
A Novel NOX Inhibitor Alleviates Parkinson’s Disease Pathology in PFF-Injected Mice

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
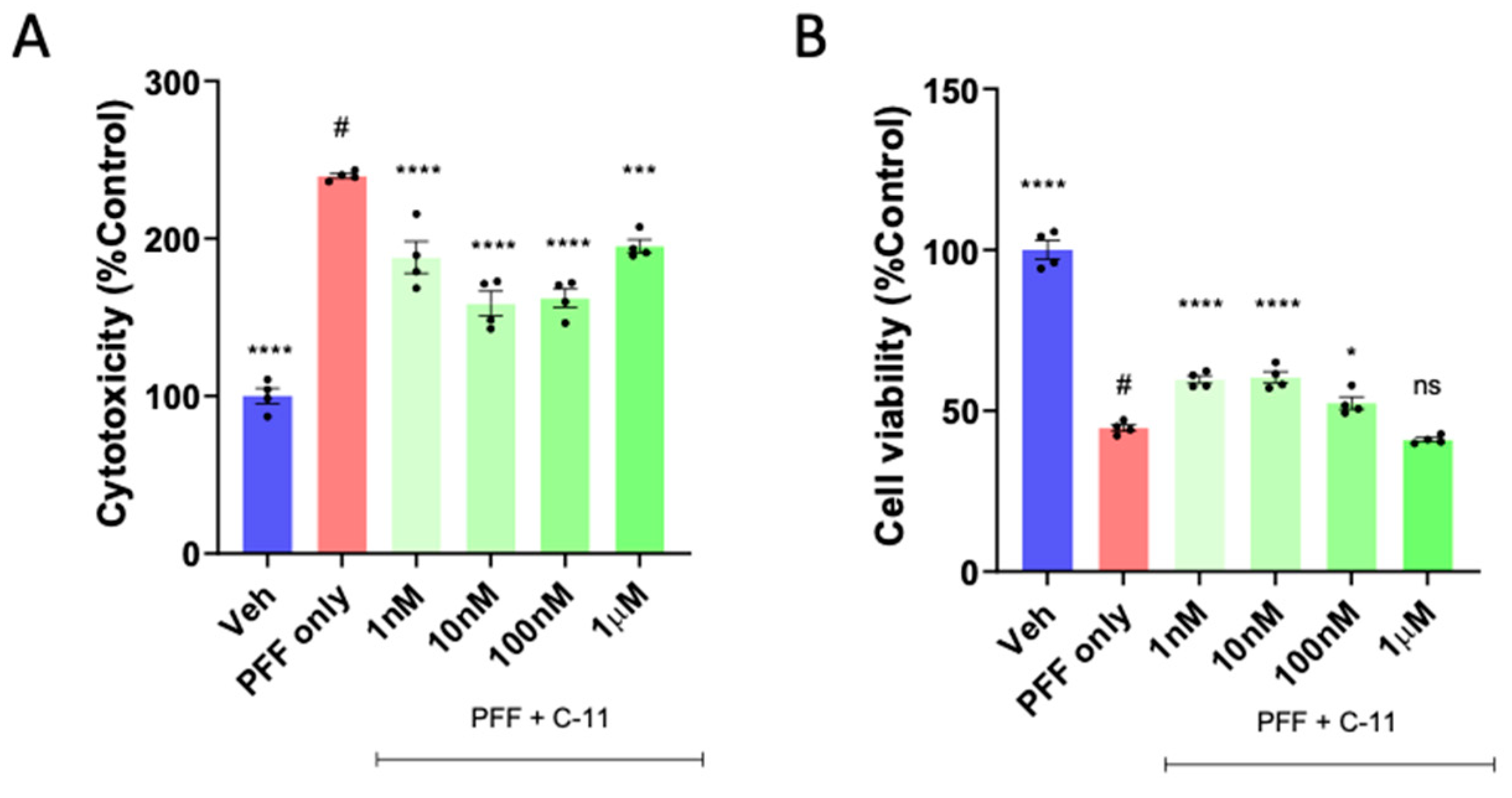
2.1. A Novel NOX Inhibitor Treatment Decreases Cytotoxicity and Increases Cell Viability in PFF Treated N27P Dopaminergic Cells
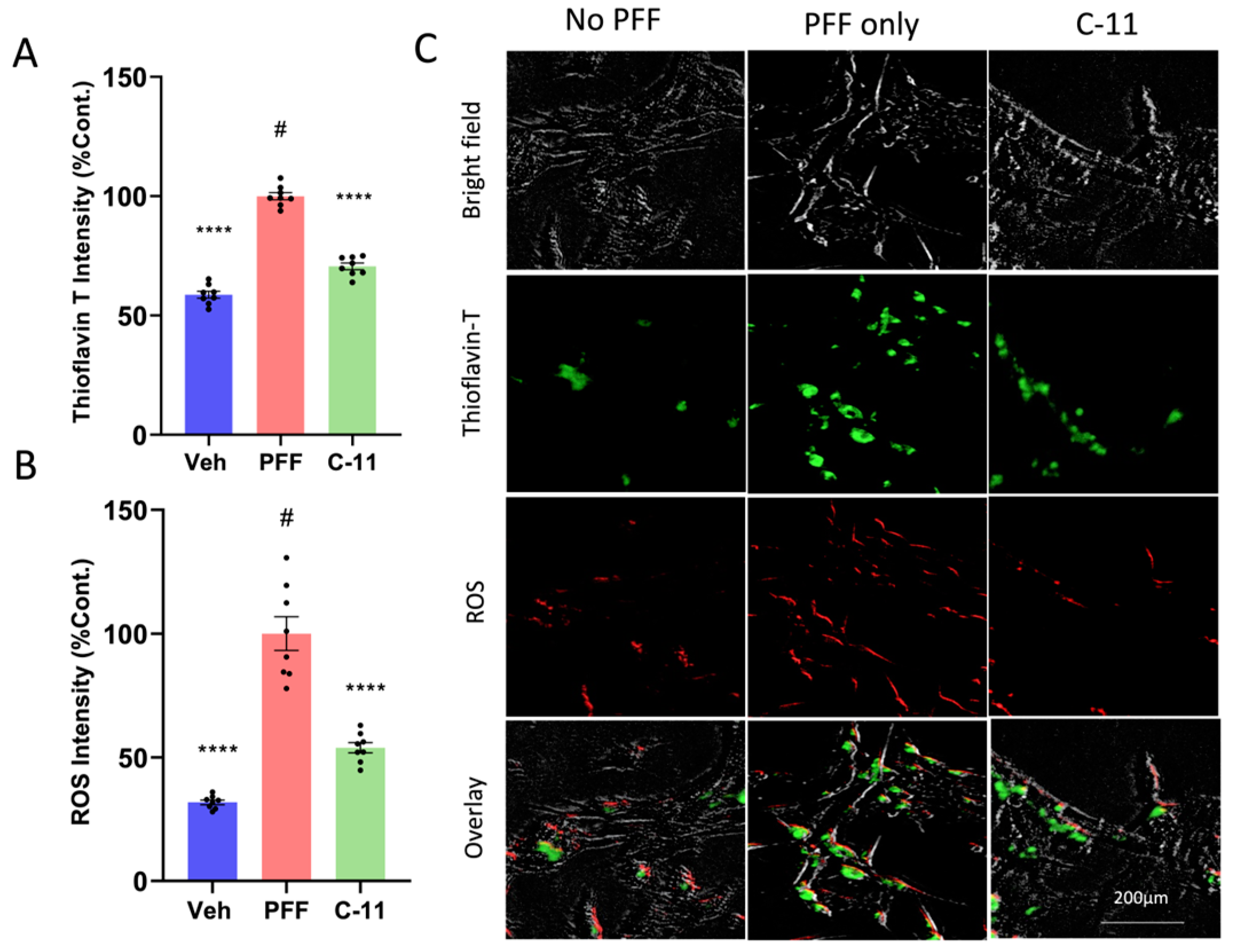
2.2. The C-11 Treatment Decreases Protein Aggregation and ROS Generation Induced by PFF
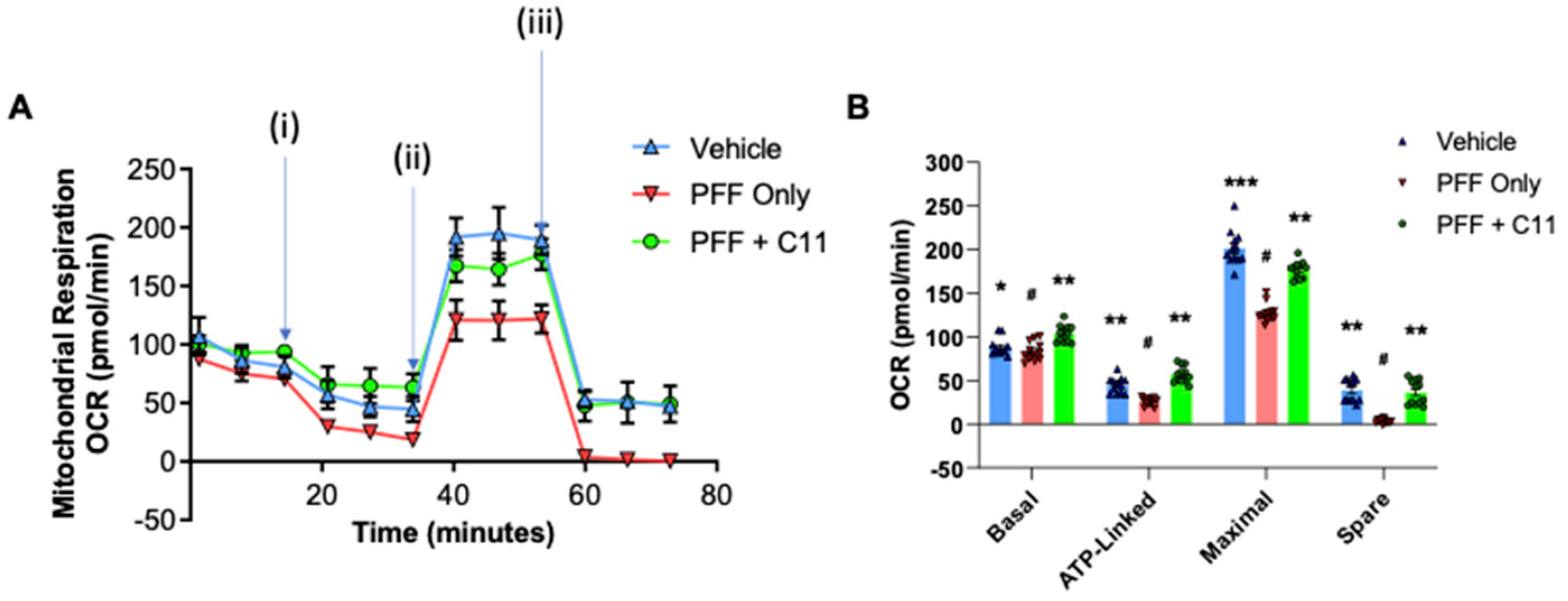
2.3. The C-11 Treatment Prevents PFF-Induced Mitochondrial Dysfunctions in N27 Cells
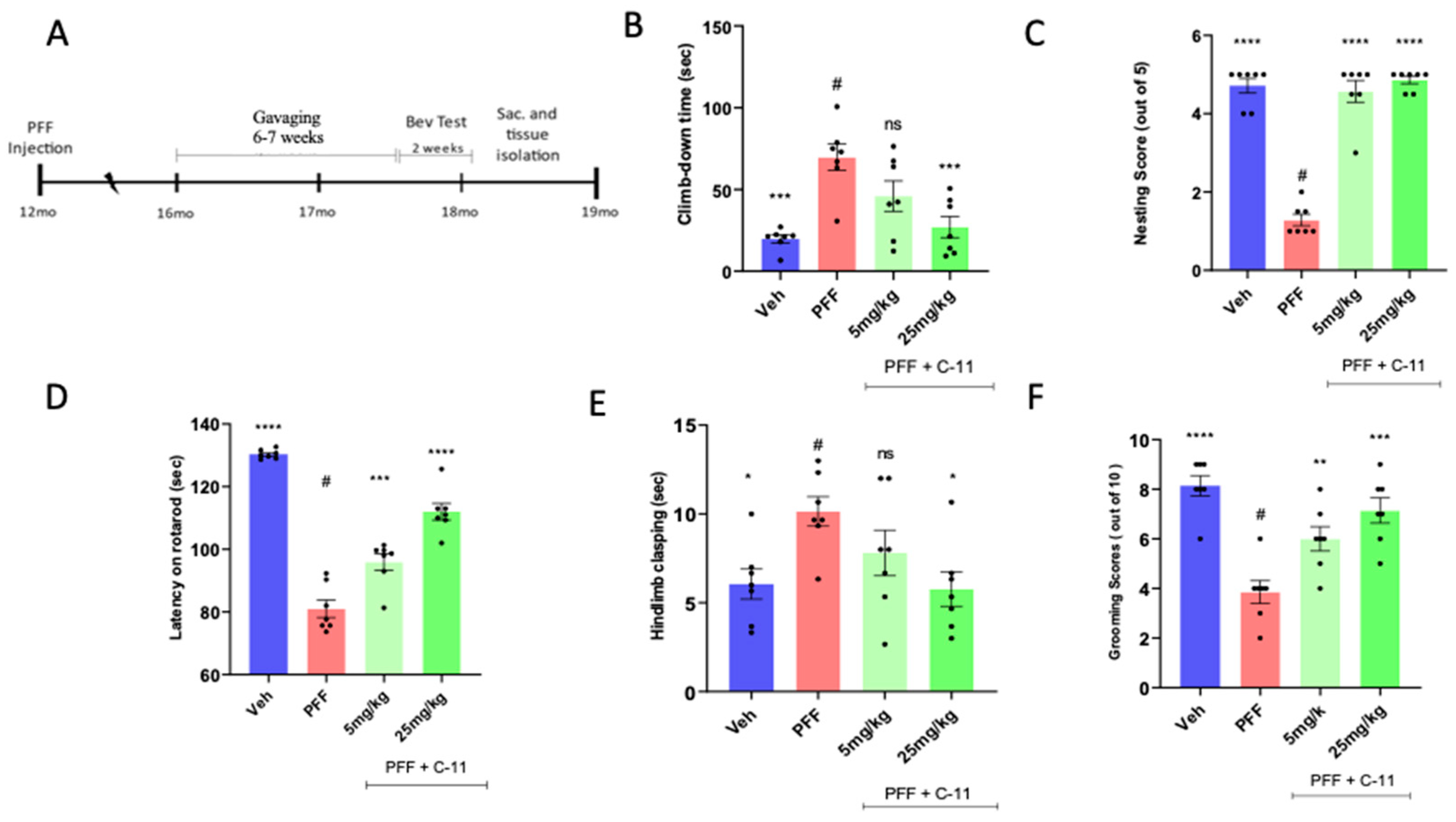
2.4. In Behavioral Analyses, the Oral C-11 Treatment Reduces Motor Deficits in PFF Injected Mice
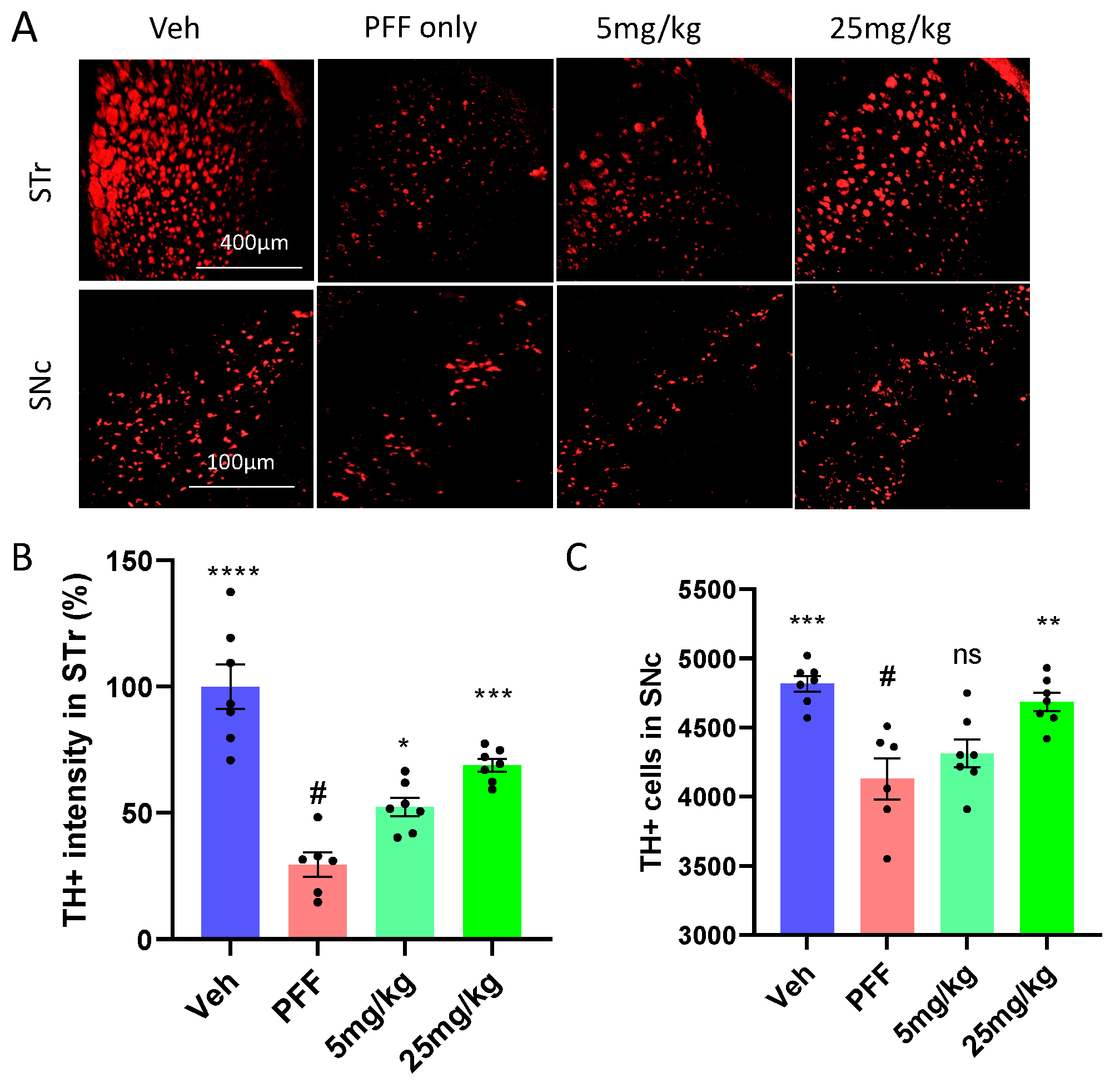
2.5. The Neuronal Loss in the STr and SNc Induced by PFF Was Prevented or Partially Reversed by the C-11 Treatment
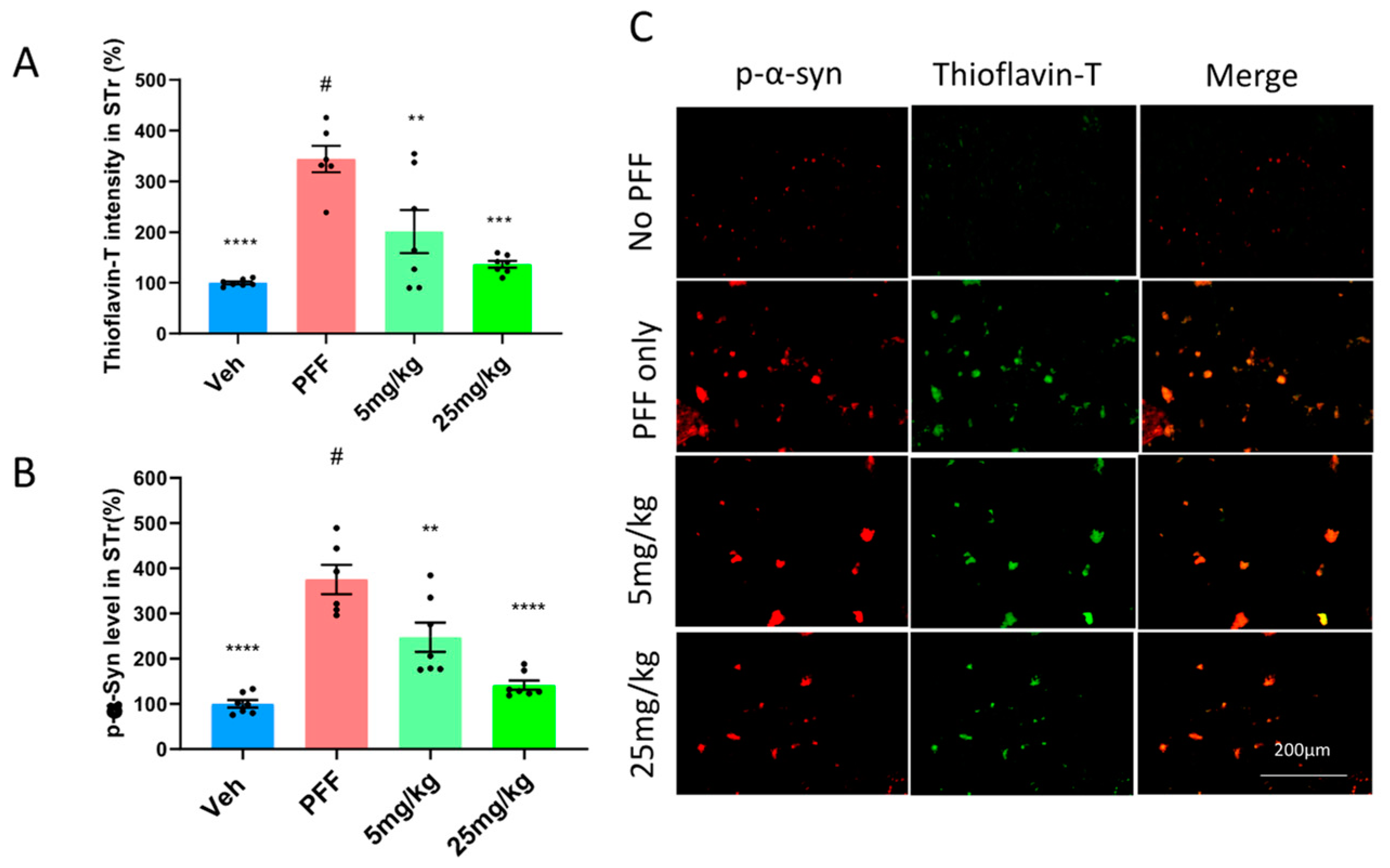
2.6. The C-11 Reduces PFF-Induced Protein Aggregates in Thioflavin-T Stain & Phospho-α-syn in the Striatum (STr) and Substantia Nigra Compacta (SNc)
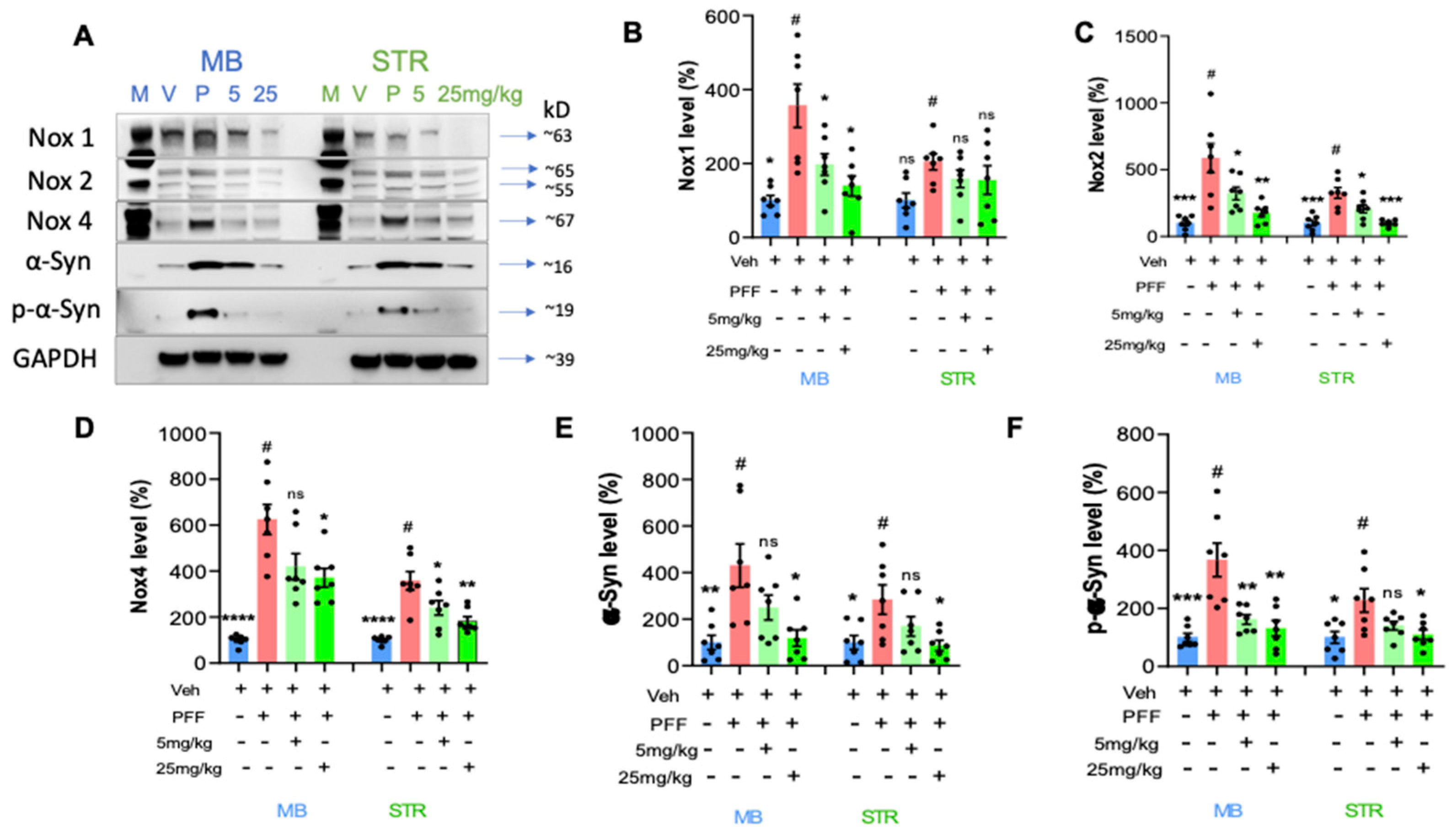
2.7. The Levels of Phospho-α-Synuclein, NOX-1, 2, & 4 in the STr and the Ventral Midbrain Were Reduced by the Treatment in Western Blot Analysis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Culture and Treatment
4.3. Cell Viability Assay
4.4. Cytotoxicity Assay
4.5. ROS Levels and Thioflavin-T assessment
4.6. Oxygen Consumption Rate (OCR) Measurements Using Agilent Seahorse XF Analyzer
4.7. Stereotaxic α-Synuclein PFF Injection and Brain Isolation
4.8. Gavaging Schedule
4.9. Tissue Isolation
4.10. Behavioral Assays
4.10.1. Hindlimb Clasping
4.10.2. Rotarod
4.10.3. Pole Test
4.10.4. Nesting
4.10.5. Grooming
4.11. Immunohistochemistry (IHC)
4.12. Immunoblot Analysis
4.13. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parkinson, J. An essay on the shaking palsy 1817. J. Neuropsychiatry Clin Neurosci. 2002, 14, 223–236. [Google Scholar] [CrossRef]
- Dias, V.; Junn, E.; Mouradian, M.M. The role of oxidative stress in Parkinson’s disease. J. Parkinson’s Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef] [PubMed]
- Angelova, P.R.; Abramov, A.Y. Role of Mitochondrial ROS in the Brain: From Physiology to Neurodegeneration. FEBS Lett. 2018, 592, 692–702. [Google Scholar] [CrossRef]
- Risiglione, P.; Zinghirino, F.; Di Rosa, M.C.; Magrì, A.; Messina, A. Alpha-Synuclein and Mitochondrial Dysfunction in Parkinson’s Disease: The Emerging Role of VDAC. Biomolecules 2021, 11, 718. [Google Scholar] [CrossRef] [PubMed]
- Payne, B.A.I.; Chinnery, P.F. Mitochondrial dysfunction in aging: Much progress but many unresolved questions. Biochim. Biophys. Acta. 2015, 1847, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Li, W.G.; Miller, F.J.; Zhang, H.J.; Spitz, D.R.; Oberley, L.W.; Weintraub, N.L. H(2)O(2)-induced O(2) production by a non-phagocytic NAD(P)H oxidase causes oxidant injury. J. Biol. Chem. 2001, 276, 29251–29256. [Google Scholar] [CrossRef] [PubMed]
- Jenner, P.; Olanow, C.W. The Pathogenesis of Cell Death in Parkinson’s Disease. Neurology 2006, 66, S24–S36. [Google Scholar] [CrossRef]
- Choi, D.H.; Cristόvão, A.C.; Guhathakurta, S.; Lee, J.; Joh, T.H.; Beal, M.F.; Kim, Y.-S. NADPH Oxidase 1-Mediated Oxidative Stress Leads to Dopamine Neuron Death in Parkinson’s Disease. Antioxid. Redox Signal. 2012, 16, 1033–1045. [Google Scholar] [CrossRef]
- Abbott, A. Levodopa: The story so far. Nature 2010, 466, S6–S7. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Bánfi, B.; Clark, R.A.; Steger, K.; Krause, K.-H. Two Novel Proteins Activate Superoxide Generation by the NADPH Oxidase NOX1. J. Biol. Chem. 2003, 278, 3510–3513. [Google Scholar] [CrossRef] [PubMed]
- Marden, J.J.; Harraz, M.M.; Williams, A.J.; Nelson, K.; Luo, M.; Paulson, H.; Engelhardt, J.F. Redox Modifier Genes in Amyotrophic Lateral Sclerosis in Mice. J. Clin. Investig. 2007, 117, 2913–2919. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.H. Mitochondria and Neuronal Death/Survival Signaling Pathways in Cerebral Ischemia. Neurochem. Res. 2004, 29, 1943–1949. [Google Scholar] [CrossRef]
- Head, E. Oxidative Damage and Cognitive Dysfunction: Antioxidant Treatments to Promote Healthy Brain Aging. Neurochem. Res. 2009, 34, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Cooney, S.J.; Bermudez-Sabogal, S.L.; Byrnes, K.R. Cellular and Temporal Expression of NADPH Oxidase (NOX) Isotypes after Brain Injury. J. Neuroinflammation 2013, 10, 917. [Google Scholar] [CrossRef]
- Verma, D.K.; Seo, B.A.; Ghosh, A.; Ma, S.X.; Hernandez-Quijada, K.; Andersen, J.K.; Ko, H.S.; Kim, Y.H. Alpha-Synuclein Preformed Fibrils Induce Cellular Senescence in Parkinson’s Disease Models. Cells 2021, 10, 1694. [Google Scholar] [CrossRef]
- Ghosh, A.A.; Verma, D.K.; Cabrera, G.; Ofori, K.; Hernandez-Quijada, K.; Kim, J.K.; Chung, J.H.; Moore, M.; Moon, S.H.; Seo, J.B.; et al. A Novel NOX Inhibitor Treatment Attenuates Parkinson’s Disease-Related Pathology in Mouse Models. Int. J. Mol. Sci. 2022, 23, 4262. [Google Scholar] [CrossRef]
- Dumont, M.; Beal, M.F. Neuroprotective strategies involving ROS in Alzheimer’s disease. Free Radic. Biol. Med. 2011, 51, 1014–1026. [Google Scholar] [CrossRef]
- Volpicelli-Daley, L.A.; Luk, K.C.; Patel, T.P.; Tanik, S.A.; Riddle, D.M.; Stieber, A.; Meaney, D.F.; Trojanowski, J.Q.; Lee, V.M.-Y. Exogenous α-Synuclein Fibrils Induce Lewy Body Pathology Leading to Synaptic Dysfunction and Neuron Death. Neuron 2011, 72, 57–71. [Google Scholar] [CrossRef]
- Polinski, N.K.; Volpicelli-Daley, L.A.; Sortwell, C.E.; Luk, K.C.; Cremades, N.; Gottler, L.M.; Froula, J.; Duffy, M.F.; Lee, V.M.Y.; Martinez, T.N.; et al. Best Practices for Generating and Using Alpha-Synuclein Pre-Formed Fibrils to Model Parkinson’s Disease in Rodents. J. Parkinsons Dis. 2018, 8, 303–322. [Google Scholar] [CrossRef]
- Murphy, M.P.; Bayir, H.; Belousov, V.; Chang, C.J.; Davies, K.J.; Davies, M.J.; Dick, T.P.; Finkel, T.; Forman, H.J.; Janssen-Heininger, Y.; et al. Guidelines for measuring reactive oxygen species and oxidative damage in cells and in vivo. Nat. Metab. 2022, 4, 651–662. [Google Scholar] [CrossRef]
- Chung, H.K.; Ho, H.A.; Pérez-Acuña, D.; Lee, S.J. Modeling α-Synuclein Propagation with Preformed Fibril Injections. J. Mov. Disord. 2019, 12, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.M.; Wilson-Delfosse, A.L.; Mieyal, J.J. Dysregulation of glutathione homeostasis in neurodegenerative diseases. Nutrients 2012, 4, 1399–1440. [Google Scholar] [CrossRef]
- Matsuura, K.; Kabuto, H.; Makino, H.; Ogawa, N. Pole test is a useful method for evaluating the mouse movement disorder caused by striatal dopamine depletion. J. Neurosci. Methods 1997, 73, 45–48. [Google Scholar] [CrossRef]
- Sato, H.; Kato, T.; Arawaka, S. The role of Ser129 phosphorylation of α-synuclein in neurodegeneration of Parkinson’s disease: A review of in vivo models. Rev. Neurosci. 2013, 24, 115–123. [Google Scholar] [CrossRef]
- Verma, D.K.; Gupta, S.; Biswas, J.; Joshi, N.; Singh, A.; Gupta, P.; Tiwari, S.; Raju, K.S.; Chaturvedi, S.; Wahajuddin, M.; et al. New therapeutic activity of metabolic enhancer piracetam in treatment of neurodegenerative disease: Participation of caspase independent death factors, oxidative stress, inflammatory responses and apoptosis. Biochim. Biophys. Acta. Mol. Basis Dis. 2018, 1864, 2078–2096. [Google Scholar] [CrossRef]
- Verma, D.K.; Singh, D.K.; Gupta, S.; Gupta, P.; Singh, A.; Biswas, J.; Singh, S. Minocycline diminishes the rotenone induced neurotoxicity and glial activation via suppression of apoptosis, nitrite levels and oxidative stress. Neurotoxicology 2018, 65, 9–21. [Google Scholar] [CrossRef]
- Verma, D.K.; Gupta, S.; Biswas, J.; Joshi, N.; Sivarama Raju, K.; Wahajuddin, M.; Singh, S. Metabolic Enhancer Piracetam Attenuates the Translocation of Mitochondrion-Specific Proteins of Caspase-Independent Pathway, Poly [ADP-Ribose] Polymerase 1 Up-regulation and Oxidative DNA Fragmentation. Neurotox. Res. 2018, 34, 198–219. [Google Scholar] [CrossRef] [PubMed]
- Biswas, J.; Gupta, S.; Verma, D.K.; Gupta, P.; Singh, A.; Tiwari, S.; Goswami, P.; Sharma, S.; Singh, S. Involvement of glucose related energy crisis and endoplasmic reticulum stress: Insinuation of streptozotocin induced Alzheimer’s like pathology. Cell. Signal. 2018, 42, 211–226. [Google Scholar] [CrossRef]
- Gupta, S.; Verma, D.K.; Biswas, J.; Raju, K.S.; Joshi, N.; Wahajuddin, M.; Singh, S. The metabolic enhancer piracetam attenuates mitochondrion-specific endonuclease G translocation and oxidative DNA fragmentation. Free Radic. Bio. Med. 2014, 73, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Boyd-Kimball, D. Mitochondrial Oxidative and Nitrosative Stress and Alzheimer Disease. Antioxidants 2020, 9, 818. [Google Scholar] [CrossRef] [PubMed]
- Verma, D.K.; Joshi, N.; Raju, K.S.; Wahajuddin, M.; Singh, R.K.; Singh, S. Metabolic enhancer piracetam attenuates rotenone induced oxidative stress: A study in different rat brain regions. Acta. Neurobiol. Exp. 2015, 75, 399–411. [Google Scholar]
- Heger, L.M.; Wise, R.M.; Hees, J.T.; Harbauer, A.B.; Burbulla, L.F. Mitochondrial Phenotypes in Parkinson’s Diseases-A Focus on Human iPSC-Derived Dopaminergic Neurons. Cells 2021, 10, 3436. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Kanthasamy, A.; Ghosh, A.; Anantharam, V.; Kalyanaraman, B.; Kanthasamy, A.G. Mitochondria-targeted antioxidants for treatment of Parkinson’s disease: Preclinical and clinical outcomes. Biochim. Biophys. Acta 2014, 1842, 1282–1294. [Google Scholar] [CrossRef]
- Muhammad, M.H.; Allam, M.M. Resveratrol and/or exercise training counteract aging-associated decline of physical endurance in aged mice; targeting mitochondrial biogenesis and function. J. Physiol Sci. 2018, 68, 681–688. [Google Scholar] [CrossRef]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The Role of Oxidative Stress in Neurodegenerative Diseases. Experimental Neurobiology. Exp. Neurobiol. 2015, 24, 325–340. [Google Scholar] [CrossRef]
- Infanger, D.W.; Sharma, R.V.; Davisson, R.L. NADPH Oxidases of the Brain: Distribution, Regulation, and Function. Antioxid Redox Signal. 2006, 8, 1583–1596. [Google Scholar] [CrossRef]
- Rey, F.E.; Cifuentes, M.E.; Kiarash, A.; Quinn, M.T.; Pagano, P.J. Novel competitive inhibitor of NAD(P)H oxidase assembly attenuates vascular O(2)(-) and systolic blood pressure in mice. Circ. Res. 2001, 89, 408–414. [Google Scholar] [CrossRef]
- Guo, S.; Chen, X. The human Nox4: Gene, structure, physiological function and pathological significance. J. Drug Target. 2015, 23, 888–896. [Google Scholar] [CrossRef]
- Masliah, E.; Rockenstein, E.; Veinbergs, I.; Mallory, M.; Hashimoto, M.; Takeda, A.; Sagara, Y.; Sisk, A.; Mucke, L. Dopaminergic Loss, and Inclusion Body Formation in Alpha-Synuclein Mice: Implications for Neurodegenerative Disorders. Science 2000, 287, 1265–1269. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Thompson, E.W.; Gunter, J.H. Studying the Metabolism of Epithelial-Mesenchymal Plasticity Using the Seahorse Xfe96 Extracellular Flux Analyzer. Methods Mol. Biol. 2021, 2179, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.H.; Wang, X.; Zhu, X. Mitochondrial defects and oxidative stress in Alzheimer disease and Parkinson disease. Free Radic. Biol. Med. 2013, 62, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Lazzara, C.A.; Riley, R.R.; Rane, A.; Andersen, J.K.; Kim, Y.H. The Combination of Lithium and L-Dopa/Carbidopa Reduces MPTP-Induced Abnormal Involuntary Movements (AIMs) via Calpain-1 Inhibition in a Mouse Model: Relevance for Parkinson׳s Disease Therapy. Brain Res. 2015, 1622, 127–136. [Google Scholar] [CrossRef]
- Aniszewska, A.; Bergström, J.; Ingelsson, M.; Ekmark-Lewén, S. Modeling Parkinson’s disease-related symptoms in alpha- synuclein overexpressing mice. Brain Behav. 2022, 12, e2628. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M. Measuring motor coordination in mice. J. Vis. Exp. 2013, 29, e2609. [Google Scholar] [CrossRef]
- Sager, T.N.; Kirchhoff, J.; Mørk, A.; Van Beek, J.; Thirstrup, K.; Didriksen, M.; Lauridsen, J.B. Nest building performance following MPTP toxicity in mice. Behav. Brain Res. 2010, 208, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, A.; Girault, J.A.; Hervé, D. Unilateral Lesion of Dopamine Neurons Induces Grooming Asymmetry in the Mouse. PLoS ONE 2015, 10, e0137185. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ofori, K.; Ghosh, A.; Verma, D.K.; Wheeler, D.; Cabrera, G.; Seo, J.-B.; Kim, Y.-H. A Novel NOX Inhibitor Alleviates Parkinson’s Disease Pathology in PFF-Injected Mice. Int. J. Mol. Sci. 2023, 24, 14278. https://doi.org/10.3390/ijms241814278
Ofori K, Ghosh A, Verma DK, Wheeler D, Cabrera G, Seo J-B, Kim Y-H. A Novel NOX Inhibitor Alleviates Parkinson’s Disease Pathology in PFF-Injected Mice. International Journal of Molecular Sciences. 2023; 24(18):14278. https://doi.org/10.3390/ijms241814278
Chicago/Turabian StyleOfori, Kwadwo, Anurupa Ghosh, Dinesh Kumar Verma, Darice Wheeler, Gabriela Cabrera, Jong-Bok Seo, and Yong-Hwan Kim. 2023. "A Novel NOX Inhibitor Alleviates Parkinson’s Disease Pathology in PFF-Injected Mice" International Journal of Molecular Sciences 24, no. 18: 14278. https://doi.org/10.3390/ijms241814278
APA StyleOfori, K., Ghosh, A., Verma, D. K., Wheeler, D., Cabrera, G., Seo, J.-B., & Kim, Y.-H. (2023). A Novel NOX Inhibitor Alleviates Parkinson’s Disease Pathology in PFF-Injected Mice. International Journal of Molecular Sciences, 24(18), 14278. https://doi.org/10.3390/ijms241814278