
Plant Cell-Engineered Gold Nanoparticles Conjugated to Quercetin Inhibit SARS-CoV-2 and HSV-1 Entry

 , ,
, ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
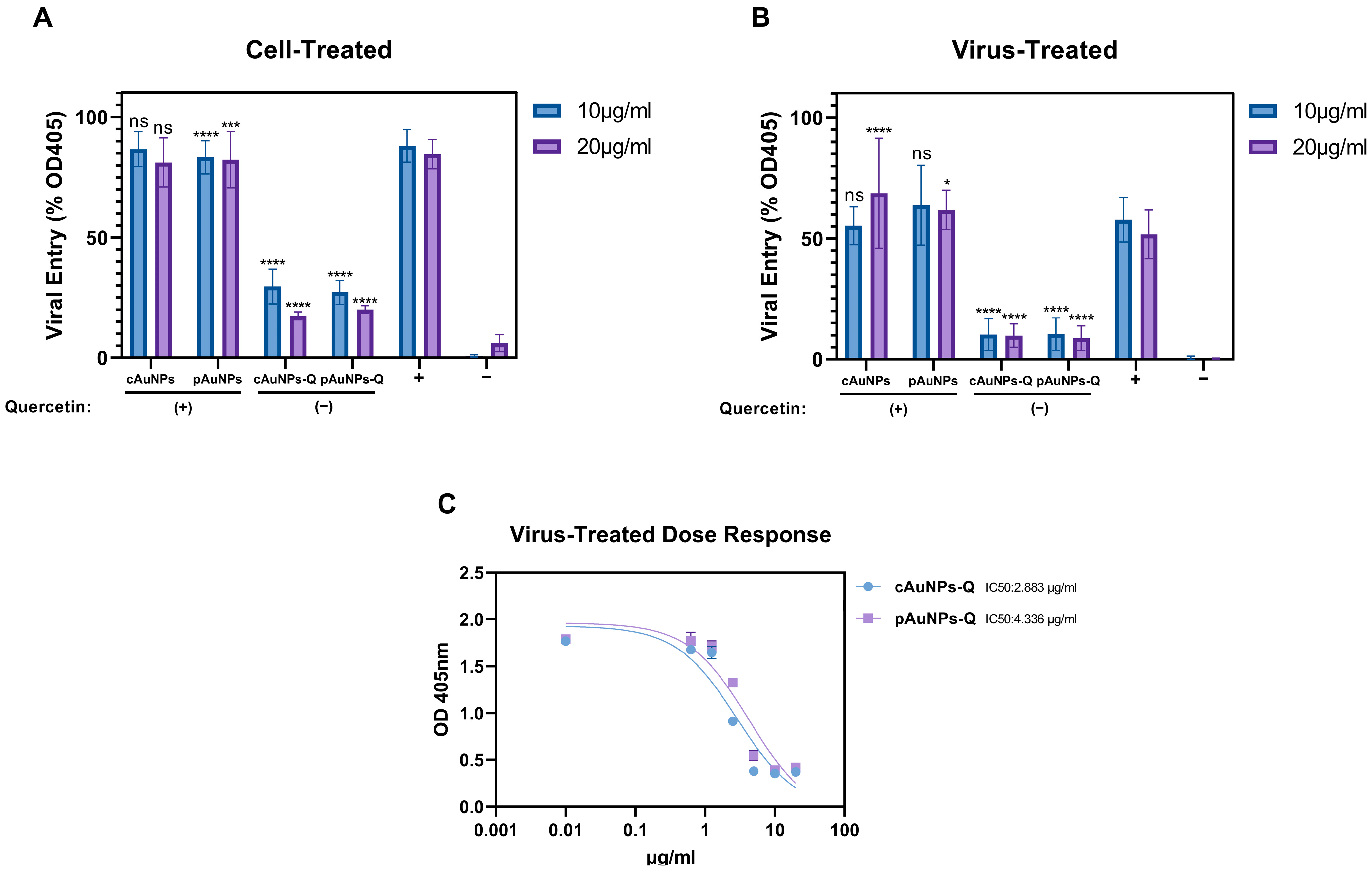
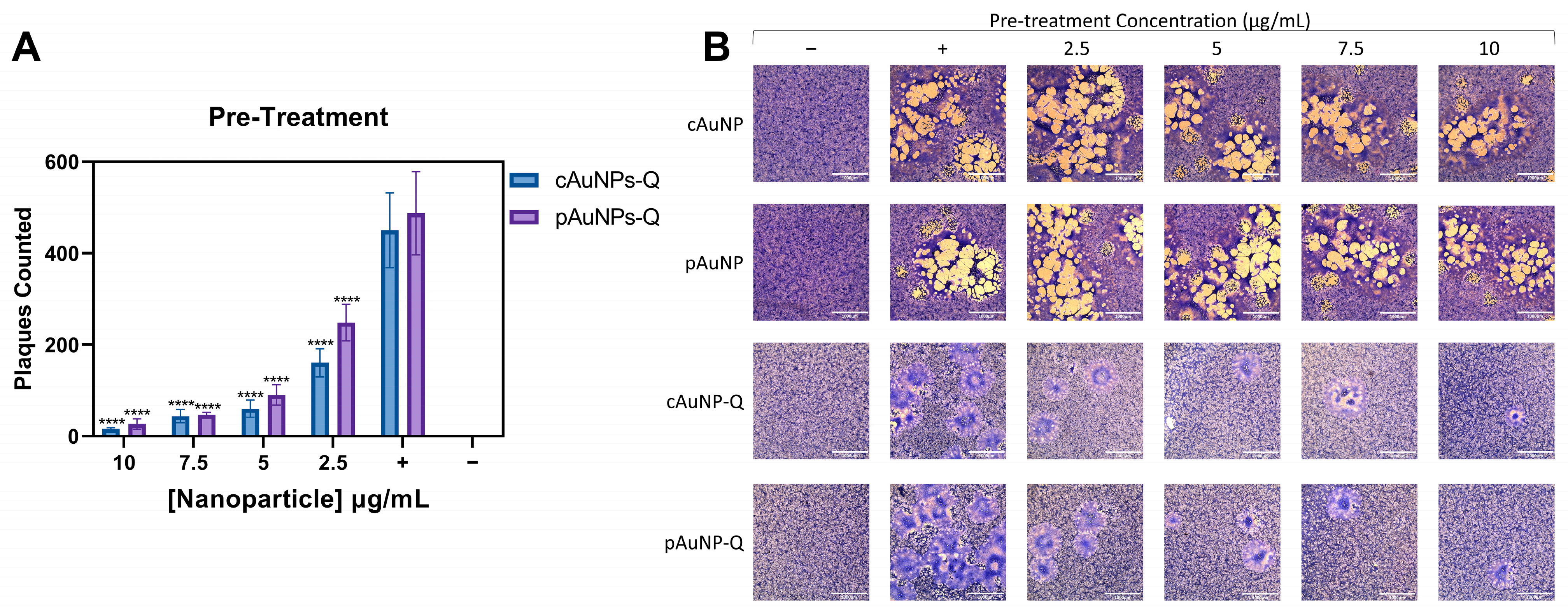
2. Results
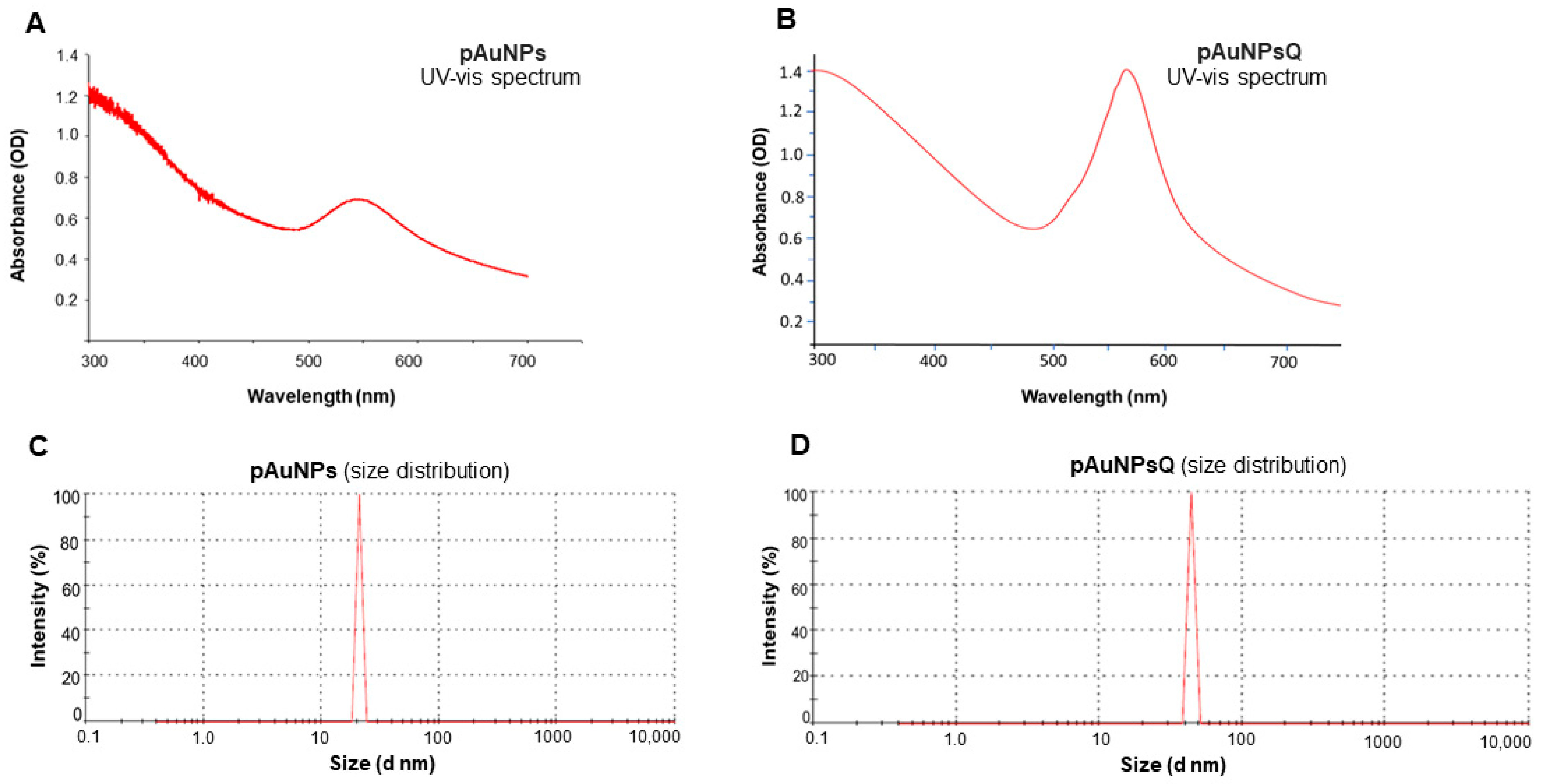
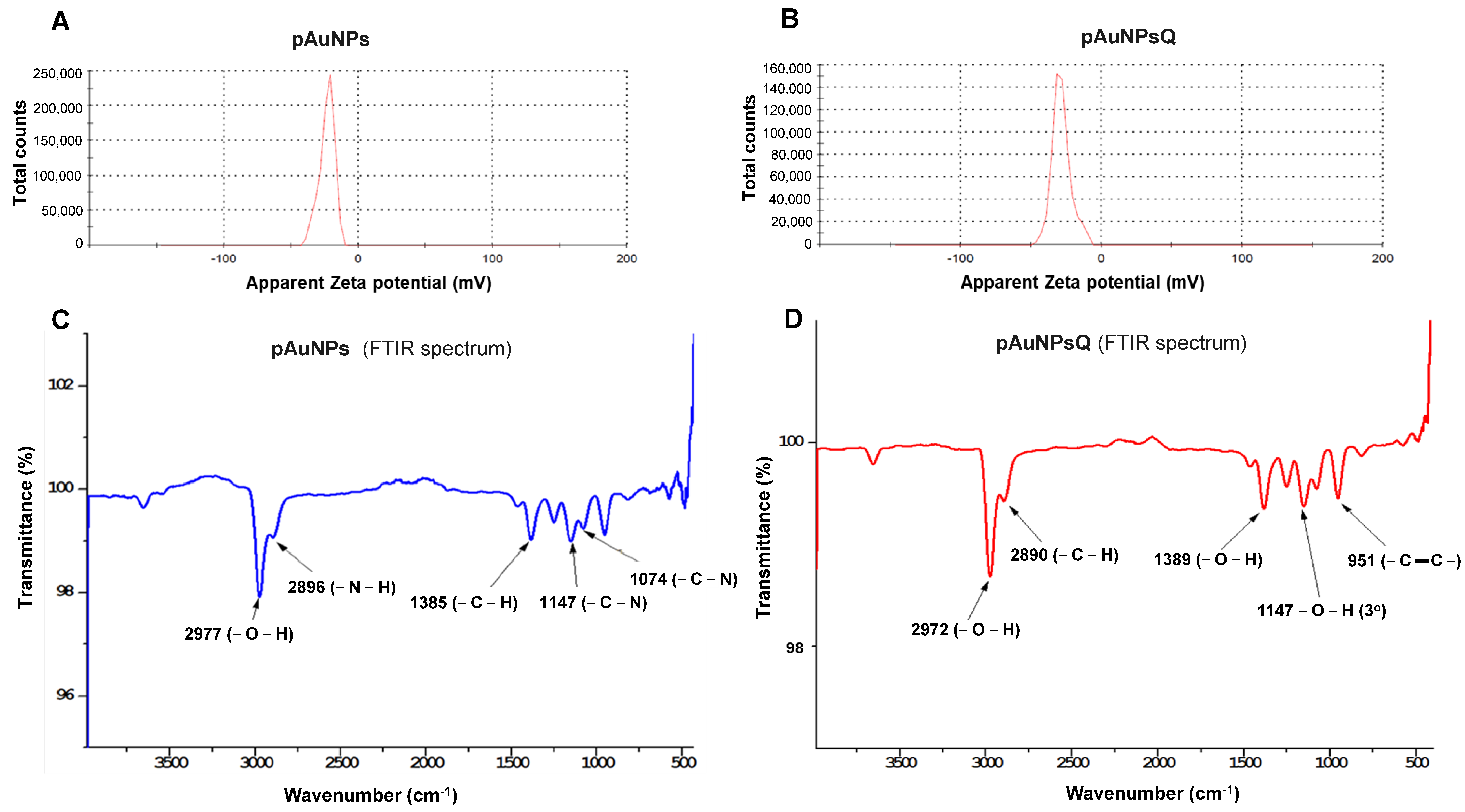
2.1. Size & Zeta (ζ) Potential
2.2. Fourier Transform Infra-Red Spectroscopy (FTRI)
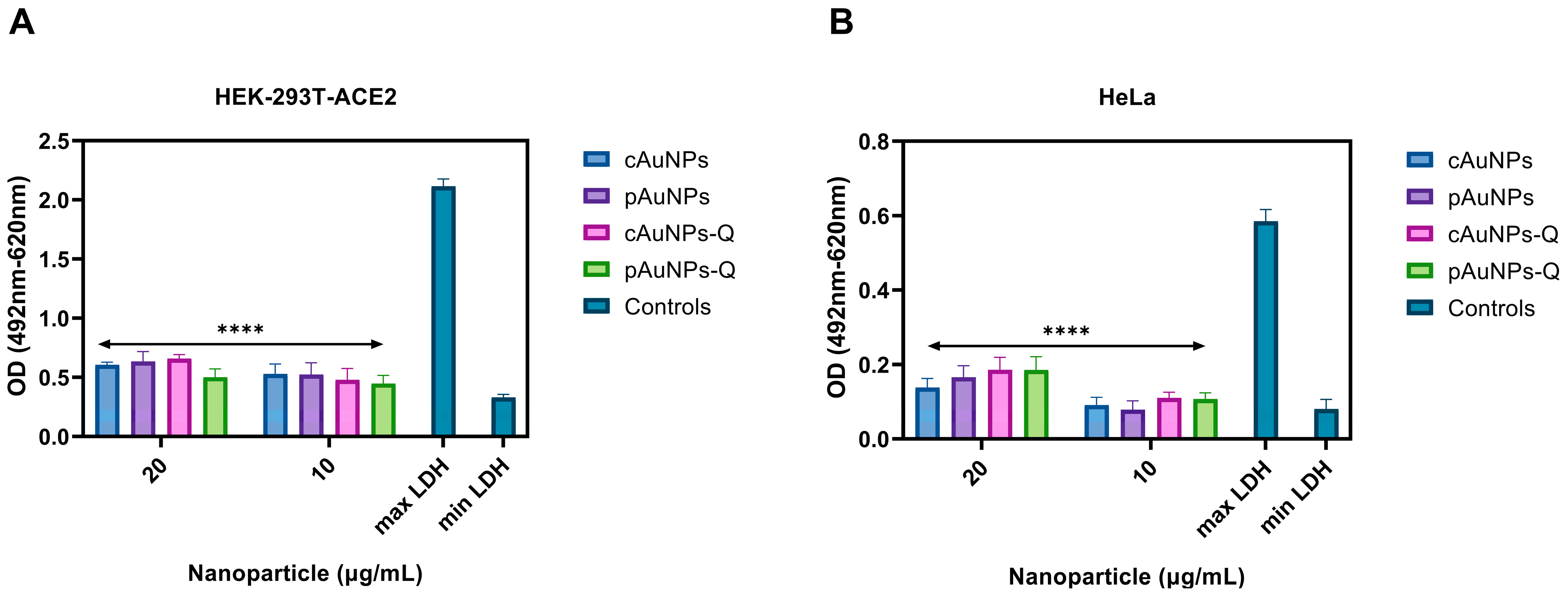
2.3. Quantification of Cytotoxicity Associated with the Nanoparticles Using LDH Assay
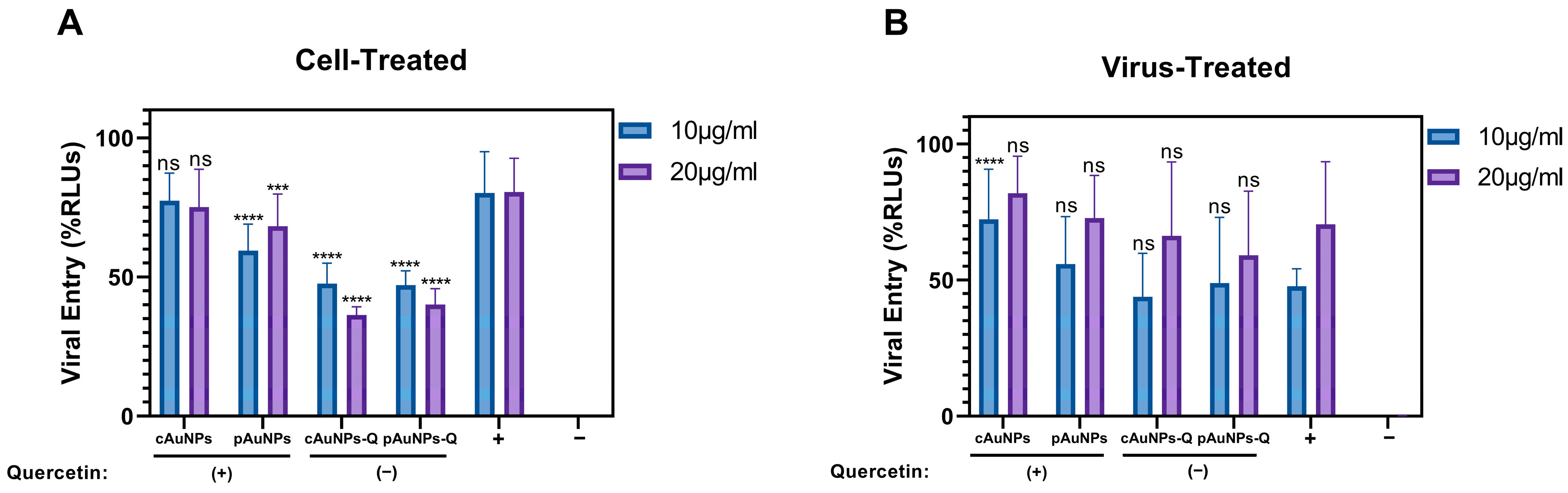
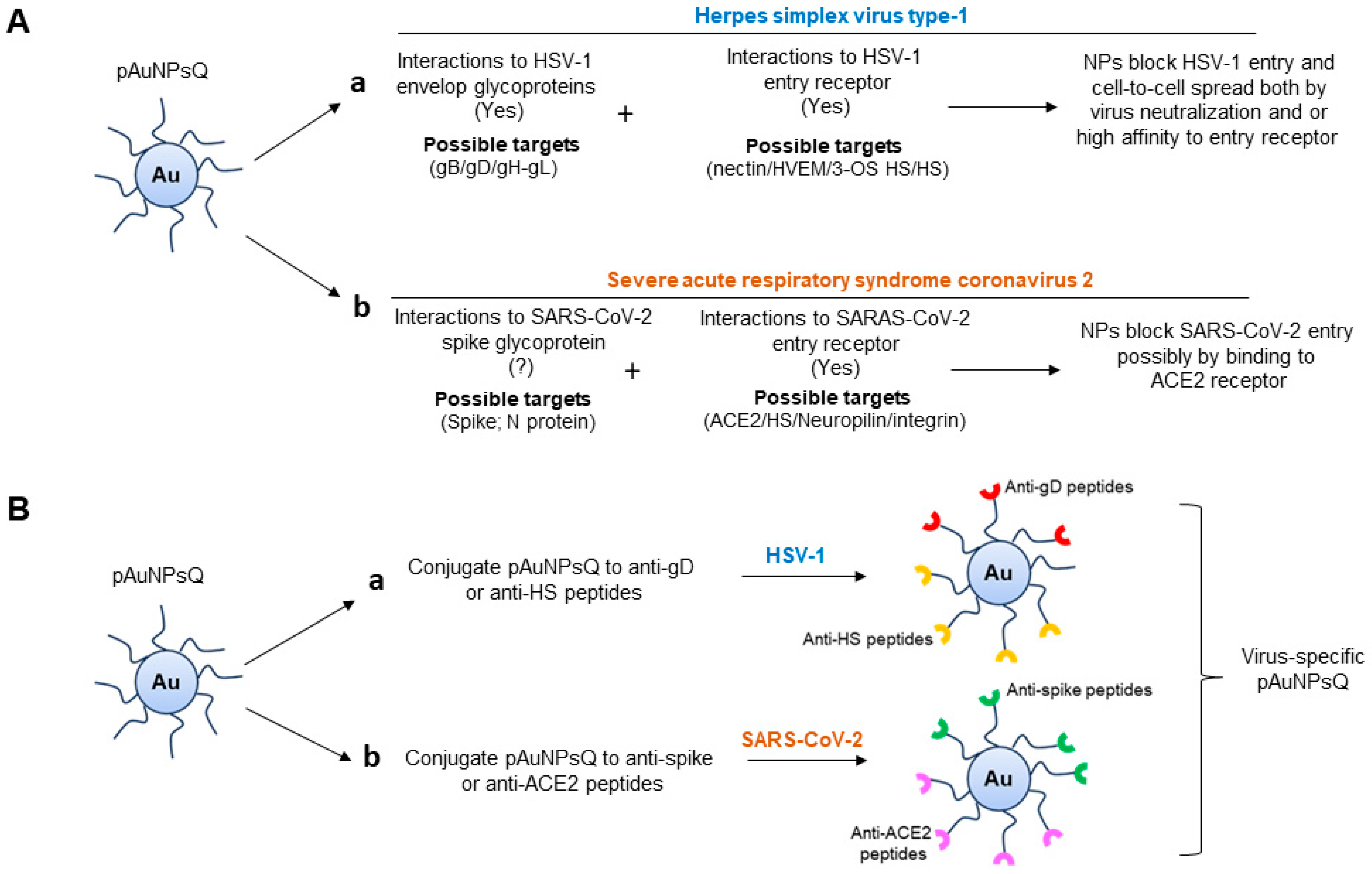
3. Discussion
4. Material and Methods
4.1. Characterization of Gold Nanoparticles
4.2. Cells and Viruses
4.3. LDH Cytotoxicity Assay
4.4. SARS-CoV-2 Spike Protein-Pseudotyped Lentivirus Entry Assay
4.5. Herpes Simplex Virus Typ-1 (HSV-1 gL86) Viral Entry Assay
4.6. Plaque Reduction Assay
4.7. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nie, C.; Stadtmüller, M.; Parshad, B.; Wallert, M.; Ahmadi, V.; Kerkhoff, Y.; Bhatia, S.; Block, S.; Cheng, C.; Wolff, T.; et al. Heteromultivalent topology-matched nanostructures as potent and broad-spectrum influenza A virus inhibitors. Sci. Adv. 2021, 7, eabd3803. [Google Scholar] [CrossRef] [PubMed]
- Krzyzowska, M.; Chodkowski, M.; Janicka, M.; Dmowska, D.; Tomaszewska, E.; Ranoszek-Soliwoda, K.; Bednarczyk, K.; Celichowski, G.; Grobelny, J. Lactoferrin-Functionalized Noble Metal Nanoparticles as New Antivirals for HSV-2 Infection. Microorganisms 2022, 10, 110. [Google Scholar] [CrossRef] [PubMed]
- Mishra, Y.K.; Adelung, R.; Röhl, C.; Shukla, D.; Spors, F.; Tiwari, V. Virostatic potential of micro-nano filopodia-like ZnO structures against herpes simplex virus-1. Antivir. Res. 2011, 92, 305–312. [Google Scholar] [CrossRef]
- Li, Y.; Xu, S.; Ye, Q.; Chi, H.; Guo, Z.; Chen, J.; Wu, M.; Fan, B.; Li, B.; Qin, C.F.; et al. Rational Development of Hypervalent Glycan Shield-Binding Nanoparticles with Broad-Spectrum Inhibition against Fatal Viruses Including SARS-CoV-2 Variants. Adv. Sci. 2023, 10, e2202689. [Google Scholar] [CrossRef]
- Morzy, D.; Bastings, M. Significance of Receptor Mobility in Multivalent Binding on Lipid Membranes. Angew. Chem. Int. Ed. Engl. 2022, 61, e202114167. [Google Scholar] [CrossRef]
- Arii, J.; Kawaguchi, Y. The Role of HSV Glycoproteins in Mediating Cell Entry. Adv. Exp. Med. Biol. 2018, 1045, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Deng, W.; Huang, B.; Gao, H.; Liu, J.; Ren, L.; Wei, Q.; Yu, P.; Xu, Y.; Qi, F.; et al. The pathogenicity of SARS-CoV-2 in hACE2 transgenic mice. Nature 2020, 583, 830–833. [Google Scholar] [CrossRef]
- Clausen, T.M.; Sandoval, D.R.; Spliid, C.B.; Pihl, J.; Perrett, H.R.; Painter, C.D.; Narayanan, A.; Majowicz, S.A.; Kwong, E.M.; McVicar, R.N.; et al. SARS-CoV-2 Infection Depends on Cellular Heparan Sulfate and ACE2. Cell 2020, 183, 1043–1057. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.Y.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef]
- Liu, J.; Lu, F.; Chen, Y.; Plow, E.; Qin, J. Integrin mediates cell entry of the SARS-CoV-2 virus independent of cellular receptor ACE2. J. Biol. Chem. 2022, 298, 101710. [Google Scholar] [CrossRef] [PubMed]
- Geraghty, R.J.; Krummenacher, C.; Cohen, G.H.; Eisenberg, R.J.; Spear, P.G. Entry of alphaherpesviruses mediated by poliovirus receptor-related protein 1 and poliovirus receptor. Science 1998, 280, 1618–1620. [Google Scholar] [CrossRef] [PubMed]
- Reske, A.; Pollara, G.; Krummenacher, C.; Chain, B.M.; Katz, D.R. Understanding HSV-1 entry glycoproteins. Rev. Med. Virol. 2007, 17, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Heldwein, E.E.; Krummenacher, C. Entry of herpesviruses into mammalian cells. Cell. Mol. Life Sci. 2008, 65, 1653–1668. [Google Scholar] [CrossRef]
- Su, S.; Xu, W.; Jiang, S. Virus Entry Inhibitors: Past, Present, and Future. Adv. Exp. Med. Biol. 2022, 1366, 1–13. [Google Scholar] [CrossRef]
- Travis, B.J.; Elste, J.; Gao, F.; Joo, B.Y.; Cuevas-Nunez, M.; Kohlmeir, E.; Tiwari, V.; Mitchell, J.C. Significance of chlorine-dioxide-based oral rinses in preventing SARS-CoV-2 cell entry. Oral. Dis. 2022, 28 (Suppl. 2), 2481–2491. [Google Scholar] [CrossRef]
- Lewis, M.A.O. Herpes simplex virus: An occupational hazard in dentistry. Int. Dent. J. 2004, 54, 103–111. [Google Scholar] [CrossRef]
- El Hayderi, L.; Delvenne, P.; Rompen, E.; Senterre, J.M.; Nikkels, A.F. Herpes simplex virus reactivation and dental procedures. Clin. Oral. Investig. 2013, 17, 1961–1964. [Google Scholar] [CrossRef]
- Bhaskaran, S.; Sharma, N.; Tiwari, P.; Singh, S.R.; Sahi, S.V. Fabrication of innocuous gold nanoparticles using plant cells in culture. Sci. Rep. 2019, 9, 12040. [Google Scholar] [CrossRef]
- Liskova, A.; Samec, M.; Koklesova, L.; Samuel, S.M.; Zhai, K.; Al-Ishaq, R.K.; Abotaleb, M.; Nosal, V.; Kajo, K.; Ashrafizadeh, M.; et al. Flavonoids against the SARS-CoV-2 induced inflammatory storm. Biomed. Pharmacother. 2021, 138, 111430. [Google Scholar] [CrossRef]
- León-Gutiérrez, G.; Elste, J.; Cabello-Gutiérrez, C.; Millán-Pacheco, C.; Martínez-Gómez, M.H.; Azuara, P.; Shalkow, J.; Mejía-Alvarez, R.; Tiwari, V.; Mejia, A. A master mix of flavonoids adsorbed onto functionalized TiO2 nanoparticles shows a potent antiviral activity against SARS-CoV-2. Appl. Microbiol. Biotechnol. 2022, 106, 5987–6002. [Google Scholar] [CrossRef] [PubMed]
- Badshah, S.L.; Faisal, S.; Muhammad, A.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Antiviral activities of flavonoids. Biomed. Pharmacother. 2021, 140, 111596. [Google Scholar] [CrossRef] [PubMed]
- Volgenant, C.M.C.; Persoon, I.F.; de Ruijter, R.A.G.; de Soet, J.J. Infection control in dental health care during and after the SARS-CoV-2 outbreak. Oral. Dis. 2021, 27, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Lo Giudice, R. The severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) in dentistry. management of biological risk in dental practice. Int. J. Environ. Res. Public Health 2020, 17, 3067. [Google Scholar] [CrossRef]
- Anjum, A.; Hosein, M.; Butt, S.A.; Fawad, B.; Abidi, F. Efficacy of Two Mouth Rinses in Reducing Aerosol Bacterial Load during Ultrasonic Scaling. J. Adv. Med. Med. Res. 2019, 31, 1–9. [Google Scholar] [CrossRef]
- Bidra, A.S.; Pelletier, J.S.; Westover, J.B.; Frank, S.; Brown, S.M.; Tessema, B. Comparison of In Vitro Inactivation of SARS-CoV-2 with Hydrogen Peroxide and Povidone-Iodine Oral Antiseptic Rinses. J. Prosthodont. 2020, 29, 599–603. [Google Scholar] [CrossRef]
- Koch-Heier, J.; Hoffmann, H.; Schindler, M.; Lussi, A.; Planz, O. Inactivation of SARS-CoV-2 through treatment with the mouth rinsing solutions viruprox® and bacterx® pro. Microorganisms 2021, 9, 521. [Google Scholar] [CrossRef]
- Paull, J.R.; Heery, G.P.; Bobardt, M.D.; Castellarnau, A.; Luscombe, C.A.; Fairley, J.K.; Gallay, P.A. Virucidal and antiviral activity of astodrimer sodium against SARS-CoV-2 in vitro. Antivir. Res. 2021, 191, 105089. [Google Scholar] [CrossRef]
- Takeda, Y.; Jamsransuren, D.; Matsuda, S.; Crea, R.; Ogawa, H. The SARS-CoV-2-inactivating activity of hydroxytyrosol-rich aqueous olive pulp extract (Hidrox®) and its use as a virucidal cream for topical application. Viruses 2021, 13, 232. [Google Scholar] [CrossRef]
- Jamil, N.; Baqar, M.; Ilyas, S.; Qadir, A.; Arslan, M.; Salman, M.; Ahsan, N.; Zahid, H. Use of Mercury in Dental Silver Amalgam: An Occupational and Environmental Assessment. Biomed. Res. Int. 2016, 2016, 6126385. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the Lactate Dehydrogenase Assay. Cold Spring Harb. Protoc. 2018, 2018, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Arduino, P.G.; Cabras, M.; Lodi, G.; Petti, S. Herpes simplex virus type 1 in subgingival plaque and periodontal diseases. Meta-analysis of observational studies. J. Periodontal. Res. 2022, 57, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Sadalage, P.S.; Patil, R.V.; Havaldar, D.V.; Gavade, S.S.; Santos, A.C.; Pawar, K.D. Optimally biosynthesized, PEGylated gold nanoparticles functionalized with quercetin and camptothecin enhance potential anti-inflammatory, anti-cancer and anti-angiogenic activities. J. Nanobiotechnol. 2021, 19, 84. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, M.; Portney, N.G.; Cui, D.; Budak, G.; Ozbay, E.; Ozkan, M.; Ozkan, C.S. Zeta potential: A surface electrical characteristic to probe the interaction of nanoparticles with normal and cancer human breast epithelial cells. Biomed. Microdevices 2008, 10, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, M.R.; Bednar, H.R.; Haes, A.J. Investigations of the mechanism of gold nanoparticle stability and surface functionalization in capillary electrophoresis. ACS. Nano 2009, 3, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Kolanthai, E.; Neal, C.J.; Kumar, U.; Fu, Y.; Seal, S. Antiviral nanopharmaceuticals: Engineered surface interactions and virus-selective activity. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2022, 14, e1823. [Google Scholar] [CrossRef] [PubMed]
- Delshadi, R.; Bahrami, A.; McClements, D.J.; Moore, M.D.; Williams, L. Development of nanoparticle-delivery systems for antiviral agents: A review. J. Control. Release 2021, 331, 30–44. [Google Scholar] [CrossRef]
- Wiesenthal, A.; Hunter, L.; Wang, S.; Wickliffe, J.; Wilkerson, M. Nanoparticles: Small and mighty. Int. J. Dermatol. 2011, 50, 247–254. [Google Scholar] [CrossRef]
- Sanna, V.; Youssef, M.F.; Pala, N.; Rogolino, D.; Carcelli, M.; Singh, P.K.; Sanchez, T.; Neamati, N.; Sechi, M. Inhibition of Human Immunodeficiency Virus-1 Integrase by β-Diketo Acid Coated Gold Nanoparticles. ACS Med. Chem. Lett. 2020, 11, 857–861. [Google Scholar] [CrossRef]
- Bawage, S.S.; Tiwari, P.M.; Singh, A.; Dixit, S.; Pillai, S.R.; Dennis, V.A.; Singh, S.R. Gold nanorods inhibit respiratory syncytial virus by stimulating the innate immune response. Nanomedicine 2016, 12, 2299–2310. [Google Scholar] [CrossRef]
- Mori, Y.; Ono, T.; Miyahira, Y.; Nguyen, V.Q.; Matsui, T.; Ishihara, M. Antiviral activity of silver nanoparticle/chitosan composites against H1N1 influenza A virus. Nanoscale Res. Lett. 2013, 8, 93. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewska, E.; Ranoszek-Soliwoda, K.; Bednarczyk, K.; Lech, A.; Janicka, M.; Chodkowski, M.; Psarski, M.; Celichowski, G.; Krzyzowska, M.; Grobelny, J. Anti-HSV Activity of Metallic Nanoparticles Functionalized with Sulfonates vs. Polyphenols. Int. J. Mol. Sci. 2022, 23, 13104. [Google Scholar] [CrossRef] [PubMed]
- Baram-Pinto, D.; Shukla, S.; Gedanken, A.; Sarid, R. Inhibition of HSV-1 attachment, entry, and cell-to-cell spread by functionalized multivalent gold nanoparticles. Small 2010, 6, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Beer, J.C.; Majmudar, H.; Mishra, Y.K.; Shukla, D.; Tiwari, V. Book Chapter—“Nanomaterial Biointeractions at the Cellular, Organismal and Systemic Levels”—Springer Nature (Published). 2021. Available online: https://www.researchgate.net/publication/349120588_Nanomaterial_Biointeractions_at_the_Cellular_Organismal_and_System_Levels (accessed on 27 September 2023).
- Dan, M.; Wen, H.; Shao, A.; Xu, L. Silver Nanoparticle Exposure Induces Neurotoxicity in the Rat Hippocampus Without Increasing the Blood-Brain Barrier Permeability. J. Biomed. Nanotechnol. 2018, 14, 1330–1338. [Google Scholar] [CrossRef]
- Pan, B.; Fang, S.; Zhang, J.; Pan, Y.; Liu, H.; Wang, Y.; Li, M.; Liu, L. Chinese herbal compounds against SARS-CoV-2: Puerarin and quercetin impair the binding of viral S-protein to ACE2 receptor. Comput. Struct. Biotechnol. J. 2020, 18, 3518–3527. [Google Scholar] [CrossRef]
- Agelidis, A.; Koujah, L.; Suryawanshi, R.; Yadavalli, T.; Mishra, Y.K.; Adelung, R.; Shukla, D. An Intra-Vaginal Zinc Oxide Tetrapod Nanoparticles (ZOTEN) and Genital Herpesvirus Cocktail Can Provide a Novel Platform for Live Virus Vaccine. Front. Immunol. 2019, 10, 500. [Google Scholar] [CrossRef]
- Wu, W.; Wang, W.; Liang, L.; Chen, J.; Wei, B.; Huang, X.R.; Wang, X.; Yu, X.; Lan, H.Y. Treatment with quercetin inhibits SARS-CoV-2 N protein-induced acute kidney injury by blocking Smad3-dependent G1 cell-cycle arrest. Mol. Ther. 2023, 31, 344–361. [Google Scholar] [CrossRef]
- Bibi, S.; Sarfraz, A.; Mustafa, G.; Ahmad, Z.; Zeb, M.A.; Wang, Y.B.; Khan, T.; Khan, M.S.; Kamal, M.A.; Yu, H. Impact of Traditional Plants and their Secondary Metabolites in the Discovery of COVID-19 Treatment. Curr. Pharm. Des. 2021, 27, 1123–1143. [Google Scholar] [CrossRef]
- Karthik, C.; Punnaivalavan, K.A.; Prabha, S.P.; Caroline, D.G. Multifarious global flora fabricated phytosynthesis of silver nanoparticles: A green nanoweapon for antiviral approach including SARS-CoV-2. Int. Nano Lett. 2022, 12, 313–344. [Google Scholar] [CrossRef]
- Punetha, V.D.; Dhali, S.; Rana, A.; Karki, N.; Tiwari, H.; Negi, P.; Basak, S.; Sahoo, N.G. Recent Advancements in Green Synthesis of Nanoparticles for Improvement of Bioactivities: A Review. Curr. Pharm. Biotechnol. 2022, 23, 904–919. [Google Scholar] [CrossRef]
- Fazal, H.; Abbasi, B.H.; Ahmad, N.; Ali, M. Elicitation of Medicinally Important Antioxidant Secondary Metabolites with Silver and Gold Nanoparticles in Callus Cultures of Prunella vulgaris L. Appl. Biochem. Biotechnol. 2016, 180, 1076–1092. [Google Scholar] [CrossRef]
- Tiwari, V.; Liu, J.; Valyi-Nagy, T.; Shukla, D. Anti-heparan sulfate peptides that block herpes simplex virus infection in vivo. J. Biol. Chem. 2011, 286, 25406–25415. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Karasneh, G.A.; Jarding, M.J.; Tiwari, V.; Shukla, D. A 3-O-sulfated heparan sulfate binding peptide preferentially targets herpes simplex virus 2-infected cells. J. Virol. 2012, 86, 6434–6443. [Google Scholar] [CrossRef] [PubMed]
- Antoine, T.E.; Yakoub, A.; Maus, E.; Shukla, D.; Tiwari, V. Zebrafish 3-O-sulfotransferase-4 generated heparan sulfate mediates HSV-1 entry and spread. PLoS ONE. 2014, 9, e87302. [Google Scholar] [CrossRef] [PubMed]
- D’Ercole, S.; Cellini, L.; Pilato, S.; Di Lodovico, S.; Iezzi, G.; Piattelli, A.; Petrini, M. Material characterization and Streptococcus oralis adhesion on Polyetheretherketone (PEEK) and titanium surfaces used in implantology. J. Mater. Sci. Mater. Med. 2020, 31, 84. [Google Scholar] [CrossRef] [PubMed]
- Majmudar, H.; Hao, M.; Sankaranarayanan, N.V.; Zanotti, B.; Volin, M.V.; Desai, U.R.; Tiwari, V. A synthetic glycosaminoglycan mimetic blocks HSV-1 infection in human iris stromal cells. Antivir. Res. 2019, 161, 154–162. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elste, J.; Kumari, S.; Sharma, N.; Razo, E.P.; Azhar, E.; Gao, F.; Nunez, M.C.; Anwar, W.; Mitchell, J.C.; Tiwari, V.; et al. Plant Cell-Engineered Gold Nanoparticles Conjugated to Quercetin Inhibit SARS-CoV-2 and HSV-1 Entry. Int. J. Mol. Sci. 2023, 24, 14792. https://doi.org/10.3390/ijms241914792
Elste J, Kumari S, Sharma N, Razo EP, Azhar E, Gao F, Nunez MC, Anwar W, Mitchell JC, Tiwari V, et al. Plant Cell-Engineered Gold Nanoparticles Conjugated to Quercetin Inhibit SARS-CoV-2 and HSV-1 Entry. International Journal of Molecular Sciences. 2023; 24(19):14792. https://doi.org/10.3390/ijms241914792
Chicago/Turabian StyleElste, James, Sangeeta Kumari, Nilesh Sharma, Erendira Palomino Razo, Eisa Azhar, Feng Gao, Maria Cuevas Nunez, Wasim Anwar, John C. Mitchell, Vaibhav Tiwari, and et al. 2023. "Plant Cell-Engineered Gold Nanoparticles Conjugated to Quercetin Inhibit SARS-CoV-2 and HSV-1 Entry" International Journal of Molecular Sciences 24, no. 19: 14792. https://doi.org/10.3390/ijms241914792
APA StyleElste, J., Kumari, S., Sharma, N., Razo, E. P., Azhar, E., Gao, F., Nunez, M. C., Anwar, W., Mitchell, J. C., Tiwari, V., & Sahi, S. (2023). Plant Cell-Engineered Gold Nanoparticles Conjugated to Quercetin Inhibit SARS-CoV-2 and HSV-1 Entry. International Journal of Molecular Sciences, 24(19), 14792. https://doi.org/10.3390/ijms241914792