
Unveiling the Genetic Complexity of Teratozoospermia: Integrated Genomic Analysis Reveals Novel Insights into lncRNAs’ Role in Male Infertility

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. WGS and RNA Expression Results—Identification of Exclusive Variants on DE lncRNAs
2.2. Exclusive Variants with a Functional Role
2.3. Exclusive Variants Affecting lncRNAs’ Structure
2.4. Exclusive Variants Affecting miRNA–lncRNA Interactions and Investigation of Target Genes of Affected miRNAs
2.5. Variants Affecting Both Structure or Function of lncRNAs and miRNA–lncRNA Interactions
2.6. lncRNAs with Multiple Variants Affecting Their Function, Structure, or Interactions with miRNAs
3. Discussion
3.1. Impact of Variants on the Functionality of lncRNAs
3.2. Impact of Variants on the lncRNAs’ Structure
3.3. Impact of Variants on miRNA–lncRNA Interactions and Processes and the Pathways Affected
3.4. Common Variants and lncRNAs with Multiple Pioritized Variants
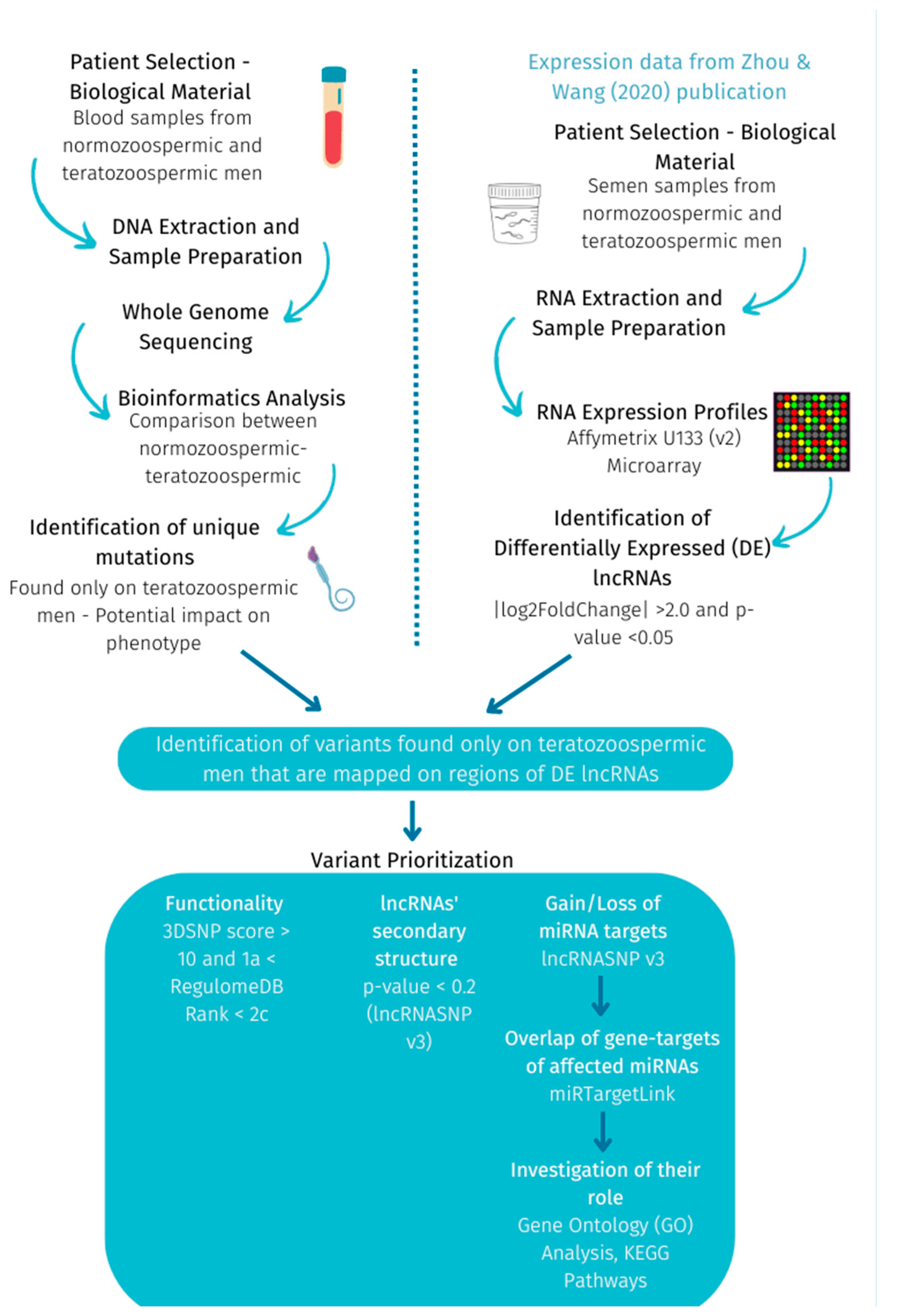
4. Materials and Methods
4.1. Whole Genome Sequencing (WGS)—Identification of Exclusive Variants on Teratozoospermic Men
4.2. RNA Expression Profiles—Identification of Differentially Expressed lncRNAs between Normozoospermic and Teratozoospermic Men
4.3. Identification of Exclusive Variants on DE lncRNAs and Variant Prioritization—Investigation of Their Role and Consequence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mascarenhas, M.N.; Flaxman, S.R.; Boerma, T.; Vanderpoel, S.; Stevens, G.A. National, Regional, and Global Trends in Infertility Prevalence Since 1990: A Systematic Analysis of 277 Health Surveys. PLoS Med. 2012, 9, e1001356. [Google Scholar] [CrossRef] [PubMed]
- Datta, J.; Palmer, M.J.; Tanton, C.; Gibson, L.J.; Jones, K.G.; Macdowall, W.; Glasier, A.; Sonnenberg, P.; Field, N.; Mercer, C.H.; et al. Prevalence of Infertility and Help Seeking among 15 000 Women and Men. Hum. Reprod. 2016, 31, 2108–2118. [Google Scholar] [CrossRef] [PubMed]
- Pathak, U.I.; Gabrielsen, J.S.; Lipshultz, L.I. Cutting-Edge Evaluation of Male Infertility. Urol. Clin. N. Am. 2020, 47, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Baskaran, S.; Parekh, N.; Cho, C.L.; Henkel, R.; Vij, S.; Arafa, M.; Panner Selvam, M.K.; Shah, R. Male Infertility. Lancet 2021, 397, 319–333. [Google Scholar] [CrossRef]
- Tü, F.; Werny, F.; Cooper, T.G.; Kliesch, S.; Simoni, M.; Nieschlag, E. Clinical Experience with Azoospermia: Aetiology and Chances for Spermatozoa Detection upon Biopsy. Wiley Online Libr. 2011, 34, 291–298. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination of Human Semen and Semen-Cervical Mucus Interaction; Press Concern: Singapore, 1980. [Google Scholar]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; Press Concern: Singapore, 2010. [Google Scholar]
- De Braekeleer, M.; Nguyen, M.H.; Morel, F.; Perrin, A. Genetic Aspects of Monomorphic Teratozoospermia: A Review. J. Assist. Reprod. Genet. 2015, 32, 615. [Google Scholar] [CrossRef]
- Coutton, C.; Escoffier, J.; Martinez, G.; Arnoult, C.; Ray, P.F. Teratozoospermia: Spotlight on the Main Genetic Actors in the Human. Hum. Reprod. Update 2015, 21, 455–485. [Google Scholar] [CrossRef]
- Jan, S.Z.; Vormer, T.L.; Jongejan, A.; Röling, M.D.; Silber, S.J.; de Rooij, D.G.; Hamer, G.; Repping, S.; van Pelt, A.M.M. Unraveling Transcriptome Dynamics in Human Spermatogenesis. Development 2017, 144, 3659–3673. [Google Scholar] [CrossRef]
- Palazzo, A.F.; Lee, E.S. Non-Coding RNA: What Is Functional and What Is Junk? Front. Genet. 2015, 5, 2. [Google Scholar] [CrossRef]
- Kyrgiafini, M.A.; Sarafidou, T.; Mamuris, Z. The Role of Long Noncoding RNAs on Male Infertility: A Systematic Review and In Silico Analysis. Biology 2022, 11, 1510. [Google Scholar] [CrossRef]
- Lekka, E.; Hall, J. Noncoding RNAs in Disease. FEBS Lett. 2018, 592, 2884. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, G.K.; Khullar, N.; Sidhu, I.S.; Navik, U.S.; Reddy, A.P.; Reddy, P.H.; Bhatti, J.S. Emerging Role of Non-coding RNA in Health and Disease. Metab. Brain Dis. 2021, 36, 1119–1134. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Li, Y.; Li, J.; Gao, Z.; Yang, Z.; Li, Y.; Liu, H.; Fan, T. Long Non-Coding RNAs: The Regulatory Mechanisms, Research Strategies, and Future Directions in Cancers. Front. Oncol. 2020, 10, 598817. [Google Scholar] [CrossRef] [PubMed]
- Balas, M.M.; Johnson, A.M. Exploring the Mechanisms behind Long Noncoding RNAs and Cancer. Non-coding RNA Res. 2018, 3, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Chang, H.Y. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef]
- Bolha, L.; Ravnik-Glavač, M.; Glavač, D. Long Noncoding RNAs as Biomarkers in Cancer. Dis. Markers 2017, 2017, 7243968. [Google Scholar] [CrossRef]
- Mukherjee, A.; Koli, S.; Reddy, K.V.R. Regulatory Non-Coding Transcripts in Spermatogenesis: Shedding Light on ‘Dark Matter’. Andrology 2014, 2, 360–369. [Google Scholar] [CrossRef]
- Joshi, M.; Rajender, S. Long Non-Coding RNAs (LncRNAs) in Spermatogenesis and Male Infertility. Reprod. Biol. Endocrinol. 2020, 18, 103. [Google Scholar] [CrossRef]
- Aznaourova, M.; Schmerer, N.; Schmeck, B.; Schulte, L.N. Disease-Causing Mutations and Rearrangements in Long Non-Coding RNA Gene Loci. Front. Genet. 2020, 11, 1485. [Google Scholar] [CrossRef]
- Hindorff, L.A.; Sethupathy, P.; Junkins, H.A.; Ramos, E.M.; Mehta, J.P.; Collins, F.S.; Manolio, T.A. Potential Etiologic and Functional Implications of Genome-Wide Association Loci for Human Diseases and Traits. Proc. Natl. Acad. Sci. USA 2009, 106, 9362–9367. [Google Scholar] [CrossRef]
- Kulkarni, S.; Lied, A.; Kulkarni, V.; Rucevic, M.; Martin, M.P.; Walker-Sperling, V.; Anderson, S.K.; Ewy, R.; Singh, S.; Nguyen, H.; et al. CCR5AS LncRNA Variation Differentially Regulates CCR5, Influencing HIV Disease Outcome. Nat. Immunol. 2019, 20, 824–834. [Google Scholar] [CrossRef]
- Olazagoitia-Garmendia, A.; Sebastian-delaCruz, M.; Castellanos-Rubio, A. Involvement of LncRNAs in Celiac Disease Pathogenesis. Int. Rev. Cell Mol. Biol. 2021, 358, 241–264. [Google Scholar] [PubMed]
- Leucci, E.; Vendramin, R.; Spinazzi, M.; Laurette, P.; Fiers, M.; Wouters, J.; Radaelli, E.; Eyckerman, S.; Leonelli, C.; Vanderheyden, K.; et al. Melanoma Addiction to the Long Non-Coding RNA SAMMSON. Nature 2016, 531, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Cabianca, D.S.; Casa, V.; Bodega, B.; Xynos, A.; Ginelli, E.; Tanaka, Y.; Gabellini, D. A Long NcRNA Links Copy Number Variation to a Polycomb/Trithorax Epigenetic Switch in FSHD Muscular Dystrophy. Cell 2012, 149, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Wang, B. Identification of Male Infertility-Related Long Non-Coding RNAs and Their Functions Based on a Competing Endogenous RNA Network. J. Int. Med. Res. 2020, 48, 0300060520961277. [Google Scholar] [CrossRef] [PubMed]
- Boyle, A.P.; Hong, E.L.; Hariharan, M.; Cheng, Y.; Schaub, M.A.; Kasowski, M.; Karczewski, K.J.; Park, J.; Hitz, B.C.; Weng, S.; et al. Annotation of Functional Variation in Personal Genomes Using RegulomeDB. Genome Res. 2012, 22, 1790–1797. [Google Scholar] [CrossRef]
- Lu, Y.; Quan, C.; Chen, H.; Bo, X.; Zhang, C. 3DSNP: A Database for Linking Human Noncoding SNPs to Their Three-Dimensional Interacting Genes. Nucleic Acids Res. 2017, 45, D643–D649. [Google Scholar] [CrossRef]
- Martens, L.; Rühle, F.; Witten, A.; Meder, B.; Katus, H.A.; Arbustini, E.; Hasenfuß, G.; Sinner, M.F.; Kääb, S.; Pankuweit, S.; et al. A Genetic Variant Alters the Secondary Structure of the LncRNA H19 and Is Associated with Dilated Cardiomyopathy. RNA Biol. 2021, 18, 409. [Google Scholar] [CrossRef]
- Halvorsen, M.; Martin, J.S.; Broadaway, S.; Laederach, A. Disease-Associated Mutations That Alter the RNA Structural Ensemble. PLoS Genet. 2010, 6, e1001074. [Google Scholar] [CrossRef]
- Wan, Y.; Qu, K.; Zhang, Q.C.; Flynn, R.A.; Manor, O.; Ouyang, Z.; Zhang, J.; Spitale, R.C.; Snyder, M.P.; Segal, E.; et al. Landscape and Variation of RNA Secondary Structure across the Human Transcriptome. Nature 2014, 505, 706–709. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, D.; Miao, Y.R.; Wu, X.; Luo, H.; Cao, W.; Yang, W.; Yang, J.; Guo, A.Y.; Gong, J. LncRNASNP v3: An Updated Database for Functional Variants in Long Non-Coding RNAs. Nucleic Acids Res. 2023, 51, D192–D198. [Google Scholar] [CrossRef]
- Karagkouni, D.; Karavangeli, A.; Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Characterizing MiRNA-LncRNA Interplay. Methods Mol. Biol. 2021, 2372, 243–262. [Google Scholar] [PubMed]
- Finotti, A.; Fabbri, E.; Lampronti, I.; Gasparello, J.; Borgatti, M.; Gambari, R. MicroRNAs and Long Non-Coding RNAs in Genetic Diseases. Mol. Diagn. Ther. 2019, 23, 155. [Google Scholar] [CrossRef] [PubMed]
- Kern, F.; Aparicio-Puerta, E.; Li, Y.; Fehlmann, T.; Kehl, T.; Wagner, V.; Ray, K.; Ludwig, N.; Lenhof, H.P.; Meese, E.; et al. MiRTargetLink 2.0—Interactive MiRNA Target Gene and Target Pathway Networks. Nucleic Acids Res. 2021, 49, W409–W416. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T.G.O.; Aleksander, S.A.; Balhoff, J.; Carbon, S.; Cherry, J.M.; Drabkin, H.J.; Ebert, D.; Feuermann, M.; Gaudet, P.; Harris, N.L.; et al. The Gene Ontology Knowledgebase in 2023. Genetics 2023, 224, iyad031. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for Taxonomy-Based Analysis of Pathways and Genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef]
- Sparber, P.; Filatova, A.; Khantemirova, M.; Skoblov, M. The Role of Long Non-Coding RNAs in the Pathogenesis of Hereditary Diseases. BMC Med. Genom. 2019, 12, 63–78. [Google Scholar] [CrossRef]
- Landi, M.T.; Bishop, D.T.; MacGregor, S.; Machiela, M.J.; Stratigos, A.J.; Ghiorzo, P.; Brossard, M.; Calista, D.; Choi, J.; Fargnoli, M.C.; et al. Genome-Wide Association Meta-Analyses Combining Multiple Risk Phenotypes Provide Insights into the Genetic Architecture of Cutaneous Melanoma Susceptibility. Nat. Genet. 2020, 52, 494–504. [Google Scholar] [CrossRef]
- Xu, J.J.; Jiang, L.; Xu, L.J.; Gao, Z.; Zhao, X.Y.; Zhang, Y.; Song, Y.; Liu, R.; Sun, K.; Gao, R.L.; et al. Association of CDKN2B-AS1 Polymorphisms with Premature Triple-Vessel Coronary Disease and Their Sex Specificity in the Chinese Population. Biomed. Environ. Sci. 2018, 31, 787–796. [Google Scholar]
- Wang, Y.; Lu, G.; Xue, X.; Xie, M.; Wang, Z.; Ma, Z.; Feng, Y.; Shao, C.; Duan, H.; Pan, M.; et al. Characterization and Validation of a Ferroptosis-Related LncRNA Signature as a Novel Prognostic Model for Lung Adenocarcinoma in Tumor Microenvironment. Front. Immunol. 2022, 13, 903758. [Google Scholar] [CrossRef]
- Yuan, L.Y.; Qin, X.; Li, L.; Zhou, J.; Zhou, M.; Li, X.; Xu, Y.; Wang, X.J.; Xing, H. The Transcriptome Profiles and Methylation Status Revealed the Potential Cancer-Related LncRNAs in Patients with Cervical Cancer. J. Cell. Physiol. 2019, 234, 9756–9763. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Song, D.; Zhao, G.; Chen, S.; Ren, H.; Zhang, B. Cross-Talk between Necroptosis-Related LncRNAs to Construct a Novel Signature and Predict the Immune Landscape of Lung Adenocarcinoma Patients. Front. Genet. 2022, 13, 966896. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Lei, X.; Lu, Q.; Wu, Q.; Ma, Q.; Huang, D.; Zhang, Y. LncRNA FRMD6-AS1 Promotes Hepatocellular Carcinoma Cell Migration and Stemness by Regulating SENP1/HIF-1α Axis. Pathol. Res. Pract. 2023, 243, 154377. [Google Scholar] [CrossRef]
- Ma, T.; Hu, Y.; Guo, Y.; Yan, B. Tumor-Promoting Activity of Long Noncoding RNA LINC00466 in Lung Adenocarcinoma via MiR-144-Regulated HOXA10 Axis. Am. J. Pathol. 2019, 189, 2154–2170. [Google Scholar] [CrossRef]
- Liu, J.; Yu, H.; Cui, H.; Wei, F.; Yan, T.; Li, T.; Liu, Y.; Chu, J. LncRNA LINC000466 Predicts the Prognosis and Promotes the Progression of Triple-Negative Breast Cancer via Modulating MiR-539-5p. Clin. Breast Cancer 2022, 22, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Shao, Y.; Xu, J.; Zhang, B.; Li, C.; Gong, J. LINC00466 Impacts Cell Proliferation, Metastasis and Sensitivity to Temozolomide of Glioma by Sponging MiR-137 to Regulate PPP1R14B Expression. Onco. Targets. Ther. 2021, 14, 1147–1159. [Google Scholar] [CrossRef]
- Zimta, A.A.; Tomuleasa, C.; Sahnoune, I.; Calin, G.A.; Berindan-Neagoe, I. Long Non-Coding RNAs in Myeloid Malignancies. Front. Oncol. 2019, 9, 1048. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, C.; Cao, S.; Zhao, H.; Jiang, R.; Li, Y. Tumor-Derived Exosomes Orchestrate the MicroRNA-128-3p/ELF4/CDX2 Axis to Facilitate the Growth and Metastasis of Gastric Cancer via Delivery of LINC01091. Cell Biol. Toxicol. 2022, 39, 519–536. [Google Scholar] [CrossRef]
- Zhou, C.; Duan, S. The Role of Long Non-Coding RNA NNT-AS1 in Neoplastic Disease. Cancers 2020, 12, 3086. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, L.; Hu, X.; Jiang, Y.; Hu, Y.; Liu, Z.; Liu, J.; Wen, T.; Ma, Y.; An, G.; et al. Upregulated NNT-AS1, a Long Noncoding RNA, Contributes to Proliferation and Migration of Colorectal Cancer Cells in Vitro and in Vivo. Oncotarget 2017, 8, 3441–3453. [Google Scholar] [CrossRef]
- Ye, H.; Lin, J.; Yao, X.; Li, Y.; Lin, X.; Lu, H. Overexpression of Long Non-Coding RNA NNT-AS1 Correlates with Tumor Progression and Poor Prognosis in Osteosarcoma. Cell. Physiol. Biochem. 2018, 45, 1904–1914. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Yuan, M.; Su, L.; Ge, M.; Dong, F. ZNF252P-AS1 Enhances Myeloid-Derived Suppressor Cell Expansion and Tumor Glycolysis to Accelerate Laryngeal Squamous Cell Carcinoma Development via the MiR-377-3p/Lactate Dehydrogenase a Axis. Mol. Cell. Toxicol. 2023, 1–11. [Google Scholar] [CrossRef]
- Wu, H.; Li, A.; Zheng, Q.; Gu, J.; Zhou, W. LncRNA LZTS1-AS1 Induces Proliferation, Metastasis and Inhibits Autophagy of Pancreatic Cancer Cells through the MiR-532/TWIST1 Signaling Pathway. Cancer Cell Int. 2023, 23, 130. [Google Scholar] [CrossRef] [PubMed]
- WANG, W.; CHENG, X.; ZHU, J. Long Non-Coding RNA OTUD6B-AS1 Overexpression Inhibits the Proliferation, Invasion and Migration of Colorectal Cancer Cells via Downregulation of MicroRNA-3171. Oncol. Lett. 2021, 21, 193. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.; Xue, H.; Li, Y.; Li, P.; Ma, F.; Liu, M.; Li, W. The Long Noncoding RNA OTUD6B-AS1 Enhances Cell Proliferation and the Invasion of Hepatocellular Carcinoma Cells through Modulating GSKIP/Wnt/β-Catenin Signalling via the Sequestration of MiR-664b-3p. Exp. Cell Res. 2020, 395, 112180. [Google Scholar] [CrossRef]
- Ferreux, L.; Boumerdassi, Y.; Dulioust, E.; Bertagna, X.; Roucher-Boulez, F.; Bourdon, M.; Thiounn, N.; Patrat, C. Testicular Impairment in Primary Adrenal Insufficiency Caused by Nicotinamide Nucleotide Transhydrogenase (NNT) Deficiency—A Case Report: Implication of Oxidative Stress and Importance of Fertility Preservation. Basic Clin. Androl. 2023, 33, 17. [Google Scholar] [CrossRef]
- Roucher-Boulez, F.; Mallet-Motak, D.; Samara-Boustani, D.; Jilani, H.; Ladjouze, A.; Souchon, P.F.; Simon, D.; Nivot, S.; Heinrichs, C.; Ronze, M.; et al. NNT Mutations: A Cause of Primary Adrenal Insufficiency, Oxidative Stress and Extra-Adrenal Defects. Eur. J. Endocrinol. 2016, 175, 73–84. [Google Scholar] [CrossRef]
- Ramírez-Bello, J.; Jiménez-Morales, M. Functional Implications of Single Nucleotide Polymorphisms (SNPs) in Protein-Coding and Non-Coding RNA Genes in Multifactorial Diseases. Gac. Med. Mex. 2017, 153, 238–250. [Google Scholar]
- Baytak, E.; Gong, Q.; Akman, B.; Yuan, H.; Chan, W.C.; Küçük, C. Whole Transcriptome Analysis Reveals Dysregulated Oncogenic LncRNAs in Natural Killer/T-Cell Lymphoma and Establishes MIR155HG as a Target of PRDM1. Tumour Biol. 2017, 39, 1010428317701648. [Google Scholar] [CrossRef]
- Kim, S.; Günesdogan, U.; Zylicz, J.J.; Hackett, J.A.; Cougot, D.; Bao, S.; Lee, C.; Dietmann, S.; Allen, G.E.; Sengupta, R.; et al. PRMT5 Protects Genomic Integrity during Global DNA Demethylation in Primordial Germ Cells and Preimplantation Embryos. Mol. Cell 2014, 56, 564–579. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Chen, M.; Chen, M.; Jiang, L.; Shen, Z.; Ma, L.; Han, C.; Guo, X.; Gao, F. PRMT5 Is Involved in Spermatogonial Stem Cells Maintenance by Regulating Plzf Expression via Modulation of Lysine Histone Modifications. Front. Cell Dev. Biol. 2021, 9, 673258. [Google Scholar] [CrossRef]
- Deng, Y.; Guo, K.; Tang, Z.; Feng, Y.; Cai, S.; Ye, J.; Xi, Y.; Li, J.; Liu, R.; Cai, C.; et al. Identification and Experimental Validation of a Tumor-Infiltrating Lymphocytes-Related Long Noncoding RNA Signature for Prognosis of Clear Cell Renal Cell Carcinoma. Front. Immunol. 2022, 13, 1046790. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Guo, Y.; Chen, Z.; Zhang, F.; Qin, Y. Accurate Prognostic Prediction for Patients with Clear Cell Renal Cell Carcinoma Using a Ferroptosis-Related Long Non-Coding RNA Risk Model. Cancer Biomark. 2023, 37, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zheng, H. LncRNA LINC00944 Promotes Tumorigenesis but Suppresses Akt Phosphorylation in Renal Cell Carcinoma. Front. Mol. Biosci. 2021, 8, 697962. [Google Scholar] [CrossRef]
- McKay, J.D.; Hung, R.J.; Han, Y.; Zong, X.; Carreras-Torres, R.; Christiani, D.C.; Caporaso, N.E.; Johansson, M.; Xiao, X.; Li, Y.; et al. Large-Scale Association Analysis Identifies New Lung Cancer Susceptibility Loci and Heterogeneity in Genetic Susceptibility across Histological Subtypes. Nat. Genet. 2017, 49, 1126–1132. [Google Scholar] [CrossRef]
- Tan, C.; Cao, J.; Chen, L.; Xi, X.; Wang, S.; Zhu, Y.; Yang, L.; Ma, L.; Wang, D.; Yin, J.; et al. Noncoding RNAs Serve as Diagnosis and Prognosis Biomarkers for Hepatocellular Carcinoma. Clin. Chem. 2019, 65, 905–915. [Google Scholar] [CrossRef]
- Huang, X.; Liu, X.; Du, B.; Liu, X.; Xue, M.; Yan, Q.; Wang, X.; Wang, Q. LncRNA LINC01305 Promotes Cervical Cancer Progression through KHSRP and Exosome-Mediated Transfer. Aging 2021, 13, 19230–19242. [Google Scholar] [CrossRef]
- Duan, C. LncRNA SLC16A1-AS1 Contributes to the Progression of Hepatocellular Carcinoma Cells by Modulating MiR-411/MITD1 Axis. J. Clin. Lab. Anal. 2022, 36, e24344. [Google Scholar] [CrossRef]
- Logotheti, S.; Marquardt, S.; Gupta, S.K.; Richter, C.; Edelhäuser, B.A.H.; Engelmann, D.; Brenmoehl, J.; Söhnchen, C.; Murr, N.; Alpers, M.; et al. LncRNA-SLC16A1-AS1 Induces Metabolic Reprogramming during Bladder Cancer Progression as Target and Co-Activator of E2F1. Theranostics 2020, 10, 9620–9643. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, T.; Xu, H.; Wang, Y.; Yang, T.; Liu, L. LncRNA FIRRE Promotes the Proliferation and Metastasis of Hepatocellular Carcinoma by Regulating the Expression of PXN through Interacting with MBNL3. Biochem. Biophys. Res. Commun. 2022, 625, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Z.; Xu, S.; Li, W.; Chen, M.; Jiang, M.; Fan, X. LncRNA FIRRE Functions as a Tumor Promoter by Interaction with PTBP1 to Stabilize BECN1 MRNA and Facilitate Autophagy. Cell Death Dis. 2022, 13, 98. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Xu, D.; Wang, P.; Sun, W.; Xue, X.; Hu, Y.; Xie, C.; Ma, Y. RNA-Sequencing and Bioinformatics Analysis of Long Noncoding RNAs and MRNAs in the Asthenozoospermia. Biosci. Rep. 2020, 40, BSR20194041. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.C.; Zhang, Y.; Yu, K.; Li, Y.; Yu, H.; Zhou, S.J.; Wang, Y.P.; Deng, S.L.; Tian, L. LncRNAs Induce Oxidative Stress and Spermatogenesis by Regulating Endoplasmic Reticulum Genes and Pathways. Aging 2021, 13, 13764–13787. [Google Scholar] [CrossRef] [PubMed]
- Kui, F.; Ye, H.; Chen, X.L.; Zhang, J. Microarray Meta-Analysis Identifies Candidate Genes for Human Spermatogenic Arrest. Andrologia 2019, 51, e13301. [Google Scholar] [CrossRef]
- Yang, F.; Deng, X.; Ma, W.; Berletch, J.B.; Rabaia, N.; Wei, G.; Moore, J.M.; Filippova, G.N.; Xu, J.; Liu, Y.; et al. The LncRNA Firre Anchors the Inactive X Chromosome to the Nucleolus by Binding CTCF and Maintains H3K27me3 Methylation. Genome Biol. 2015, 16, 52. [Google Scholar] [CrossRef]
- Hatok, J.; Racay, P. Bcl-2 Family Proteins: Master Regulators of Cell Survival. Biomol. Concepts 2016, 7, 259–270. [Google Scholar] [CrossRef]
- Mostafa, T.; Rashed, L.; Nabil, N.; Amin, R. Seminal BAX and BCL2 Gene and Protein Expressions in Infertile Men With Varicocele. Urology 2014, 84, 590–595. [Google Scholar] [CrossRef]
- Chen, H.; Lam Fok, K.; Jiang, X.; Chan, H.C. New Insights into Germ Cell Migration and Survival/Apoptosis in Spermatogenesis. Spermatogenesis 2012, 2, 264–272. [Google Scholar] [CrossRef]
- Eisenberg, M.L.; Li, S.; Brooks, J.D.; Cullen, M.R.; Baker, L.C. Increased Risk of Cancer in Infertile Men: Analysis of U.S. Claims Data. J. Urol. 2015, 193, 1596–1601. [Google Scholar] [CrossRef]
- Anderson, R.E.; Hanson, H.A.; Lowrance, W.T.; Redshaw, J.; Oottamasathien, S.; Schaeffer, A.; Johnstone, E.; Aston, K.I.; Carrell, D.T.; Cartwright, P.; et al. Childhood Cancer Risk in the Siblings and Cousins of Men with Poor Semen Quality. J. Urol. 2017, 197, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, M.L.; Betts, P.; Herder, D.; Lamb, D.J.; Lipshultz, L.I. Increased Risk of Cancer among Azoospermic Men. Fertil. Steril. 2013, 100, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Hotaling, J.M.; Walsh, T.J. Male Infertility: A Risk Factor for Testicular Cancer. Nat. Rev. Urol. 2009, 6, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Nagirnaja, L.; Aston, K.I.; Conrad, D.F. Genetic Intersection of Male Infertility and Cancer. Fertil. Steril. 2018, 109, 20–26. [Google Scholar] [CrossRef]
- Tvrda, E.; Agarwal, A.; Alkuhaimi, N. Male Reproductive Cancers and Infertility: A Mutual Relationship. Int. J. Mol. Sci. 2015, 16, 7230–7260. [Google Scholar] [CrossRef]
- Neto, F.T.L.; Bach, P.V.; Najari, B.B.; Li, P.S.; Goldstein, M. Spermatogenesis in Humans and Its Affecting Factors. Semin. Cell Dev. Biol. 2016, 59, 10–26. [Google Scholar] [CrossRef]
- Link, W. Introduction to FOXO Biology. Methods Mol. Biol. 2019, 1890, 1–9. [Google Scholar]
- Zhang, X.; Tang, N.; Hadden, T.J.; Rishi, A.K. Akt, FoxO and Regulation of Apoptosis. Biochim. Biophys. Acta-Mol. Cell Res. 2011, 1813, 1978–1986. [Google Scholar] [CrossRef]
- Luo, D.; He, Z.; Yu, C.; Guan, Q. Role of P38 MAPK Signalling in Testis Development and Male Fertility. Oxid. Med. Cell. Longev. 2022, 2022, 6891897. [Google Scholar] [CrossRef]
- Ni, F.D.; Hao, S.L.; Yang, W.X. Multiple Signaling Pathways in Sertoli Cells: Recent Findings in Spermatogenesis. Cell Death Dis. 2019, 10, 541. [Google Scholar] [CrossRef]
- Li, M.W.M.; Mruk, D.D.; Cheng, C.Y. Mitogen-Activated Protein Kinases in Male Reproductive Function. Trends Mol. Med. 2009, 15, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.-Y.; Lv, M.; Luo, B.-H.; Zhao, S.-Z.; Mo, Z.-C.; Xie, Y.-J. The Role of the PI3K/AKT/MTOR Signalling Pathway in Male Reproduction. Curr. Mol. Med. 2021, 21, 539–548. [Google Scholar]
- Yao, P.L.; Lin, Y.C.; Richburg, J.H. TNF Alpha-Mediated Disruption of Spermatogenesis in Response to Sertoli Cell Injury in Rodents Is Partially Regulated by MMP2. Biol. Reprod. 2009, 80, 581. [Google Scholar] [CrossRef] [PubMed]
- Masola, V.; Greco, N.; Tozzo, P.; Caenazzo, L.; Onisto, M. The Role of SPATA2 in TNF Signaling, Cancer, and Spermatogenesis. Cell Death Dis. 2022, 13, 977. [Google Scholar] [CrossRef] [PubMed]
- Kierszenbaum, A.L. Tyrosine Protein Kinases and Spermatogenesis: Truncation Matters. Mol. Reprod. Dev. 2006, 73, 399–403. [Google Scholar] [CrossRef]
- Jenardhanan, P.; Mathur, P.P. Kinases as Targets for Chemical Modulators: Structural Aspects and Their Role in Spermatogenesis. Spermatogenesis 2015, 4, e979113. [Google Scholar] [CrossRef]
- Rothzerg, E.; Ho, X.D.; Xu, J.; Wood, D.; Märtson, A.; Kõks, S. Upregulation of 15 Antisense Long Non-Coding RNAs in Osteosarcoma. Genes 2021, 12, 1132. [Google Scholar] [CrossRef]
- Liu, L.; Li, X.; Wu, H.; Tang, Y.; Li, X.; Shi, Y. The COX10-AS1/MiR-641/E2F6 Feedback Loop Is Involved in the Progression of Glioma. Front. Oncol. 2021, 11, 648152. [Google Scholar] [CrossRef]
- Zhou, C.; Jiang, X.; Liang, A.; Zhu, R.; Yang, Y.; Zhong, L.; Wan, D. COX10-AS1 Facilitates Cell Proliferation and Inhibits Cell Apoptosis in Glioblastoma Cells at Post-Transcription Level. Neurochem. Res. 2020, 45, 2196–2203. [Google Scholar] [CrossRef]
- Xing, C.; Cai, Z.; Gong, J.; Zhou, J.; Xu, J.; Guo, F. Identification of Potential Biomarkers Involved in Gastric Cancer Through Integrated Analysis of Non-Coding RNA Associated Competing Endogenous RNAs Network. Clin. Lab. 2018, 64, 1661–1669. [Google Scholar] [CrossRef]
- Xu, H.; Jia, J. Immune-Related Hub Genes and the Competitive Endogenous RNA Network in Alzheimer’s Disease. J. Alzheimers. Dis. 2020, 77, 1255–1265. [Google Scholar] [CrossRef]
- Xu, Y.; Yu, X.; Zhang, M.; Zheng, Q.; Sun, Z.; He, Y.; Guo, W. Promising Advances in LINC01116 Related to Cancer. Front. cell Dev. Biol. 2021, 9, 736927. [Google Scholar] [CrossRef]
- Zhang, E.; He, X.; Zhang, C.; Su, J.; Lu, X.; Si, X.; Chen, J.; Yin, D.; Han, L.; De, W. A Novel Long Noncoding RNA HOXC-AS3 Mediates Tumorigenesis of Gastric Cancer by Binding to YBX1. Genome Biol. 2018, 19, 154. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Wang, W.; Mo, J.; Liu, F.; Zhang, H.; Liu, Y.; Chen, X.; Liao, Z.; Zhang, B.; Zhu, P. Long Noncoding RNA HOXC-AS3 Interacts with CDK2 to Promote Proliferation in Hepatocellular Carcinoma. Biomark. Res. 2022, 10, 65. [Google Scholar] [CrossRef]
- Cai, S.; Zhou, Y.; Pan, Y.; Liu, P.; Yu, K.; Chen, S. Long Non-Coding RNA A1BG-AS1 Promotes Tumorigenesis in Breast Cancer by Sponging MicroRNA-485-5p and Consequently Increasing Expression of FLOT1 Expression. Hum. Cell 2021, 34, 1517–1531. [Google Scholar] [CrossRef]
- Bai, J.; Yao, B.; Wang, L.; Sun, L.; Chen, T.; Liu, R.; Yin, G.; Xu, Q.; Yang, W. LncRNA A1BG-AS1 Suppresses Proliferation and Invasion of Hepatocellular Carcinoma Cells by Targeting MiR-216a-5p. J. Cell. Biochem. 2019, 120, 10310–10322. [Google Scholar] [CrossRef]
- Yao, J.; Chen, X.; Liu, X.; Li, R.; Zhou, X.; Qu, Y. Characterization of a Ferroptosis and Iron-Metabolism Related LncRNA Signature in Lung Adenocarcinoma. Cancer Cell Int. 2021, 21, 340. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhao, J.; Liu, J.N.; Sun, C. Mechanism and Role of the Neuropeptide LGI1 Receptor ADAM23 in Regulating Biomarkers of Ferroptosis and Progression of Esophageal Cancer. Dis. Markers 2021, 2021, 9227897. [Google Scholar] [CrossRef] [PubMed]
- Zhi, Y.; Du, J.; Qian, M.; Song, N. Long Non-Coding RNA RP11-342L8.2, Derived from RNA Sequencing and Validated via RT-QPCR, Is Upregulated and Correlates with Disease Severity in Psoriasis Patients. Ir. J. Med. Sci. 2022, 191, 2643–2649. [Google Scholar] [CrossRef]
- Yuan, H.; Ren, Q.; Du, Y.; Ma, Y.; Gu, L.; Zhou, J.; Tian, W.; Deng, D. LncRNA MiR663AHG Represses the Development of Colon Cancer in a MiR663a-Dependent Manner. Cell Death Discov. 2023, 9, 220. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Q.; Lou, Q.; Zhang, X.; Cui, Y.; Wang, P.; Yang, F.; Wu, F.; Wang, J.; Fan, T.; et al. Differential Analysis of LncRNA, MiRNA and MRNA Expression Profiles and the Prognostic Value of LncRNA in Esophageal Cancer. Pathol. Oncol. Res. 2020, 26, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhu, J.; Zhong, Y.; Liu, C.; Yao, M.; Sun, Y.; Yao, W.; Ni, X.; Zhou, F.; Yao, J.; et al. Targeting Long Noncoding RNA-AQP4-AS1 for the Treatment of Retinal Neurovascular Dysfunction in Diabetes Mellitus. EBioMedicine 2022, 77, 103857. [Google Scholar] [CrossRef] [PubMed]
- Andrew, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 25 August 2023).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Garrison, E.; Marth, G. Haplotype-Based Variant Detection from Short-Read Sequencing. 2012. Available online: https://arxiv.org/abs/1207.3907 (accessed on 25 August 2023).
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef]
- Auton, A.; Abecasis, G.R.; Altshuler, D.M.; Durbin, R.M.; Bentley, D.R.; Chakravarti, A.; Clark, A.G.; Donnelly, P.; Eichler, E.E.; Flicek, P.; et al. A Global Reference for Human Genetic Variation. Nature 2015, 526, 68–74. [Google Scholar]
- Platts, A.E.; Dix, D.J.; Chemes, H.E.; Thompson, K.E.; Goodrich, R.; Rockett, J.C.; Rawe, V.Y.; Quintana, S.; Diamond, M.P.; Strader, L.F.; et al. Success and Failure in Human Spermatogenesis as Revealed by Teratozoospermic RNAs. Hum. Mol. Genet. 2007, 16, 763–773. [Google Scholar] [CrossRef]
- Srinivasan, S.; Clements, J.A.; Batra, J. Single Nucleotide Polymorphisms in Clinics: Fantasy or Reality for Cancer? Crit. Rev. Clin. Lab. Sci. 2016, 53, 29–39. [Google Scholar] [CrossRef]
- Kyrgiafini, M.A.; Sarafidou, T.; Giannoulis, T.; Chatziparasidou, A.; Christoforidis, N.; Mamuris, Z. Gene-by-Sex Interactions: Genome-Wide Association Study Reveals Five SNPs Associated with Obesity and Overweight in a Male Population. Genes 2023, 14, 799. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Huang, H.; Gao, Y.; Huang, Q. Functional Annotation of Alzheimer’s Disease Associated Loci Revealed by GWASs. PLoS ONE 2017, 12, e0179677. [Google Scholar] [CrossRef]
- Shahin, M.H.; Sá, A.C.; Webb, A.; Gong, Y.; Langaee, T.; McDonough, C.W.; Riva, A.; Beitleshees, A.L.; Chapman, A.B.; Gums, J.G.; et al. Genome-Wide Prioritization and Transcriptomics Reveal Novel Signatures Associated with Thiazide Diuretics Blood Pressure Response. Circ. Cardiovasc. Genet. 2017, 10, e001404. [Google Scholar] [CrossRef] [PubMed]
- Mesbah-Uddin, M.; Elango, R.; Banaganapalli, B.; Shaik, N.A.; Al-Abbasi, F.A. In-Silico Analysis of Inflammatory Bowel Disease (IBD) GWAS Loci to Novel Connections. PLoS ONE 2015, 10, e0119420. [Google Scholar] [CrossRef] [PubMed]
- Sabarinathan, R.; Tafer, H.; Seemann, S.E.; Hofacker, I.L.; Stadler, P.F.; Gorodkin, J. RNAsnp: Efficient Detection of Local RNA Secondary Structure Changes Induced by SNPs. Hum. Mutat. 2013, 34, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Jung, D.; Jung, D.; Yao, R. ShinyGO: A Graphical Gene-Set Enrichment Tool for Animals and Plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Variant | lncRNA | 3DSNP Score | RegulomeDB Rank |
|---|---|---|---|
| rs11170045 | LINC00592 | 42.35 | 1b |
| rs17126450 | LINC00592 | 34.82 | 1b |
| rs7972661 | LINC00592 | 28.40 | 1f |
| rs936329 | LINC00592 | 26.09 | 1f |
| rs1870213 | LINC00592 | 27.76 | 1f |
| rs143637901 | LINC00592 | 27.63 | 1f |
| rs7301136 | LINC00592 | 23.79 | 1f |
| rs7315889 | LINC00592 | 44.57 | 1b |
| rs10783499 | LINC00592 | 45.49 | 1f |
| rs7316130 | LINC00592 | 45.37 | 1f |
| rs10876245 | LINC00592 | 46.98 | 1f |
| rs10876246 | LINC00592 | 14.97 | 1f |
| rs10876247 | LINC00592 | 16.98 | 1f |
| rs10876248 | LINC00592 | 12.32 | 1f |
| rs10783500 | LINC00592 | 12.40 | 1f |
| rs7303604 | LINC00592 | 10.60 | 1f |
| rs10783503 | LINC00592 | 25.87 | 1f |
| rs10783504 | LINC00592 | 125.41 | 1f |
| rs11170048 | LINC00592 | 125.70 | 1f |
| rs11170049 | LINC00592 | 125.39 | 1f |
| rs11170050 | LINC00592 | 123.71 | 1f |
| rs56035420 | HOXC-AS3 | 48.25 | 1b |
| rs1956568 | FRMD6-AS1 | 25.01 | 1b |
| rs1956567 | FRMD6-AS1 | 22.74 | 1f |
| rs566495825 | FRMD6-AS1 | 16.98 | 2b |
| rs71266965 | AQP4-AS1 | 23.77 | 2b |
| rs953369619 | AQP4-AS1 | 38.38 | 2b |
| rs1265960 | A1BG-AS1 | 23.04 | 1f |
| rs56822355 | LINC01350 | 136.62 | 1b |
| rs41453048 | LINC01350 | 18.39 | 1b |
| rs12029108 | LINC01350 | 16.82 | 1f |
| rs72727706 | LINC01350 | 12.63 | 1f |
| rs80202485 | LINC00466 | 126.85 | 1f |
| rs72781367 | LINC00276 | 11.09 | 1b |
| rs2969359 | LINC01116 | 84.20 | 1b |
| rs2969358 | LINC01116 | 84.25 | 1f |
| rs2969357 | LINC01116 | 74.60 | 1f |
| rs6723379 | DCTN1-AS1 | 44.15 | 1f |
| rs1727884 | ARHGEF26-AS1 | 18.68 | 1f |
| rs1713828 | ARHGEF26-AS1 | 55.43 | 1f |
| rs1713827 | ARHGEF26-AS1 | 125.75 | 1f |
| rs71281391 | PRICKLE2-AS3 | 23.94 | 2b |
| rs901028815 | LINC00877 | 11.67 | 2b |
| rs59058260 | LINC00877 | 15.06 | 2a |
| rs10016533 | SEC24B-AS1 | 111.37 | 1f |
| rs553142360 | LINC01091 | 56.16 | 2b |
| rs17491036 | LINC01091 | 24.33 | 2b |
| rs1560738 | LINC01091 | 21.52 | 1b |
| rs58833496 | LINC01091 | 10.36 | 1f |
| rs78146337 | NNT-AS1 | 12.66 | 1b |
| rs3734931 | C7orf69 | 50.17 | 2b |
| rs4595031 | C7orf69 | 10.44 | 1f |
| rs7823346 | ZNF252P-AS1 | 205.05 | 2b |
| rs2978419 | ZNF252P-AS1 | 106.11 | 1f |
| rs7832026 | ZNF252P-AS1 | 54.79 | 1f |
| rs10107110 | LZTS1-AS1 | 20.92 | 1b |
| rs12681283 | STAU2-AS1 | 52.30 | 2b |
| rs199889712 | OTUD6B-AS1 | 75.01 | 1b |
| rs3217986 | CDKN2B-AS1 | 75.44 | 2b |
| rs1368574245 | CDKN2B-AS1 | 92.75 | 2b |
| rs73652847 | CDKN2B-AS1 | 12.19 | 2b |
| rs17694555 | CDKN2B-AS1 | 14.83 | 1f |
| rs564311323 | CDKN2B-AS1 | 17.41 | 2b |
| rs112111321 | RAP2C-AS1 | 216.01 | 1a |
| Variant | lncRNA | Transcript | p-Value |
|---|---|---|---|
| rs139506376 | LINC00944 | NONHSAT031816.2 | 0.1688 |
| rs200712585 | PRMT5-AS1 | NONHSAT035833.2 | 0.0739 |
| PRMT5-AS1 | NONHSAT168171.1 | 0.0814 | |
| PRMT5-AS1 | NONHSAT168172.1 | 0.1409 | |
| PRMT5-AS1 | NONHSAT035834.2 | 0.0818 | |
| rs4792409 | PRMT5-AS1 | NONHSAT145734.2 | 0.1078 |
| rs150364102 | COX10-AS1 | NONHSAT145753.2 | 0.0946 |
| COX10-AS1 | NONHSAT145752.2 | 0.0946 | |
| COX10-AS1 | NONHSAT145754.2 | 0.0946 | |
| COX10-AS1 | NONHSAT145751.2 | 0.0921 | |
| COX10-AS1 | NONHSAT175957.1 | 0.0946 | |
| COX10-AS1 | NONHSAT175958.1 | 0.0946 | |
| rs75133618 | COX10-AS1 | NONHSAT175957.1 | 0.1591 |
| COX10-AS1 | NONHSAT175958.1 | 0.1591 | |
| rs142992890 | MIR663AHG | NONHSAT189547.1 | 0.1669 |
| rs1285843394 | MIR663AHG | NONHSAT189547.1 | 0.1941 |
| Variants | lncRNAs | Impact on lncRNAs |
|---|---|---|
| rs150364102 | COX10-AS1 | Structure and miRNA–lncRNA interactions |
| rs56035420 | HOXC-AS3 | Function and miRNA–lncRNA interactions |
| rs2969359 | LINC01116 | Function and miRNA–lncRNA interactions |
| rs2969358 | LINC01116 | Function and miRNA–lncRNA interactions |
| rs2969357 | LINC01116 | Function and miRNA–lncRNA interactions |
| rs1713827 | ARHGEF26-AS1 | Function and miRNA–lncRNA interactions |
| lncRNAs | Number of Variants | Impact of Variants |
|---|---|---|
| COX10-AS1 | 7 | Structure and miRNAs–lncRNAs |
| MIR663AHG | 6 | Structure and miRNAs–lncRNAs |
| HOXC-AS3 | 2 | Function and miRNAs–lncRNAs |
| AQP4-AS1 | 5 | Function and miRNAs–lncRNAs |
| A1BG-AS1 | 6 | Function and miRNAs–lncRNAs |
| LINC01116 | 7 | Function and miRNAs–lncRNAs |
| DCTN1-AS1 | 2 | Function and miRNAs–lncRNAs |
| ARHGEF26-AS1 | 6 | Function and miRNAs–lncRNAs |
| PRICKLE2-AS3 | 2 | Function and miRNAs–lncRNAs |
| STAU2-AS1 | 2 | Function and miRNAs–lncRNAs |
| RAP2C-AS1 | 2 | Function and miRNAs–lncRNAs |
| CDKN2B-AS1 | 7 | Function and miRNAs–lncRNAs |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyrgiafini, M.-A.; Giannoulis, T.; Chatziparasidou, A.; Christoforidis, N.; Mamuris, Z. Unveiling the Genetic Complexity of Teratozoospermia: Integrated Genomic Analysis Reveals Novel Insights into lncRNAs’ Role in Male Infertility. Int. J. Mol. Sci. 2023, 24, 15002. https://doi.org/10.3390/ijms241915002
Kyrgiafini M-A, Giannoulis T, Chatziparasidou A, Christoforidis N, Mamuris Z. Unveiling the Genetic Complexity of Teratozoospermia: Integrated Genomic Analysis Reveals Novel Insights into lncRNAs’ Role in Male Infertility. International Journal of Molecular Sciences. 2023; 24(19):15002. https://doi.org/10.3390/ijms241915002
Chicago/Turabian StyleKyrgiafini, Maria-Anna, Themistoklis Giannoulis, Alexia Chatziparasidou, Nikolaos Christoforidis, and Zissis Mamuris. 2023. "Unveiling the Genetic Complexity of Teratozoospermia: Integrated Genomic Analysis Reveals Novel Insights into lncRNAs’ Role in Male Infertility" International Journal of Molecular Sciences 24, no. 19: 15002. https://doi.org/10.3390/ijms241915002
APA StyleKyrgiafini, M.-A., Giannoulis, T., Chatziparasidou, A., Christoforidis, N., & Mamuris, Z. (2023). Unveiling the Genetic Complexity of Teratozoospermia: Integrated Genomic Analysis Reveals Novel Insights into lncRNAs’ Role in Male Infertility. International Journal of Molecular Sciences, 24(19), 15002. https://doi.org/10.3390/ijms241915002