
Pulsatilla vulgaris Inhibits Cancer Proliferation in Signaling Pathways of 12 Reporter Genes

 ,
,  , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
2.1. Cytotoxicity Studies
2.2. The Chemical Composition of the Studied P. vulgaris Methanolic Extracts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comp. No | Tentative Identification | Rt (min) | Molecular Formula | MW | [M − H]− | Fragments (m/z) | L | R | Ref |
|---|---|---|---|---|---|---|---|---|---|
| 1. | Hydroxymelilotic acid | 8.99 | C9H10O4 | 182.0544 | 181.0544 | 163.0395; 149.0229; 135.0431; 119.0505 | + | + | [6] |
| 2. | Hydroxybenzoic acid isomer 1 | 9.560 | C7H6O3 | 138.0317 | 137.0247 | 119.0112; 109.0319; 93.0344 | + | + | |
| 3. | Caffeic acid hexoside | 10.382 | C15H18O9 | 342.0951 | 341.0900 | 179.0305; 161.0221; 133.0258; 135.0419 | + | + | |
| 4. | Dihydroxycoumarin-O-glucoside | 11.307 | C15H16O9 | 340.0794 | 339.0750 | 177.0236 161.0324 133.0333 | + | + | [76,77] |
| 5. | Caftaric acid | 12.582 | C13H12O9 | 312.0481 | 311.0373 | 179.0345; 149.0084; 135.0476; 112.9942 | + | + | [6,78] |
| 6. | Dihydroxy-methoxycoumarin | 12.260 | C10H8O5 | 208.0340 | 207.0340 | 163.0403; 135.0434; 109.0296 | + | + | [76,77] |
| 7. | Tartaric acid | 12.455 | C4H6O6 | 150.0143 | 149.0143 | 149.0143; 121.0241; 87.0046; 72.9914 | + | + | [76,78] |
| 8. | Dihydroxycoumarin | 15.653 | C9H6O4 | 178.0266 | 177.0202 | 133.0280; 105.0343 | + | + | [76,77] |
| 9. | Caffeic acid | 16.600 | C9H8O4 | 180.0423 | 179.0322 | 135.0459 | + | + | [6,78] |
| 10. | Dihydroxy-methoxycoumarin-O-glucoside | 17.706 | C16H18O10 | 370.0900 | 369.0894 | 207.0318; 192.0059; 163.0046 | + | + | [76,77] |
| 11. | Coumaric acid hexoside | 17.756 | C15H18O8 | 326.0982 | 325.0982 | 265.0663; 235.0568; 205.0486; 163.0359; 145.0282 | - | + | |
| 12. | Vanillic acid | 18.387 | C8H8O4 | 168.0423 | 167.0338 | 152.0089; 108.0209 | + | + | [6] |
| 13. | Ferulic acid derivative | 19.474 | - | - | 225.0790 | 193.0580; 161.0244; 135.0492 | + | + | [76] |
| 14. | Ferulic acid hexoside | 20.733 | C16H20O10 | 356.1107 | 355.1045 | 193.0522; 178.0223; 134.0387 | + | + | |
| 15. | Quercetin-O-deoxyhexoside-O-hexoside | 23.391 | C27H30O16 | 610.1534 | 609.1486 | 301.0351; 300.0194; 271.0257; 255.0332; 151.0051; 117.8824 | + | - | [76] |
| 16. | Hydroxybenzoic acid isomer 2 | 23.793 | C7H6O3 | 138.0317 | 137.0261 | 93.0365 | + | + | [6] |
| 17. | Ferulic acid isomer | 24.273 | C10H10O4 | 194.0579 | 193.0522 | 161.0259; 134.0383 | + | + | [6] |
| 18. | Ferulic acid | 25.276 | C10H10O4 | 194.0579 | 193.0520 | 134.0424; 161.0307; 178.0260 | + | + | [6,79] |
| 19. | Ferulic acid di-hexoside | 26.386 | C22H29O1 | 518.1752 | 517.1752 | 355.1153 | + | + | [80] |
| 20. | Dicaffeoyltartaric acid isomer 1 (=Chicoric acid isomer 1) | 28.361 | C22H18O12 | 474.0798 | 473.0712 | 311.0373; 179.0310; 149.0059; 135.0430 | + | + | [6,81] |
| 21. | 3-O-glucopyranosyl-arabinopyranosyl—bayogenin 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl ester | 28.687 | C59H96O28 | 1252.6044 | 1251.6044 | 781.4519; 619.3788; 487.3556; 471.0539; 469.1561 | - | traces | [6,73] |
| 22. | 3-O-glucopyranosyl-arabinopyranosyl—23-hydroxybetulinic acid 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl ester | 29.491 | C59H96O27 | 1236.6139 | 1235.5926 | 765.4470; 603.3781; 469.1522 | - | + | [6] |
| 23. | 3-O- arabinopyranosyl-bayogenin 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl ester | 29.760 | C53H86O23 | 1090.556 | 1089.5527 | 767.3829; 619.3829; 469.1502; 471.1573; 487.9452 | - | + | [6] |
| 24. | 3-O-glucopyranosyl-arabinopyranosyl—hederagenin 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl ester | 30.381 | C59H96O27 | 1236.6139 | 1235.6016 | 765.4402; 603.3883; 469.1542; 367.1197; 471.4602; 451.1425 | - | + | [6] |
| 25. | Dicaffeoyltartaric acid isomer 2 (=Chicoric acid isomer 2) | 30.586 | C22H18O12 | 474.0798 | 473.0712 | 311.0399; 179.0327; 149.0062; 135.0412 | + | + | [6,81] |
| 26. | Caffeoyl feruloyltartaric acid | 30.630 | C23H20O12 | 488.0957 | 487.0957 | 325.0517; 293.0265; 193.0431; 179.0310; 149.0115; 135.0437 | + | + | [81,82] |
| 27. | 3-O-xylopyranosyl (1-2)-α-L-arabinopyranosyl-23-hydroxybetulinic acid 28-O- rhamnopyranosyl (1-4)-glucopyranosyl (1-6)-glucopyranosyl ester | 30.993 | C58H94O26 | 1206.6033 | 1205.5922 | 735.3446; 603.4000; 469.1599 | - | + | [74] |
| 28. | Hederacoside C (3-O-arabinopyranosyl-rhamnopyranosyl-hederagenin 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl ester)/ Anemoside B4 (3-O-arabinopyranosyl-rhamnopyranosyl-23-hydroxybetulinic acid 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl ester) | 30.993 | C59H96O26 | 1220.619 | 1219.6033 | 749.4449; 603.4249; 469.1550; 471.0370 | - | + | [74] |
| 29. | 3-O-arabinopyranosyl-23-hydroxybetulinic acid 28-O-rhamnopyranosyl(1→4)–glucopyranosyl (1→6)-glucopyranosyl ester | 31.369 | C53H86O22 | 1074.5611 | 1119.5586 [M + HCOO]− | 1073.5510; 603.3963; 469.1590 | - | + | [6] |
| 30. | 3-O-rhamnopyranosyl-glucopyranosyl-hederagenin 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl ester /3-O-rhamnopyranosyl-glucopyranosyl-23-hydroxybetulinic acid 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl ester | 32.220 | C60H96O27 | 1248.6204 | 1247.6204 | 1187.6022; 777.4205; 469.1467 | - | + | |
| 31. | 3-O-arabinopyranosyl—hederagenin 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl ester | 33.829 | C53H86O22 | 1074.5611 | 1073.5628 | 603.4048; 471.1944; 469.1651; 409.1363 | - | + | [6] |
| 32. | 3β-[(O-β-D-xylopyranosyl (1→2)-α-L-arabinopyranosyl) oxy]lup-20-(29)-en-28-oic acid 28-O-α-L-rhamnopyranosyl-(1→4)-O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranosyl ester | 34.267 | C58H94O25 | 1190.6084 | 1189.6050 | 719.4455; 587.3973; 469.1608 | - | + | [74] |
| 33. | 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl 23- hydroxybetulinic acid ester | 34.502 | C48H78O18 | 942.5223 | 987.519 [M + HCOO]− | 941.4754; 471.3436; 469.1527 | - | + | [6] |
| 34. | derivative of 23-hydroxybetulinic acid 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl ester/ derivative of 28-O-rhamnopyranosyl(1-4)-glucopyranosyl(1-6)-glucopyranosyl hederagenin ester | 34.502 | - | 1027.5420 | 1026.5420 | 941.5067; 471.3329; 469.1383 | - | + | |
| 35. | Trihydroxy-octadecenoic acid | 35.697 | C18H34O5 | 330.2292 | 329.2292 | 293.2082; 229.1428; 211.1264; 171.0929 | + | + | [76] |
| 36. | 3-O-arabinopyranosyl–28-O-glucopyranosyl hederagenin ester | 39.886 | C41H66O13 | 766.4503 | 811.4472 [M + HCOO]− | 765.4380; 603.3835; 471.3425 | - | + | [6] |
| 37. | 3-O-Arabinopyranosyl –28-O-arabinopyranosyl hederagenin ester | 42.089 | C40H64O12 | 736.4486 | 781.4397 [M + HCOO]− | 735.4486; 603.3976; 471.3566 | - | + | |
| 38. | Fatty acid | 46.663 | C18H30O3 | 294.2175 | 293.2175 | 275.1987; 224.1362; 195.1427; 171.0968 | tr | + | [76] |
| 39. | Fatty acid | 48.944 | C18H32O3 | 296.2344 | 295.2334 | 277.2158; 195.1374; 177.1248 | tr | + | [76] |
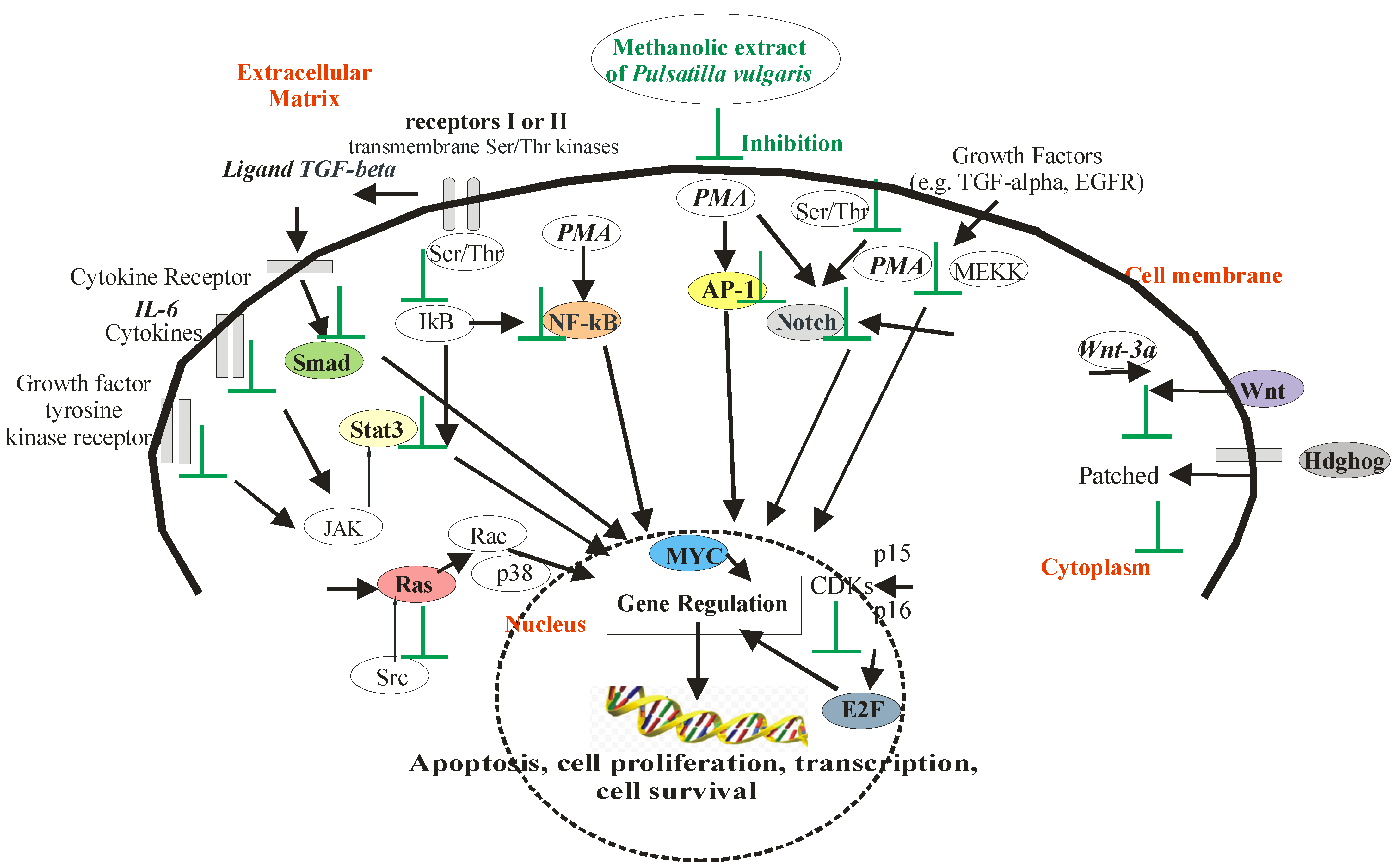
2.3. The Cancer Activity of P. vulgaris in Signaling Pathways in HeLa Cells
3. Materials and Methods
3.1. Plant Material
3.2. LC/ESI-QTOF-MS Procedure
3.3. Cell Cultures and Media
3.4. Cytotoxicity Testing
3.5. Transfection and Luciferase Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Wink, M.; Van Wyk, B.E. Mind-Altering and Poisonous Plants of the World. A Scientifically Accurate Guide to 1200 Toxic and Intoxicating Plants, 1st ed.; Timber Press: Portland, OR, USA, 2008; pp. 1–464. [Google Scholar]
- Jiangsu New Medicine College. Encyclopedia of Chinese Materia Medica, 1st ed.; Shanghai Science and Technology Press: Shanghai, China, 1977; pp. 1–294. [Google Scholar]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China, 1st ed.; People’s Medical Publishing House: Beijing, China, 2005; pp. 1–946.
- Kumar, S.; Madaan, R.; Farooq, A.; Sharma, A. The genus Pulsatilla: A review. Pharmacogn. Rev. 2008, 2, 116. [Google Scholar]
- Li, W.; Ding, Y.; Sun, Y.N.; Yan, X.T.; Yang, S.Y.; Choi, G.W.; Kim, E.-J.; Kang, H.K.; Kim, Y.H. Oleanane-type triterpenoid saponins from the roots of Pulsatilla koreana and their apoptosis-inducing effects on HL-60 human promyelocytic leukemia cells. Arch. Pharm. Res. 2013, 36, 768–774. [Google Scholar] [CrossRef]
- Łaska, G.; Maciejewska-Turska, M.; Sieniawska, E.; Świątek, Ł.; Pasco, D.S.; Balachandran, P. Extracts from Pulsatilla patens target cancer-related signaling pathways in HeLa cells. Sci. Rep. 2021, 11, 10654. [Google Scholar] [CrossRef]
- Hensen, I.; Oberprieler, C.; Wesche, K. Genetic structure, population size, and seed production of Pulsatilla vulgaris Mill. (Ranunculaceae) in Central Germany. Flora Morphol. Distrib. Funct. Ecol. Plants 2005, 200, 3–14. [Google Scholar] [CrossRef]
- Kaźmierczakowa, R.; Zarzycki, K.; Mirek, Z. Polish Red Data Book of Plants. Pteridophytes and Flowering Plants, 3rd ed.; Institute of Nature Conservation, Polish Academy of Sciences: Cracow, Poland, 2014; pp. 1–895. [Google Scholar]
- Łaska, G.; Sienkiewicz, A. Antifungal activity of the rhizome extracts of Pulsatilla vulgaris against Candida glabrata. Eur. J. Biol. Res. 2019, 9, 93–103. [Google Scholar]
- Zhang, Y.; Bao, J.; Wang, K.; Jia, X.; Zhang, C.; Huang, B.; Chen, M.; Wan, J.B.; Su, H.; Wang, Y.; et al. Pulsatilla saponin D inhibits autophagic flux and synergistically enhances the anticancer activity of chemotherapeutic agents against HeLa cells. Am. J. Chin. Med. 2015, 43, 1657–1670. [Google Scholar] [CrossRef]
- Łaska, G.; Sienkiewicz, A.; Stocki, M.; Sharma, V.; Zjawiony, J.K.; Jacob, M. Secondary metabolites from Pulsatilla patens and Pulsatilla vulgaris and their biological activity. Planta Med. 2015, 81, 122. [Google Scholar] [CrossRef]
- Łaska, G.; Sienkiewicz, A.; Stocki, M.; Zjawiony, J.K.; Sharma, V.; Bajguz, A.; Piotrowska-Niczyporuk, A.; Jacob, M.; Khan, S. Phytochemical screening of Pulsatilla species and investigation of their biological activities. Acta Soc. Bot. Pol. 2019, 88, 3613. [Google Scholar] [CrossRef]
- Ye, W.C.; Ji, N.N.; Zhao, S.X.; Che, C.T. A new cytotoxic saponin from Pulsatilla patens var. multifida. Pharm. Biol. 2001, 39, 7–10. [Google Scholar] [CrossRef]
- Ye, W.C.; Zhang, Q.W.; Hsiao, W.L.W.; Zhao, S.X.; Che, C.T. Novel cytotoxic lupane-type glycosides from Pulsatilla chinensis. Planta Med. 2002, 68, 183–186. [Google Scholar] [CrossRef]
- Shu, Z.; Chen, Z.; Liu, Y.L.; Zhu, W.F.; Feng, Y.L.; Xu, Q.M.; Li, X.R.; Yang, S.L. A new oleanane-type triterpenoidal saponin from Pulsatilla chinensis. Nat. Prod. Res. 2013, 27, 2196–2201. [Google Scholar] [CrossRef]
- Xu, Q.M.; Shu, Z.; He, W.J.; Chen, L.Y.; Yang, S.L.; Yang, G.; Liu, Y.L.; Li, X.R. Antitumor activity of Pulsatilla chinensis (Bunge) Regel saponins in human liver tumor 7402 cells in vitro and in vivo. Phytomedicine 2012, 19, 293–300. [Google Scholar] [CrossRef]
- Bi, Y.L.; Wang, B.; Huang, B.H.; Zhang, W.T.; Zhang, Y.H. Antifungal activity of botanical extracts against Botrytis cinerea and Alternaria solani. J. Agric. Sci. Technol. 2011, 12, 862–864. [Google Scholar]
- Ma, C. Treatment methods of traditional Chinese medicines against intestinal protozoan infections. In Treatment of Human Parasitosis in Traditional Chinese Medicine, 1st ed.; Mehlhorn, H., Zhongdao, W., Ye, B., Eds.; Springer: Berlin, Germany, 2014; Volume 6, pp. 11–21. [Google Scholar] [CrossRef]
- Li, W.C.; Shi, W.J.; Gu, Y.F.; Chen, H.L.; Chen, W.; Zhang, Y. Antibacterial effect of different extract of Pulsatilla chinensis (Bunge) Regel in vitro. J. Tradit. Chin. Vet. Med. 2011, 2, 38–40. [Google Scholar]
- Li, L.D.; Li, W.C.; Liu, C.W.; Shi, W.J.; Gong, P.T.; Li, J.H.; Zhang, G.C.; Yang, J.; Li, H.; Zhang, X.C. Giardia intestinalis: Effects of Pulsatilla chinensis extracts on trophozoites. Parasitol. Res. 2012, 111, 1929–1935. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Xu, Q.M.; Liu, Y.L.; Li, X.R.; Yang, S.L.; Zhuge, H.X. Laboratory evaluation of the molluscicidal activity of Pulsatilla chinensis (Bunge) Regel saponins against the snail Oncomelania hupensis. Biomed. Environ. Sci. 2012, 25, 224–229. [Google Scholar] [CrossRef]
- Kim, Y.; Bang, S.C.; Lee, J.H.; Ahn, B.Z. Pulsatilla saponin D: The antitumor principle from Pulsatilla koreana. Arch. Pharm. Res. 2004, 27, 915–918. [Google Scholar] [CrossRef]
- Yang, H.; Cho, Y.W.; Kim, S.H.; Kim, Y.C.; Sung, S.H. Triterpenoidal saponins of Pulsatilla koreana roots. Phytochemistry 2010, 71, 1892–1899. [Google Scholar] [CrossRef]
- Yoo, H.H.; Lee, S.K.; Lim, S.Y.; Kim, Y.; Kang, M.J.; Kim, E.J.; Park, Y.H.; Im, G.J.; Lee, B.Y.; Kim, D.H. LC-MS/MS method for determination of hederacolchiside E, a neuroactive saponin from Pulsatilla koreana extract in rat plasma for pharmacokinetic study. J. Pharm. Biomed. Anal. 2008, 48, 1425–1429. [Google Scholar] [CrossRef]
- Bang, S.C.; Kim, Y.; Lee, J.H.; Ahn, B.Z. Triterpenoid saponins from the roots of Pulsatilla koreana. J. Nat. Prod. 2005, 68, 268–272. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, S.B.; You, Y.J.; Ahn, B.Z. Deoxypodophyllotoxin; the cytotoxic and antiangiogenic komponent from Pulsatilla koreana. Planta Med. 2002, 68, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.W.; Chung, C.H.; Lim, S.B.; Kim, J.K.; So, E.H. The antimicrobial effect of Pulsatilla koreana extracts to oral micro-organism. J. Korean Acad. Periodontol. 2000, 30, 661. [Google Scholar] [CrossRef]
- Li, W.; Sun, Y.N.; Yan, X.Y.; Yang, S.Y.; Lee, S.J.; Byun, H.J.; Moon, C.S.; Han, B.S.; Kim, Y.H. Isolation of nematicidal triterpenoid saponins from Pulsatilla koreana root and their activities against Meloidogyne incognita. Molecules 2013, 18, 5306–5316. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.A.; Choi, B.K.; Jeong, C.S.; Li, D.W.; Lee, E.B. The anti-inflammatory and analgesic actions of the fractions from Pulsatilla koreana root extract. Korean J. Clin. Pharm. 2000, 31, 174–184. [Google Scholar]
- Schrenk, D. Feasibility study of nonclinical safety assessments on homeopathic preparations using the example of protoanemonin in Pulsatilla pratensis L. Regul. Toxicol. Pharmacol. 2013, 66, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Samadder, A.; Das, J.; Das, S.; Khuda-Bukhsh, A.R. Dihydroxy-isosteviol-methyl-ester, an active biological component of Pulsatilla nigricans, reduces arsenic induced cellular dysfunction in testis of male mice. Environ. Toxicol. Pharmacol. 2012, 34, 743–752. [Google Scholar] [CrossRef]
- Fan, W.; Liu, J.; Gong, Y.; Ma, J.; Zhou, N.; Xu, Y. A new triterpenoid saponin from Pulsatilla cernua. Nat. Prod. Sci. 2013, 19, 150–154. [Google Scholar]
- Zhang, Q.W.; Ye, W.C.; Che, C.T.; Zhao, S.X. Triterpene saponins from Pulsatilla cernua. Acta Pharm. Sin. 2000, 35, 756–759. [Google Scholar]
- Xu, H.J.; Shi, X.W.; Ji, X.; Du, Y.F.; Zhu, H.; Zhang, L.T. A rapid method for simultaneous determination of triterpenoid saponins in Pulsatilla turczaninovii using microwave-assisted extraction and high performance liquid chromatography-tandem mass spectrometry. Food Chem. 2012, 135, 251–258. [Google Scholar] [CrossRef]
- Sun, H.; Wang, Y.; Zhang, X.Q.; Zhao, S.X.; Ye, W.C. Chemical constituents of Pulsatilla dahurica. Chem. Nat. Compd. 2009, 45, 764–765. [Google Scholar] [CrossRef]
- Li, H.B.; Wong, C.C.; Cheng, K.W.; Chen, F. Antioxidant properties in vitro and total phenolic contents in methanol extracts from medicinal plants. LWT—Food Sci. Technol. 2008, 41, 385–390. [Google Scholar] [CrossRef]
- Lee, H.S.; Beon, M.S.; Kim, M.K. Selective growth inhibitor toward human intestinal bacteria derived from Pulsatilla cernua root. J. Agric. Food Chem. 2001, 49, 4656–4661. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Wang, H.; Chen, Y. The immune-enhancing effect of PcG-A-a glycoprotein isolated from dried root of Pulsatilla chinensis (Bunge) Regel. Chin. J. Biochem. Pharm. 2000, 21, 230–231. [Google Scholar]
- Liu, J.Y.; Guan, Y.L.; Zou, L.B.; Gong, Y.X.; Hua, H.M.; Xu, Y.N.; Zhang, H.; Yu, Z.G.; Fan, W.H. Saponins with neuroprotective effects from the roots of Pulsatilla cernua. Molecules 2012, 17, 5520–5531. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.S.; Yun, J.H.; Baek, I.S.; Leem, Y.H.; Kang, H.W.; Cho, H.K.; Lyu, Y.S.; Son, H.J.; Han, P.L. Oriental medicine Jangwonhwan reduces Aβ(1–42) level and β-amyloid deposition in the brain of Tg- APPswe/PS1dE9 mouse model of Alzheimer disease. J. Ethnopharmacol. 2010, 128, 206–212. [Google Scholar] [CrossRef]
- Chen, Z.; Guan, Y.; Zhou, L.; Xu, Y.; Yang, M.; Liu, H. Preparation and characterization of colon-targeted particles of Pulsatilla chinensis saponins. Nat. Prod. Commun. 2015, 10, 237–238. [Google Scholar]
- Xu, K.; Shu, Z.; Xu, Q.M.; Liu, Y.L.; Li, X.R.; Wang, Y.L.; Yang, S.L. Cytotoxic activity of Pulsatilla chinensis saponins and their structure-activity relationship. J. Asian Nat. Prod. Res. 2013, 15, 680–686. [Google Scholar] [CrossRef]
- Borenfreund, E.; Babich, H.; Martin-Alguacil, N. Rapid chemosensitivity assay with human normal and tumor cells in vitro. In Vitro Cell Dev. Biol. 1990, 26, 1030. [Google Scholar] [CrossRef]
- Zaki, M.A.; Balachandran, P.; Khan, S.; Wang, M.; Mohammed, R.; Hetta, M.H.; Pasco, D.S.; Muhammad, I. Cytotoxicity and modulation of cancer-related signaling by (Z)- and (E)-3,4,3′,5′-tetramethoxystilbene isolated from Eugenia brigida. J. Nat. Prod. 2013, 76, 679–684. [Google Scholar] [CrossRef]
- Fotakis, G.; Timbrell, J.A. In vitro cytotoxicity assays: Comparison of LDH, neutral red, MTT and protein assay in hepatoma cell lines following exposure to cadmium chloride. Toxicol. Lett. 2006, 160, 171–177. [Google Scholar] [CrossRef]
- Weyermann, J.; Lochmann, D.; Zimmer, A. A practical note on the use of cytotoxicity assays. Int. J. Pharm. 2005, 288, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Danova, K.; Bertoli, A.; Pistelli, L.; Dimitrow, D.; Pistelli, L. In vitro culture of Balkan endemic and rare Pulsatilla species for conservational purposes and secondary metabolites production. Bot. Serb. 2009, 33, 157–162. [Google Scholar]
- Xu, H.J.; Shi, X.W.; Ji, X.; Du, Y.F.; Zhu, H.; Zhang, L.T. Qualitative and quantitative determination of nine main active constituents in Pulsatilla cernua by high-performance liquid chromatography coupled to electrospray ionization tandem mass spectrometry. J. Sep. Sci. 2011, 34, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Goyal, S.; Kumar, S. Anti-anxiety activity studies of various extracts of Pulsatilla nigricans Stoerck. Int. J. Pharm. Sci. Drug Res. 2010, 2, 291–293. [Google Scholar]
- Nikolova, M.; Nikolov, S.; Koeva, Y. Flavonoids in the overground part of Pulsatilla slaviankae (Rumm.) D. Yord. et Koz. (Ranunculaceae). Phytol. Balc. 1998, 4, 165–170. [Google Scholar]
- Nikolova, M.; Asenov, A. Surface flavonoid aglycones in newly studied plant species. Nat. Prod. Res. 2006, 20, 103–106. [Google Scholar] [CrossRef]
- Luft, G. The distribution of saponins and tannins in the plant. Mon. Chem. 1926, 47, 259–284. [Google Scholar] [CrossRef]
- Raymond-Hamet, L. Pharmacodynamic properties of Anemone pulsatilla. Bull. Sci. Pharmacol. 1927, 34, 143–151. [Google Scholar]
- Kroeber, L. On the pharmacology and therapeutic use of the anemonine drugs from Ranunculaceous plants. Pharmazie 1949, 4, 181–190. [Google Scholar]
- Walker, K.J.; Pinches, C.E. Reduced grazing and the decline of Pulsatilla vulgaris Mill. (Ranunculaceae) in England, UK. Biol. Conserv. 2011, 144, 3098–3105. [Google Scholar] [CrossRef]
- Piqueray, J.; Saad, L.; Bizoux, J.P.; Mahy, G. Why some species cannot colonise restored habitats? The effects of seed and microsite availability. J. Nat. Conserv. 2013, 21, 189–197. [Google Scholar] [CrossRef]
- Kricsfalusy, V. Variations in the life cycle of Anemone patens L. (Ranunculaceae) in wild populations of Canada. Plants 2016, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, E.; Holderegger, R.; Matthies, D.; Rutishauser, R. Investigation on the population biology of a flagship species of dry meadows: Pulsatilla vulgaris Mill. in north-eastern Switzerland. Bot. Helv. 2002, 112, 153–172. [Google Scholar]
- Liu, D.D.; Chao, W.M.; Turgeon, R. Transport of sucrose, not hexose, in the phloem. J. Exp. Bot. 2012, 63, 4315–4320. [Google Scholar] [CrossRef]
- DiLeo, M.F.; Graf, R.; Holderegger, R.; Rico, Y.; Wagner, H.H. Highly polymorphic microsatellite markers in Pulsatilla vulgaris (Ranunculaceae) using next-generation sequencing. Appl. Plant Sci. 2015, 3, 1500031. [Google Scholar] [CrossRef]
- Niklfeld, H.; Schratt-Ehrendorfer, L. Red List of Threatened Plants of Austria, 1st ed.; Grüne Reihe; Bundesministerium für Umwelt, Jugend und Familie: Vienna, Austria, 1999; pp. 1–291.
- Ludwig, G.; Schnittler, M. Red List of Threatened Plants in Germany, 1st ed.; Bundesamt für Naturschutz: Bonn, Germany, 1996; pp. 1–72.
- Moser, D.; Gygax, A.; Bäumler, B.; Wyler, N.; Palese, R. Red List of the Threatened Ferns and Flowering Plants of Switzerland, 1st ed.; Bundesamt für Umwelt, Wald und Landschaft, Zentrum des Datenverbundnetzes der Schweizer Flora: Bern, Switzerland, 2002; pp. 1–120.
- IUCN. The IUCN Red List of Threatened Species, Version 2021. Available online: http://www.iucnredlist.org/ (accessed on 10 August 2021).
- Cheffings, C.; Farrell, L. The Vascular Plant Red Data List for Great Britain, 1st ed.; Joint Nature Conservation Committee: Peterborough, UK, 2005; pp. 1–120.
- Gärdenfors, U. The 2010 Red List of Swedish Species, 1st ed.; ArtDatabanken: Uppsala, Sweden, 2010. [Google Scholar]
- Witkowski, Z.J.; Król, W.; Solarz, W. Carpathian List of Endangered Species, 1st ed.; WWF and Institute of Nature Conservation, Polish Academy of Sciences: Vienna, Austria, 2003; pp. 1–84. [Google Scholar]
- Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Flowering Plants and Pteridophytes of Poland. A Checklist; W. Szafer Institute of Botany, Polish Academy of Sciences: Cracow, Poland, 2002; pp. 1–442. [Google Scholar]
- Thienthiti, K.; Tuchind, P.; Wongnoppavich, A.; Anantachoke, N.; Soonthornchareonnon, N. Cytotoxic effect of compounds isolated from Goniothalam usmarcanii Craib stem barks. Pharm. Sci. Asia 2017, 44, 86–95. [Google Scholar] [CrossRef]
- Figueiró, L.R.; Comerlato, L.C.; Da Silva, M.V.; Zuanazzi, J.; Von Poser, G.L.; Ziulkoski, A.L. Toxicity of Glandularia selloi (Spreng.) Tronc. leave extract by MTT and neutral red assays: Influence of the test medium procedure. Interdiscip. Toxicol. 2016, 9, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Zuang, V. The Neutral Red Release Assay: A Review. Altern. Lab. Anim. 2001, 29, 575–599. [Google Scholar] [CrossRef]
- Jin, M.-M.; Zhang, W.-D.; Jiang, H.-H.; Du, Y.-F.; Guo, W.; Cao, L.; Xu, H.-J. UPLC-Q-TOF-MS/MS-guided dereplication of Pulsatilla chinensis to identify triterpenoid saponins. Phytochem. Anal. 2018, 29, 516–527. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, H.; Yang, J.; Song, G.; Wen, D.; Liu, W.; Jin, M.; Wang, Q.; Du, Y.; Sun, Q.; et al. A high-throughput metabolomics approach for the comprehensive differentiation of four Pulsatilla Adans herbs combined with a nontargeted bidirectional screen for rapid identification of triterpenoid saponins. Anal. Bioanal. Chem. 2019, 411, 2071–2088. [Google Scholar] [CrossRef]
- Ling, Y.; Lin, Z.; Zha, W.; Lian, T.; You, S. Rapid detection and characterisation of triterpene saponins from the root of Pulsatilla chinensis (Bunge) Regel by HPLC-ESI-QTOF-MS/MS. Phytochem. Anal. 2016, 27, 174–183. [Google Scholar] [CrossRef]
- Lee, K.Y.; Cho, Y.W.; Park, J.; Lee, D.Y.; Kim, S.H.; Kim, Y.C.; Sung, S.H. Quality control of Pulsatilla koreana based on the simultaneous determination of triterpenoidal saponins by HPLC-ELSD and principal component analysis. Phytochem. Anal. 2010, 21, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Pubchem Database. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 10 September 2022).
- Zhao, M.; Ding, W.; Wang, S.; Wang, C.; Du, Y.; Xu, H.; Wang, Q.; Jin, S. Simultaneous determination of nine coumarins in rat plasma by HPLC-MS/MS for pharmacokinetics studies following oral administration of Fraxini Cortex extract. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1025, 25–32. [Google Scholar] [CrossRef] [PubMed]
- HMDB Human Metabolome Database. Available online: https://hmdb.ca/ (accessed on 10 September 2022).
- Hossain, M.B.; Rai, D.K.; Brunton, N.P.; Martin-Diana, A.B.; Barry-Ryan, A.C. Characterization of phenolic composition in lamiaceae spices by LC-ESI-MS/MS. J. Agric. Food Chem. 2010, 58, 10576–10581. [Google Scholar] [CrossRef] [PubMed]
- Usui, A.; Matsuo, Y.; Tanaka, T.; Ohshima, K.; Fukuda, S.; Mine, T.; Nakayama, H.; Ishimaru, K. Ferulic acid esters of oligo-glucose from Allium macrostemon. Nat. Prod. Commun. 2017, 12, 89–91. [Google Scholar] [CrossRef]
- Zeb, A.; Haq, A.; Murkovic, M. Effects of microwave cooking on carotenoids, phenolic compounds and antioxidant activity of Cichorium intybus L. (chicory) leaves. Eur. Food Res. Technol. 2019, 245, 365–374. [Google Scholar] [CrossRef]
- Mulinacci, N.; Innocenti, M.; Gallori, S.; Romani, A.; La Marca, G.; Vincieri, F.F. Optimization of the chromatographic determination of polyphenols in the aerial parts of Cichorium intybus L. Chromatographia 2001, 54, 455–461. [Google Scholar] [CrossRef]
- Said, R.B.; Hamed, A.I.; Mahalel, U.A.; Al-Ayed, A.S.; Kowalczyk, M.; Moldoch, J.; Oleszek, W.; Stochmal, A. Tentative characterization of polyphenolic compounds in the male flowers of Phoenix dactylifera by liquid chromatography coupled with mass spectrometry and DFT. Int. J. Mol. Sci. 2017, 18, 512. [Google Scholar] [CrossRef]
- Mahomoodally, M.F.; Sieniawska, E.; Sinan, K.I.; Nancy Picot-Allain, M.C.; Yerlikaya, S.; Cengiz Baloglu, M.; Altunoglu, Y.C.; Senkardes, I.; Rengasamy, K.R.; Zengin, G. Utilisation of Rhododendron luteum Sweet bioactive compounds as valuable source of enzymes inhibitors, antioxidant, and anticancer agents. Food Chem. Toxicol. 2020, 135, 111052. [Google Scholar] [CrossRef]
- Tardugno, R.; Pozzebon, M.; Beggio, M.; Del Turco, P.; Pojana, G. Polyphenolic profile of Cichorium intybus L. endemic varieties from the Veneto region of Italy. Food Chem. 2018, 266, 175–182. [Google Scholar] [CrossRef]
- Shumova, G.; Savelieva, E.V.; Vladymyrova, I.M.; Tishakova, T.S. Phenolic compound composition of herb of Pulsatilla pratensis (L.) Mill. Vet. Sci. Pharm. 2017, 4, 35–38. [Google Scholar]
- Zhao, Z.; Nian, M.; Qiao, H.; Li, B.; Zheng, X. Pulsatilla chinensis: A review of traditional uses, phytochemistry and pharmacology research progress. Arab. J. Chem. 2021, 14, 103403. [Google Scholar] [CrossRef]
- Hamilton, K.L.; Sheehan, S.A.; Retzbach, E.P.; Timmerman, C.A.; Gianneschi, G.B.; Tempera, P.J.; Balachandran, P.; Goldberg, G.S. Effects of Maackia amurensis seed lectin (MASL) on oral squamous cell carcinoma (OSCC) gene expression and transcriptional signaling pathways. J. Cancer Res. Clin. Oncol. 2021, 147, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Allard, S.T.M.; Kopish, K. Luciferase reporter assays: Powerful, adaptable tools for cell biology research. Cell Notes 2008, 21, 23–26. [Google Scholar]
- Allard, S.T.M. Bioluminescent Reporter Genes. Adv. Biochem. 2008, 54, 350–353. [Google Scholar]
- Cappellini, A.; Tabellini, G.; Zweyer, M.; Bortul, R.; Tazzari, P.L.; Billi, A.M.; Fala, F.; Cocco, L.; Martelli, A.M. The phosphoinositide 3-kinase/Akt pathways regulates cell cycle progression of HL60 human leukemia cells through cytoplasmic relocalization of the cyclin-dependent kinase inhibitor p27 (Kip1) and control of cyclin D1 expression. Leukemia 2003, 17, 2157–2167. [Google Scholar] [CrossRef]
- Burgering, B.M.; Kops, G.J. Cell cycle and death control: Long live Forkheads. Trends Biochem. Sci. 2002, 27, 352–360. [Google Scholar] [CrossRef]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef]
- Tang, T.T.; Dowbenko, D.; Jackson, A.; Toney, L.; Lewin, D.A.; Dent, A.L.; Lasky, L.A. The forkhead transcription factor AFX activates apoptosis by induction of the BCL-6 transcriptional repressor. J. Biol. Chem. 2002, 277, 14255–14265. [Google Scholar] [CrossRef]
- Dijkers, P.F.; Medema, R.H.; Lammers, J.W.; Koenderman, L.; Coffer, P.J. Expression of the pro-apoptotic Bcl-2 family member Bim is regulated by the forkhead transcription factor FKHR-L1. Curr. Biol. 2000, 10, 1201–1204. [Google Scholar] [CrossRef]
- Gilley, J.; Coffer, P.J.; Ham, J. FOXO transcription factors directly activate bim gene expression and promote apoptosis in sympathetic neurons. J. Cell Biol. 2003, 162, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, P.; Osipo, C.; Foreman, K.; Golde, T.; Osborne, B.; Miele, L. Rational targeting of Notch signaling in cancer. Oncogene 2008, 27, 5124–5131. [Google Scholar] [CrossRef]
- Cavin, L.G.; Wang, F.; Factor, V.M.; Kaur, S.; Venkatraman, M.; Thorgeirsson, S.S.; Arsura, M. Transforming growth factoralpha inhibits the intrinsic pathway of c-Myc-induced apoptosis through activation of nuclear factor-kappaB in murine hepatocellular carcinomas. Mol. Cancer Res. 2005, 3, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Z.; Tsai, S.Y.; Leone, G. Emerging roles of E2Fs in cancer: An exit from cell cycle control. Nat. Rev. Cancer 2009, 9, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Viatour, P.; Ehmer, U.; Saddic, L.A.; Dorrell, C.; Andersen, J.B.; Lin, C.; Zmoos, A.F.; Mazur, P.K.; Schaffer, B.E.; Ostermeier, A.; et al. Notch signaling inhibits hepatocellular carcinoma following inactivation of the RB pathway. J. Exp. Med. 2011, 208, 1963–1976. [Google Scholar] [CrossRef] [PubMed]
- Chanvorachote, P.; Sriratanasak, N.; Nonpanya, N. C-myc contributes to malignancy of lung cancer. A potential anticancer drug target. Anticancer Res. 2020, 40, 609–618. [Google Scholar] [CrossRef]
- Katoh, M. Regulation of WNT3 and WNT3A mRNAs in human cancer cell lines NT2, MCF-7, and MKN45. Int. J. Oncol. 2002, 20, 373–377. [Google Scholar] [CrossRef]
- Maubant, S.; Tesson, B.; Maire, V.; Ye, M.; Rigaill, G.; Gentien, D.; Cruzalegui, F.; Tucker, G.C.; Roman-Roman, S.; Dubois, T. Transcriptome Analysis of Wnt3a-Treated Triple-Negative Breast Cancer Cells. PLoS ONE 2015, 10, e0122333. [Google Scholar] [CrossRef]
- Zheng, J.; Xue, H.; Wang, T.; Jiang, Y.; Liu, B.; Li, J.; Liu, Y.; Wang, W.; Zhang, B.; Sun, M. MiR-21 downregulates the tumor suppressor P12 CDK2AP1 and stimulates cell proliferation and invasion. J. Cell. Biochem. 2011, 112, 872–880. [Google Scholar] [CrossRef]
- Asangani, I.A.; Rasheed, S.A.K.; Nikolova, D.A.; Leupold, J.H.; Colburn, N.H.; Post, S.; Allgayer, H. MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal Cancer. Oncogene 2008, 27, 2128–2136. [Google Scholar] [CrossRef]
- Medema, R.H.; Kops, G.J.; Bos, J.L.; Burgering, B.M. AFX-like Forkhead transcription factors mediate cell-cycle regulation by Ras and PKB through p27kip1. Nature 2000, 404, 782–787. [Google Scholar] [CrossRef]
- Nouri, M.; Massah, S.; Caradec, J.; Lubik, A.A.; Li, N.; Truong, S.; Lee, A.R.; Fazli, L.; Ramnarine, V.R.; Lovnicki, J.M.; et al. Transient Sox9 exspression facilitates resistance to androgen-targeted therapy in prostate cancer. Clin. Cancer Res. 2020, 26, 1678–1689. [Google Scholar] [CrossRef] [PubMed]
- Vlahopoulos, S.A. Aberrant control of NF-κB in cancer permits transcriptional and phenotypic plasticity, to curtail dependence on host tissue: Molecular mode. Cancer Biol. Med. 2017, 14, 254–270. [Google Scholar] [CrossRef] [PubMed]
- Tewari, D.; Nabavi, S.F.; Nabavi, S.M.; Sureda, A.; Farooqi, A.A.; Atanasov, A.G.; Vacca, R.A.; Sethi, G.; Bishayee, A. Targeting activator protein 1 signaling pathway by bioactive natural agents: Possible therapeutic strategy for cancer prevention and intervention. Pharmacol. Res. 2018, 128, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Massagué, J. TGF-beta in cancer. Cell 2008, 134, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Skoda, A.M.; Simovic, D.; Karin, V.; Kardum, V.; Vranic, S.; Serman, L. The role of the Hedgehog signaling pathway in cancer: A comprehensive review. Bosn. J. Basic Med. Sci. 2018, 18, 8–20. [Google Scholar] [CrossRef]
- Qi, T.; Qu, Q.; Li, G.; Wang, J.; Zhu, H.; Yang, Z.; Sun, Y.; Lu, Q.; Qu, J. Function and regulation of the PEA3 subfamily of ETS transcription factors in cancer. Am. J. Cancer Res. 2020, 10, 3083–3105. [Google Scholar]
- Oikawa, T.; Yamada, T. Molecular biology of the Ets family of transcription factors. Gene 2003, 303, 11–34. [Google Scholar] [CrossRef]
- Hollenhorst, P.C.; McIntosh, L.P.; Graves, B.J. Genomic and biochemical insights into the specificity of ETS transcription factors. Annu. Rev. Biochem. 2011, 80, 437–471. [Google Scholar] [CrossRef]
- Tkach, M.; Rosemblit, C.; Rivas, M.A.; Proietti, C.J.; Díaz Flaqué, M.C.; Mercogliano, M.F.; Beguelin, W.; Maronna, E.; Guzmán, P.; Gercovich, F.G.; et al. P42/p44 MAPK-mediated Stat3Ser727 phosphorylation is required for progestin-induced full activation of Stat3 and breast cancer growth. Endocr.-Relat. Cancer 2013, 20, 197–212. [Google Scholar] [CrossRef]
- Klampfer, L. Signal transducers and activators of transcription (STATs): Novel targets of chemopreventive and chemotherapeutic drugs. Curr. Cancer Drug Targets 2006, 6, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, H.; Naseri, G.; Rezaee, R.; Mohammadi, M.; Banikazemi, Z.; Mirzaei, H.R.; Salehi, H.; Peyvandi, M.; Pawelek, J.M.; Sahebkar, A. Curcumin: A new candidate for melanoma therapy? Int. J. Cancer 2016, 139, 1683–1695. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Mandal, D.; Saha, B.; Sen, G.S.; Das, T.; Sa, G. Curcumin prevents tumor-induced T cell apoptosis through Stat-5a-mediated Bcl-2 induction. BioMed Cent. J. Biol. Chem. 2007, 282, 15954e64. [Google Scholar] [CrossRef] [PubMed]
- Terlikowska, K.; Witkowska, A.; Zujko, M.; Dobrzycka, B.; Terlikowski, S. Potential application of curcumin and its analogues in the treatment strategy of patients with primary epithelial ovarian Cancer. Int. J. Mol. Sci. 2014, 15, 21703–21722. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Rane, G.; Kanchi, M.M.; Arfuso, F.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; Tan, B.K.H.; Kumar, A.P.; Sethi, G. The multifaceted role of curcumin in cancer prevention and treatment. Molecules 2015, 20, 2728–2769. [Google Scholar] [CrossRef]
- Wondrak, G.T. Redox-directed cancer therapeutics: Molecular mechanisms and opportunities. Antioxid. Redox Signal. 2009, 11, 3013–3069. [Google Scholar] [CrossRef]
- Snezhkina, V.A.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS generation and antioxidant defense systems in normal and malignant cells. Oxid. Med. Cell. Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef]
- Son, M.K.; Jung, K.H.; Lee, H.S.; Lee, H.; Kim, S.J.; Yan, H.H.; Ryu, Y.L.; Hong, S.S. SB365, Pulsatilla saponin D suppresses proliferation and induces apoptosis of pancreatic cancer cells. Oncol. Rep. 2013, 30, 801–808. [Google Scholar] [CrossRef]
- Landry, J.J.M.; Pyl, P.T.; Rausch, T.; Zichner, T.; Tekkedil, M.M.; Stütz, A.M.; Jauch, A.; Aiyar, R.S.; Pau, G.; Delhomme, N.; et al. The genomic and transcriptomic landscape of a HeLa cell line. G3 2013, 3, 1213–1224. [Google Scholar] [CrossRef]
- Adey, A.; Burton, J.N.; Kitzman, J.O.; Hiatt, J.B.; Lewis, A.P.; Martin, B.K.; Qiu, R.; Lee, C.; Shendure, J. The haplotype-resolved genome and epigenome of the aneuploid HeLa cancer cell line. Nature 2013, 500, 207–211. [Google Scholar] [CrossRef]
- Forment, J.V.; Abderrahmane, K.; Jackson, S.P. Chromothrypsis and cancer: Causes and consequences of chromosome shattering. Nat. Rev. Cancer 2012, 12, 663–670. [Google Scholar] [CrossRef]
- Korbel, J.O.; Campbell, P.J. Criteria for inference of chromothripsis in cancer genomes. Cell 2013, 152, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.W.; Jung, K.H.; Lee, H.S.; Son, M.K.; Yan, H.H.; Kang, N.S.; Lee, J.; Hong, S.S. SB365, Pulsatilla saponin D, targets c-Met and exerts antiangiogenic and antitumor activities. Carcinogenesis 2013, 34, 2156–2169. [Google Scholar] [CrossRef] [PubMed]
- Zając, A.; Zając, M. Distribution Atlas of Vascular Plants in Poland, 1st ed.; Laboratory of Computer Chorology, Institute of Botany, Jagiellonian University: Cracow, Poland, 2001; pp. 1–319. [Google Scholar]
- Den Virtuella Floran—Naturhistoriska Riksmuseet. 1996. Available online: http://linnaeus.nrm.se/flora/di (accessed on 1 August 2017).
- Zengin, G.; Sieniawska, E.; Senkardes, I.; Picot-Allain, M.C.N.; Sinan, I.; Fawzi, K.; Mahomoodally, M. Antioxidant abilities, key enzyme inhibitory potential and phytochemical profile of Tanacetum poteriifolium Grierson. Ind. Crops Prod. 2019, 140, 111629. [Google Scholar] [CrossRef]
- Łaska, G.; Sieniawska, E.; Świątek, Ł.; Zjawiony, J.; Khan, S.; Boguszewska, A.; Stocki, M.; Angielczyk, M.; Polz-Dacewicz, M. Phytochemistry and biological activities of Polemonium caeruleum L. Phytochem. Lett. 2019, 30, 314–323. [Google Scholar] [CrossRef]






| Cytotoxic Activity (CC50 μg/mL) 1 | |||||||
|---|---|---|---|---|---|---|---|
| Mammalian Cell Lines | Normal Cell Lines | Cancer Cell Lines | |||||
| Sample Name | VERO | LLC-PK1 | HeLa | SK-OV-3 | KB | SK-MEL | BT-549 |
| PVR | 39 | 42 | 31 | 35 | 42 | 44 | 57 |
| PVL | 52 | 70 | 50 | 60 | 73 | 60 | 68 |
| Doxorubicin 2 | >5 | 1.6 | 3.9 | 2.3 | 1.7 | 1.7 | 2.2 |
| Extracts | CC50 1 µg/mL (±SD) | Statistical Significance | SI | |
|---|---|---|---|---|
| VERO | HeLa | |||
| PVR | 188.3 ± 7.11 | 71.87 ± 0.79 | ** | 2.62 |
| PVL | 17.57 ± 1.77 | 21.88 ± 1.73 | ns | 0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łaska, G.; Sieniawska, E.; Maciejewska-Turska, M.; Świątek, Ł.; Pasco, D.S.; Balachandran, P. Pulsatilla vulgaris Inhibits Cancer Proliferation in Signaling Pathways of 12 Reporter Genes. Int. J. Mol. Sci. 2023, 24, 1139. https://doi.org/10.3390/ijms24021139
Łaska G, Sieniawska E, Maciejewska-Turska M, Świątek Ł, Pasco DS, Balachandran P. Pulsatilla vulgaris Inhibits Cancer Proliferation in Signaling Pathways of 12 Reporter Genes. International Journal of Molecular Sciences. 2023; 24(2):1139. https://doi.org/10.3390/ijms24021139
Chicago/Turabian StyleŁaska, Grażyna, Elwira Sieniawska, Magdalena Maciejewska-Turska, Łukasz Świątek, David S. Pasco, and Premalatha Balachandran. 2023. "Pulsatilla vulgaris Inhibits Cancer Proliferation in Signaling Pathways of 12 Reporter Genes" International Journal of Molecular Sciences 24, no. 2: 1139. https://doi.org/10.3390/ijms24021139
APA StyleŁaska, G., Sieniawska, E., Maciejewska-Turska, M., Świątek, Ł., Pasco, D. S., & Balachandran, P. (2023). Pulsatilla vulgaris Inhibits Cancer Proliferation in Signaling Pathways of 12 Reporter Genes. International Journal of Molecular Sciences, 24(2), 1139. https://doi.org/10.3390/ijms24021139