
Facade-Based Bicelles as a New Tool for Production of Active Membrane Proteins in a Cell-Free System

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
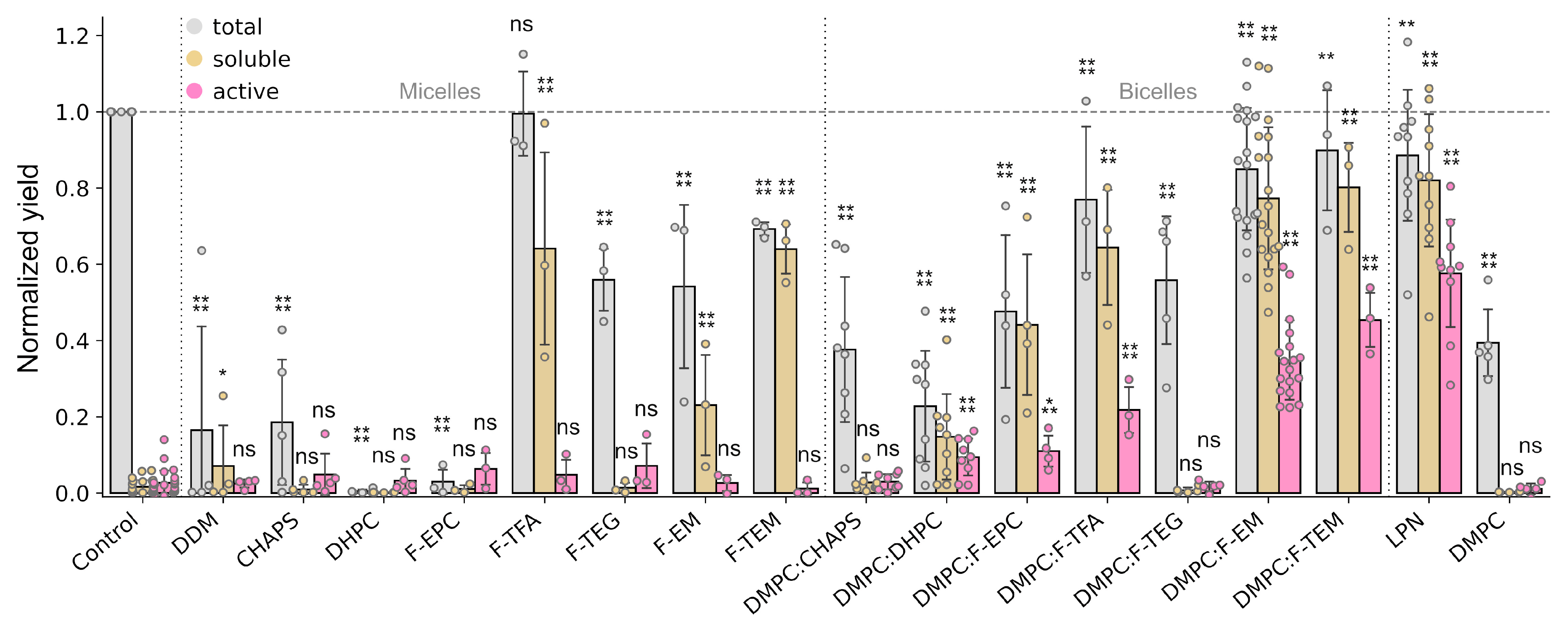
2.1. Facade-Based Bicelles Reveal Prospects as a Membrane Mimetic Additive for Cell-Free Systems
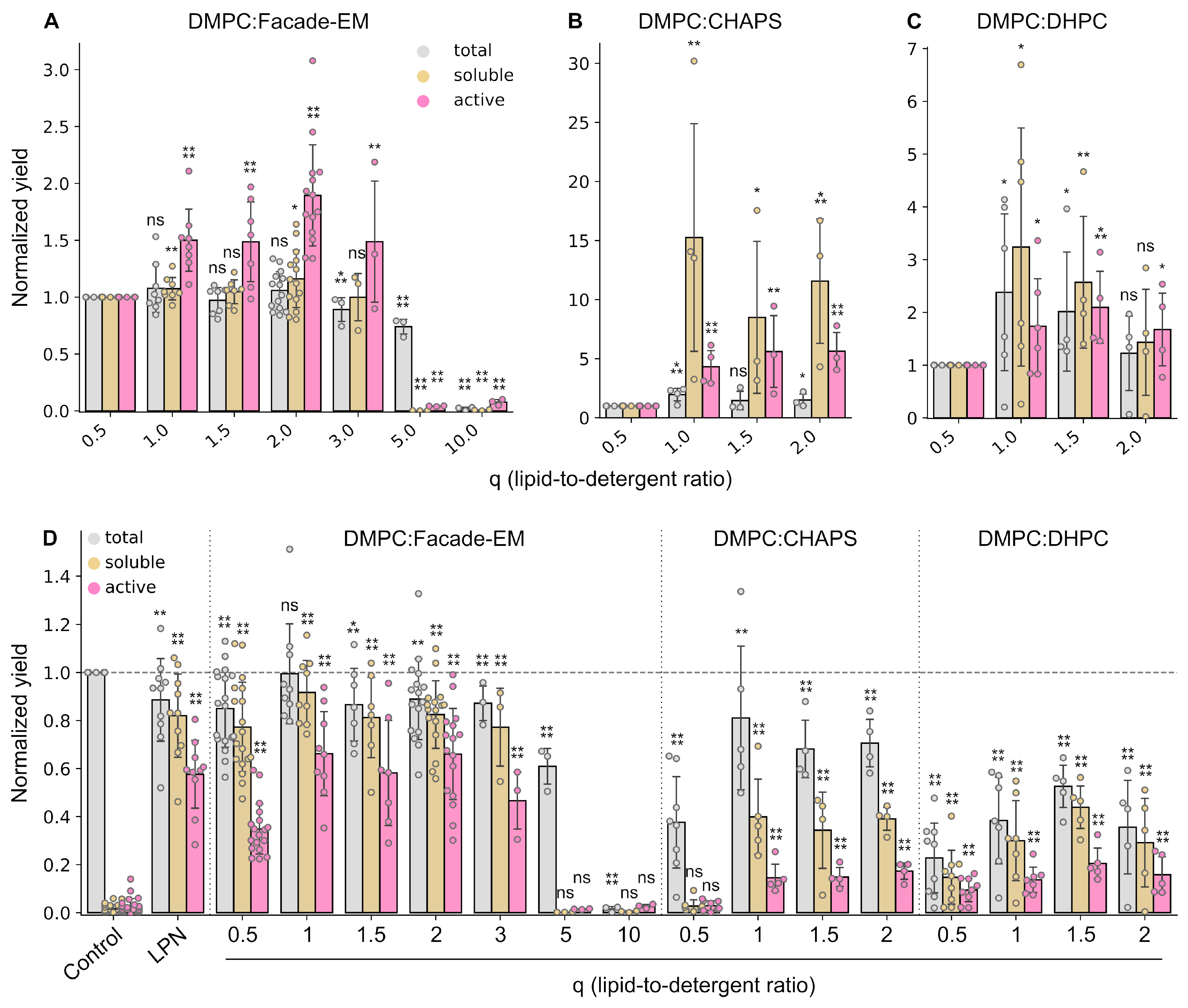
2.2. Size of Bicelles Is an Important Parameter of a Cell-Free Reaction
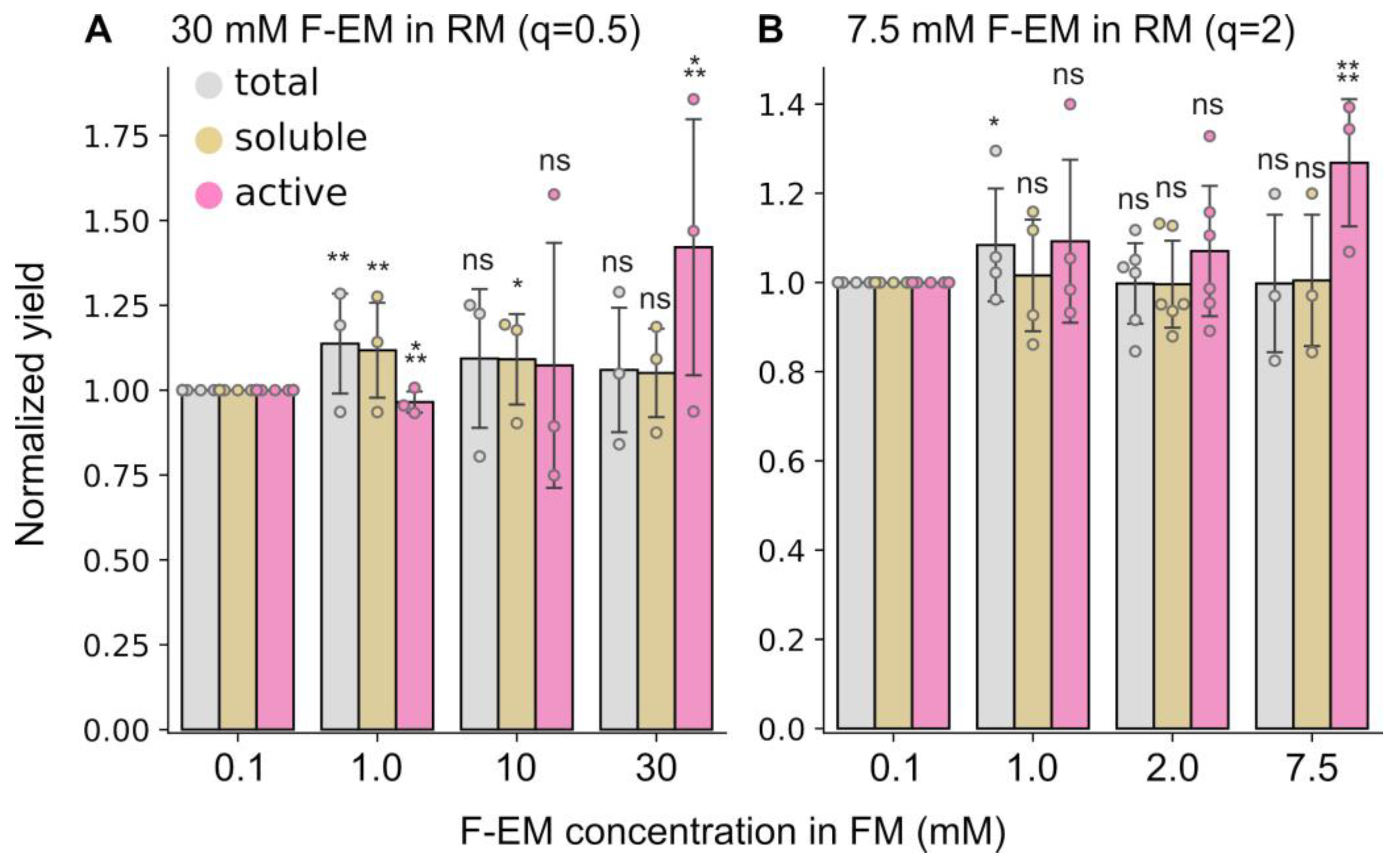
2.3. The Detergent Concentration in the Feeding Mixture Has a Limited Influence on the Yield of Active Protein
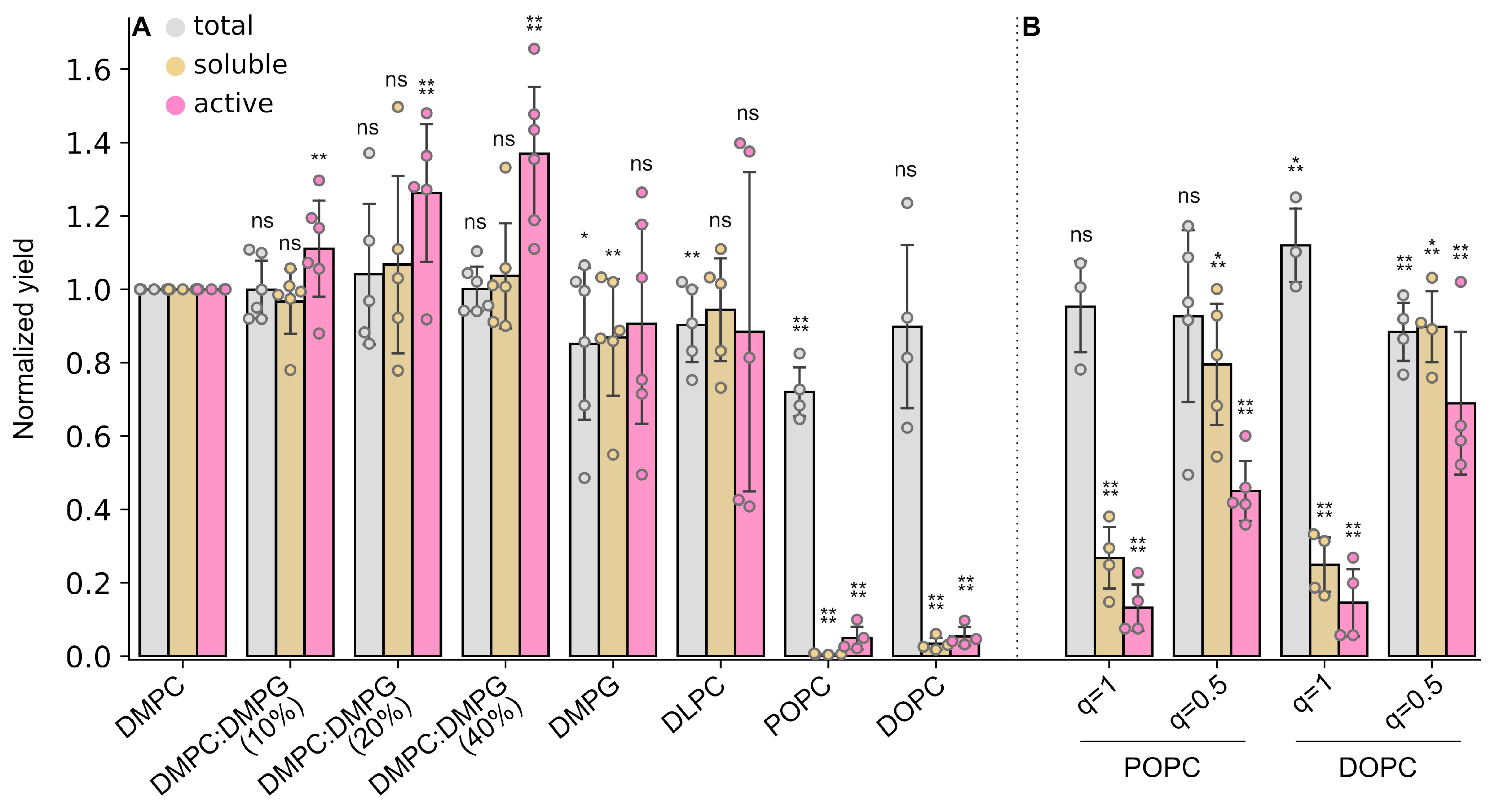
2.4. Facade-EM Is a Versatile Rim-Forming Detergent for Different Lipids
2.5. Facade-EM Bicelles Are the Most Promising Additives for the Co-Translational Folding of Membrane Proteins
3. Discussion
4. Materials and Methods
4.1. Cell-Free Synthesis and Sample Preparation
4.2. Quantification of ESR Yields
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Wong, T.-S.; Li, G.; Li, S.; Gao, W.; Chen, G.; Gan, S.; Zhang, M.; Li, H.; Wu, S.; Du, Y. G Protein-Coupled Receptors in Neurodegenerative Diseases and Psychiatric Disorders. Signal Transduct. Target. Ther. 2023, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Marinko, J.T.; Huang, H.; Penn, W.D.; Capra, J.A.; Schlebach, J.P.; Sanders, C.R. Folding and Misfolding of Human Membrane Proteins in Health and Disease: From Single Molecules to Cellular Proteostasis. Chem. Rev. 2019, 119, 5537–5606. [Google Scholar] [CrossRef] [PubMed]
- Meldolesi, J. Neurotrophin Receptors in the Pathogenesis, Diagnosis and Therapy of Neurodegenerative Diseases. Pharmacol. Res. 2017, 121, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, S.; Han, J.; Li, S.; Gao, X.; Wang, M.; Zhu, J.; Jin, T. Role of Toll-Like Receptors in Neuroimmune Diseases: Therapeutic Targets and Problems. Front. Immunol. 2021, 12, 777606. [Google Scholar] [CrossRef]
- He, Y.; He, G.; He, T. Specifically Targeted Transport of Plasma Membrane Transporters: From Potential Mechanisms for Regulating Cell Health or Disease to Applications. Membranes 2021, 11, 736. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-R.; Jung, S.; Ryu, H.; Yoo, Y.-E.; Kim, S.M.; Jeon, T.-J. Synthetic Biomimetic Membranes and Their Sensor Applications. Sensors 2012, 12, 9530–9550. [Google Scholar] [CrossRef]
- Santos, R.; Ursu, O.; Gaulton, A.; Bento, A.P.; Donadi, R.S.; Bologa, C.G.; Karlsson, A.; Al-Lazikani, B.; Hersey, A.; Oprea, T.I.; et al. A Comprehensive Map of Molecular Drug Targets. Nat. Rev. Drug Discov. 2017, 16, 19–34. [Google Scholar] [CrossRef]
- Overington, J.P.; Al-Lazikani, B.; Hopkins, A.L. How Many Drug Targets Are There? Nat. Rev. Drug Discov. 2006, 5, 993–996. [Google Scholar] [CrossRef]
- Birch, J.; Cheruvara, H.; Gamage, N.; Harrison, P.J.; Lithgo, R.; Quigley, A. Changes in Membrane Protein Structural Biology. Biology 2020, 9, 401. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Shin, K.; Patterson, R.E.; Liu, X.-Q.; Rainey, J.K. Current Strategies for Protein Production and Purification Enabling Membrane Protein Structural Biology. Biochem. Cell Biol. 2016, 94, 507–527. [Google Scholar] [CrossRef]
- Motov, V.V.; Kot, E.F.; Shabalkina, A.V.; Goncharuk, S.A.; Arseniev, A.S.; Goncharuk, M.V.; Mineev, K.S. Investigation of Lipid/Protein Interactions in Trifluoroethanol-Water Mixtures Proposes the Strategy for the Refolding of Helical Transmembrane Domains. J. Biomol. NMR 2023, 77, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Barrera, F.N.; Renart, M.L.; Molina, M.L.; Poveda, J.A.; Encinar, J.A.; Fernández, A.M.; Neira, J.L.; González-Ros, J.M. Unfolding and Refolding in Vitro of a Tetrameric, Alpha-Helical Membrane Protein: The Prokaryotic Potassium Channel KcsA. Biochemistry 2005, 44, 14344–14352. [Google Scholar] [CrossRef] [PubMed]
- Gorzelle, B.M.; Nagy, J.K.; Oxenoid, K.; Lonzer, W.L.; Cafiso, D.S.; Sanders, C.R. Reconstitutive Refolding of Diacylglycerol Kinase, an Integral Membrane Protein. Biochemistry 1999, 38, 16373–16382. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Junge, F.; Durst, F.; Frölich, N.; Schneider, B.; Reckel, S.; Sobhanifar, S.; Dötsch, V.; Bernhard, F. Preparative Scale Expression of Membrane Proteins in Escherichia Coli-Based Continuous Exchange Cell-Free Systems. Nat. Protoc. 2007, 2, 2945–2957. [Google Scholar] [CrossRef] [PubMed]
- Pardee, K.; Slomovic, S.; Nguyen, P.Q.; Lee, J.W.; Donghia, N.; Burrill, D.; Ferrante, T.; McSorley, F.R.; Furuta, Y.; Vernet, A.; et al. Portable, On-Demand Biomolecular Manufacturing. Cell 2016, 167, 248–259.e12. [Google Scholar] [CrossRef]
- Henrich, E.; Hein, C.; Dötsch, V.; Bernhard, F. Membrane Protein Production in Escherichia Coli Cell-Free Lysates. FEBS Lett. 2015, 589, 1713–1722. [Google Scholar] [CrossRef] [PubMed]
- Manzer, Z.A.; Selivanovitch, E.; Ostwalt, A.R.; Daniel, S. Membrane Protein Synthesis: No Cells Required. Trends Biochem. Sci. 2023, 48, 642–654. [Google Scholar] [CrossRef]
- Sachse, R.; Dondapati, S.K.; Fenz, S.F.; Schmidt, T.; Kubick, S. Membrane Protein Synthesis in Cell-Free Systems: From Bio-Mimetic Systems to Bio-Membranes. FEBS Lett. 2014, 588, 2774–2781. [Google Scholar] [CrossRef]
- Kuruma, Y.; Ueda, T. The PURE System for the Cell-Free Synthesis of Membrane Proteins. Nat. Protoc. 2015, 10, 1328–1344. [Google Scholar] [CrossRef] [PubMed]
- Ezure, T.; Nanatani, K.; Sato, Y.; Suzuki, S.; Aizawa, K.; Souma, S.; Ito, M.; Hohsaka, T.; von Heijine, G.; Utsumi, T.; et al. A Cell-Free Translocation System Using Extracts of Cultured Insect Cells to Yield Functional Membrane Proteins. PLoS ONE 2014, 9, e112874. [Google Scholar] [CrossRef]
- Wuu, J.J.; Swartz, J.R. High Yield Cell-Free Production of Integral Membrane Proteins without Refolding or Detergents. Biochim. Biophys. Acta BBA Biomembr. 2008, 1778, 1237–1250. [Google Scholar] [CrossRef] [PubMed]
- Lyukmanova, E.N.; Shenkarev, Z.O.; Khabibullina, N.F.; Kopeina, G.S.; Shulepko, M.A.; Paramonov, A.S.; Mineev, K.S.; Tikhonov, R.V.; Shingarova, L.N.; Petrovskaya, L.E.; et al. Lipid-Protein Nanodiscs for Cell-Free Production of Integral Membrane Proteins in a Soluble and Folded State: Comparison with Detergent Micelles, Bicelles and Liposomes. Biochim. Biophys. Acta BBA Biomembr. 2012, 1818, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Bruni, R.; Laguerre, A.; Kaminska, A.-M.; McSweeney, S.; Hendrickson, W.A.; Liu, Q. High-Throughput Cell-Free Screening of Eukaryotic Membrane Protein Expression in Lipidic Mimetics. Protein Sci. 2022, 31, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Nakai, H.; Isshiki, K.; Hattori, M.; Maehira, H.; Yamaguchi, T.; Masuda, K.; Shimizu, Y.; Watanabe, T.; Hohsaka, T.; Shihoya, W.; et al. Cell-Free Synthesis of Human Endothelin Receptors and Its Application to Ribosome Display. Anal. Chem. 2022, 94, 3831–3839. [Google Scholar] [CrossRef]
- Kornilov, F.D.; Shabalkina, A.V.; Lin, C.; Volynsky, P.E.; Kot, E.F.; Kayushin, A.L.; Lushpa, V.A.; Goncharuk, M.V.; Arseniev, A.S.; Goncharuk, S.A.; et al. The Architecture of Transmembrane and Cytoplasmic Juxtamembrane Regions of Toll-like Receptors. Nat. Commun. 2023, 14, 1503. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, J.; Ohlhoff, C.; Dondapati, S.K.; Zemella, A.; Kubick, S. Evaluation of the Ion Channel Assembly in a Eukaryotic Cell-Free System Focusing on Two-Pore Domain Potassium Channels K2P. Int. J. Mol. Sci. 2023, 24, 6299. [Google Scholar] [CrossRef]
- Cuevas Arenas, R.; Danielczak, B.; Martel, A.; Porcar, L.; Breyton, C.; Ebel, C.; Keller, S. Fast Collisional Lipid Transfer among Polymer-Bounded Nanodiscs. Sci. Rep. 2017, 7, 45875. [Google Scholar] [CrossRef]
- Köck, Z.; Ermel, U.; Martin, J.; Morgner, N.; Frangakis, A.S.; Dötsch, V.; Hilger, D.; Bernhard, F. Biochemical Characterization of Cell-Free Synthesized Human Β1 Adrenergic Receptor Cotranslationally Inserted into Nanodiscs. J. Mol. Biol. 2022, 434, 167687. [Google Scholar] [CrossRef]
- Zhang, Q.; Ma, X.; Ward, A.; Hong, W.-X.; Jaakola, V.-P.; Stevens, R.C.; Finn, M.G.; Chang, G. Designing Facial Amphiphiles for the Stabilization of Integral Membrane Proteins. Angew. Chem. Int. Ed Engl. 2007, 46, 7023–7025. [Google Scholar] [CrossRef]
- Lee, S.C.; Bennett, B.C.; Hong, W.-X.; Fu, Y.; Baker, K.A.; Marcoux, J.; Robinson, C.V.; Ward, A.B.; Halpert, J.R.; Stevens, R.C.; et al. Steroid-Based Facial Amphiphiles for Stabilization and Crystallization of Membrane Proteins. Proc. Natl. Acad. Sci. USA 2013, 110, E1203–E1211. [Google Scholar] [CrossRef]
- Khan, A.K.; Jagielnicki, M.; Bennett, B.C.; Purdy, M.D.; Yeager, M. Cryo-EM Structure of an Open Conformation of a Gap Junction Hemichannel in Lipid Bilayer Nanodiscs. Struct. Lond. Engl. 1993 2021, 29, 1040–1047.e3. [Google Scholar] [CrossRef]
- Hildebrandt, E.; Zhang, Q.; Cant, N.; Ding, H.; Dai, Q.; Peng, L.; Fu, Y.; DeLucas, L.J.; Ford, R.; Kappes, J.C.; et al. A Survey of Detergents for the Purification of Stable, Active Human Cystic Fibrosis Transmembrane Conductance Regulator (CFTR). Biochim. Biophys. Acta BBA Biomembr. 2014, 1838, 2825–2837. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.; Miu, A.; Alexander, M.K.; Garcia, N.K.; Oh, A.; Zilberleyb, I.; Reichelt, M.; Austin, C.D.; Tam, C.; Shriver, S.; et al. Structural Basis for Dual-Mode Inhibition of the ABC Transporter MsbA. Nature 2018, 557, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Padayatti, P.S.; Lee, S.C.; Stanfield, R.L.; Wen, P.-C.; Tajkhorshid, E.; Wilson, I.A.; Zhang, Q. Structural Insights into the Lipid a Transport Pathway in MsbA. Structure 2019, 27, 1114–1123.e3. [Google Scholar] [CrossRef]
- Xu, H.; Li, T.; Rohou, A.; Arthur, C.P.; Tzakoniati, F.; Wong, E.; Estevez, A.; Kugel, C.; Franke, Y.; Chen, J.; et al. Structural Basis of Nav1.7 Inhibition by a Gating-Modifier Spider Toxin. Cell 2019, 176, 702–715.e14. [Google Scholar] [CrossRef] [PubMed]
- Mineev, K.S.; Nadezhdin, K.D.; Goncharuk, S.A.; Arseniev, A.S. Façade Detergents as Bicelle Rim-Forming Agents for Solution NMR Spectroscopy. Nanotechnol. Rev. 2016, 6, 93–103. [Google Scholar] [CrossRef]
- Kot, E.F.; Goncharuk, S.A.; Arseniev, A.S.; Mineev, K.S. Phase Transitions in Small Isotropic Bicelles. Langmuir 2018, 34, 3426–3437. [Google Scholar] [CrossRef]
- Petrovskaya, L.E.; Lukashev, E.P.; Chupin, V.V.; Sychev, S.V.; Lyukmanova, E.N.; Kryukova, E.A.; Ziganshin, R.H.; Spirina, E.V.; Rivkina, E.M.; Khatypov, R.A.; et al. Predicted Bacteriorhodopsin from Exiguobacterium Sibiricum Is a Functional Proton Pump. FEBS Lett. 2010, 584, 4193–4196. [Google Scholar] [CrossRef] [PubMed]
- Petrovskaya, L.E.; Balashov, S.P.; Lukashev, E.P.; Imasheva, E.S.; Gushchin, I.Y.; Dioumaev, A.K.; Rubin, A.B.; Dolgikh, D.A.; Gordeliy, V.I.; Lanyi, J.K.; et al. ESR—A Retinal Protein with Unusual Properties from Exiguobacterium Sibiricum. Biochem. Biokhimiia 2015, 80, 688–700. [Google Scholar] [CrossRef]
- Klammt, C.; Schwarz, D.; Fendler, K.; Haase, W.; Dötsch, V.; Bernhard, F. Evaluation of Detergents for the Soluble Expression of α-Helical and β-Barrel-Type Integral Membrane Proteins by a Preparative Scale Individual Cell-Free Expression System. FEBS J. 2005, 272, 6024–6038. [Google Scholar] [CrossRef] [PubMed]
- Mineev, K.S.; Nadezhdin, K.D.; Goncharuk, S.A.; Arseniev, A.S. Characterization of Small Isotropic Bicelles with Various Compositions. Langmuir 2016, 32, 6624–6637. [Google Scholar] [CrossRef]
- Beaugrand, M.; Arnold, A.A.; Hénin, J.; Warschawski, D.E.; Williamson, P.T.F.; Marcotte, I. Lipid Concentration and Molar Ratio Boundaries for the Use of Isotropic Bicelles. Langmuir 2014, 30, 6162–6170. [Google Scholar] [CrossRef]
- Caldwell, T.A.; Baoukina, S.; Brock, A.T.; Oliver, R.C.; Root, K.T.; Krueger, J.K.; Glover, K.J.; Tieleman, D.P.; Columbus, L. Low- q Bicelles Are Mixed Micelles. J. Phys. Chem. Lett. 2018, 9, 4469–4473. [Google Scholar] [CrossRef] [PubMed]
- Liebau, J.; Ye, W.; Mäler, L. Characterization of Fast-Tumbling Isotropic Bicelles by PFG Diffusion NMR. Magn. Reson. Chem. MRC 2017, 55, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Lind, J.; Eriksson, J.; Mäler, L. Characterization of the Morphology of Fast-Tumbling Bicelles with Varying Composition. Langmuir 2014, 30, 5488–5496. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Van Horn, W.D.; Chen, J.; Mathew, S.; Zent, R.; Sanders, C.R. Bicelles at Low Concentrations. Mol. Pharm. 2012, 9, 752–761. [Google Scholar] [CrossRef]
- Glover, K.J.; Whiles, J.A.; Wu, G.; Yu, N.; Deems, R.; Struppe, J.O.; Stark, R.E.; Komives, E.A.; Vold, R.R. Structural Evaluation of Phospholipid Bicelles for Solution-State Studies of Membrane-Associated Biomolecules. Biophys. J. 2001, 81, 2163–2171. [Google Scholar] [CrossRef]
- Bogdanov, M.; Dowhan, W.; Vitrac, H. Lipids and Topological Rules Governing Membrane Protein Assembly. Biochim. Biophys. Acta BBA Mol. Cell Res. 2014, 1843, 1475–1488. [Google Scholar] [CrossRef]
- Subramaniam, S.; Greenhalgh, D.A.; Rath, P.; Rothschild, K.J.; Khorana, H.G. Replacement of Leucine-93 by Alanine or Threonine Slows down the Decay of the N and O Intermediates in the Photocycle of Bacteriorhodopsin: Implications for Proton Uptake and 13-Cis-Retinal----All-Trans-Retinal Reisomerization. Proc. Natl. Acad. Sci. USA 1991, 88, 6873–6877. [Google Scholar] [CrossRef]
- Dioumaev, A.K.; Savransky, V.V.; Tkachenko, N.V.; Chukharev, V.I. Quantum Yield and Extinction Measurements in Strongly Overlapping Reactant and Photoproduct Absorption Bands: II: Bathointermediate Formation in Bacteriorhodopsin Photocycle at Room Temperature. J. Photochem. Photobiol. B 1989, 3, 397–410. [Google Scholar] [CrossRef]
- Kot, E.F.; Arseniev, A.S.; Mineev, K.S. Behavior of Most Widely Spread Lipids in Isotropic Bicelles. Langmuir 2018, 34, 8302–8313. [Google Scholar] [CrossRef] [PubMed]
- Denisov, I.G.; Sligar, S.G. Nanodiscs for Structural and Functional Studies of Membrane Proteins. Nat. Struct. Mol. Biol. 2016, 23, 481–486. [Google Scholar] [CrossRef]
- Katzen, F.; Fletcher, J.E.; Yang, J.-P.; Kang, D.; Peterson, T.C.; Cappuccio, J.A.; Blanchette, C.D.; Sulchek, T.; Chromy, B.A.; Hoeprich, P.D.; et al. Insertion of Membrane Proteins into Discoidal Membranes Using a Cell-Free Protein Expression Approach. J. Proteome Res. 2008, 7, 3535–3542. [Google Scholar] [CrossRef]
- Sligar, S.G.; Denisov, I.G. Nanodiscs: A Toolkit for Membrane Protein Science. Protein Sci. 2021, 30, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, A.; Shadiac, N.; Amalraj, A.; Garajová, S.; Nagarajan, Y.; Waters, S.; Mertens, H.D.T.; Hrmova, M. Cell-Free Protein Synthesis of Membrane (1,3)-β-d-Glucan (Curdlan) Synthase: Co-Translational Insertion in Liposomes and Reconstitution in Nanodiscs. Biochim. Biophys. Acta BBA Biomembr. 2013, 1828, 743–757. [Google Scholar] [CrossRef] [PubMed]
- Glueck, D.; Grethen, A.; Das, M.; Mmeka, O.P.; Patallo, E.P.; Meister, A.; Rajender, R.; Kins, S.; Räschle, M.; Victor, J.; et al. Electroneutral Polymer Nanodiscs Enable Interference-Free Probing of Membrane Proteins in a Lipid-Bilayer Environment. Small 2022, 18, 2202492. [Google Scholar] [CrossRef]
- Bragin, P.E.; Kuznetsov, A.S.; Bocharova, O.V.; Volynsky, P.E.; Arseniev, A.S.; Efremov, R.G.; Mineev, K.S. Probing the Effect of Membrane Contents on Transmembrane Protein-Protein Interaction Using Solution NMR and Computer Simulations. Biochim. Biophys. Acta BBA Biomembr. 2018, 1860, 2486–2498. [Google Scholar] [CrossRef]
- Kot, E.F.; Goncharuk, S.A.; Franco, M.L.; Arseniev, A.S.; Benito-Martínez, A.; Costa, M.; Cattaneo, A.; Vilar, M.; Mineev, K.S. Structural Basis for the Transmembrane Signaling and Antidepressant-Induced Activation of the Receptor Tyrosine Kinase TrkB|bioRxiv. Available online: https://www.biorxiv.org/content/10.1101/2023.06.06.543881v1 (accessed on 13 September 2023).
- Laguerre, A.; Löhr, F.; Henrich, E.; Hoffmann, B.; Abdul-Manan, N.; Connolly, P.J.; Perozo, E.; Moore, J.M.; Bernhard, F.; Dötsch, V. From Nanodiscs to Isotropic Bicelles: A Procedure for Solution NMR Studies of Detergent Sensitive Integral Membrane Proteins. Structure 2016, 24, 1830–1841. [Google Scholar] [CrossRef]
- Takeda, M.; Kainosho, M. Cell-Free Protein Synthesis Using E. Coli Cell Extract for NMR Studies. Adv. Exp. Med. Biol. 2012, 992, 167–177. [Google Scholar] [CrossRef]
- Bernhard, F.; Tozawa, Y. Cell-Free Expression—Making a Mark. Curr. Opin. Struct. Biol. 2013, 23, 374–380. [Google Scholar] [CrossRef]
- Pellowe, G.A.; Booth, P.J. Structural Insight into Co-Translational Membrane Protein Folding. Biochim. Biophys. Acta BBA Biomembr. 2020, 1862, 183019. [Google Scholar] [CrossRef]
- Harris, N.J.; Reading, E.; Booth, P.J. Cell-Free Synthesis Strategies to Probe Co-Translational Folding of Proteins Within Lipid Membranes. Methods Mol. Biol. Clifton NJ 2022, 2433, 273–292. [Google Scholar] [CrossRef]
- Mercier, E.; Wintermeyer, W.; Rodnina, M.V. Co-Translational Insertion and Topogenesis of Bacterial Membrane Proteins Monitored in Real Time. EMBO J. 2020, 39, e104054. [Google Scholar] [CrossRef]
- Niesen, M.J.M.; Zimmer, M.H.; Miller, T.F. Dynamics of Co-Translational Membrane Protein Integration and Translocation via the Sec Translocon. J. Am. Chem. Soc. 2020, 142, 5449–5460. [Google Scholar] [CrossRef]
- Tinafar, A.; Jaenes, K.; Pardee, K. Synthetic Biology Goes Cell-Free. BMC Biol. 2019, 17, 64. [Google Scholar] [CrossRef]
- Schneider, B.; Junge, F.; Shirokov, V.A.; Durst, F.; Schwarz, D.; Dötsch, V.; Bernhard, F. Membrane Protein Expression in Cell-Free Systems. Methods Mol. Biol. Clifton NJ 2010, 601, 165–186. [Google Scholar] [CrossRef]
- Hagn, F.; Etzkorn, M.; Raschle, T.; Wagner, G. Optimized Phospholipid Bilayer Nanodiscs Facilitate High-Resolution Structure Determination of Membrane Proteins. J. Am. Chem. Soc. 2013, 135, 1919–1925. [Google Scholar] [CrossRef]
- Zhang, Z.-M.; Chen, S.; Liang, Y.-Z. Baseline Correction Using Adaptive Iteratively Reweighted Penalized Least Squares. Analyst 2010, 135, 1138–1146. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goncharuk, M.V.; Vasileva, E.V.; Ananiev, E.A.; Gorokhovatsky, A.Y.; Bocharov, E.V.; Mineev, K.S.; Goncharuk, S.A. Facade-Based Bicelles as a New Tool for Production of Active Membrane Proteins in a Cell-Free System. Int. J. Mol. Sci. 2023, 24, 14864. https://doi.org/10.3390/ijms241914864
Goncharuk MV, Vasileva EV, Ananiev EA, Gorokhovatsky AY, Bocharov EV, Mineev KS, Goncharuk SA. Facade-Based Bicelles as a New Tool for Production of Active Membrane Proteins in a Cell-Free System. International Journal of Molecular Sciences. 2023; 24(19):14864. https://doi.org/10.3390/ijms241914864
Chicago/Turabian StyleGoncharuk, Marina V., Ekaterina V. Vasileva, Egor A. Ananiev, Andrey Y. Gorokhovatsky, Eduard V. Bocharov, Konstantin S. Mineev, and Sergey A. Goncharuk. 2023. "Facade-Based Bicelles as a New Tool for Production of Active Membrane Proteins in a Cell-Free System" International Journal of Molecular Sciences 24, no. 19: 14864. https://doi.org/10.3390/ijms241914864
APA StyleGoncharuk, M. V., Vasileva, E. V., Ananiev, E. A., Gorokhovatsky, A. Y., Bocharov, E. V., Mineev, K. S., & Goncharuk, S. A. (2023). Facade-Based Bicelles as a New Tool for Production of Active Membrane Proteins in a Cell-Free System. International Journal of Molecular Sciences, 24(19), 14864. https://doi.org/10.3390/ijms241914864