
Long-Term Effects of Suramin on Renal Function in Streptozotocin-Induced Diabetes in Rats
 , , , , and
, , , , and
Abstract
:1. Introduction
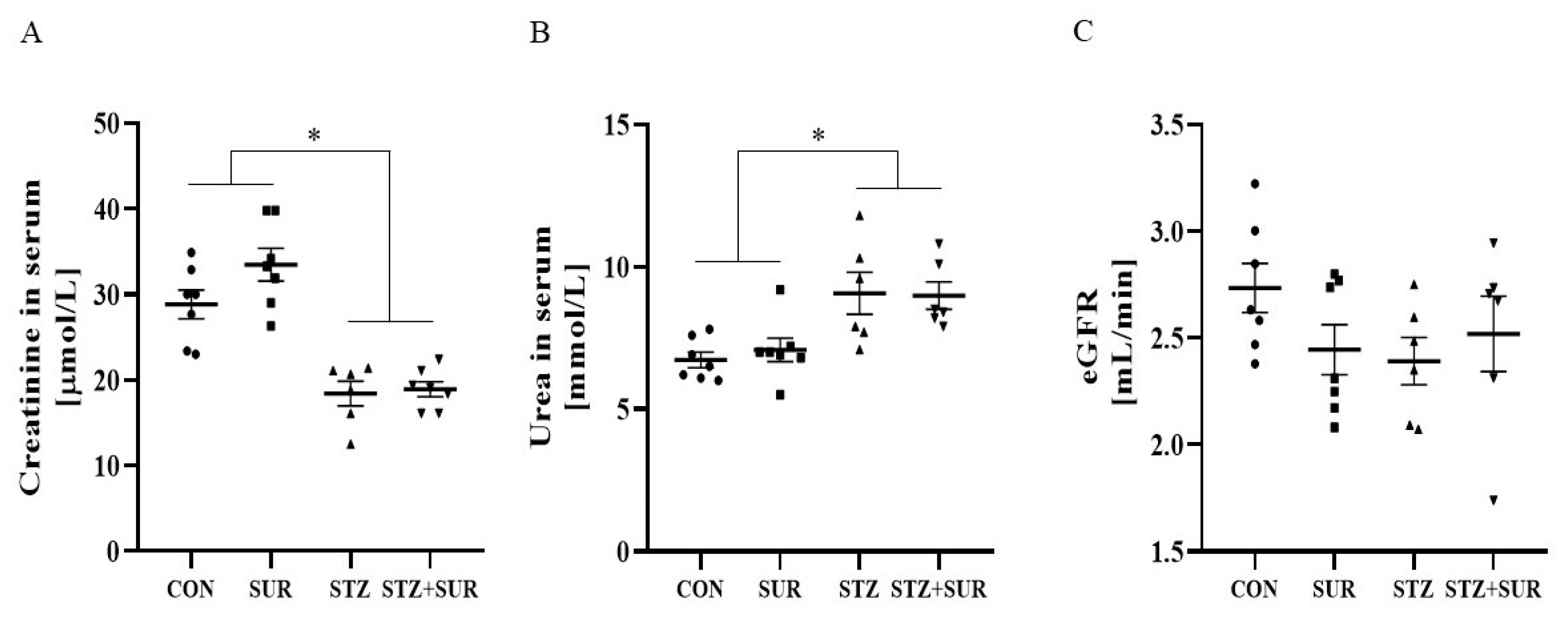
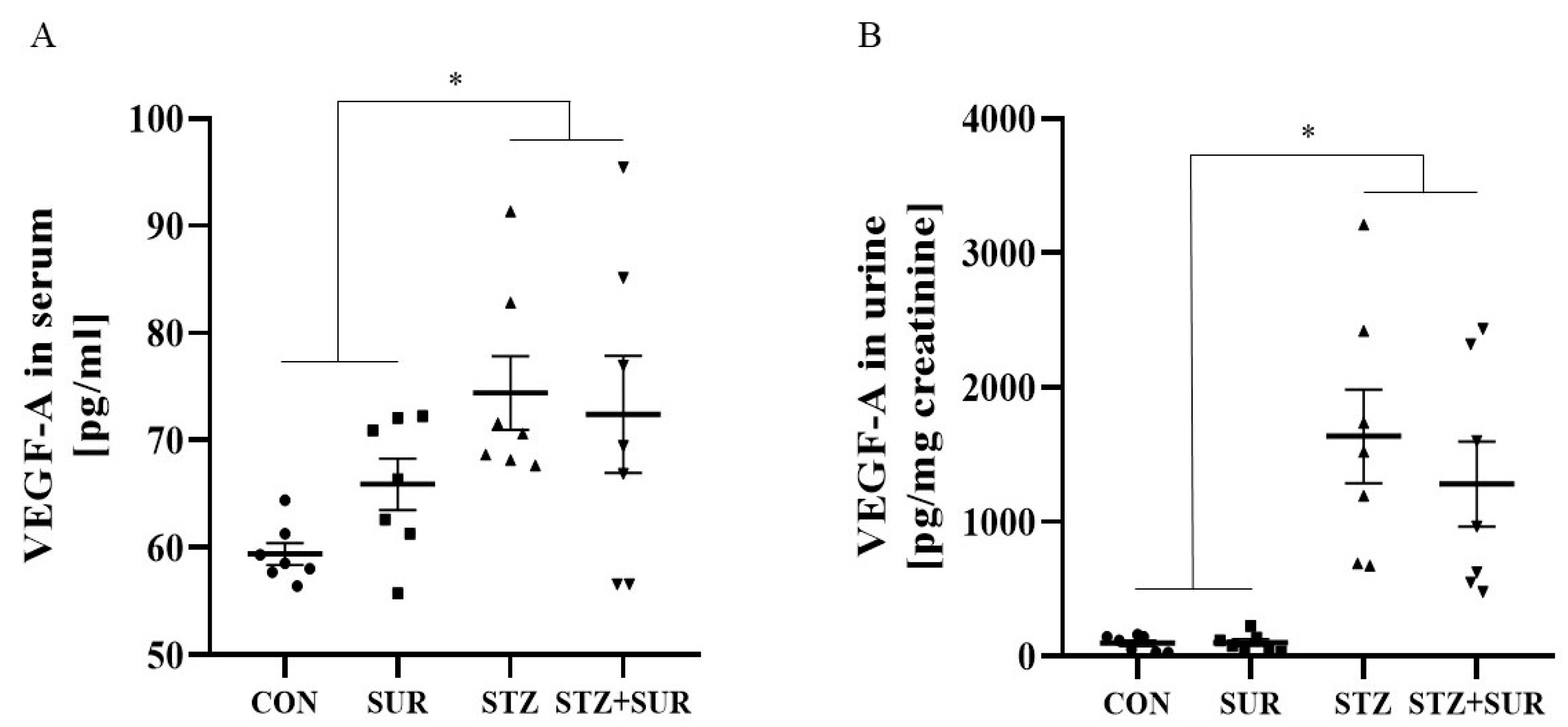
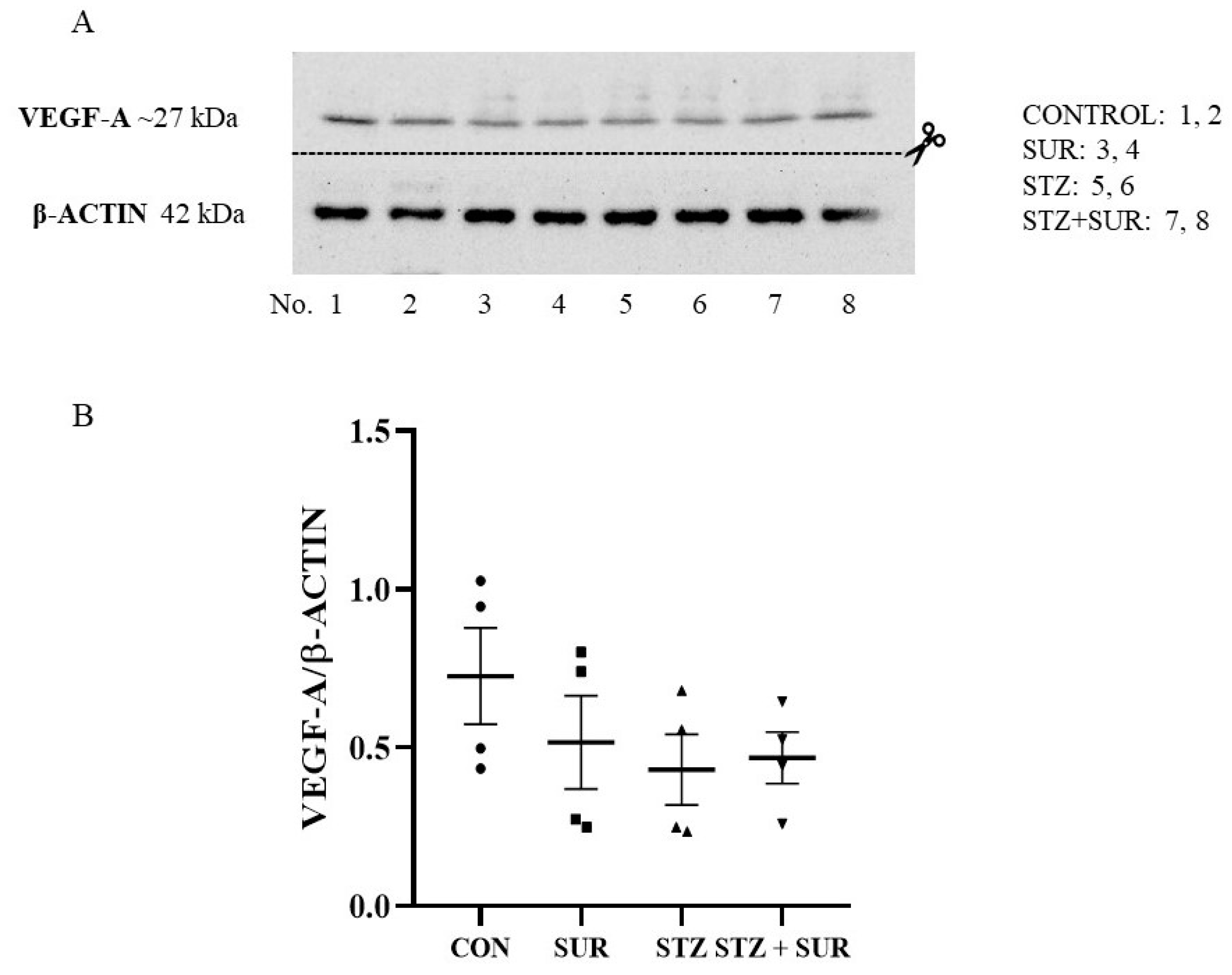
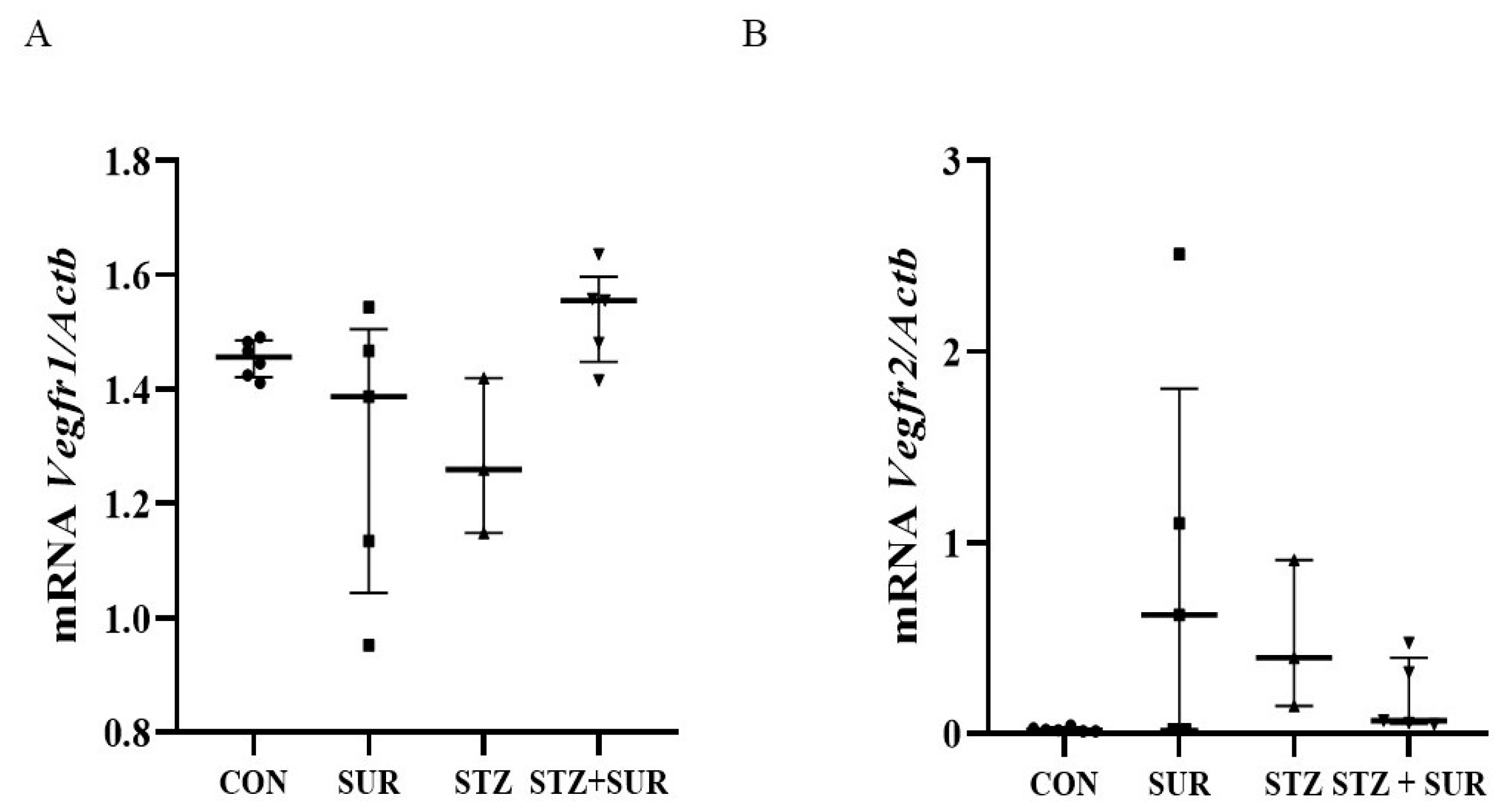
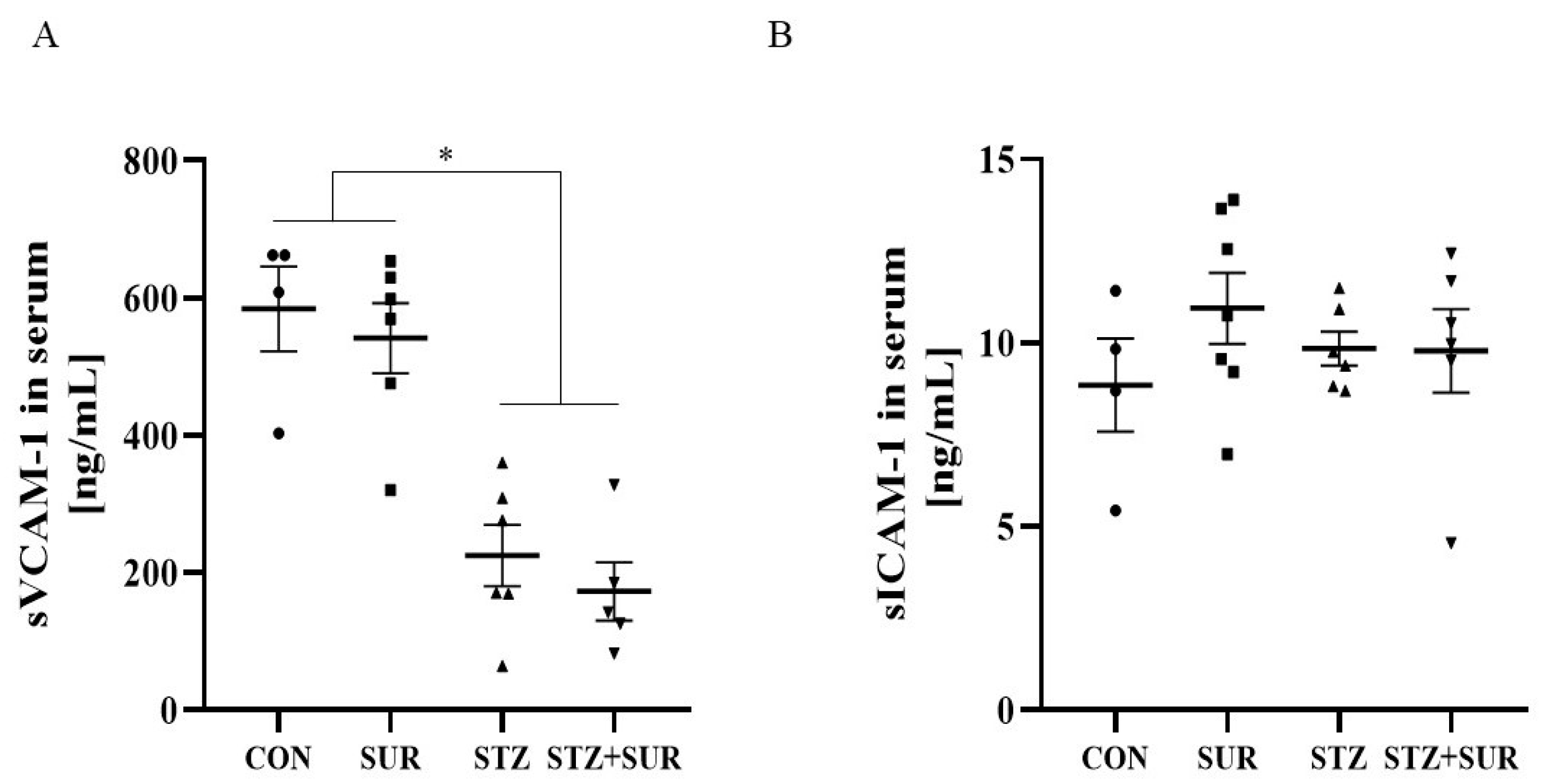
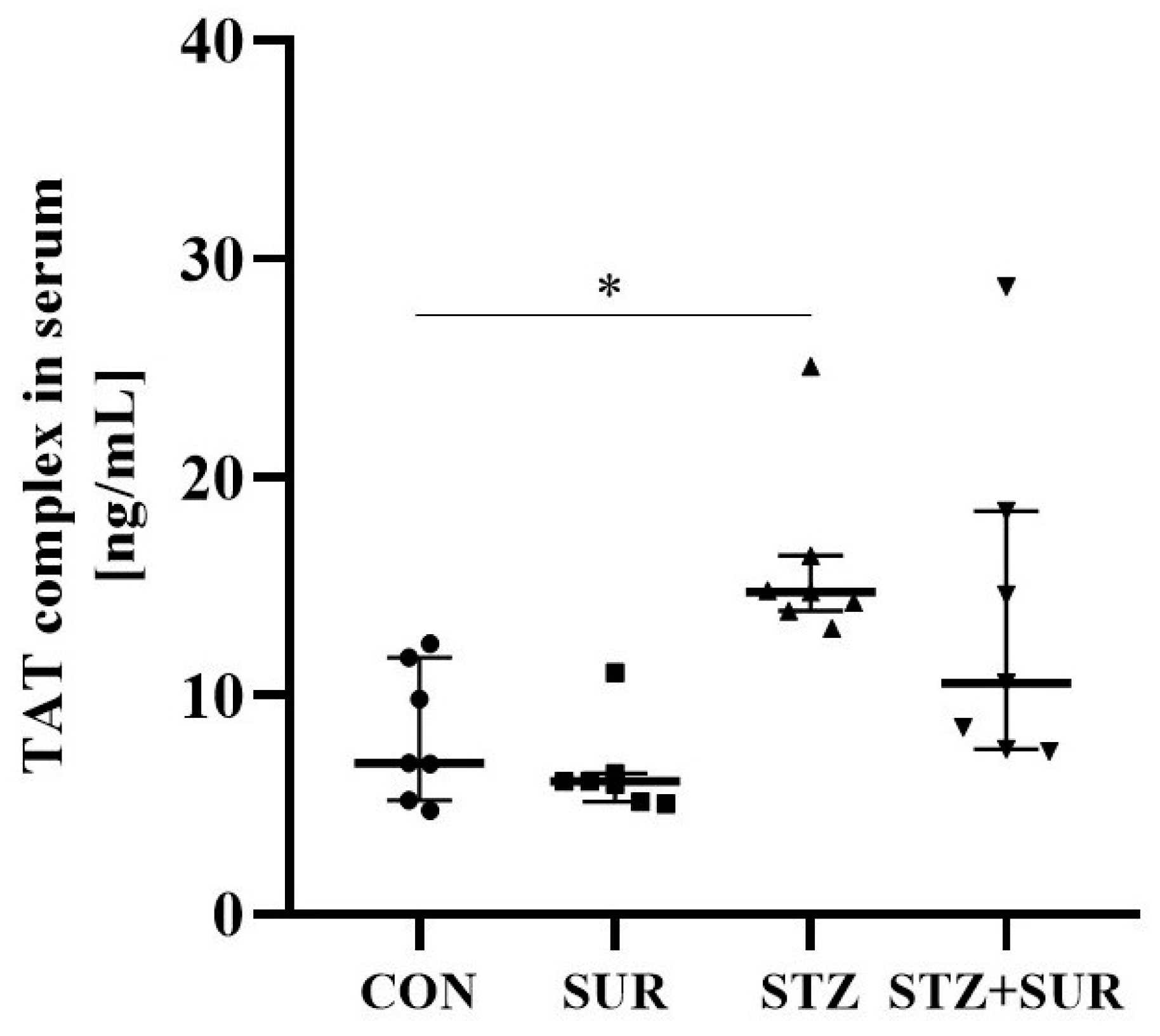
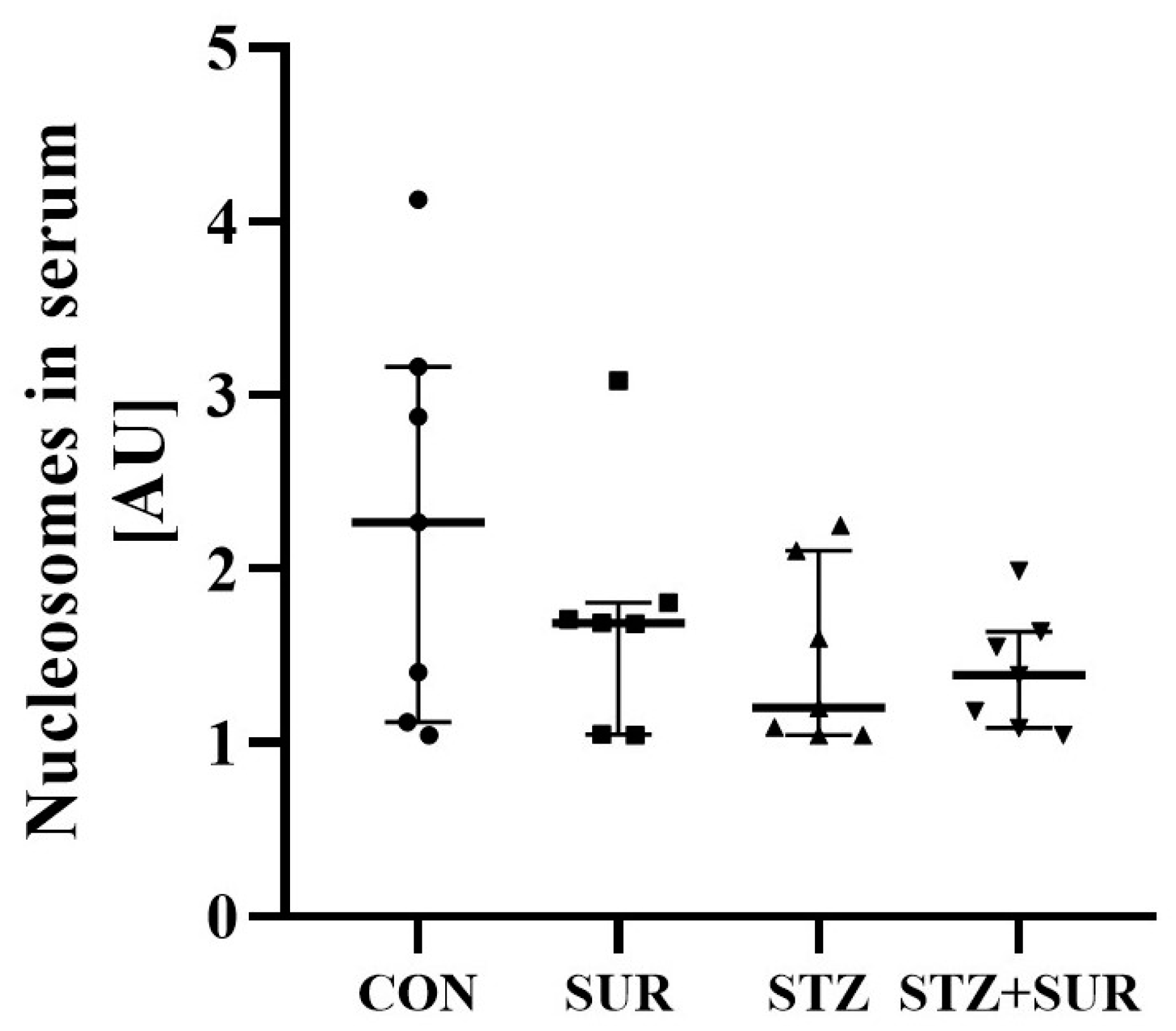
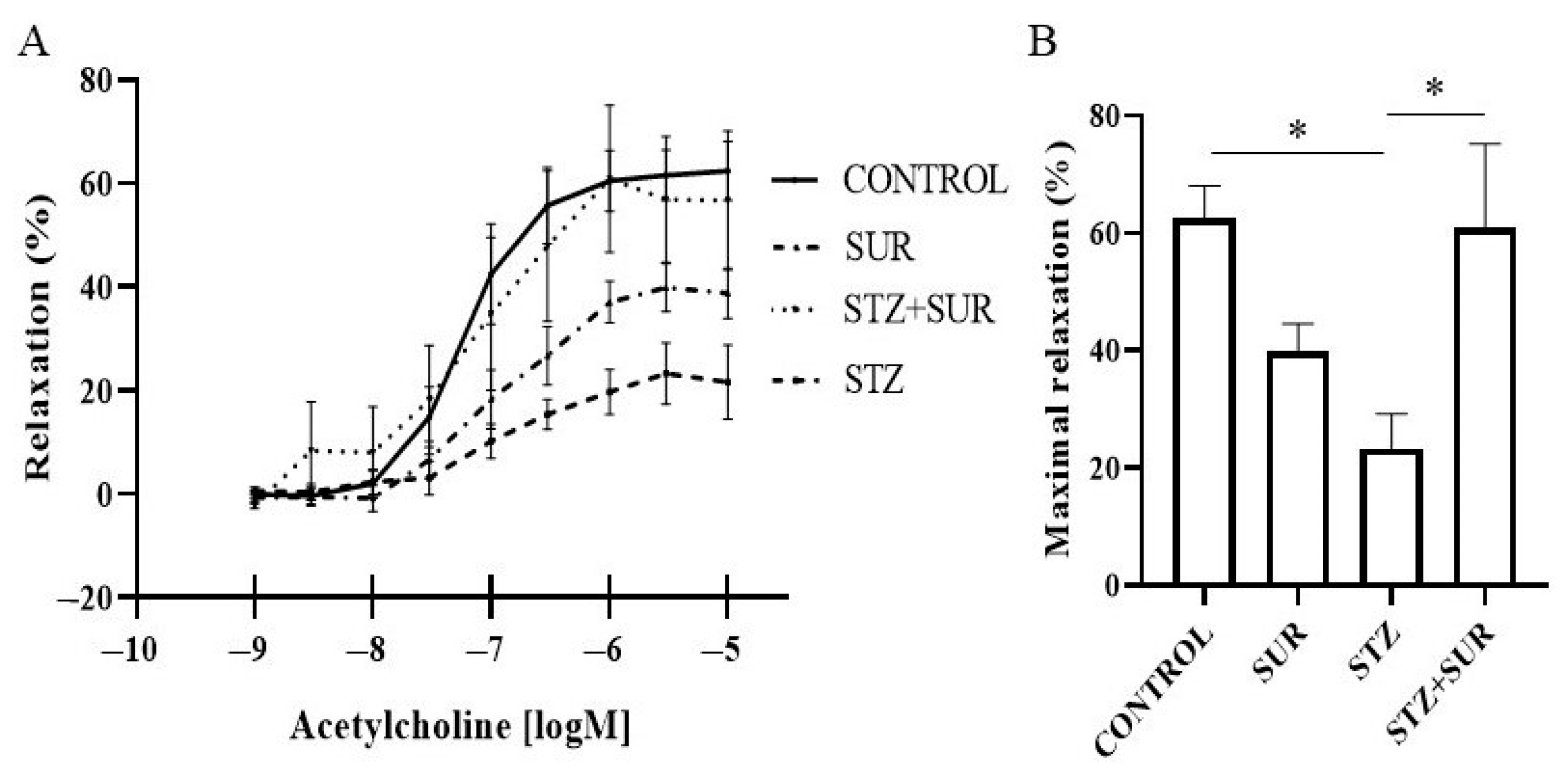
2. Results
2.1. Experiments In Vivo
2.2. Experiment In Vitro
2.3. Experiments Ex Vivo
3. Discussion
4. Materials and Methods
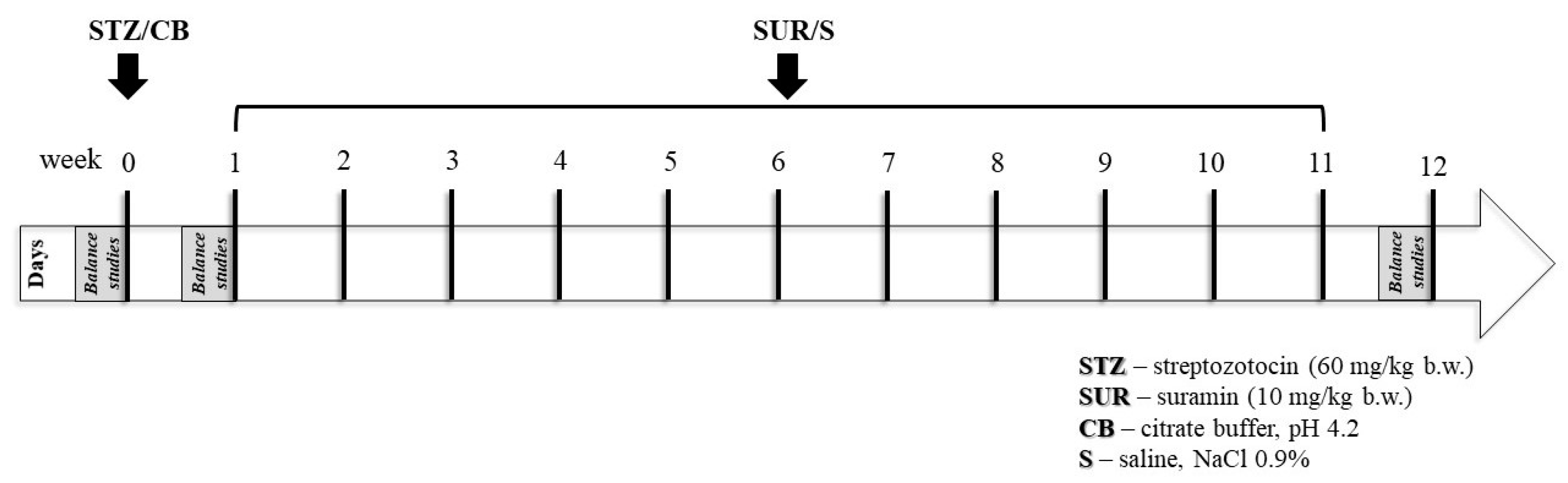
4.1. Animals
- Non-diabetic group (CON); citrate buffer was injected on the first day of the experiment (week 0), followed by weekly saline injections for 11 weeks.
- Suramin-treated non-diabetic group (SUR); suramin (10 mg/kg, i.p.) was injected weekly for 11 weeks, starting 1 week after citrate buffer administration.
- Diabetes group (STZ); streptozotocin was injected on the first day of the experiment (week 0), followed by weekly saline injections for 11 weeks.
- Suramin-treated diabetes group (STZ + SUR); suramin (10 mg/kg, i.p.) was injected weekly for 11 weeks, starting one week after streptozotocin administration.
4.2. Renal Interlobar Artery Preparation
4.3. Vascular Response Measurements
4.4. Relative Quantitative Real-Time RT-PCR Assay
4.5. Analysis by Western Blotting
4.6. Analytical Methods
4.7. eGFR Analysis
4.8. Nutrient Variable Analysis
4.9. Statistical Analysis
4.10. Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brun, R.; Blum, J.; Chappuis, F.; Burri, C. Human African Trypanosomiasis. Lancet 2010, 375, 148–159. [Google Scholar] [CrossRef]
- Naviaux, R.K.; Curtis, B.; Li, K.; Naviaux, J.C.; Bright, A.T.; Reiner, G.E.; Westerfield, M.; Goh, S.; Alaynick, W.A.; Wang, L.; et al. Low-Dose Suramin in Autism Spectrum Disorder: A Small, Phase I/II, Randomized Clinical Trial. Ann. Clin. Transl. Neurol. 2017, 4, 491–505. [Google Scholar] [CrossRef]
- Villalona-Calero, M.A.; Wientjes, M.G.; Otterson, G.A.; Kanter, S.; Young, D.; Murgo, A.J.; Fischer, B.; DeHoff, C.; Chen, D.; Yeh, T.-K.; et al. Phase I Study of Low-Dose Suramin as a Chemosensitizer in Patients with Advanced Non-Small Cell Lung Cancer. Clin. Cancer Res. 2003, 9, 3303–3311. [Google Scholar]
- Wiedemar, N.; Hauser, D.A.; Mäser, P. 100 Years of Suramin. Antimicrob. Agents Chemother. 2020, 64, 1100–1128. [Google Scholar] [CrossRef]
- Mitsuya, H.; Popovic, M.; Yarchoan, R.; Matsushita, S.; Gallo, R.C.; Broder, S. Suramin Protection of T Cells in Vitro against Infectivity and Cytopathic Effect of HTLV-III. Science 1984, 226, 174–177. [Google Scholar] [CrossRef] [PubMed]
- Villalba, N.; Sackheim, A.M.; Lawson, M.A.; Haines, L.; Chen, Y.-L.; Sonkusare, S.K.; Ma, Y.-T.; Li, J.; Majumdar, D.; Bouchard, B.A.; et al. The Polyanionic Drug Suramin Neutralizes Histones and 1 Prevents Endotheliopathy 2. J. Immunol. 2023, 211, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Waltenberger, J.; Mayr, U.; Frank, H.; Hombach, V. Suramin Is a Potent Inhibitor of Vascular Endothelial Growth Factor. A Contribution to the Molecular Basis of Its Antiangiogenic Action. J. Mol. Cell Cardiol. 1996, 28, 1523–1529. [Google Scholar] [CrossRef]
- Vempati, P.; Popel, A.S.; mac Gabhann, F. Extracellular Regulation of VEGF: Isoforms, Proteolysis, and Vascular Patterning. Cytokine Growth Factor. Rev. 2014, 25, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Chen, T.T.; Barber, C.L.; Jordan, M.C.; Murdock, J.; Desai, S.; Ferrara, N.; Nagy, A.; Roos, K.P.; Iruela-Arispe, M.L. Autocrine VEGF Signaling Is Required for Vascular Homeostasis. Cell 2007, 130, 691–703. [Google Scholar] [CrossRef]
- Kamba, T.; Tam, B.Y.Y.; Hashizume, H.; Haskell, A.; Sennino, B.; Mancuso, M.R.; Norberg, S.M.; O’Brien, S.M.; Davis, R.B.; Gowen, L.C.; et al. VEGF-Dependent Plasticity of Fenestrated Capillaries in the Normal Adult Microvasculature. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H560–H576. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Moon, S.O.; Kim, S.H.; Kim, H.J.; Koh, Y.S.; Koh, G.Y. Vascular Endothelial Growth Factor Expression of Intercellular Adhesion Molecule 1 (ICAM-1), Vascular Cell Adhesion Molecule 1 (VCAM-1), and E-Selectin through Nuclear Factor-ΚB Activation in Endothelial Cells. J. Biol. Chem. 2001, 276, 7614–7620. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yang, Q.; Ma, B.; Wang, X.; Bove, A.M.; Simone, G. Molecular Bases of VEGFR-2-Mediated Physiological Function and Pathological Role. Front. Cell Dev. Biol. 2020, 8, 599281. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The Biology of VEGF and Its Receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Bussolati, B.; Dunk, C.; Grohman, M.; Kontos, C.D.; Mason, J.; Ahmed, A. Vascular Endothelial Growth Factor Receptor-1 Modulates Vascular Endothelial Growth Factor-Mediated Angiogenesis via Nitric Oxide. Am. J. Pathol. 2001, 159, 993–1008. [Google Scholar] [CrossRef]
- Persson, F.; Rossing, P. Diagnosis of Diabetic Kidney Disease: State of the Art and Future Perspective. Kidney Int. Suppl. 2018, 8, 2–7. [Google Scholar] [CrossRef]
- Hempel, A.; Maasch, C.; Heintze, U.; Lindschau, C.; Dietz, R.; Luft, F.C.; Haller, H. High Glucose Concentrations Increase Endothelial Cell Permeability via Activation of Protein Kinase Cα. Circ. Res. 1997, 81, 363–371. [Google Scholar] [CrossRef]
- Varma, S.; Lal, B.K.; Zheng, R.; Breslin, J.W.; Saito, S.; Pappas, P.J.; Hobson, R.W.; Durán, W.N. Hyperglycemia Alters PI3k and Akt Signaling and Leads to Endothelial Cell Proliferative Dysfunction. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H1744–H1751. [Google Scholar] [CrossRef]
- Bartlett, C.S.; Jeansson, M.; Quaggin, S.E. Vascular Growth Factors and Glomerular Disease. Annu. Rev. Physiol. 2016, 78, 437–461. [Google Scholar] [CrossRef]
- Chyła, G.; Sałaga-Zaleska, K.; Dąbkowski, K.; Kuchta, A.; Jankowski, M. Suramin Enhances the Urinary Excretion of VEGF-A in Normoglycemic and Streptozotocin-Induced Diabetic Rats. Pharmacol. Rep. 2021, 73, 841–846. [Google Scholar] [CrossRef]
- Chyła-Danił, G.; Sałaga-Zaleska, K.; Kreft, E.; Krzesí Nska, A.; Herman, S.; Kuchta, A.; Sakowicz-Burkiewicz, M.; Lenartowicz, M.; Jankowski, M. Suramin Affects the Renal VEGF-A/VEGFR Axis in Short-Term Streptozotocin-Induced Diabetes. Pharmaceuticals 2023, 16, 470. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Tolbert, E.; Pang, M.; Ponnusamy, M.; Yan, H.; Zhuang, S. Suramin Inhibits Renal Fibrosis in Chronic Kidney Disease. J. Am. Soc. Nephrol. 2011, 22, 1064. [Google Scholar] [CrossRef] [PubMed]
- Korrapati, M.C.; Shaner, B.E.; Neely, B.A.; Alge, J.L.; Arthur, J.M.; Schnellmann, R.G. Diabetes-Induced Renal Injury in Rats Is Attenuated by Suramin. J. Pharmacol. Exp. Ther. 2012, 343, 34–43. [Google Scholar] [CrossRef]
- Korrapati, M.C.; Howell, L.H.; Shaner, B.E.; Megyesi, J.K.; Siskind, L.J.; Schnellmann, R.G. Suramin: A Potential Therapy for Diabetic Nephropathy. PLoS ONE 2013, 8, e73655. [Google Scholar] [CrossRef]
- Voogd, T.E.; Vansterkenburg, E.L.M.; Wilting, J.; Janssen, L.H.M. Recent Research on the Biological Activity of Suramin. Pharmacol. Rev. 1993, 45, 177–203. [Google Scholar]
- Cooper, M.E.; Vranes, D.; Youssef, S.; Stacker, S.A.; Cox, A.J.; Rizkalla, B.; Casley, D.J.; Bach, L.A.; Kelly, D.J.; Gilbert, R.E. Increased Renal Expression of Vascular Endothelial Growth Factor (VEGF) and Its Receptor VEGFR-2 in Experimental Diabetes. Diabetes 1999, 48, 2229–2239. [Google Scholar] [CrossRef] [PubMed]
- Zafar, M.I.; Mills, K.; Ye, X.; Blakely, B.; Min, J.; Kong, W.; Zhang, N.; Gou, L.; Regmi, A.; Hu, S.Q.; et al. Association between the Expression of Vascular Endothelial Growth Factors and Metabolic Syndrome or Its Components: A Systematic Review and Meta-Analysis. Diabetol. Metab. Syndr. 2018, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Eremina, V.; Sood, M.; Haigh, J.; Nagy, A.; Lajoie, G.; Ferrara, N.; Gerber, H.P.; Kikkawa, Y.; Miner, J.H.; Quaggin, S.E. Glomerular-Specific Alterations of VEGF-A Expression Lead to Distinct Congenital and Acquired Renal Diseases. J. Clin. Investig. 2003, 111, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Sivaskandarajah, G.A.; Jeansson, M.; Maezawa, Y.; Eremina, V.; Baelde, H.J.; Quaggin, S.E. Vegfa Protects the Glomerular Microvasculature in Diabetes. Diabetes 2012, 61, 2958–2966. [Google Scholar] [CrossRef]
- Hohenstein, B.; Hausknecht, B.; Boehmer, K.; Riess, R.; Brekken, R.A.; Hugo, C.P.M. Local VEGF Activity but Not VEGF Expression Is Tightly Regulated during Diabetic Nephropathy in Man. Kidney Int. 2006, 69, 1654–1661. [Google Scholar] [CrossRef]
- Tanabe, K.; Maeshima, Y.; Sato, Y.; Wada, J. Antiangiogenic Therapy for Diabetic Nephropathy. Biomed. Res. Int. 2017, 2017, 5724069. [Google Scholar] [CrossRef] [PubMed]
- McNally, W.P.; DeHart, P.D.; Lathia, C.; Whitfield, L.R. Distribution of [14C]Suramin in Tissues of Male Rats Following a Single Intravenous Dose. Life Sci. 2000, 67, 1847–1857. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xiang, H.; Lu, Y.; Wu, T.; Ji, G. New Progress in Drugs Treatment of Diabetic Kidney Disease. Biomed. Pharmacother. 2021, 141, 111918. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Baena, G.; Armstrong, S.D.; Halstead, J.O.; Prescott, M.; Roberts, S.A.; Mclean, L.; Mudge, J.M.; Hurst, J.L.; Beynon, R.J. Molecular Complexity of the Major Urinary Protein System of the Norway Rat, Rattus Kingdom. Cient. Rep. 2019, 9, 10757. [Google Scholar] [CrossRef]
- Omez-Baena, G.G.; Armstrong, S.D.; Phelan, M.M.; Hurst, J.L.; Beynon, R.J. The Major Urinary Protein System in the Rat. Biochem. Soc. Trans. 2014, 42, 886–892. [Google Scholar] [CrossRef]
- Maric-Bilkan, C.; Flynn, E.R.; Chade, A.R. Microvascular Disease Precedes the Decline in Renal Function in the Streptozotocin-Induced Diabetic Rat. Am. J. Physiol. Ren. Physiol. 2012, 302, F308–F319. [Google Scholar] [CrossRef]
- Edgley, A.J.; Tare, M.; Evans, R.G.; Skordilis, C.; Parkington, H.C. In Vivo Regulation of Endothelium-Dependent Vasodilation in the Rat Renal Circulation and the Effect of Streptozotocin-Induced Diabetes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R829–R839. [Google Scholar] [CrossRef]
- Johnstone, M.T.; Creager, S.J.; Scales, K.M.; Cusco, J.A.; Lee, B.K.; Creager, M.A. Impaired Endothelium-Dependent Vasodilation in Patients with Insulin-Dependent Diabetes Mellitus. Circulation 1993, 88, 5210–5216. [Google Scholar] [CrossRef]
- Kamata, K.; Miyata, N.; Kasuya, Y. Impairment of Endothelium-Dependent Relaxation and Changes in Levels of Cyclic GMP in Aorta from Streptozotocin-Induced Diabetic Rats. Br. J. Pharmacol. 1989, 97, 614. [Google Scholar] [CrossRef]
- Jourde-Chiche, N.; Fakhouri, F.; Dou, L.; Bellien, J.; Burtey, S.; Frimat, M.; Jarrot, P.A.; Kaplanski, G.; le Quintrec, M.; Pernin, V.; et al. Endothelium Structure and Function in Kidney Health and Disease. Nat. Rev. Nephrol. 2019, 15, 87–108. [Google Scholar] [CrossRef]
- Sahu, D.; Saroha, A.; Roy, S.; Das, S.; Srivastava, P.S.; Das, H.R. Suramin Ameliorates Collagen Induced Arthritis. Int. Immunopharmacol. 2012, 12, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Bardal, S.; Misurski, D.; Qiu, X.; Desai, K.; McNeill, J.R. Chronic Treatment with Vascular Endothelial Growth Factor Preserves Agonist-Evoked Vascular Responses in the Streptozotocin-Induced Diabetic Rat. Diabetologia 2006, 49, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Undas, A.; Wiek, I.; Stêpień, E.; Zmudka, K.; Tracz, W. Hyperglycemia Is Associated with Enhanced Thrombin Formation, Platelet Activation, and Fibrin Clot Resistance to Lysis in Patients with Acute Coronary Syndrome. Diabetes Care 2008, 31, 1590–1595. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Shou, W.; Wu, W.; Wang, G.; Cui, W. Performance Evaluation of Thrombomodulin, Thrombin-Antithrombin Complex, Plasmin-A2-Antiplasmin Complex, and t-PA: PAI-1 Complex. J. Clin. Lab. Anal. 2019, 33, e22913. [Google Scholar] [CrossRef]
- Majumder, S.; Advani, A. VEGF and the Diabetic Kidney: More than Too Much of a Good Thing. J. Diabetes Complicat. 2017, 31, 273–279. [Google Scholar] [CrossRef]
- Carranza, K.; Veron, D.; Cercado, A.; Bautista, N.; Pozo, W.; Tufro, A.; Veron, D. Cellular and Molecular Aspects of Diabetic Nephropathy; the Role of VEGF-A. Nefrol. Engl. Ed. 2015, 35, 131–138. [Google Scholar] [CrossRef]
- Oates, J.C.; Russell, D.L.; van Beusecum, J.P. Endothelial Cells: Potential Novel Regulators of Renal Inflammation. Am. J. Physiol. Renal Physiol. 2022, 322, F309–F321. [Google Scholar] [CrossRef]
- Marfella, R.; Esposito, K.; Giunta, R.; Coppola, G.; de Angelis, L.; Farzati, B.; Paolisso, G.; Giugliano, D. Circulating Adhesion Molecules in Humans: Role of Hyperglycemia and Hyperinsalinemia. Circulation 2000, 101, 2247–2251. [Google Scholar] [CrossRef]
- Gu, H.F.; Ma, J.; Gu, K.T.; Brismar, K. Association of Intercellular Adhesion Molecule 1 (ICAM1) with Diabetes and Diabetic Nephropathy. Front. Endocrinol. 2013, 3, 179. [Google Scholar] [CrossRef]
- Leeuwenberg, J.F.; Smeets, E.F.; Neefjes, J.J.; Shaffer, M.A.; Cinek, T.; Jeunhomme, T.M.; Ahern, T.J.; Buurman, W.A. E-Selectin and Intercellular Adhesion Molecule-1 Are Released by Activated Human Endothelial Cells in Vitro. Immunology 1992, 77, 543. [Google Scholar]
- Nacci, C.; Tarquinio, M.; De Benedictis, L.; Mauro, A.; Zigrino, A.; Carratù, M.R.; Quon, M.J.; Montagnani, M. Endothelial Dysfunction in Mice with Streptozotocin-Induced Type 1 Diabetes Is Opposed by Compensatory Overexpression of Cyclooxygenase-2 in the Vasculature. Endocrinology 2009, 150, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Lhotta, K.; Schlögl, A.; Kronenberg, F.; Joannidis, M.; König, P. Soluble Intercellular Adhesion Molecule-1 (ICAM-1) in Serum and Urine: Correlation with Renal Expression of ICAM-1 in Patients with Kidney Disease. Clin. Nephrol. 1997, 48, 85–91. [Google Scholar] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Besseling, P.J.; Pieters, T.T.; Nguyen, I.T.N.; de Bree, P.M.; Willekes, N.; Dijk, A.H.; Bovee, D.M.; Hoorn, E.J.; Rookmaaker, M.B.; Gerritsen, K.G.; et al. A Plasma Creatinine- And Urea-Based Equation to Estimate Glomerular Filtration Rate in Rats. Am. J. Physiol. Ren. Physiol. 2021, 320, F518–F524. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Week | Experimental Groups | |||
|---|---|---|---|---|---|
| CON | SUR | STZ | STZ + SUR | ||
| Body weight, (g) | 0 | 266 ±10 | 277 ± 10 | 301 ± 12 | 282 ± 15 |
| 1 | 290 ± 9 * | 302 ± 10 * | 279 ± 11 | 278 ± 9 | |
| 12 | 416 ± 6 *# | 420 ± 12 *# | 268 ± 14 *Δ | 297 ± 25 | |
| Diuresis, (mL/24 h) | 0 | 5.8 <4.3; 9.2> | 7.5 <6.6; 8.4> | 5.1 <4.7; 6.5> | 6.9 <6.1; 8.3> |
| 1 | 8.4 ± 1.5 * | 7.1 ± 0.6 | 93.5 ± 11.2 *Δ | 90.6 ± 12.2 * | |
| 12 | 9.3 ± 0.6 | 8.5 ± 0.6 # | 119.5 ± 10.2 *Δ | 130.8 ± 11.6 *# | |
| Water intake, (mL) | 0 | 21.0 <20.6; 25.4> | 21.3 <19.0; 24.7> | 21.2 <16.1; 25.9> | 26.1 <22.8; 26.7> |
| 1 | 26.5 ± 1.8 * | 22.8 ± 3.9 | 101.9 ± 9.9 *Δ | 107.0 ± 11.5 * | |
| 12 | 21.0 ± 1.3 # | 20.4 ± 3.5 | 128.2 ± 10.9 *Δ | 142.0 ± 11.4 *# | |
| Blood glucose, (mg/dL) | 0 | 123 ± 3 | 125 ± 7 | 116 ± 5 | 123 ± 5 |
| 1 | 116 ± 1 * | 125 ± 2 | 486 ± 33 *Δ | 484 ± 24 * | |
| 12 | 115 <106; 122> | 126 <109; 134> | 600 <525; 600> *Δ | 586 <551; 600> *# | |
| Parameter | Week | Experimental Groups | |||
|---|---|---|---|---|---|
| CON | SUR | STZ | STZ + SUR | ||
| Food intake, (g) | 0 | 18.5 <17.0; 21.0> | 20.3 <17.9; 22.5> | 19.3 <18.4; 29.8> | 26.3 <24.8; 28.1> |
| 12 | 16.2 ± 1.1 | 18.1 ± 2.0 | 31.9 ± 1.7 *Δ | 34.5 ± 1.4 * | |
| Specific rate of body mass gain, (g/kg) | 12 | 443.0 ± 42.6 | 396.1 ± 54.2 | −40.4 ± 25.7 & | 80.6 ± 111.1 |
| Feed efficiency ratio | 12 | 52.61 ± 5.03 | 45.55 ± 5.71 | −3.15 ± 1.95 & | 4.42 ± 6.86 |
| Efficiency of food utilization for body weight, (g/cal) | 12 | 0.19 ± 0.02 | 0.17 ± 0.02 | −0.01 ± 0.01 & | 0.02 ± 0.02 |
| Gene Transcript | Accession No. | Oligonucleotide Sequence 5′-3′ | Universal ProbeLibrary Probe |
|---|---|---|---|
| rFlt1 (Vegfr1) | D28498 | (F) cagtttccaagtggccagag | #22 |
| (R) aggtcgcgatgaatgcac | |||
| rFlk1 (Vegfr2) | U93306 | (F) gagacccgcgttttcaga | #65 |
| (R) aagaacaatatagtctttgccatcc | |||
| Actb | Universal ProbeLibrary Rat Actb Gene Assay (Roche, Cat #05046203001) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chyła-Danił, G.; Sałaga-Zaleska, K.; Kreft, E.; Stumski, O.; Krzesińska, A.; Sakowicz-Burkiewicz, M.; Kuchta, A.; Jankowski, M. Long-Term Effects of Suramin on Renal Function in Streptozotocin-Induced Diabetes in Rats. Int. J. Mol. Sci. 2023, 24, 14671. https://doi.org/10.3390/ijms241914671
Chyła-Danił G, Sałaga-Zaleska K, Kreft E, Stumski O, Krzesińska A, Sakowicz-Burkiewicz M, Kuchta A, Jankowski M. Long-Term Effects of Suramin on Renal Function in Streptozotocin-Induced Diabetes in Rats. International Journal of Molecular Sciences. 2023; 24(19):14671. https://doi.org/10.3390/ijms241914671
Chicago/Turabian StyleChyła-Danił, Gabriela, Kornelia Sałaga-Zaleska, Ewelina Kreft, Olaf Stumski, Aleksandra Krzesińska, Monika Sakowicz-Burkiewicz, Agnieszka Kuchta, and Maciej Jankowski. 2023. "Long-Term Effects of Suramin on Renal Function in Streptozotocin-Induced Diabetes in Rats" International Journal of Molecular Sciences 24, no. 19: 14671. https://doi.org/10.3390/ijms241914671
APA StyleChyła-Danił, G., Sałaga-Zaleska, K., Kreft, E., Stumski, O., Krzesińska, A., Sakowicz-Burkiewicz, M., Kuchta, A., & Jankowski, M. (2023). Long-Term Effects of Suramin on Renal Function in Streptozotocin-Induced Diabetes in Rats. International Journal of Molecular Sciences, 24(19), 14671. https://doi.org/10.3390/ijms241914671