, which mimics the lipid II pyrophosphate group, in water solution. The initial conformation of Lchα was retrieved from the recalculated NMR structure with corrected chirality of Cβ atom of Abu residues. The force-field parameters for the non-canonical amino acids such as unsaturated Dha, Dhb, lanthionine, and methyllanthionine were taken from our previous work on nisin [21]. The N-terminal 2-oxobutyryl group was replaced with the standard aliphatic valine residue. The peptide was solvated with water, and one or three DMPPi ions were randomly placed in a cubic box with a minimum distance of 7 Å to Lchα, in order to determine the preferable Lchα binding site, as well as the mutual adaptation of the peptide and its target.
- Truncated Lchα analogs containing potential lipid II binding sites: nisin-like N-terminal Lchα1–21 fragment and mersacidin-like C-terminal Lchα22–32 fragment in the presence of DMPPi in water. The starting coordinates for Lchα1–21 and Lchα22–32 for MD calculations were extracted from equilibrated states of the full-length molecule. Several Ca2+-containing trajectories were calculated for this and the previous group of simulations to investigate the role of this ion in intermolecular interactions of Lchα (see Table 1).
- Full-length Lchα in the presence of the hydrated lipid bilayer (with the peptide initially randomly placed outside the bilayer interfacial region): each bilayer leaflet contained 144 lipid molecules and was composed of the palmitoyl-oleoyl-phosphatidylglycerol(POPG)/palmitoyl-oleoyl-phosphatidylethanolamine (POPE) mixture at a ratio of 3:1, which mimics the Gram-positive bacteria membrane [68]. The semi-isotropic pressure coupling in the bilayer plane and along the membrane normal was used in the simulation. MD-averaged values of the area per lipid and the POPG/POPE bilayer thickness were 54.8 ± 0.3 Å2 and 39.2 ± 0.4 Å, respectively.
4.6. Accessing Codes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Angelopoulou, A.; Field, D.; Pérez-Ibarreche, M.; Warda, A.K.; Hill, C.; Ross, R.P. Vancomycin and nisin A are effective against biofilms of multi-drug resistant Staphylococcus aureus isolates from human milk. PLoS ONE 2020, 15, e0233284. [Google Scholar] [CrossRef] [PubMed]
- Henderson, L.O.; Erazo Flores, B.J.; Skeens, J.; Kent, D.; Murphy, S.I.; Wiedmann, M.; Guariglia-Oropeza, V. Nevertheless, she resisted–role of the environment on Listeria monocytogenes sensitivity to nisin treatment in a laboratory cheese model. Front. Microbiol. 2020, 11, 635. [Google Scholar] [CrossRef]
- Singh, A.P.; Preet, S.; Rishi, P. Nisin/β-lactam adjunct therapy against Salmonella enterica serovar typhimurium: A mechanistic approach. J. Antimicrob. Chemother. 2014, 69, 1877–1887. [Google Scholar] [CrossRef]
- El-Kazzaz, S.S.; Abou El-Khier, N.T. Effect of the lantibiotic nisin on inhibitory and bactericidal activities of antibiotics used against vancomycin-resistant Enterococci. J. Glob. Antimicrob. Resist. 2020, 22, 263–269. [Google Scholar] [CrossRef]
- Zhou, L.; van Heel, A.J.; Montalban-Lopez, M.; Kuipers, O.P. Potentiating the activity of nisin against Escherichia coli. Front. Cell Dev. Biol. 2016, 4, 7. [Google Scholar] [CrossRef]
- Breukink, E.; de Kruijff, B. Lipid II as a target for antibiotics. Nat. Rev. Drug Discov. 2006, 5, 321–332. [Google Scholar] [CrossRef]
- Oppedijk, S.F.; Martin, N.I.; Breukink, E. Hit ’em where it hurts: The growing and structurally diverse family of peptides that target lipid-II. Biochim. Biophys. Acta 2016, 1858, 947–957. [Google Scholar] [CrossRef]
- Müller, A.; Klöckner, A.; Schneider, T. Targeting a cell wall biosynthesis hot spot. Nat. Prod. Rep. 2017, 34, 909–932. [Google Scholar] [CrossRef]
- Grein, F.; Schneider, T.; Sahl, H.-G. Docking on lipid II—A widespread mechanism for potent bactericidal activities of antibiotic peptides. J. Mol. Biol. 2019, 431, 3520–3530. [Google Scholar] [CrossRef]
- Medeiros-Silva, J.; Jekhmane, S.; Breukink, E.; Weingarth, M. Towards the native binding modes of antibiotics that target lipid II. Chembiochem. Eur. J. Chem. Biol. 2019, 20, 1731–1738. [Google Scholar] [CrossRef]
- Hsu, S.-T.D.; Breukink, E.; Tischenko, E.; Lutters, M.A.G.; de Kruijff, B.; Kaptein, R.; Bonvin, A.M.J.J.; van Nuland, N.A.J. The nisin-lipid II complex reveals a pyrophosphate cage that provides a blueprint for novel antibiotics. Nat. Struct. Mol. Biol. 2004, 11, 963–967. [Google Scholar] [CrossRef]
- Münch, D.; Müller, A.; Schneider, T.; Kohl, B.; Wenzel, M.; Bandow, J.E.; Maffioli, S.; Sosio, M.; Donadio, S.; Wimmer, R.; et al. The lantibiotic NAI-107 binds to bactoprenol-bound cell wall precursors and impairs membrane functions. J. Biol. Chem. 2014, 289, 12063–12076. [Google Scholar] [CrossRef]
- Müller, A.; Ulm, H.; Reder-Christ, K.; Sahl, H.-G.; Schneider, T. Interaction of type A lantibiotics with undecaprenol-bound cell envelope precursors. Microb. Drug Resist. Larchmt. N 2012, 18, 261–270. [Google Scholar] [CrossRef]
- Islam, M.R.; Nishie, M.; Nagao, J.; Zendo, T.; Keller, S.; Nakayama, J.; Kohda, D.; Sahl, H.-G.; Sonomoto, K. Ring A of nukacin ISK-1: A lipid ii-binding motif for type-A(II) lantibiotic. J. Am. Chem. Soc. 2012, 134, 3687–3690. [Google Scholar] [CrossRef] [PubMed]
- Brötz, H.; Bierbaum, G.; Leopold, K.; Reynolds, P.E.; Sahl, H.G. The lantibiotic mersacidin inhibits peptidoglycan synthesis by targeting lipid II. Antimicrob. Agents Chemother. 1998, 42, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Oman, T.J.; Lupoli, T.J.; Wang, T.-S.A.; Kahne, D.; Walker, S.; van der Donk, W.A. Haloduracin α binds the peptidoglycan precursor lipid II with 2:1 stoichiometry. J. Am. Chem. Soc. 2011, 133, 17544–17547. [Google Scholar] [CrossRef] [PubMed]
- Wiedemann, I.; Böttiger, T.; Bonelli, R.R.; Wiese, A.; Hagge, S.O.; Gutsmann, T.; Seydel, U.; Deegan, L.; Hill, C.; Ross, P.; et al. The mode of action of the lantibiotic lacticin 3147–a complex mechanism involving specific interaction of two peptides and the cell wall precursor lipid II. Mol. Microbiol. 2006, 61, 285–296. [Google Scholar] [CrossRef]
- Shenkarev, Z.O.; Finkina, E.I.; Nurmukhamedova, E.K.; Balandin, S.V.; Mineev, K.S.; Nadezhdin, K.D.; Yakimenko, Z.A.; Tagaev, A.A.; Temirov, Y.V.; Arseniev, A.S.; et al. Isolation, structure elucidation, and synergistic antibacterial activity of a novel two-component lantibiotic lichenicidin from Bacillus licheniformis VK21. Biochemistry 2010, 49, 6462–6472. [Google Scholar] [CrossRef]
- Caetano, T.; Krawczyk, J.M.; Mösker, E.; Süssmuth, R.D.; Mendo, S. Heterologous expression, biosynthesis, and mutagenesis of type II lantibiotics from Bacillus licheniformis in Escherichia coli. Chem. Biol. 2011, 18, 90–100. [Google Scholar] [CrossRef]
- Wiedemann, I.; Breukink, E.; van Kraaij, C.; Kuipers, O.P.; Bierbaum, G.; de Kruijff, B.; Sahl, H.G. Specific binding of nisin to the peptidoglycan precursor lipid II combines pore formation and inhibition of cell wall biosynthesis for potent antibiotic activity. J. Biol. Chem. 2001, 276, 1772–1779. [Google Scholar] [CrossRef]
- Panina, I.; Krylov, N.; Nolde, D.; Efremov, R.; Chugunov, A. Environmental and dynamic effects explain how nisin captures membrane-bound lipid II. Sci. Rep. 2020, 10, 8821. [Google Scholar] [CrossRef]
- Szekat, C.; Jack, R.W.; Skutlarek, D.; Färber, H.; Bierbaum, G. Construction of an expression system for site-directed mutagenesis of the lantibiotic mersacidin. Appl. Environ. Microbiol. 2003, 69, 3777–3783. [Google Scholar] [CrossRef]
- Bakhtiary, A.; Cochrane, S.A.; Mercier, P.; McKay, R.T.; Miskolzie, M.; Sit, C.S.; Vederas, J.C. Insights into the mechanism of action of the two-peptide lantibiotic lacticin 3147. J. Am. Chem. Soc. 2017, 139, 17803–17810. [Google Scholar] [CrossRef] [PubMed]
- Medeiros-Silva, J.; Jekhmane, S.; Paioni, A.L.; Gawarecka, K.; Baldus, M.; Swiezewska, E.; Breukink, E.; Weingarth, M. High-resolution NMR studies of antibiotics in cellular membranes. Nat. Commun. 2018, 9, 3963. [Google Scholar] [CrossRef] [PubMed]
- Martin, N.I.; Sprules, T.; Carpenter, M.R.; Cotter, P.D.; Hill, C.; Ross, R.P.; Vederas, J.C. Structural characterization of lacticin 3147, a two-peptide lantibiotic with synergistic activity. Biochemistry 2004, 43, 3049–3056. [Google Scholar] [CrossRef] [PubMed]
- Holo, H.; Jeknic, Z.; Daeschel, M.; Stevanovic, S.; Nes, I.F. Plantaricin W from Lactobacillus plantarum belongs to a new family of two-peptide lantibiotics. Microbiol. Read. Engl. 2001, 147, 643–651. [Google Scholar] [CrossRef]
- McClerren, A.L.; Cooper, L.E.; Quan, C.; Thomas, P.M.; Kelleher, N.L.; van der Donk, W.A. Discovery and in vitro biosynthesis of haloduracin, a two-component lantibiotic. Proc. Natl. Acad. Sci. USA 2006, 103, 17243–17248. [Google Scholar] [CrossRef] [PubMed]
- Begley, M.; Cotter, P.D.; Hill, C.; Ross, R.P. Identification of a novel two-peptide lantibiotic, lichenicidin, following rational genome mining for LanM proteins. Appl. Environ. Microbiol. 2009, 75, 5451–5460. [Google Scholar] [CrossRef]
- Sawa, N.; Wilaipun, P.; Kinoshita, S.; Zendo, T.; Leelawatcharamas, V.; Nakayama, J.; Sonomoto, K. Isolation and characterization of enterocin W, a novel two-peptide lantibiotic produced by Enterococcus faecalis NKR-4-1. Appl. Environ. Microbiol. 2012, 78, 900–903. [Google Scholar] [CrossRef]
- Zhao, X.; van der Donk, W.A. Structural characterization and bioactivity analysis of the two-component lantibiotic Flv system from a ruminant bacterium. Cell Chem. Biol. 2016, 23, 246–256. [Google Scholar] [CrossRef][Green Version]
- Huo, L.; van der Donk, W.A. Discovery and characterization of bicereucin, an unusual D-amino acid-containing mixed two-component lantibiotic. J. Am. Chem. Soc. 2016, 138, 5254–5257. [Google Scholar] [CrossRef] [PubMed]
- Xin, B.; Zheng, J.; Liu, H.; Li, J.; Ruan, L.; Peng, D.; Sajid, M.; Sun, M. Thusin, a novel two-component lantibiotic with potent antimicrobial activity against several gram-positive pathogens. Front. Microbiol. 2016, 7, 1115. [Google Scholar] [CrossRef]
- Singh, M.; Chaudhary, S.; Sareen, D. Roseocin, a novel two-component lantibiotic from an actinomycete. Mol. Microbiol. 2020, 113, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Cooper, L.E.; McClerren, A.L.; Chary, A.; van der Donk, W.A. Structure-activity relationship studies of the two-component lantibiotic haloduracin. Chem. Biol. 2008, 15, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Oman, T.J.; van der Donk, W.A. Insights into the mode of action of the two-peptide lantibiotic haloduracin. ACS Chem. Biol. 2009, 4, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Field, D.; Collins, B.; Cotter, P.D.; Hill, C.; Ross, R.P. A System for the random mutagenesis of the two-peptide lantibiotic lacticin 3147: Analysis of mutants producing reduced antibacterial activities. J. Mol. Microbiol. Biotechnol. 2007, 13, 226–234. [Google Scholar] [CrossRef]
- Barbosa, J.C.; Gonçalves, S.; Makowski, M.; Silva, Í.C.; Caetano, T.; Schneider, T.; Mösker, E.; Süssmuth, R.D.; Santos, N.C.; Mendo, S. Insights into the mode of action of the two-peptide lantibiotic lichenicidin. Coll. Surf. B Biointerfaces 2022, 211, 112308. [Google Scholar] [CrossRef]
- Elliott, A.G.; Huang, J.X.; Neve, S.; Zuegg, J.; Edwards, I.A.; Cain, A.K.; Boinett, C.J.; Barquist, L.; Lundberg, C.V.; Steen, J.; et al. An Amphipathic Peptide with Antibiotic Activity against Multidrug-Resistant Gram-Negative Bacteria. Nat. Commun. 2020, 11, 3184. [Google Scholar] [CrossRef]
- Repka, L.M.; Chekan, J.R.; Nair, S.K.; van der Donk, W.A. Mechanistic understanding of lanthipeptide biosynthetic enzymes. Chem. Rev. 2017, 117, 5457–5520. [Google Scholar] [CrossRef]
- Sarksian, R.; Hegemann, J.D.; Simon, M.A.; Acedo, J.Z.; van der Donk, W.A. Unexpected methyllanthionine stereochemistry in the morphogenetic lanthipeptide SapT. J. Am. Chem. Soc. 2022, 144, 6373–6382. [Google Scholar] [CrossRef]
- Shenkarev, Z.O.; Nadezhdin, K.D.; Sobol, V.A.; Sobol, A.G.; Skjeldal, L.; Arseniev, A.S. Conformation and mode of membrane interaction in cyclotides. Spatial structure of kalata B1 bound to a dodecylphosphocholine micelle. FEBS J. 2006, 273, 2658–2672. [Google Scholar] [CrossRef] [PubMed]
- Kneller, J.M.; Lu, M.; Bracken, C. An Effective method for the discrimination of motional anisotropy and chemical exchange. J. Am. Chem. Soc. 2002, 124, 1852–1853. [Google Scholar] [CrossRef]
- Mineev, K.S.; Lesovoy, D.M.; Usmanova, D.R.; Goncharuk, S.A.; Shulepko, M.A.; Lyukmanova, E.N.; Kirpichnikov, M.P.; Bocharov, E.V.; Arseniev, A.S. NMR-based approach to measure the free energy of transmembrane helix-helix interactions. Biochim. Biophys. Acta 2014, 1838, 164–172. [Google Scholar] [CrossRef]
- Panina, I.; Taldaev, A.; Efremov, R.; Chugunov, A. Molecular dynamics insight into the lipid II recognition by type A lantibiotics: Nisin, epidermin, and gallidermin. Micromachines 2021, 12, 1169. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.; Caetano, T.; Mösker, E.; Süssmuth, R.; Mendo, S. Lichenicidin rational site-directed mutagenesis library: A tool to generate bioengineered lantibiotics. Biotechnol. Bioeng. 2019, 116, 3053–3062. [Google Scholar] [CrossRef]
- Barbosa, J.C.; Silva, Í.C.; Caetano, T.; Mösker, E.; Seidel, M.; Lourenço, J.; Süssmuth, R.D.; Santos, N.C.; Gonçalves, S.; Mendo, S. Assessing the potential of the two-peptide lantibiotic lichenicidin as a new generation antimicrobial. World J. Microbiol. Biotechnol. 2022, 38, 18. [Google Scholar] [CrossRef]
- Yuan, J.; Zhang, Z.-Z.; Chen, X.-Z.; Yang, W.; Huan, L.-D. Site-Directed Mutagenesis of the Hinge Region of NisinZ and Properties of NisinZ Mutants. Appl. Microbiol. Biotechnol. 2004, 64, 806–815. [Google Scholar] [CrossRef] [PubMed]
- Field, D.; Connor, P.M.O.; Cotter, P.D.; Hill, C.; Ross, R.P. The Generation of Nisin Variants with Enhanced Activity against Specific Gram-Positive Pathogens. Mol. Microbiol. 2008, 69, 218–230. [Google Scholar] [CrossRef]
- Balandin, S.V.; Ovchinnikova, T.V. Antimicrobial peptides of invertebrates. Part 2. Biological functions and mechanisms of action. Russ. J. Bioorgan. Chem. 2016, 42, 343–360. [Google Scholar] [CrossRef]
- Finkina, E.I.; Melnikova, D.N.; Bogdanov, I.V.; Ovchinnikova, T.V. Peptides of the innate immune system of plants. Part I. Structure, biological activity, and mechanisms of action. Russ. J. Bioorgan. Chem. 2018, 44, 573–585. [Google Scholar] [CrossRef]
- Barrett, M.S.; Wenzel, R.P.; Jones, R.N. In vitro activity of mersacidin (M87-1551), an investigational peptide antibiotic tested against gram-positive bloodstream isolates. Diagn. Microbiol. Infect. Dis. 1992, 15, 641–644. [Google Scholar] [CrossRef]
- Böttiger, T.; Schneider, T.; Martínez, B.; Sahl, H.-G.; Wiedemann, I. Influence of Ca(2+) ions on the activity of lantibiotics containing a mersacidin-like lipid II binding motif. Appl. Environ. Microbiol. 2009, 75, 4427–4434. [Google Scholar] [CrossRef]
- Hsu, S.-T.D.; Breukink, E.; Bierbaum, G.; Sahl, H.-G.; de Kruijff, B.; Kaptein, R.; van Nuland, N.A.J.; Bonvin, A.M.J.J. NMR study of mersacidin and lipid II interaction in dodecylphosphocholine micelles. Conformational changes are a key to antimicrobial activity. J. Biol. Chem. 2003, 278, 13110–13117. [Google Scholar] [CrossRef]
- Taku, A.; Stuckey, M.; Fan, D.P. Purification of the peptidoglycan transglycosylase of Bacillus megaterium. J. Biol. Chem. 1982, 257, 5018–5022. [Google Scholar] [CrossRef]
- Münch, D.; Sahl, H.-G. Structural variations of the cell wall precursor lipid II in Gram-positive bacteria-Impact on binding and efficacy of antimicrobial peptides. Biochim. Biophys. Acta 2015, 1848, 3062–3071. [Google Scholar] [CrossRef]
- Kohlrausch, U.; Höltje, J.V. Analysis of murein and murein precursors during antibiotic-induced lysis of Escherichia coli. J. Bacteriol. 1991, 173, 3425–3431. [Google Scholar] [CrossRef]
- Brötz, H.; Bierbaum, G.; Reynolds, P.E.; Sahl, H.G. The Lantibiotic mersacidin inhibits peptidoglycan biosynthesis at the level of transglycosylation. Eur. J. Biochem. 1997, 246, 193–199. [Google Scholar] [CrossRef]
- Breukink, E.; van Heusden, H.E.; Vollmerhaus, P.J.; Swiezewska, E.; Brunner, L.; Walker, S.; Heck, A.J.R.; de Kruijff, B. Lipid II is an intrinsic component of the pore induced by nisin in bacterial membranes. J. Biol. Chem. 2003, 278, 19898–19903. [Google Scholar] [CrossRef]
- Schneider, T.; Senn, M.M.; Berger-Bächi, B.; Tossi, A.; Sahl, H.-G.; Wiedemann, I. In vitro assembly of a complete, pentaglycine interpeptide bridge containing cell wall precursor (Lipid II-Gly5) of Staphylococcus aureus. Mol. Microbiol. 2004, 53, 675–685. [Google Scholar] [CrossRef]
- Rouser, G.; Fkeischer, S.; Yamamoto, A. Two dimensional then layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 1970, 5, 494–496. [Google Scholar] [CrossRef]
- Ovchinnikova, T.V.; Shenkarev, Z.O.; Nadezhdin, K.D.; Balandin, S.V.; Zhmak, M.N.; Kudelina, I.A.; Finkina, E.I.; Kokryakov, V.N.; Arseniev, A.S. Recombinant expression, synthesis, purification, and solution structure of arenicin. Biochem. Biophys. Res. Commun. 2007, 360, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Gunsteren, W.F.V.; Billeter, S.R.; Eising, A.A.; Hünenberger, P.H.; Krueger, P.; Mark, A.E.; Scott, W.R.P.; Tironi, I.G. Biomolecular Simulation: The GROMOS96 Manual and User Guide; Biomos: Zürich, Switzerland, 1996; ISBN 978-3-7281-2422-7. [Google Scholar]
- Berger, O.; Edholm, O.; Jähnig, F. Molecular dynamics simulations of a fluid bilayer of dipalmitoylphosphatidylcholine at full hydration, constant pressure, and constant temperature. Biophys. J. 1997, 72, 2002–2013. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical sampling through velocity rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Prossnigg, F.; Hickel, A.; Pabst, G.; Lohner, K. Packing behaviour of two predominant anionic phospholipids of bacterial cytoplasmic membranes. Biophys. Chem. 2010, 150, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Krylov, N.A.; Efremov, R.G. Libxtc: An Efficient library for reading XTC-compressed MD trajectory data. BMC Res. Notes 2021, 14, 124. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System Composition | MD Length (ns) | Number of Trajectories |
|---|---|---|
| Lichenicidin with DMPPi in solution | ||
| Lchα/DMPPi1/Water10336/Na+1 | 500 | 5 |
| Lchα/DMPPi3/Water9704/Na+5 | 500 | 2 |
| Lchα/DMPPi3/Water10321/Ca2+3/Cl−1 | 500 | 3 |
| Lchα1–21/DMPPi3/Water7648/Na+5 | 500 | 3 |
| Lchα1–21/DMPPi3/Water7648 /Ca2+3/Cl−1 | 500 | 2 |
| Lchα22–32/DMPPi3/Water9242/Na+6 | 500 | 2 |
| Lchα22–32/DMPPi3/Water10216/Ca2+3 | 500 | 3 |
| Lichenicidin in the membrane environment | ||
| Lchα/POPG126/POPE96/Water20358/Na+191 | 500 | 3 |
| Lichenicidin/lipid II complex in bacterial membrane | ||
| C-terminal complex | ||
| Lchα/Lipid II/POPG186/POPE66/Water11966/Na+188 | 500 | 3 |
| N-terminal complex | ||
| Lchα/Lipid II/POPG186/POPE66/Water11985/Na+188 | 500 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panina, I.S.; Balandin, S.V.; Tsarev, A.V.; Chugunov, A.O.; Tagaev, A.A.; Finkina, E.I.; Antoshina, D.V.; Sheremeteva, E.V.; Paramonov, A.S.; Rickmeyer, J.; et al. Specific Binding of the α-Component of the Lantibiotic Lichenicidin to the Peptidoglycan Precursor Lipid II Predetermines Its Antimicrobial Activity. Int. J. Mol. Sci. 2023, 24, 1332. https://doi.org/10.3390/ijms24021332
Panina IS, Balandin SV, Tsarev AV, Chugunov AO, Tagaev AA, Finkina EI, Antoshina DV, Sheremeteva EV, Paramonov AS, Rickmeyer J, et al. Specific Binding of the α-Component of the Lantibiotic Lichenicidin to the Peptidoglycan Precursor Lipid II Predetermines Its Antimicrobial Activity. International Journal of Molecular Sciences. 2023; 24(2):1332. https://doi.org/10.3390/ijms24021332
Chicago/Turabian StylePanina, Irina S., Sergey V. Balandin, Andrey V. Tsarev, Anton O. Chugunov, Andrey A. Tagaev, Ekaterina I. Finkina, Daria V. Antoshina, Elvira V. Sheremeteva, Alexander S. Paramonov, Jasmin Rickmeyer, and et al. 2023. "Specific Binding of the α-Component of the Lantibiotic Lichenicidin to the Peptidoglycan Precursor Lipid II Predetermines Its Antimicrobial Activity" International Journal of Molecular Sciences 24, no. 2: 1332. https://doi.org/10.3390/ijms24021332
APA StylePanina, I. S., Balandin, S. V., Tsarev, A. V., Chugunov, A. O., Tagaev, A. A., Finkina, E. I., Antoshina, D. V., Sheremeteva, E. V., Paramonov, A. S., Rickmeyer, J., Bierbaum, G., Efremov, R. G., Shenkarev, Z. O., & Ovchinnikova, T. V. (2023). Specific Binding of the α-Component of the Lantibiotic Lichenicidin to the Peptidoglycan Precursor Lipid II Predetermines Its Antimicrobial Activity. International Journal of Molecular Sciences, 24(2), 1332. https://doi.org/10.3390/ijms24021332