

The Effects of Combined Exposure to Bisphenols and Perfluoroalkyls on Human Perinatal Stem Cells and the Potential Implications for Health Outcomes

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
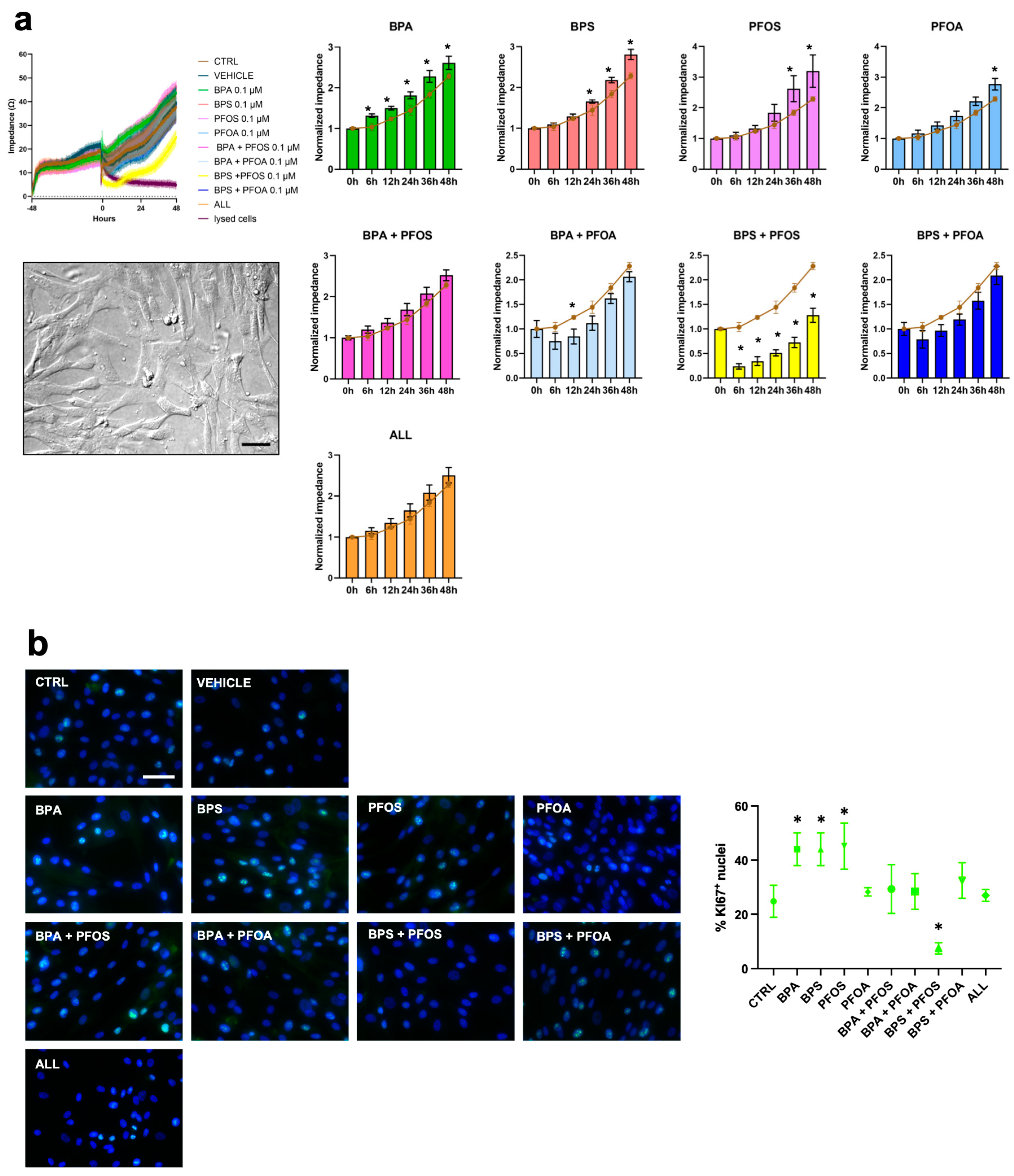
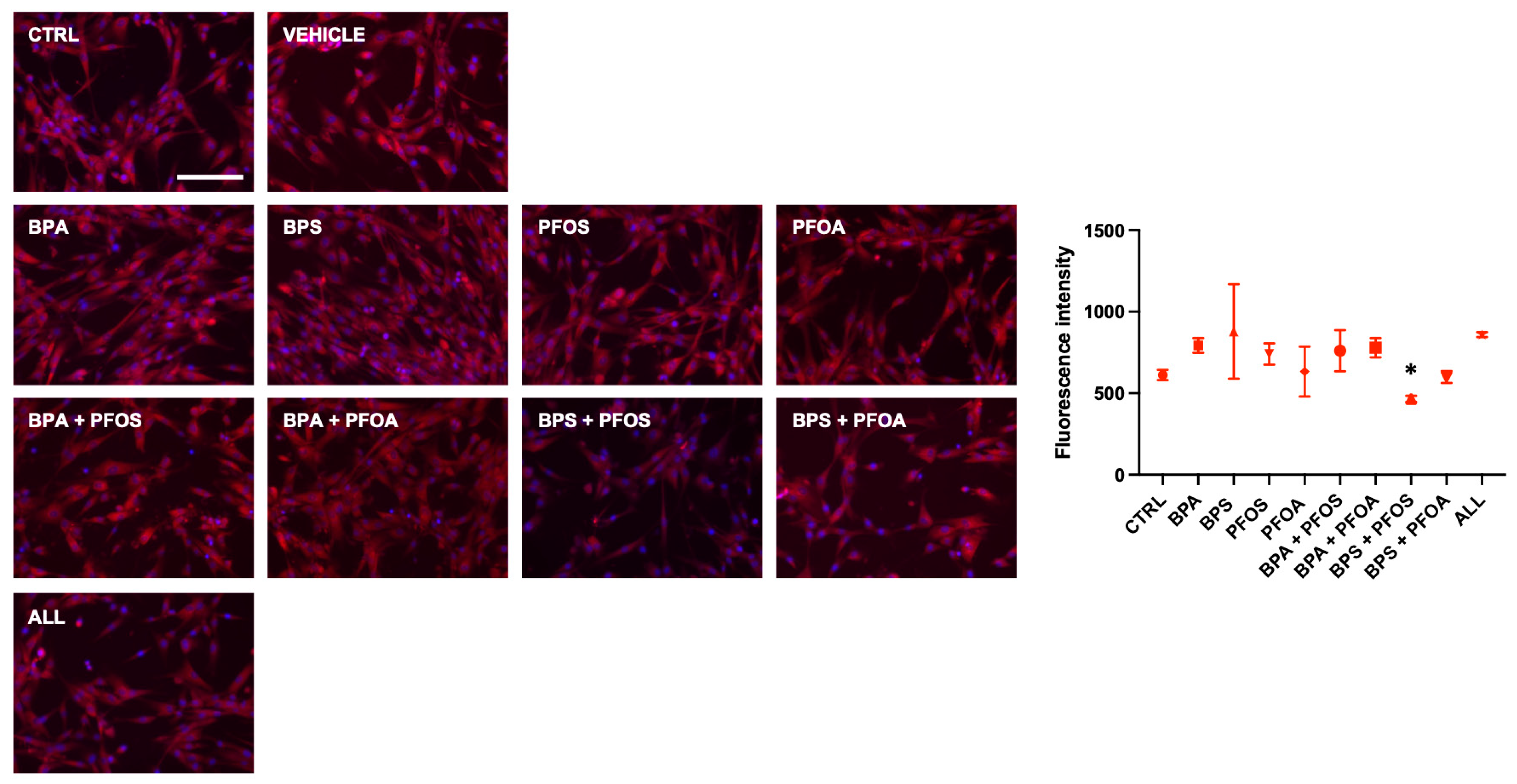
2.1. BPs and PFs Effects on hFM-MSCs Biology
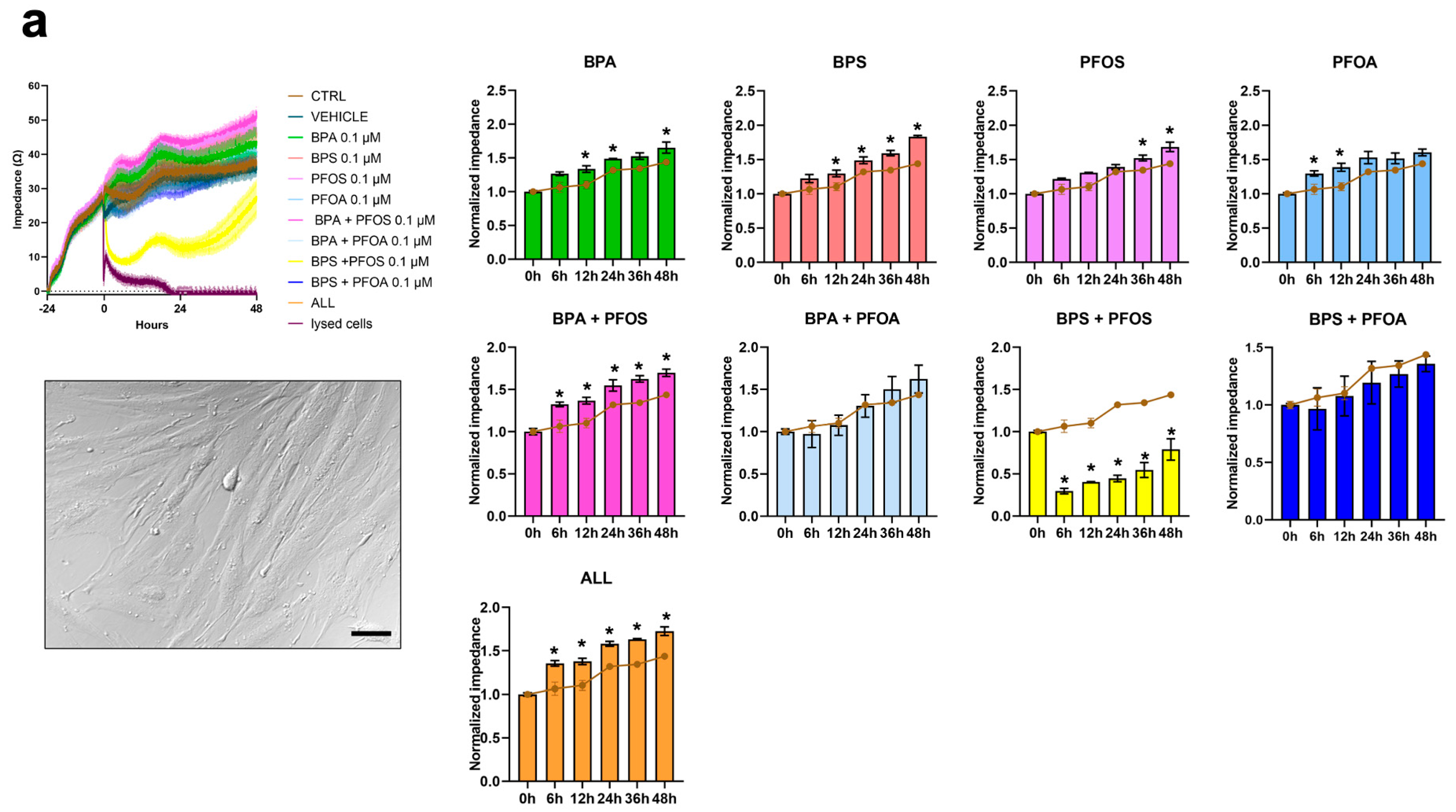
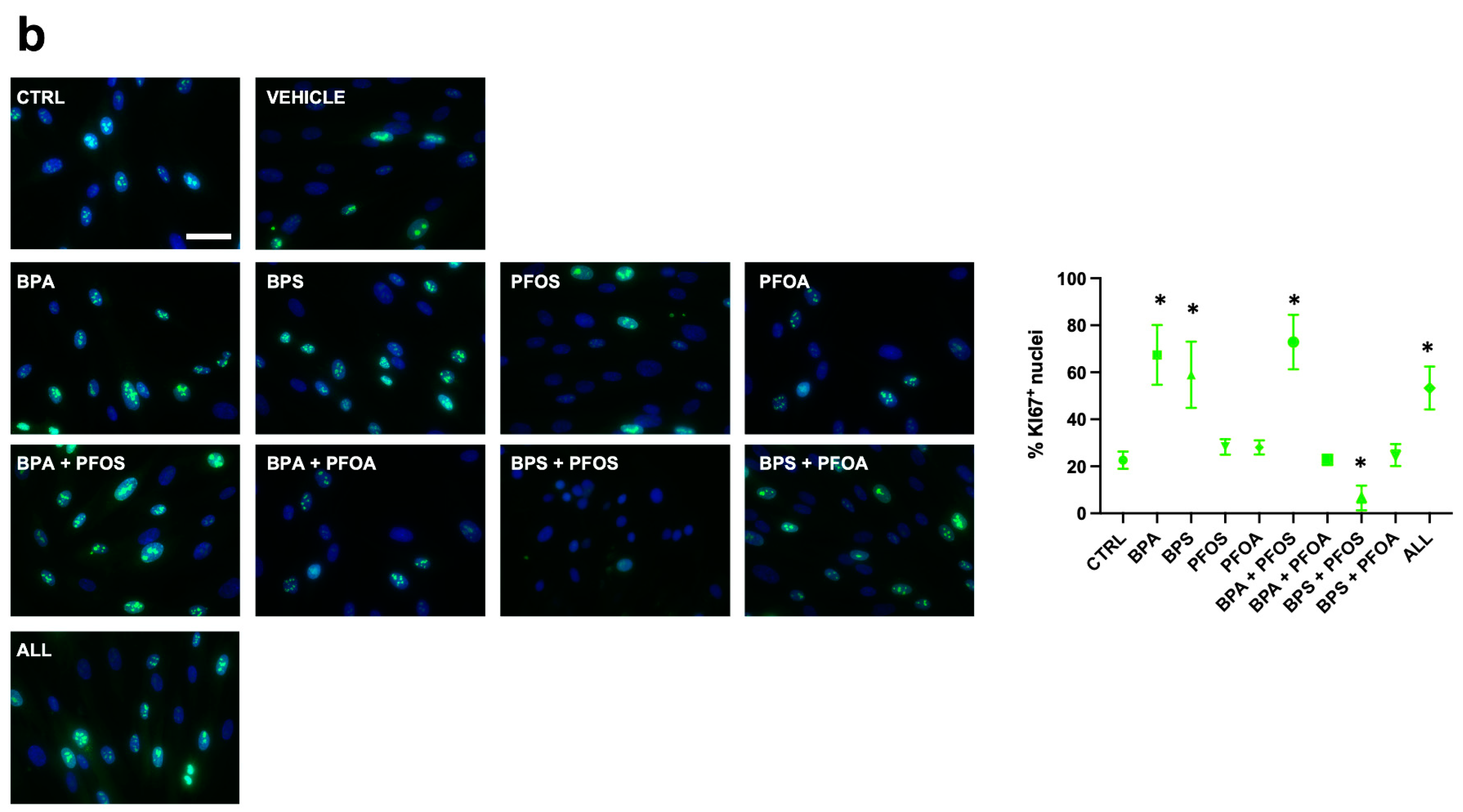
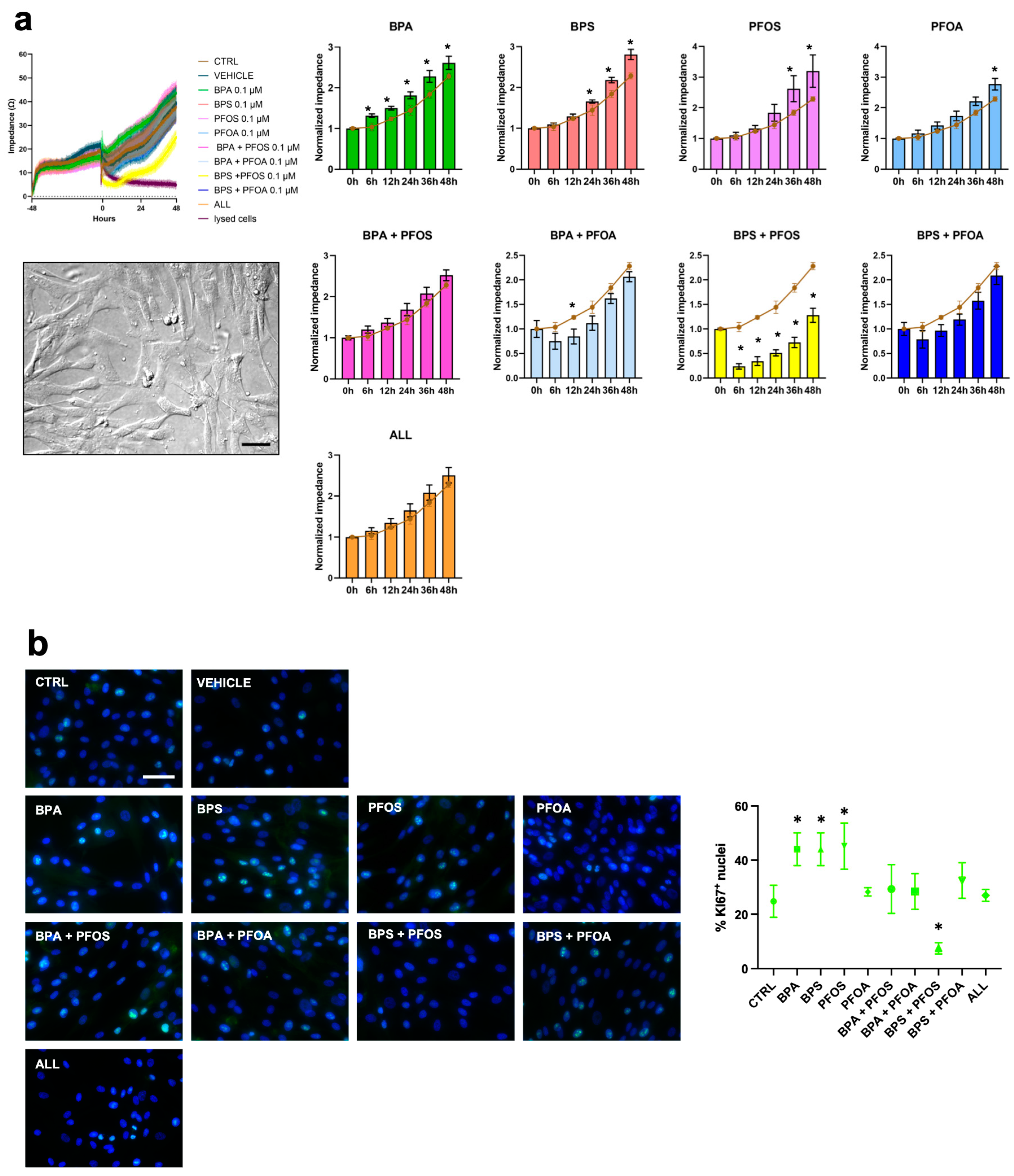
2.1.1. Proliferation and Cytotoxicity
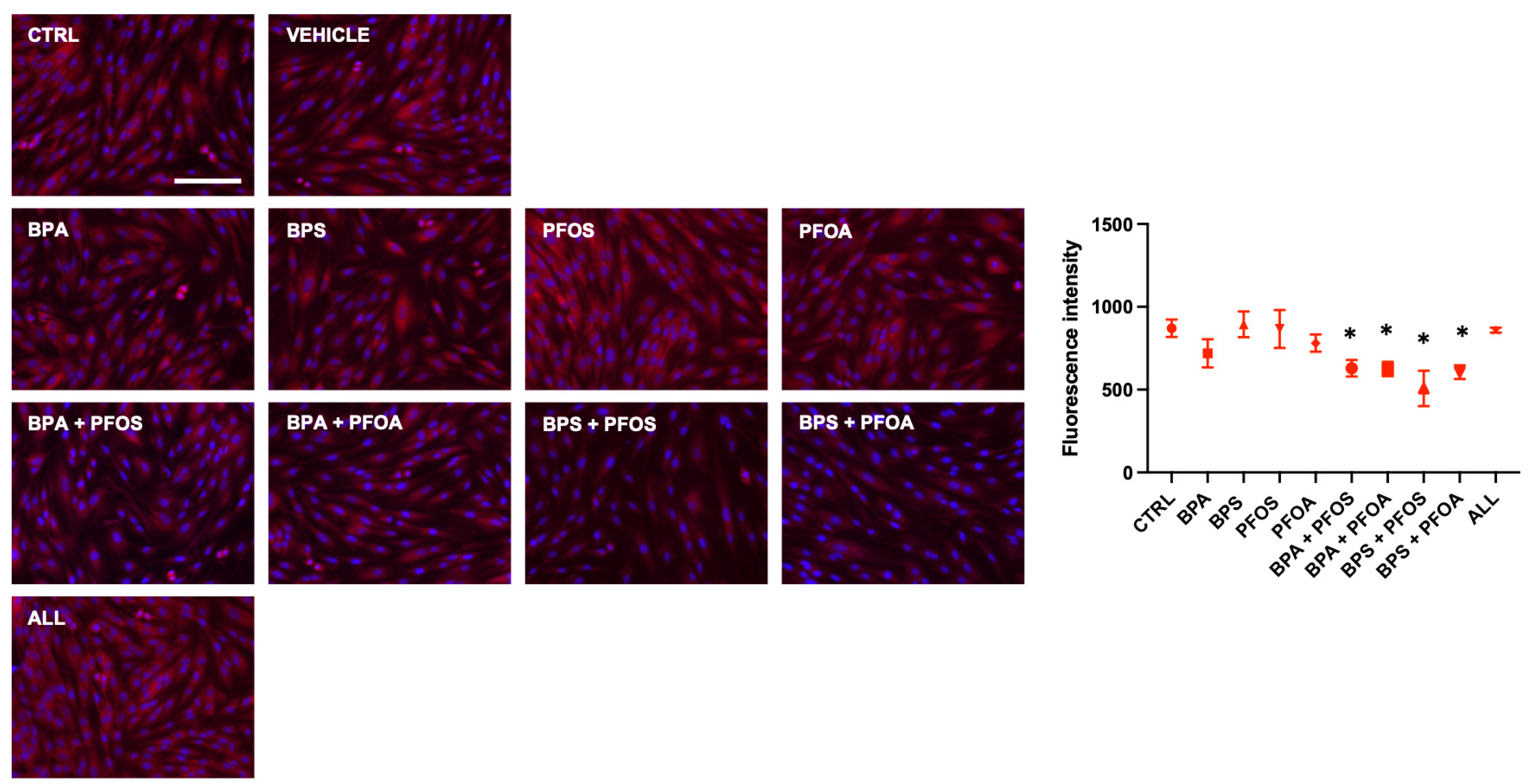
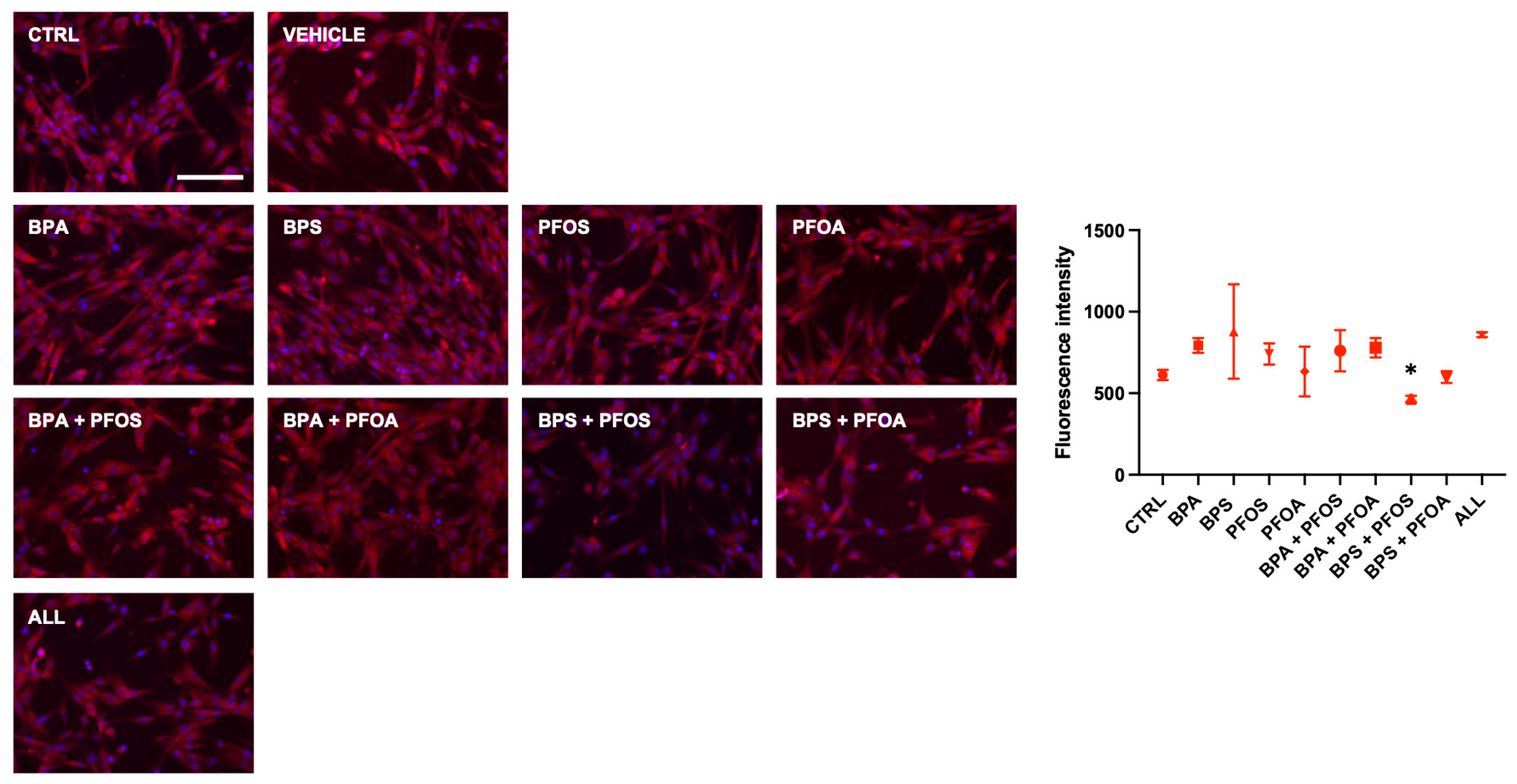
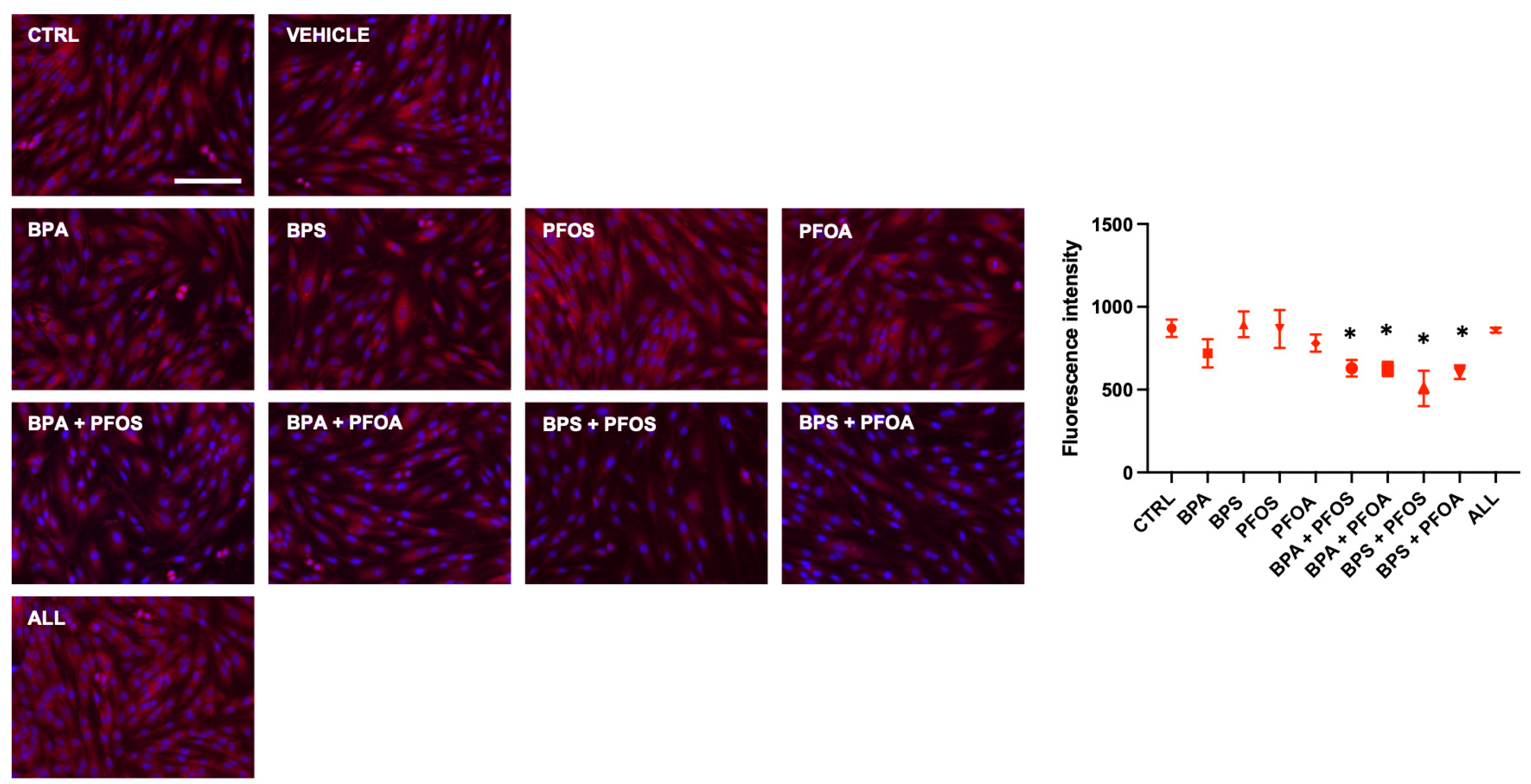
2.1.2. BP and PF Effects on hFM-MSCs Biology: Effects on Mitochondrial Membrane Potential
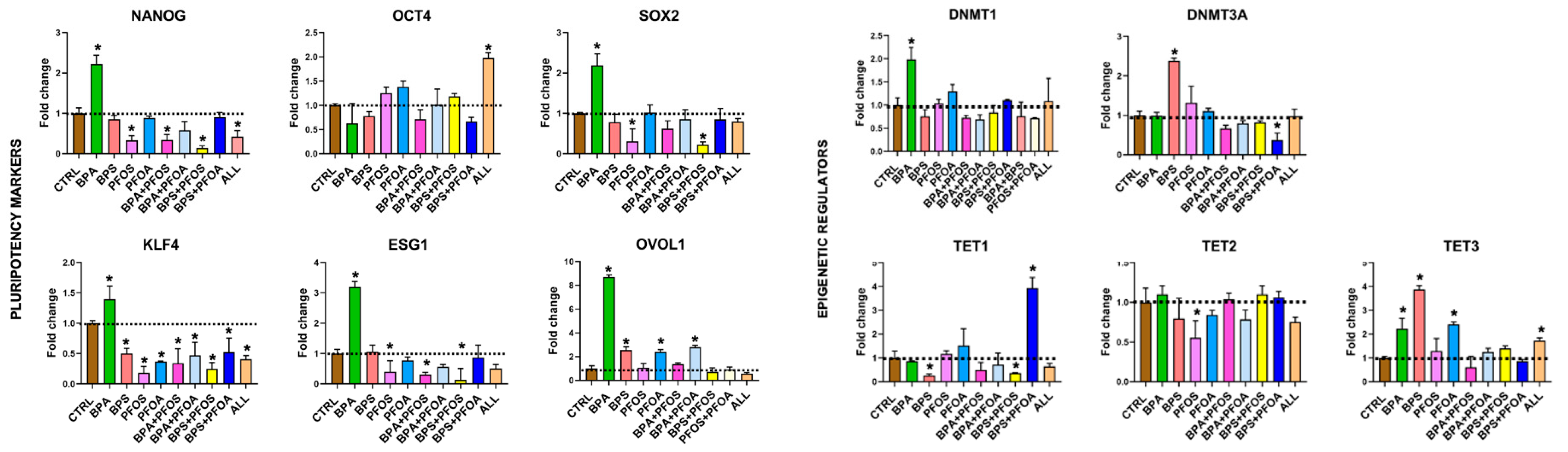
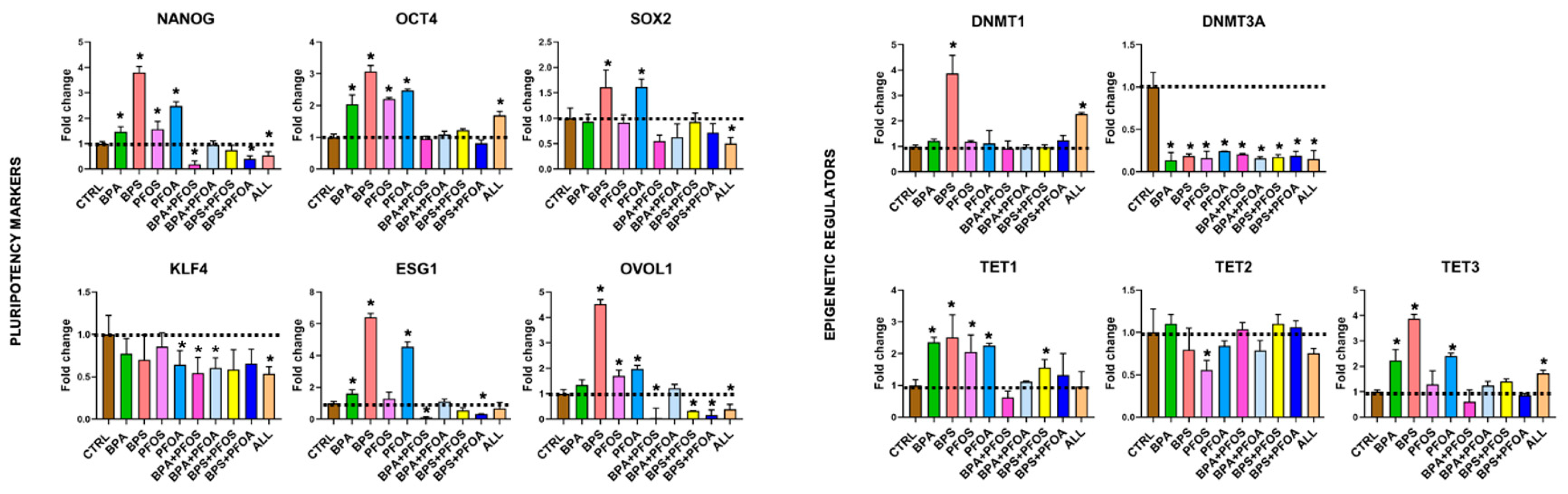
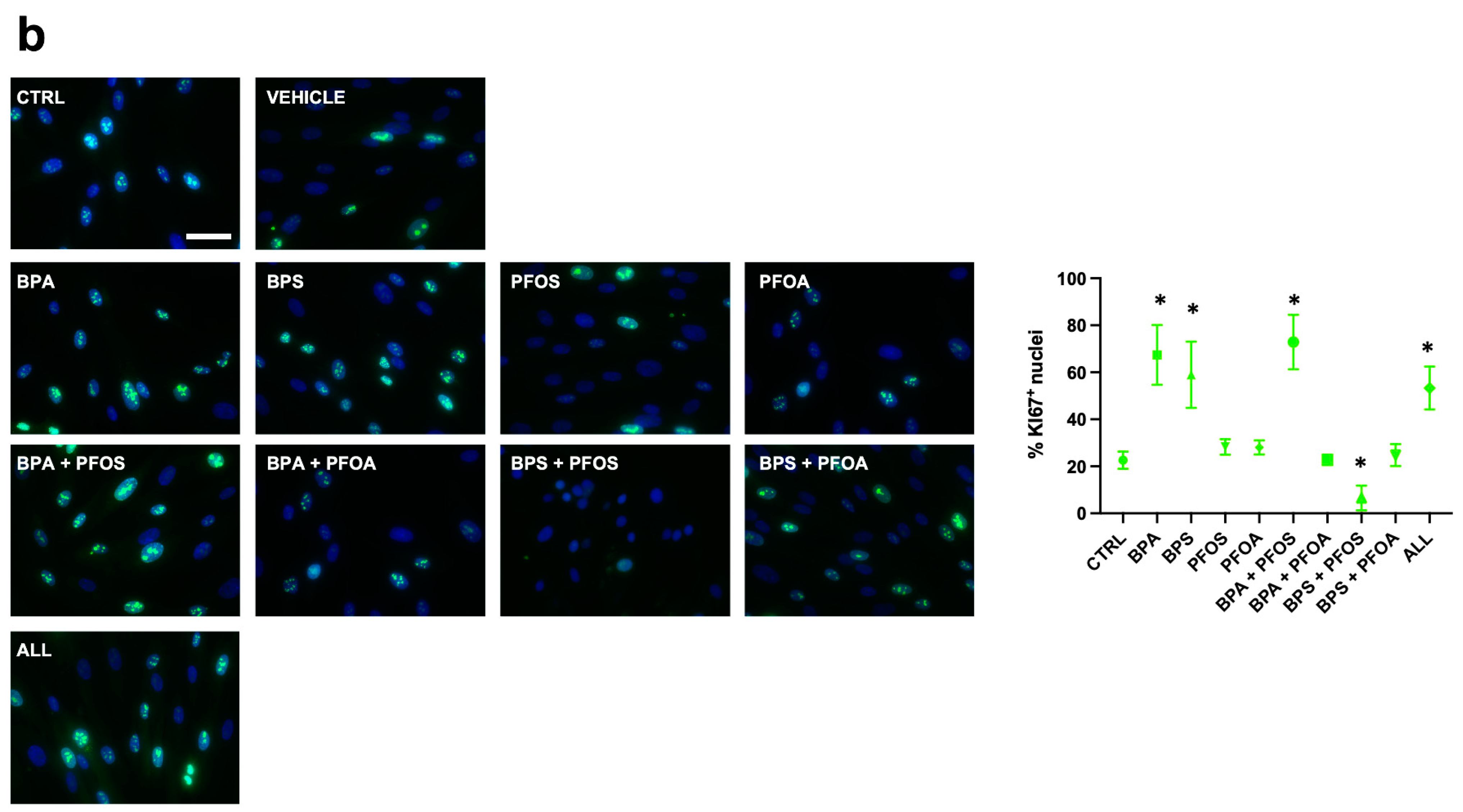
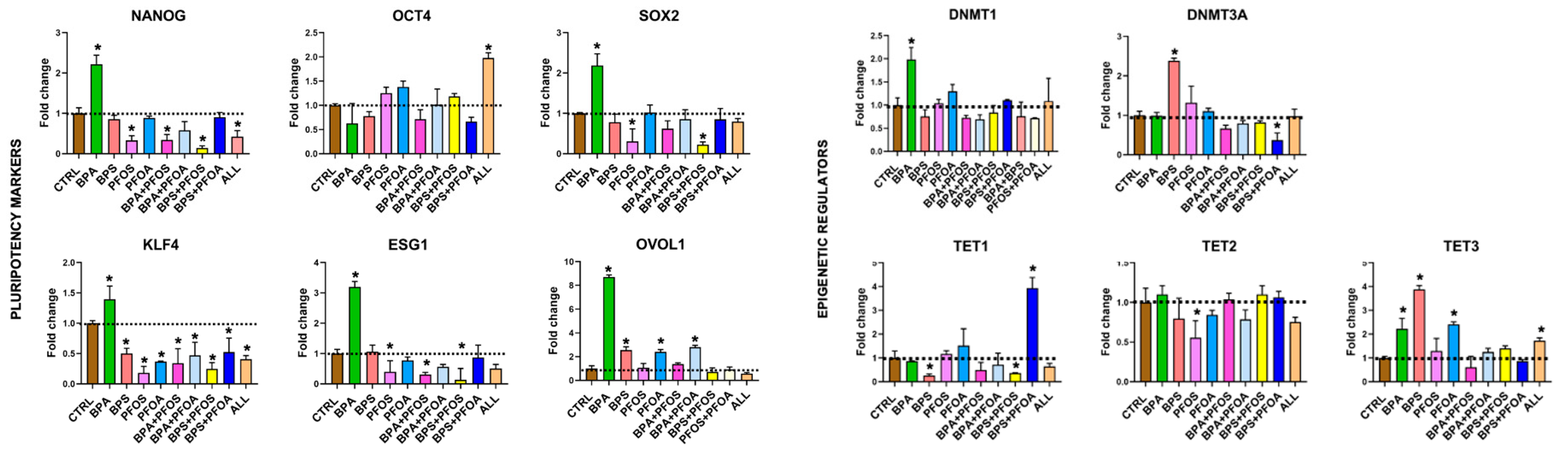
2.1.3. Consequences on Stemness Markers Profile
2.1.4. Modifications of Epigenetic Regulatory Enzymes
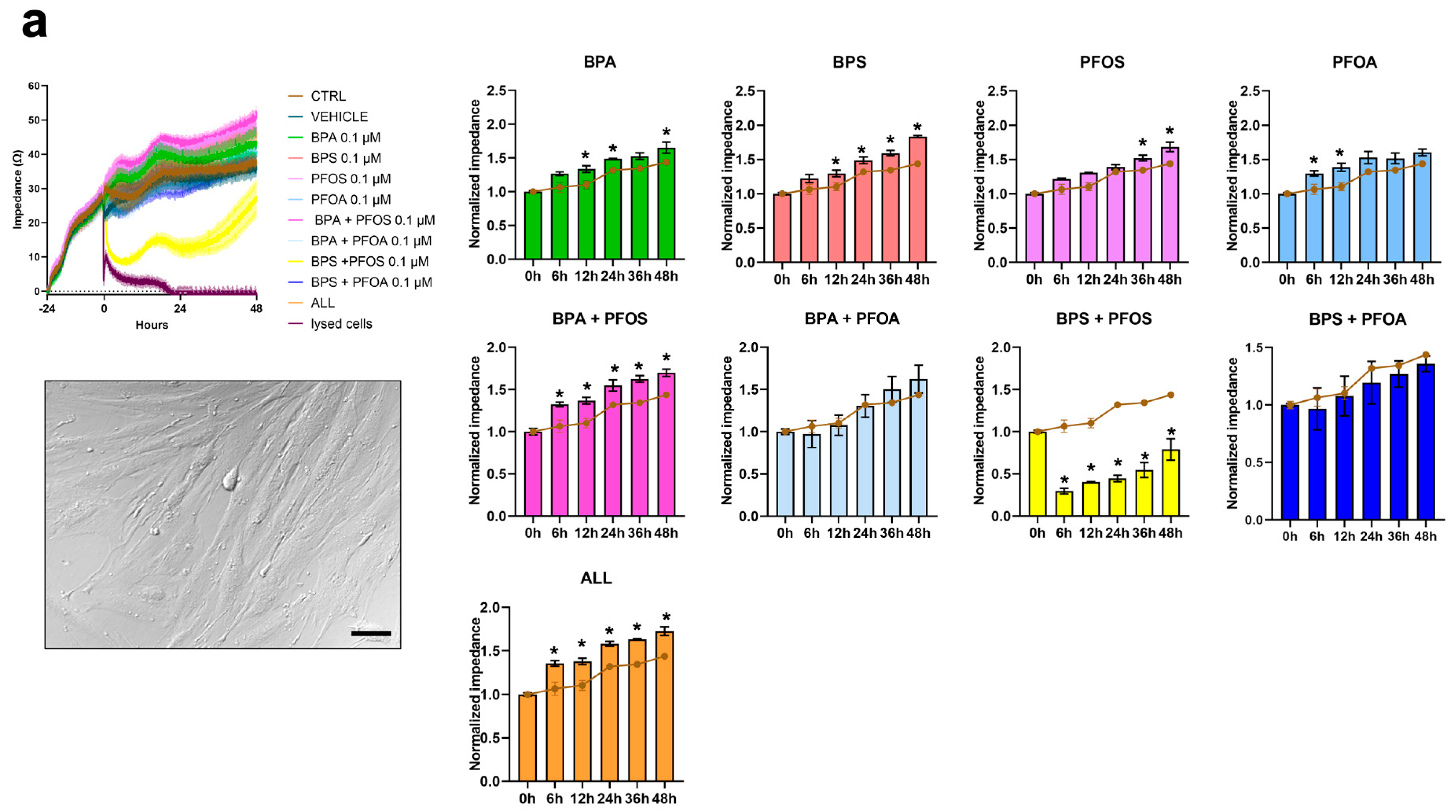
2.2. BPs and PFs Effects on hAFSCs Biology
2.2.1. Proliferation and Cytotoxicity
2.2.2. Effects on Mitochondrial Membrane Potential
2.2.3. Consequences on Stemness Markers Profile
2.2.4. Modifications of Epigenetic Regulatory Enzymes
3. Discussion
Study Limitations
4. Materials and Methods
4.1. Human Cell Cultures and Treatments
4.2. Analysis of Impedance
4.3. Immunofluorescence Analysis of Ki67 Expression
4.4. Immunofluorescence Analysis of Mitochondrial Membrane Potential (MMP)
4.5. RNA Extraction and Reverse Transcription
4.6. Real-Time PCR (qPCR)
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Diamanti-Kandarakis, E.; Bourguignon, J.-P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-Disrupting Chemicals: An Endocrine Society Scientific Statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef]
- Rochester, J.R.; Bolden, A.L. Bisphenol S and F: A Systematic Review and Comparison of the Hormonal Activity of Bisphenol A Substitutes. Environ. Health Perspect. 2015, 123, 643–650. [Google Scholar] [CrossRef]
- Kitamura, S. Comparative Study of the Endocrine-Disrupting Activity of Bisphenol A and 19 Related Compounds. Toxicol. Sci. 2005, 84, 249–259. [Google Scholar] [CrossRef]
- Maqbool, F.; Mostafalou, S.; Bahadar, H.; Abdollahi, M. Review of Endocrine Disorders Associated with Environmental Toxicants and Possible Involved Mechanisms. Life Sci. 2016, 145, 265–273. [Google Scholar] [CrossRef]
- Behr, A.-C.; Lichtenstein, D.; Braeuning, A.; Lampen, A.; Buhrke, T. Perfluoroalkylated Substances (PFAS) Affect Neither Estrogen and Androgen Receptor Activity nor Steroidogenesis in Human Cells in Vitro. Toxicol. Lett. 2018, 291, 51–60. [Google Scholar] [CrossRef]
- Shafei, A.; Ramzy, M.M.; Hegazy, A.I.; Husseny, A.K.; EL-hadary, U.G.; Taha, M.M.; Mosa, A.A. The Molecular Mechanisms of Action of the Endocrine Disrupting Chemical Bisphenol A in the Development of Cancer. Gene 2018, 647, 235–243. [Google Scholar] [CrossRef]
- Miranda, A.; Sousa, N. Maternal Hormonal Milieu Influence on Fetal Brain Development. Brain Behav. 2018, 8, e00920. [Google Scholar] [CrossRef]
- Kelley, A.S.; Banker, M.; Goodrich, J.M.; Dolinoy, D.C.; Burant, C.; Domino, S.E.; Smith, Y.R.; Song, P.X.K.; Padmanabhan, V. Early Pregnancy Exposure to Endocrine Disrupting Chemical Mixtures Are Associated with Inflammatory Changes in Maternal and Neonatal Circulation. Sci. Rep. 2019, 9, 5422. [Google Scholar] [CrossRef]
- Horikoshi, T.; Nishimura, T.; Nomura, Y.; Iwabuchi, T.; Itoh, H.; Takizawa, T.; Tsuchiya, K.J. Umbilical Cord Serum Concentrations of Perfluorooctane Sulfonate, Perfluorooctanoic Acid, and the Body Mass Index Changes from Birth to 5 1/2 Years of Age. Sci. Rep. 2021, 11, 19789. [Google Scholar] [CrossRef]
- Gerona, R.R.; Woodruff, T.J.; Dickenson, C.A.; Pan, J.; Schwartz, J.M.; Sen, S.; Friesen, M.W.; Fujimoto, V.Y.; Hunt, P.A. Bisphenol-A (BPA), BPA Glucuronide, and BPA Sulfate in Midgestation Umbilical Cord Serum in a Northern and Central California Population. Environ. Sci. Technol. 2013, 47, 12477–12485. [Google Scholar] [CrossRef]
- Li, A.; Zhuang, T.; Shi, W.; Liang, Y.; Liao, C.; Song, M.; Jiang, G. Serum Concentration of Bisphenol Analogues in Pregnant Women in China. Sci. Total Environ. 2020, 707, 136100. [Google Scholar] [CrossRef] [PubMed]
- Adu-Gyamfi, E.A.; Rosenfeld, C.S.; Tuteja, G. The Impact of Bisphenol A on the Placenta. Biol. Reprod. 2022, 106, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Berkane, N.; Liere, P.; Oudinet, J.-P.; Hertig, A.; Lefèvre, G.; Pluchino, N.; Schumacher, M.; Chabbert-Buffet, N. From Pregnancy to Preeclampsia: A Key Role for Estrogens. Endocr. Rev. 2017, 38, 123–144. [Google Scholar] [CrossRef]
- Braun, J.M.; Hauser, R. Bisphenol A and Children’s Health. Curr. Opin. Pediatr. 2011, 23, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Segovia-Mendoza, M.; Palacios-Arreola, M.I.; Monroy-Escamilla, L.M.; Soto-Piña, A.E.; Nava-Castro, K.E.; Becerril-Alarcón, Y.; Camacho-Beiza, R.; Aguirre-Quezada, D.E.; Cardoso-Peña, E.; Amador-Muñoz, O.; et al. Association of Serum Levels of Plasticizers Compounds, Phthalates and Bisphenols, in Patients and Survivors of Breast Cancer: A Real Connection? Int. J. Environ. Res. Public Health 2022, 19, 8040. [Google Scholar] [CrossRef]
- Gaggi, G.; Izzicupo, P.; Di Credico, A.; Sancilio, S.; Di Baldassarre, A.; Ghinassi, B. Spare Parts from Discarded Materials: Fetal Annexes in Regenerative Medicine. Int. J. Mol. Sci. 2019, 20, 1573. [Google Scholar] [CrossRef]
- Gaggi, G.; Di Credico, A.; Izzicupo, P.; Antonucci, I.; Crescioli, C.; Di Giacomo, V.; Di Ruscio, A.; Amabile, G.; Alviano, F.; Di Baldassarre, A.; et al. Epigenetic Features of Human Perinatal Stem Cells Redefine Their Stemness Potential. Cells 2020, 9, 1304. [Google Scholar] [CrossRef] [PubMed]
- Di Baldassarre, A.; D’Amico, M.A.; Izzicupo, P.; Gaggi, G.; Guarnieri, S.; Mariggiò, M.A.; Antonucci, I.; Corneo, B.; Sirabella, D.; Stuppia, L.; et al. Cardiomyocytes Derived from Human CardiopoieticAmniotic Fluids. Sci. Rep. 2018, 8, 12028. [Google Scholar] [CrossRef]
- Gaggi, G.; Di Credico, A.; Izzicupo, P.; Alviano, F.; Di Mauro, M.; Di Baldassarre, A.; Ghinassi, B. Human Mesenchymal Stromal Cells Unveil an Unexpected Differentiation Potential toward the Dopaminergic Neuronal Lineage. Int. J. Mol. Sci. 2020, 21, 6589. [Google Scholar] [CrossRef]
- Gaggi, G.; Di Credico, A.; Guarnieri, S.; Mariggiò, M.A.; Di Baldassarre, A.; Ghinassi, B. Human Mesenchymal Amniotic Fluid Stem Cells Reveal an Unexpected Neuronal Potential Differentiating into Functional Spinal Motor Neurons. Front. Cell Dev. Biol. 2022, 10, 936990. [Google Scholar] [CrossRef]
- Arand, J.; Chiang, H.R.; Martin, D.; Snyder, M.P.; Sage, J.; Reijo Pera, R.A.; Wossidlo, M. Tet Enzymes Are Essential for Early Embryogenesis and Completion of Embryonic Genome Activation. EMBO Rep. 2022, 23, e53968. [Google Scholar] [CrossRef]
- Uysal, F.; Ozturk, S.; Akkoyunlu, G. DNMT1, DNMT3A and DNMT3B Proteins Are Differently Expressed in Mouse Oocytes and Early Embryos. J. Mol. Hist. 2017, 48, 417–426. [Google Scholar] [CrossRef]
- Yang, J.; Bashkenova, N.; Zang, R.; Huang, X.; Wang, J. The Roles of TET Family Proteins in Development and Stem Cells. Development 2020, 147, dev183129. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Xie, J.; Mao, L.; Zhao, M.; Bai, X.; Wen, J.; Shen, T.; Wu, P. Bisphenol Analogue Concentrations in Human Breast Milk and Their Associations with Postnatal Infant Growth. Environ. Pollut. 2020, 259, 113779. [Google Scholar] [CrossRef] [PubMed]
- Pinney, S.E.; Mesaros, C.A.; Snyder, N.W.; Busch, C.M.; Xiao, R.; Aijaz, S.; Ijaz, N.; Blair, I.A.; Manson, J.M. Second Trimester Amniotic Fluid Bisphenol A Concentration Is Associated with Decreased Birth Weight in Term Infants. Reprod. Toxicol. 2017, 67, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kundakovic, M.; Gudsnuk, K.; Franks, B.; Madrid, J.; Miller, R.L.; Perera, F.P.; Champagne, F.A. Sex-Specific Epigenetic Disruption and Behavioral Changes Following Low-Dose in Utero Bisphenol A Exposure. Proc. Natl. Acad. Sci. USA 2013, 110, 9956–9961. [Google Scholar] [CrossRef]
- Aimuzi, R.; Luo, K.; Huang, R.; Huo, X.; Nian, M.; Ouyang, F.; Du, Y.; Feng, L.; Wang, W.; Zhang, J. Perfluoroalkyl and Polyfluroalkyl Substances and Maternal Thyroid Hormones in Early Pregnancy. Environ. Pollut. 2020, 264, 114557. [Google Scholar] [CrossRef]
- Rivera-Núñez, Z.; Kinkade, C.W.; Khoury, L.; Brunner, J.; Murphy, H.; Wang, C.; Kannan, K.; Miller, R.K.; O’Connor, T.G.; Barrett, E.S. Prenatal Perfluoroalkyl Substances Exposure and Maternal Sex Steroid Hormones across Pregnancy. Environ. Res. 2023, 220, 115233. [Google Scholar] [CrossRef]
- Wan, H.T.; Wong, A.Y.-M.; Feng, S.; Wong, C.K.-C. Effects of In Utero Exposure to Perfluorooctane Sulfonate on Placental Functions. Environ. Sci. Technol. 2020, 54, 16050–16061. [Google Scholar] [CrossRef]
- Jiang, W.; Deng, Y.; Song, Z.; Xie, Y.; Gong, L.; Chen, Y.; Kuang, H. Gestational Perfluorooctanoic Acid Exposure Inhibits Placental Development by Dysregulation of Labyrinth Vessels and UNK Cells and Apoptosis in Mice. Front. Physiol. 2020, 11, 51. [Google Scholar] [CrossRef]
- Gaggi, G.; Di Credico, A.; Barbagallo, F.; Ghinassi, B.; Di Baldassarre, A. Bisphenols and Perfluoroalkyls Alter Human Stem Cells Integrity: A Possible Link with Infertility. Environ. Res. 2023, 116487. [Google Scholar] [CrossRef] [PubMed]
- Gaggi, G.; Di Credico, A.; Barbagallo, F.; Ballerini, P.; Ghinassi, B.; Di Baldassarre, A. Antenatal Exposure to Plastic Pollutants: Study of the Bisphenols and Perfluoroalkyls Effects on Human Stem Cell Models. Expo. Health 2023. [Google Scholar] [CrossRef]
- Lorigo, M.; Cairrao, E. Fetoplacental Vasculature as a Model to Study Human Cardiovascular Endocrine Disruption. Mol. Asp. Med. 2022, 87, 101054. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, H.B.; Larose, T.L.; Øien, T.; Odland, J.Ø.; Van De Bor, M.; Jacobsen, G.W.; Sandanger, T.M. Factors Associated with Maternal Serum Levels of Perfluoroalkyl Substances and Organochlorines: A Descriptive Study of Parous Women in Norway and Sweden. PLoS ONE 2016, 11, e0166127. [Google Scholar] [CrossRef] [PubMed]
- Pierozan, P.; Cattani, D.; Karlsson, O. Perfluorooctane Sulfonate (PFOS) and Perfluorooctanoic Acid (PFOA) Induce Epigenetic Alterations and Promote Human Breast Cell Carcinogenesis In Vitro. Arch. Toxicol. 2020, 94, 3893–3906. [Google Scholar] [CrossRef]
- Profita, M.; Fabbri, E.; Spisni, E.; Valbonesi, P. Comparing Effects and Action Mechanisms of BPA and BPS on HTR-8/SVneo Placental Cells. Biol. Reprod. 2021, 105, 1355–1364. [Google Scholar] [CrossRef]
- Zhang, A.; Wang, X.; Fan, C.; Mao, X. The Role of Ki67 in Evaluating Neoadjuvant Endocrine Therapy of Hormone Receptor-Positive Breast Cancer. Front. Endocrinol. 2021, 12, 687244. [Google Scholar] [CrossRef]
- Nayak, D.; Adiga, D.; Khan, N.G.; Rai, P.S.; Dsouza, H.S.; Chakrabarty, S.; Gassman, N.R.; Kabekkodu, S.P. Impact of Bisphenol A on Structure and Function of Mitochondria: A Critical Review. Rev. Environ. Contam. Toxicol. 2022, 260, 10. [Google Scholar] [CrossRef]
- Zhou, Y.-T.; Li, R.; Li, S.-H.; Ma, X.; Liu, L.; Niu, D.; Duan, X. Perfluorooctanoic Acid (PFOA) Exposure Affects Early Embryonic Development and Offspring Oocyte Quality via Inducing Mitochondrial Dysfunction. Environ. Int. 2022, 167, 107413. [Google Scholar] [CrossRef]
- Osellame, L.D.; Blacker, T.S.; Duchen, M.R. Cellular and Molecular Mechanisms of Mitochondrial Function. Best Pract. Res. Clin. Endocrinol. Metab. 2012, 26, 711–723. [Google Scholar] [CrossRef]
- Bisaccia, G.; Ricci, F.; Gallina, S.; Di Baldassarre, A.; Ghinassi, B. Mitochondrial Dysfunction and Heart Disease: Critical Appraisal of an Overlooked Association. Int. J. Mol. Sci. 2021, 22, 614. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.C.; Acsadi, G.; Brenner, C.A. Mitochondria: Determinants of Stem Cell Fate? Stem Cells Dev. 2009, 18, 803–806. [Google Scholar] [CrossRef]
- Qiao, W.; Zhang, Y.; Xie, Z.; Luo, Y.; Zhang, X.; Sang, C.; Xie, S.; Huang, J. Toxicity of Perfluorooctane Sulfonate on Phanerochaete Chrysosporium: Growth, Pollutant Degradation and Transcriptomics. Ecotoxicol. Environ. Saf. 2019, 174, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Kleszczyński, K.; Składanowski, A.C. Mechanism of Cytotoxic Action of Perfluorinated Acids. Toxicol. Appl. Pharmacol. 2009, 234, 300–305. [Google Scholar] [CrossRef]
- Napso, T.; Zhao, X.; Lligoña, M.I.; Sandovici, I.; Kay, R.G.; George, A.L.; Gribble, F.M.; Reimann, F.; Meek, C.L.; Hamilton, R.S.; et al. Placental Secretome Characterization Identifies Candidates for Pregnancy Complications. Commun. Biol. 2021, 4, 701. [Google Scholar] [CrossRef]
- Ghassabian, A.; Vandenberg, L.; Kannan, K.; Trasande, L. Endocrine-Disrupting Chemicals and Child Health. Annu. Rev. Pharmacol. Toxicol. 2022, 62, 573–594. [Google Scholar] [CrossRef] [PubMed]
- Campolo, F.; Gori, M.; Favaro, R.; Nicolis, S.; Pellegrini, M.; Botti, F.; Rossi, P.; Jannini, E.A.; Dolci, S. Essential Role of Sox2 for the Establishment and Maintenance of the Germ Cell Line. Stem Cells 2013, 31, 1408–1421. [Google Scholar] [CrossRef] [PubMed]
- Western, P.; Maldonado-Saldivia, J.; van den Bergen, J.; Hajkova, P.; Saitou, M.; Barton, S.; Surani, M.A. Analysis of Esg1 Expression in Pluripotent Cells and the Germline Reveals Similarities with Oct4 and Sox2 and Differences Between Human Pluripotent Cell Lines. Stem Cells 2005, 23, 1436–1442. [Google Scholar] [CrossRef]
- Li, B.; Nair, M.; Mackay, D.R.; Bilanchone, V.; Hu, M.; Fallahi, M.; Song, H.; Dai, Q.; Cohen, P.E.; Dai, X. Ovol1 Regulates Meiotic Pachytene Progression during Spermatogenesis by Repressing Id2 Expression. Development 2005, 132, 1463–1473. [Google Scholar] [CrossRef] [PubMed]
- Antonucci, I.; Di Pietro, R.; Alfonsi, M.; Centurione, M.A.; Centurione, L.; Sancilio, S.; Pelagatti, F.; D’amico, M.A.; Di Baldassarre, A.; Piattelli, A.; et al. Human Second Trimester Amniotic Fluid Cells Are Able to Create Embryoid Body-Like Structures in Vitro and to Show Typical Expression Profiles of Embryonic and Primordial Germ Cells. Cell Transplant. 2014, 23, 1501–1515. [Google Scholar] [CrossRef]
- Gaggi, G.; Di Credico, A.; Guarnieri, S.; Mariggiò, M.A.; Ballerini, P.; Di Baldassarre, A.; Ghinassi, B. Human Fetal Membrane-Mesenchymal Stromal Cells Generate Functional Spinal Motor Neurons In Vitro. iScience 2022, 25, 105197. [Google Scholar] [CrossRef] [PubMed]
- D’amico, M.A.; Ghinassi, B.; Izzicupo, P.; Di Ruscio, A.; Di Baldassarre, A. IL-6 Activates PI3K and PKCζ Signaling and Determines Cardiac Differentiation in Rat Embryonic H9c2 Cells. J. Cell. Physiol. 2016, 231, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Gaggi, G.; Di Credico, A.; Izzicupo, P.; Sancilio, S.; Di Mauro, M.; Iannetti, G.; Dolci, S.; Amabile, G.; Di Baldassarre, A.; Ghinassi, B. Decellularized Extracellular Matrices and Cardiac Differentiation: Study on Human Amniotic Fluid-Stem Cells. Int. J. Mol. Sci. 2020, 21, 6317. [Google Scholar] [CrossRef]
- Di Giacomo, V.; Matteucci, A.; Stellacci, E.; Battistini, A.; Di Baldassarre, A.; Capitani, S.; Alfani, E.; Migliaccio, A.R.; Cocco, L.; Migliaccio, G. Expression of Signal Transduction Proteins during the Differentiation of Primary Human Erythroblasts. J. Cell. Physiol. 2005, 202, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Izzicupo, P.; Ghinassi, B.; D’Amico, M.A.; Di Blasio, A.; Gesi, M.; Napolitano, G.; Gallina, S.; Di Baldassarre, A. Effects of ACE I/D Polymorphism and Aerobic Training on the Immune–Endocrine Network and Cardiovascular Parameters of Postmenopausal Women. J. Clin. Endocrinol. Metab. 2013, 98, 4187–4194. [Google Scholar] [CrossRef]
- Aasen, T.; Raya, A.; Barrero, M.J.; Garreta, E.; Consiglio, A.; Gonzalez, F.; Vassena, R.; Bilić, J.; Pekarik, V.; Tiscornia, G.; et al. Efficient and Rapid Generation of Induced Pluripotent Stem Cells from Human Keratinocytes. Nat. Biotechnol. 2008, 26, 1276–1284. [Google Scholar] [CrossRef]
- Rubino, E.; Cruciani, M.; Tchitchek, N.; Le Tortorec, A.; Rolland, A.D.; Veli, Ö.; Vallet, L.; Gaggi, G.; Michel, F.; Dejucq-Rainsford, N.; et al. Human Ubiquitin-Specific Peptidase 18 Is Regulated by MicroRNAs via the 3’Untranslated Region, A Sequence Duplicated in Long Intergenic Non-Coding RNA Genes Residing in Chr22q11.21. Front. Genet. 2021, 11, 627007. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pollutant | Impedance Value (Fold Change vs. Control) | |
|---|---|---|
| hFM-MSC | hAFSCs | |
| BPA | 1.15 | 1.15 |
| BPS | 1.27 | 1.23 |
| PFOS | 1.17 | 1.40 |
| PFOA | 1.11 | 1.21 |
| BPA + PFOS | 1.17 | 0.92 |
| BPA + PFOA | 1.13 | 1.11 |
| BPS + PFOS | 0.55 | 0.56 |
| BPS + PFOA | 0.94 | 0.93 |
| ALL | 1.20 | 0.92 |
| hFM-MSCs | BPA | BPS | PFOS | PFOA | BPA + PFOS | BPA + PFOA | BPS + PFOS | BPS + PFOA | ALL | ||
| Proliferation Rate | Impedance | + | + | + | + | − | + | ||||
| Ki67+ | ++ | + | ++ | + | |||||||
| Mitochondrial Health | MMP | ||||||||||
| Pluripotency Markers | NANOG | + | |||||||||
| OCT4 | + | ||||||||||
| SOX2 | + | ||||||||||
| ESG1 | ++ | ||||||||||
| KLF4 | + | ||||||||||
| OVOL1 | +++ | + | + | + | |||||||
| Epigenetic Regulators | DNMT1 | + | |||||||||
| DNMT3A | + | ||||||||||
| TET1 | ++ | ||||||||||
| TET2 | |||||||||||
| TET3 | + | ++ | + | + | |||||||
| hAFSCs | Proliferation Rate | Impedance | + | + | + | + | |||||
| Ki67+ | + | + | + | + | |||||||
| Mitochondrial Health | MMP | − | |||||||||
| Pluripotency Markers | NANOG | + | ++ | + | + | ||||||
| OCT4 | + | ++ | + | + | + | ||||||
| SOX2 | + | + | |||||||||
| ESG1 | + | +++ | +++ | ||||||||
| KLF4 | |||||||||||
| REX1 | |||||||||||
| OVOL1 | ++ | + | + | ||||||||
| Epigenetic Regulators | DNMT1 | ++ | + | ||||||||
| DNMT3A | |||||||||||
| TET1 | + | + | + | + | + | ||||||
| TET2 | |||||||||||
| TET3 | + | ++ | + | + |
| Gene | Sequence (5′ to 3′) | Reference |
|---|---|---|
| NANOG_FW | CCAGACCCAGAACATCCAGTC | [17] |
| NANOG_REV | CACTGGCAGGAGAATTTGGC | |
| OCT4_FW | GGGTTTTTGGGATTAAGTTCTTC | [56] |
| OCT4_REV | GCCCCCACCCTTTGTGTT | |
| SOX2_FW | CAAAAATGGCCATGCAGGTT | [56] |
| SOX2_REV | AGTTGGGATCGAACAAAAGCTATT | |
| ESG1_FW | CCATGAATGCCCTCGAACTAGG | [19] |
| ESG1_REV | CCTTAACTCTTTAGGCTGGAGCA | |
| KLF4_FW | AGCCTAAATGATGGTGCTTGGT | [56] |
| KLF4_REV | TTGAAAACTTTGGCTTCCTTGTT | |
| OVOL1_FW | AGAGCAGAGACCATGGCTTC | [50] |
| OVOL1_REV | GACGTGTCTCTTGAGGTCGA | |
| DNMT1_FW | GCCAGAGATAGAGATCAAGCTG | Primer blast |
| DNMT1_REV | CACAGCGTGTCAGAGATGCC | |
| DNTM3A_FW | CCATCGTCAACCCTGCTCG | Primer blast |
| DNMT3A_REV | CACCACATTCTCAAAGAGCCAG | |
| DNTM3B_FW | GACTCGTTCAGAAAGCCCAG | Primer blast |
| DNMT3B_REV | GGACTCGTCCACATGGTTGC | |
| TET1_FW | ACTCCCTGAGGTCTGTCCTG | Primer blast |
| TET1_REV | CAGGTAGGGCTGCATGACTT | |
| TET2_FW | CTCAGCAGCAGCCAATAGGA | Primer blast |
| TET2_REV | CTGTCTGGCAAATGGGAGGT | |
| TET3_FW | AACTGCTCACTCAGCTCTGC | Primer blast |
| TET3_REV | GCAGCCCTCAGAAAAGGGAT | |
| 18S_FW | CATGGCCGTTCTTAGTTGGT | [57] |
| 18S_REV | CGCTGAGCCAGTCAGTGTAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Credico, A.; Gaggi, G.; Bucci, I.; Ghinassi, B.; Di Baldassarre, A. The Effects of Combined Exposure to Bisphenols and Perfluoroalkyls on Human Perinatal Stem Cells and the Potential Implications for Health Outcomes. Int. J. Mol. Sci. 2023, 24, 15018. https://doi.org/10.3390/ijms241915018
Di Credico A, Gaggi G, Bucci I, Ghinassi B, Di Baldassarre A. The Effects of Combined Exposure to Bisphenols and Perfluoroalkyls on Human Perinatal Stem Cells and the Potential Implications for Health Outcomes. International Journal of Molecular Sciences. 2023; 24(19):15018. https://doi.org/10.3390/ijms241915018
Chicago/Turabian StyleDi Credico, Andrea, Giulia Gaggi, Ines Bucci, Barbara Ghinassi, and Angela Di Baldassarre. 2023. "The Effects of Combined Exposure to Bisphenols and Perfluoroalkyls on Human Perinatal Stem Cells and the Potential Implications for Health Outcomes" International Journal of Molecular Sciences 24, no. 19: 15018. https://doi.org/10.3390/ijms241915018
APA StyleDi Credico, A., Gaggi, G., Bucci, I., Ghinassi, B., & Di Baldassarre, A. (2023). The Effects of Combined Exposure to Bisphenols and Perfluoroalkyls on Human Perinatal Stem Cells and the Potential Implications for Health Outcomes. International Journal of Molecular Sciences, 24(19), 15018. https://doi.org/10.3390/ijms241915018