
Identification of Candidate Biomarkers of Alzheimer’s Disease via Multiplex Cerebrospinal Fluid and Serum Proteomics

Abstract
:1. Introduction
2. Results
2.1. Demographic and Clinical Characteristics of the Study Subjects
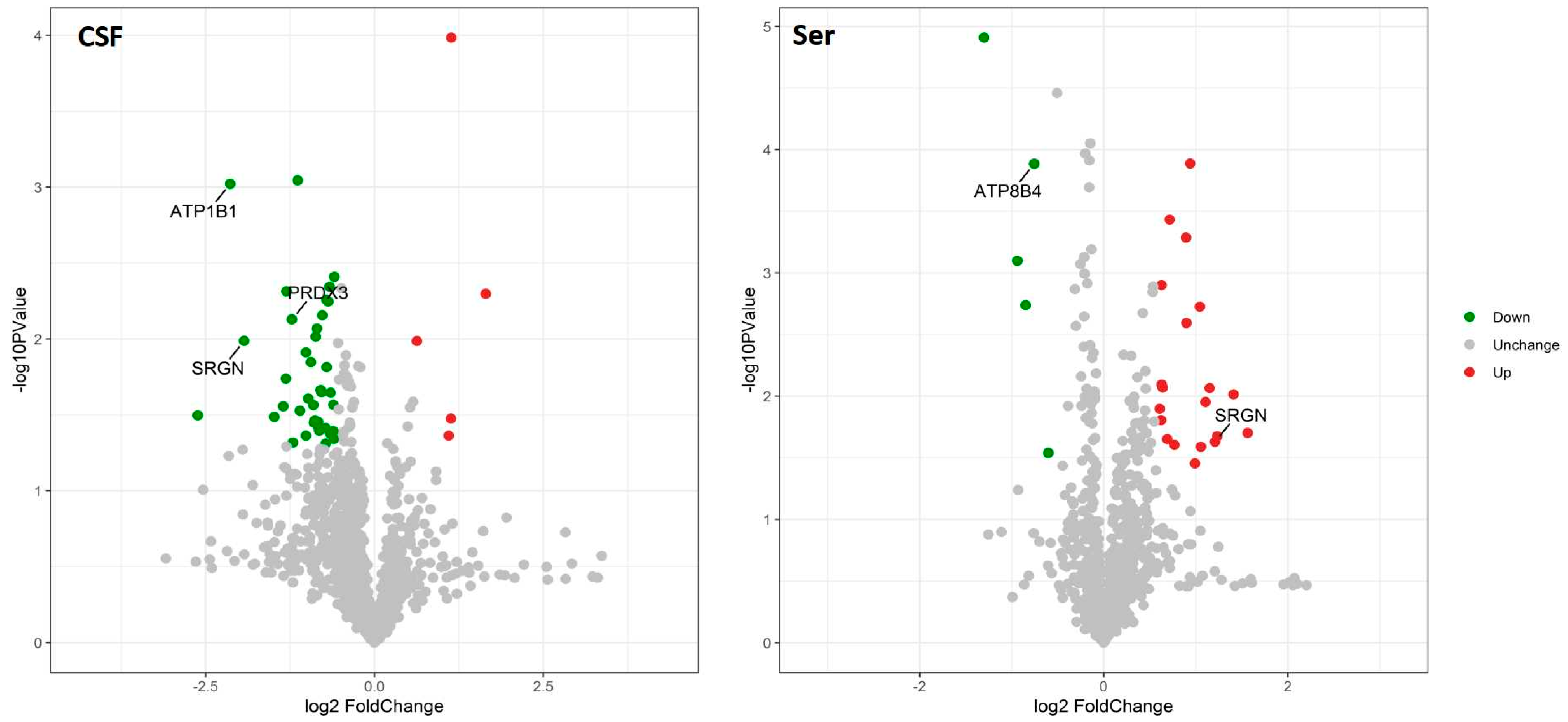
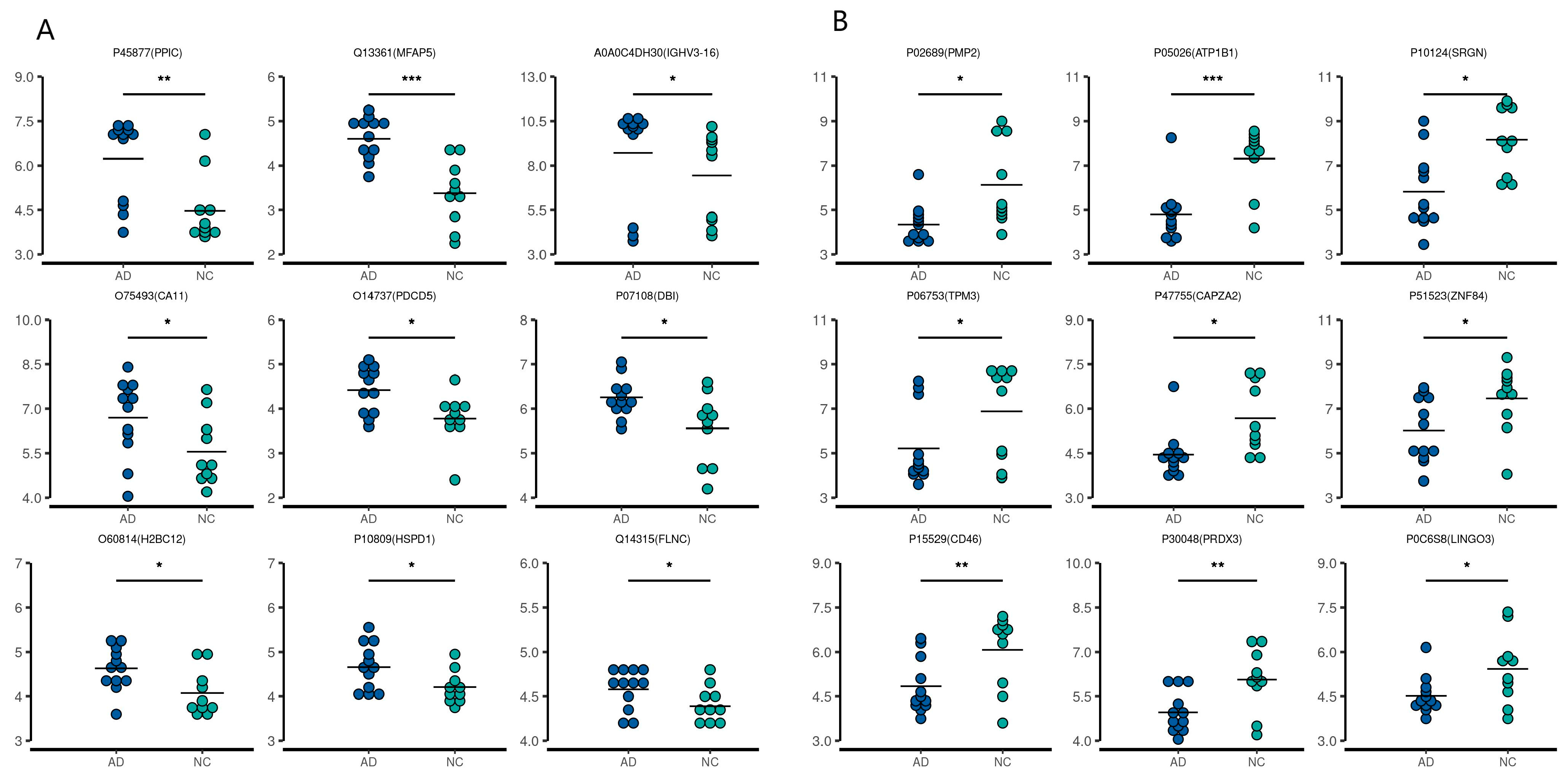
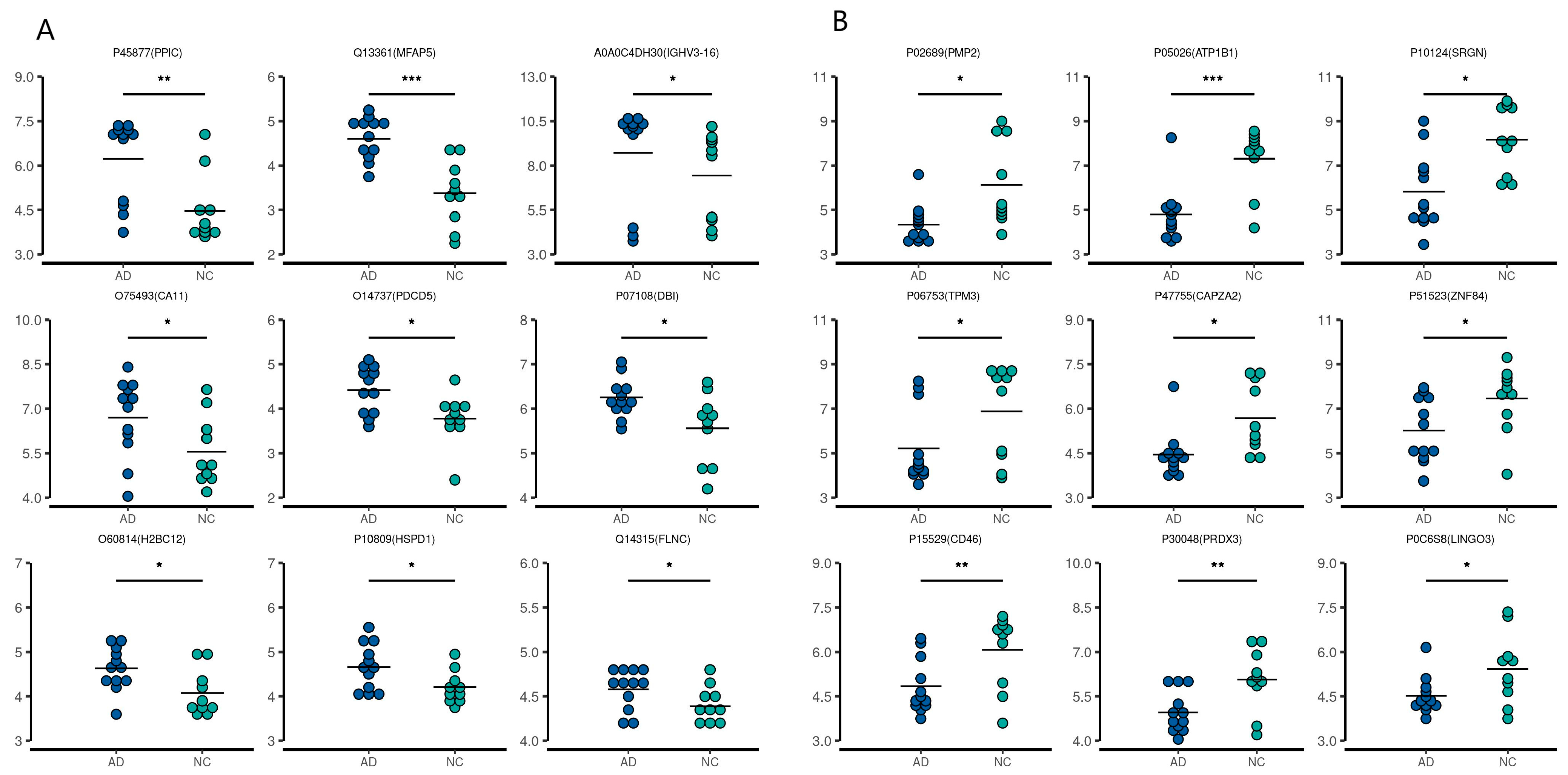
2.2. Quantitative Proteomic Profiling of CSF and Serum Samples from AD and NC Subjects
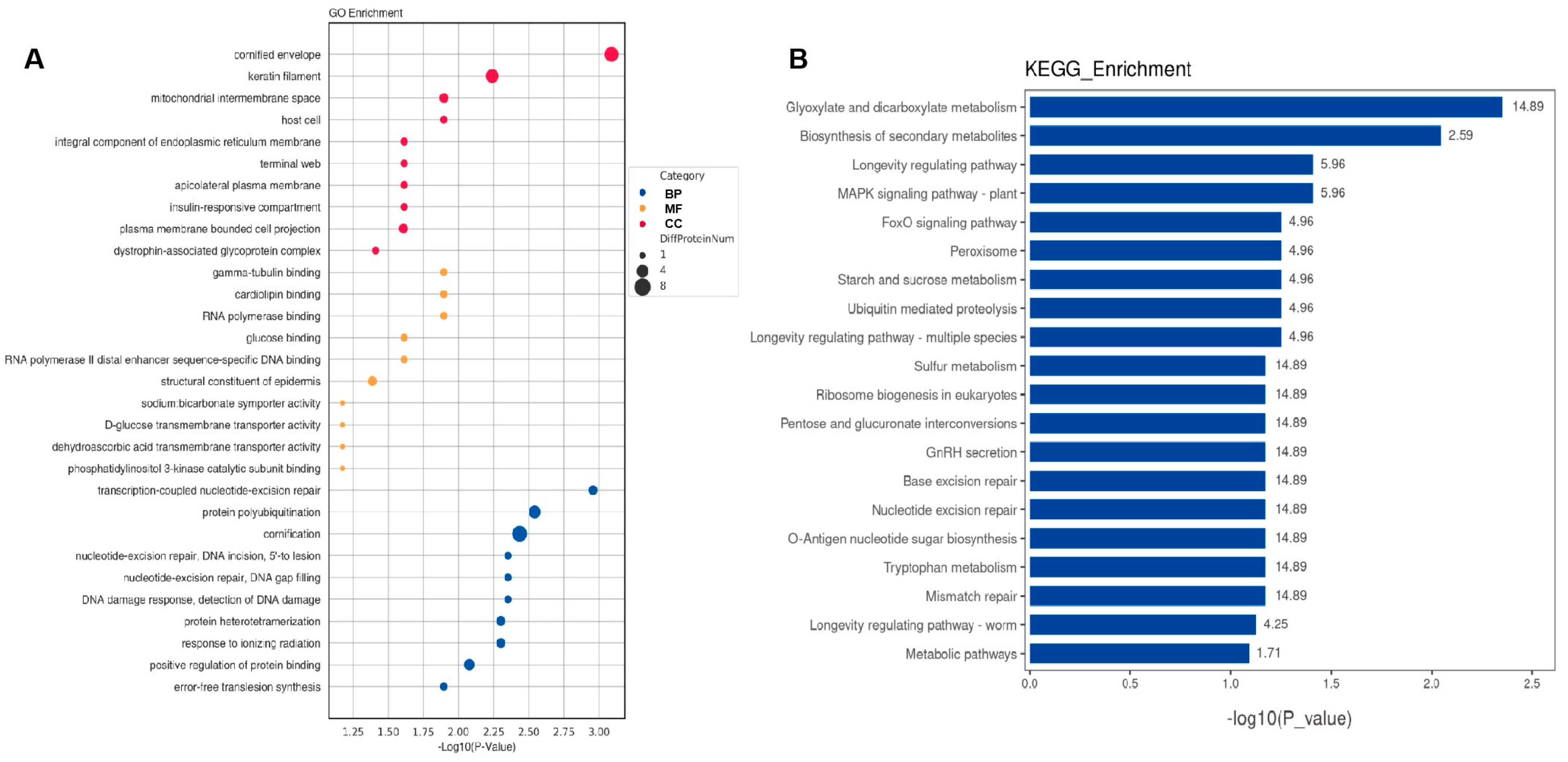
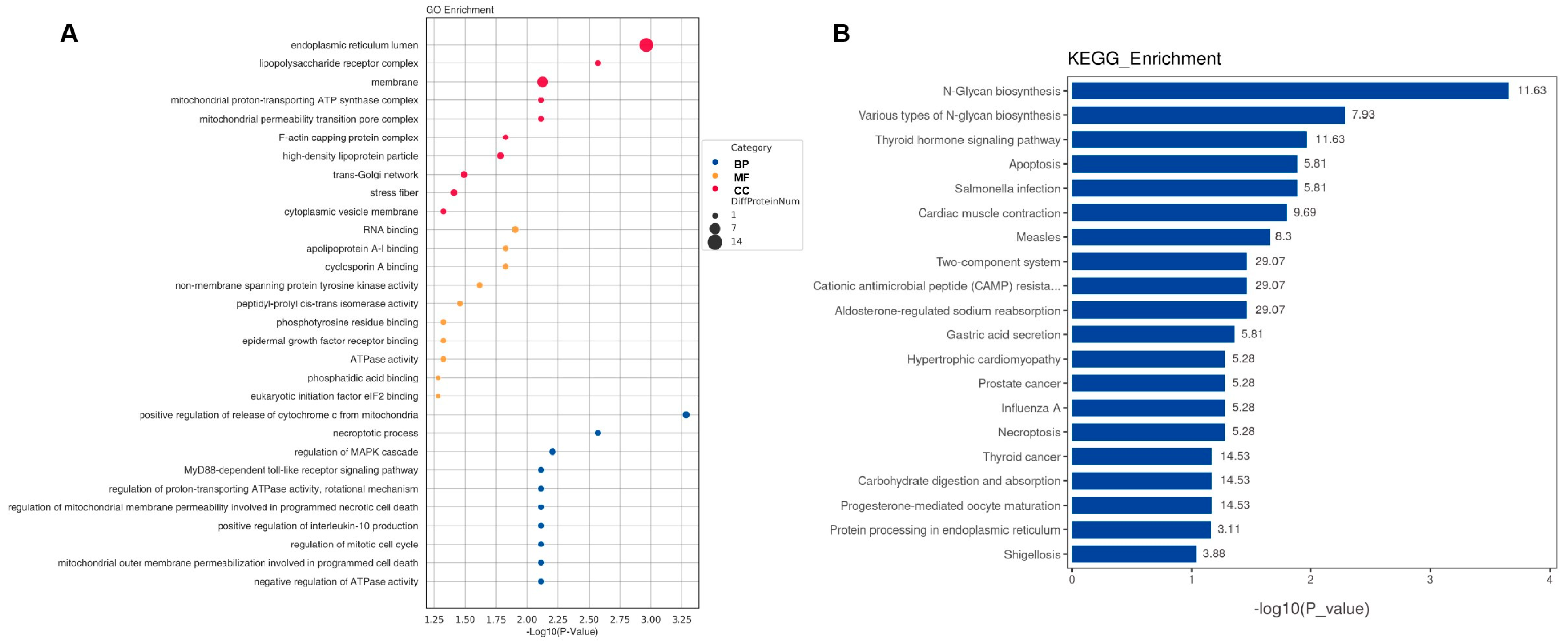
2.3. Bioinformatic Analysis of Specific Proteins Expressed in the AD Group
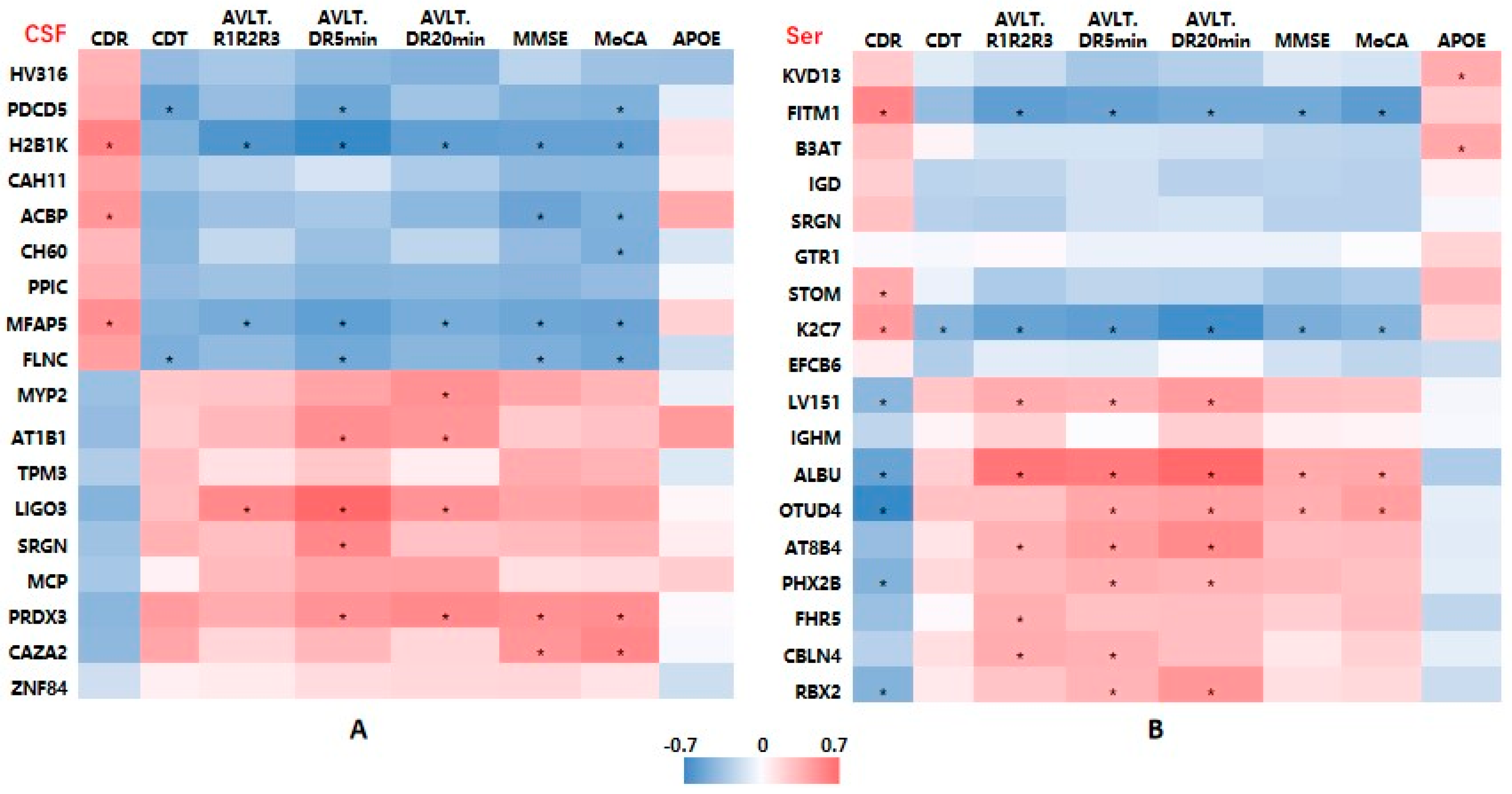
2.4. Associations between DEP Levels and Cognitive Function
2.5. Assessment of the Discriminative Ability of Candidate Biomarkers Using AUC Analysis
3. Discussion
4. Materials and Methods
4.1. Study Subjects
4.2. Clinical Samples
4.3. Mass Spectrometry (MS) Analysis
4.4. Bioinformatic Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jia, J.; Wang, F.; Wei, C.; Zhou, A.; Jia, X.; Li, F.; Tang, M.; Chu, L.; Zhou, Y.; Zhou, C.; et al. The prevalence of dementia in urban and rural areas of China. Alzheimers Dement. 2014, 10, 1–9. [Google Scholar] [CrossRef]
- Jack, C.J.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimers Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef]
- Hampel, H.; Cummings, J.; Blennow, K.; Gao, P.; Jack, C.J.; Vergallo, A. Developing the ATX(N) classification for use across the Alzheimer disease continuum. Nat. Rev. Neurol. 2021, 17, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; Vanderwall, D.; Li, Y.; Wang, X.; Poudel, S.; Wang, H.; Dey, K.K.; Chen, P.C.; Yang, K.; Peng, J. Proteomic landscape of Alzheimer’s Disease: Novel insights into pathogenesis and biomarker discovery. Mol. Neurodegener. 2021, 16, 55. [Google Scholar] [CrossRef]
- Teunissen, C.E.; Verberk, I.; Thijssen, E.H.; Vermunt, L.; Hansson, O.; Zetterberg, H.; van der Flier, W.M.; Mielke, M.M.; Del, C.M. Blood-based biomarkers for Alzheimer’s disease: Towards clinical implementation. Lancet Neurol. 2022, 21, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Dey, K.K.; Chen, P.C.; Li, Y.; Niu, M.; Cho, J.H.; Wang, X.; Bai, B.; Jiao, Y.; Chepyala, S.R.; et al. Integrated analysis of ultra-deep proteomes in cortex, cerebrospinal fluid and serum reveals a mitochondrial signature in Alzheimer’s disease. Mol. Neurodegener. 2020, 15, 43. [Google Scholar] [CrossRef] [PubMed]
- Sathe, G.; Na, C.H.; Renuse, S.; Madugundu, A.K.; Albert, M.; Moghekar, A.; Pandey, A. Quantitative Proteomic Profiling of Cerebrospinal Fluid to Identify Candidate Biomarkers for Alzheimer’s Disease. Proteom. Clin. Appl. 2019, 13, e1800105. [Google Scholar] [CrossRef]
- Bader, J.M.; Geyer, P.E.; Muller, J.B.; Strauss, M.T.; Koch, M.; Leypoldt, F.; Koertvelyessy, P.; Bittner, D.; Schipke, C.G.; Incesoy, E.I.; et al. Proteome profiling in cerebrospinal fluid reveals novel biomarkers of Alzheimer’s disease. Mol. Syst. Biol. 2020, 16, e9356. [Google Scholar] [CrossRef]
- Higginbotham, L.; Ping, L.; Dammer, E.B.; Duong, D.M.; Zhou, M.; Gearing, M.; Hurst, C.; Glass, J.D.; Factor, S.A.; Johnson, E.; et al. Integrated proteomics reveals brain-based cerebrospinal fluid biomarkers in asymptomatic and symptomatic Alzheimer’s disease. Sci. Adv. 2020, 6, eaaz9360. [Google Scholar] [CrossRef]
- Wesenhagen, K.; Teunissen, C.E.; Visser, P.J.; Tijms, B.M. Cerebrospinal fluid proteomics and biological heterogeneity in Alzheimer’s disease: A literature review. Crit. Rev. Clin. Lab. Sci. 2020, 57, 86–98. [Google Scholar] [CrossRef]
- Rebelo, A.P.; Eidhof, I.; Cintra, V.P.; Guillot-Noel, L.; Pereira, C.V.; Timmann, D.; Traschutz, A.; Schols, L.; Coarelli, G.; Durr, A.; et al. Biallelic loss-of-function variations in PRDX3 cause cerebellar ataxia. Brain 2021, 144, 1467–1481. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Na, R.; Ran, Q. Enhanced defense against mitochondrial hydrogen peroxide attenuates age-associated cognition decline. Neurobiol. Aging 2014, 35, 2552–2561. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Gao, C.; Zhang, H.; Huang, X.; Yang, X.; Yang, C.; Liu, W.; Wu, D.; Liu, J. A quantitative proteomic analysis reveals the potential roles of PRDX3 in neurite outgrowth in N2a-APPswe cells. Biochem. Biophys. Res. Commun. 2022, 604, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Kolset, S.O.; Pejler, G. Serglycin: A structural and functional chameleon with wide impact on immune cells. J. Immunol. 2011, 187, 4927–4933. [Google Scholar] [CrossRef]
- van Horssen, J.; Wesseling, P.; van den Heuvel, L.P.; de Waal, R.M.; Verbeek, M.M. Heparan sulphate proteoglycans in Alzheimer’s disease and amyloid-related disorders. Lancet Neurol. 2003, 2, 482–492. [Google Scholar] [CrossRef]
- Lorente-Gea, L.; Garcia, B.; Martin, C.; Quiros, L.M.; Fernandez-Vega, I. Heparan sulfate proteoglycans and heparanases in Alzheimer’s disease: Current outlook and potential therapeutic targets. Neural Regen. Res. 2017, 12, 914–915. [Google Scholar] [CrossRef]
- Lorente-Gea, L.; Garcia, B.; Martin, C.; Ordiales, H.; Garcia-Suarez, O.; Pina-Batista, K.M.; Merayo-Lloves, J.; Quiros, L.M.; Fernandez-Vega, I. Heparan Sulfate Proteoglycans Undergo Differential Expression Alterations in Alzheimer Disease Brains. J. Neuropathol. Exp. Neurol. 2020, 79, 474–483. [Google Scholar] [CrossRef]
- D’Ascola, A.; Scuruchi, M.; Avenoso, A.; Bruschetta, G.; Campo, S.; Mandraffino, G.; Campo, G.M. Serglycin is involved in inflammatory response in articular mouse chondrocytes. Biochem. Biophys. Res. Commun. 2018, 499, 506–512. [Google Scholar] [CrossRef]
- Reine, T.M.; Vuong, T.T.; Jenssen, T.G.; Kolset, S.O. Serglycin secretion is part of the inflammatory response in activated primary human endothelial cells in vitro. Biochim. Biophys. Acta 2014, 1840, 2498–2505. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.D.; Karamanos, N.K. Proteoglycans remodeling in cancer: Underlying molecular mechanisms. Matrix Biol. 2019, 75–76, 220–259. [Google Scholar] [CrossRef]
- Kolseth, I.B.; Reine, T.M.; Vuong, T.T.; Meen, A.J.; Fan, Q.; Jenssen, T.G.; Gronning-Wang, L.M.; Kolset, S.O. Serglycin is part of the secretory repertoire of LPS-activated monocytes. Immun. Inflamm. Dis. 2015, 3, 23–31. [Google Scholar] [CrossRef]
- Wang, L.; Han, Y.; Wang, X. The relationship between plasma serglycin levels and the diagnosis of diabetic retinopathy. J. Clin. Lab. Anal. 2021, 35, e23663. [Google Scholar] [CrossRef]
- Lee, C.S.; Larson, E.B.; Gibbons, L.E.; Lee, A.Y.; McCurry, S.M.; Bowen, J.D.; McCormick, W.C.; Crane, P.K. Associations between recent and established ophthalmic conditions and risk of Alzheimer’s disease. Alzheimers Dement. 2019, 15, 34–41. [Google Scholar] [CrossRef]
- Geering, K. Functional roles of Na,K-ATPase subunits. Curr. Opin. Nephrol. Hypertens 2008, 17, 526–532. [Google Scholar] [CrossRef]
- Sun, J.; Zheng, Y.; Chen, Z.; Wang, Y. The role of Na+-K+-ATPase in the epileptic brain. CNS Neurosci. Ther. 2022, 28, 1294–1302. [Google Scholar] [CrossRef]
- de Lores, A.G.; Ordieres, M.G. Brain Na+, K+-ATPase Activity In Aging and Disease. Int. J. Biomed. Sci. 2014, 10, 85–102. [Google Scholar] [CrossRef]
- Hattori, N.; Kitagawa, K.; Higashida, T.; Yagyu, K.; Shimohama, S.; Wataya, T.; Perry, G.; Smith, M.A.; Inagaki, C. CI−-ATPase and Na+/K+-ATPase activities in Alzheimer’s disease brains. Neurosci. Lett. 1998, 254, 141–144. [Google Scholar] [CrossRef]
- Zhang, L.N.; Sun, Y.J.; Pan, S.; Li, J.X.; Qu, Y.E.; Li, Y.; Wang, Y.L.; Gao, Z.B. Na+-K+-ATPase, a potent neuroprotective modulator against Alzheimer disease. Fundam. Clin. Pharmacol. 2013, 27, 96–103. [Google Scholar] [CrossRef]
- Li, H.; Wetten, S.; Li, L.; St, J.P.; Upmanyu, R.; Surh, L.; Hosford, D.; Barnes, M.R.; Briley, J.D.; Borrie, M.; et al. Candidate single-nucleotide polymorphisms from a genomewide association study of Alzheimer disease. Arch. Neurol. 2008, 65, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Holstege, H.; Hulsman, M.; Charbonnier, C.; Grenier-Boley, B.; Quenez, O.; Grozeva, D.; van Rooij, J.; Sims, R.; Ahmad, S.; Amin, N.; et al. Exome sequencing identifies rare damaging variants in ATP8B4 and ABCA1 as risk factors for Alzheimer’s disease. Nat. Genet. 2022, 54, 1786–1794. [Google Scholar] [CrossRef]
- Dubois, B.; Feldman, H.H.; Jacova, C.; Hampel, H.; Molinuevo, J.L.; Blennow, K.; DeKosky, S.T.; Gauthier, S.; Selkoe, D.; Bateman, R.; et al. Advancing research diagnostic criteria for Alzheimer’s disease: The IWG-2 criteria. Lancet Neurol. 2014, 13, 614–629. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.W.; Liu, H.C.; Wong, P.F. Chinese version and norms of the Mini-Mental State Examination. Biomicrofluidics 1988, 16, e59. [Google Scholar]
- Yu, J.; Li, J.; Huang, X. The Beijing version of the Montreal Cognitive Assessment as a brief screening tool for mild cognitive impairment: A community-based study. BMC Psychiatry 2012, 12, 156. [Google Scholar] [CrossRef]
- Zhao, Q.; Lv, Y.; Zhou, Y.; Hong, Z.; Guo, Q. Short-term delayed recall of auditory verbal learning test is equivalent to long-term delayed recall for identifying amnestic mild cognitive impairment. PLoS ONE 2012, 7, e51157. [Google Scholar] [CrossRef]
- Waring, J.F.; Tang, Q.; Robieson, W.Z.; King, D.P.; Das, U.; Dubow, J.; Dutta, S.; Marek, G.J.; Gault, L.M. APOE-varepsilon4 Carrier Status and Donepezil Response in Patients with Alzheimer’s Disease. J. Alzheimers Dis. 2015, 47, 137–148. [Google Scholar] [CrossRef]
- Vanhoutte, M.; Landeau, B.; Sherif, S.; de la Sayette, V.; Dautricourt, S.; Abbas, A.; Manrique, A.; Chocat, A.; Chetelat, G. Evaluation of the early-phase [(18)F]AV45 PET as an optimal surrogate of [(18)F]FDG PET in ageing and Alzheimer’s clinical syndrome. Neuroimage Clin. 2021, 31, 102750. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AD (n = 30) | NC (n = 30) | t/χ2 | p Value | |
|---|---|---|---|---|
| Age (years) | 65.17 ± 11.34 | 65.63 ± 7.08 | 0.246 | 0.620 |
| Female (%) | 14 (46.67%) | 13 (43.33%) | 0.067 | 0.795 |
| Education (years) | 9.07 ± 4.08 | 8.47 ± 3.15 | 0.346 | 0.557 |
| Diabetes (%) | 9 (30.0%) | 8 (26.67%) | 8.208 | 0.774 |
| Hypertension (%) | 11 (36.67%) | 10 (33.33%) | 7.326 | 0.786 |
| MMSE | 18.17 ± 6.10 | 26.43 ± 1.63 | 35.101 | <0.001 |
| MoCA | 13.93 ± 5.27 | 23.70 ± 2.73 | 38.823 | <0.001 |
| CDT | 19.96 ± 4.56 | 24.78 ± 2.61 | 116.43 | <0.001 |
| CDR | 1.15 ± 0.51 | 0.00 ± 0.00 | - | - |
| CSF Aβ42 (pg/mL) | 452.99 ± 101.25 | 1062.29 ± 101.25 | −5.495 | <0.001 |
| CSF Aβ40 (pg/mL) | 9902.66 ± 2431.47 | 12,280.38 ± 1904.07 | −2.513 | 0.021 |
| CSF Aβ42/Aβ40 | 0.047 ± 0.011 | 0.086 ± 0.019 | −6.029 | <0.001 |
| CSF t-tau (pg/mL) | 413.27 ± 166.92 | 81.27 ± 82.79 | 5.714 | <0.001 |
| CSF p-tau (pg/mL) | 195.1 ± 233.98 | 42.86 ± 16.13 | 2.045 | 0.054 |
| APOE genotype group | 3.314 | 0.191 | ||
| E2 | 7 (23.3%) | 13 (43.3%) | ||
| E3 | 12 (40.0%) | 11 (36.7%) | ||
| E4 | 11 (36.7%) | 6 (20.0%) |
| Accession Number | Protein Name | Protein | Gene Name | Change | p Value |
|---|---|---|---|---|---|
| A0A0C4DH30 | Probable non-functional immunoglobulin heavy variable 3–16 | HV316 | IGHV3-16 | ↑ | 0.000546667 |
| O14737 | Programmed cell death protein 5 | PDCD5 | PDCD5 | ↑ | 0.005184637 |
| O60814 | Histone H2B type 1-K | H2B1K | H2BC12 | ↑ | 0.007706908 |
| O75493 | Carbonic anhydrase-related protein 11 | CAH11 | CA11 | ↑ | 8.88922 × 10−26 |
| P07108 | Acyl-CoA-binding protein | ACBP | DBI | ↑ | 3.75501 × 10−59 |
| P10809 | 60 kDa heat shock protein, mitochondrial | CH60 | HSPD1 | ↑ | 1.4658 × 10−25 |
| P45877 | Peptidyl-prolyl cis-trans isomerase C | PPIC | PPIC | ↑ | 0.003508082 |
| Q13361 | Microfibrillar-associated protein 5 | MFAP5 | MFAP5 | ↑ | 5.04183 × 10−8 |
| Q14315 | Filamin-C | FLNC | FLNC | ↑ | 0.000943945 |
| P02689 | Myelin P2 protein | MYP2 | PMP2 | ↓ | 5.57 × 10−24 |
| P05026 | Sodium/potassium-transporting ATPase subunit beta-1 | AT1B1 | ATP1B1 | ↓ | 0.000577 |
| P06753 | Tropomyosin alpha-3 chain; | TPM3 | TPM3 | ↓ | 1.02 × 10−16 |
| P0C6S8 | Leucine-rich repeat and immunoglobulin-like domain-containing nogo receptor-interacting protein 3 | LIGO3 | LINGO3 | ↓ | 0.002143 |
| P10124 | Serglycin | SRGN | SRGN | ↓ | 1.11 × 10−48 |
| P15529 | Membrane cofactor protein | MCP | CD46 | ↓ | 1.5 × 10−26 |
| P30048 | Thioredoxin-dependent peroxide reductase, mitochondrial | PRDX3 | PRDX3 | ↓ | 0.000903 |
| P47755 | F-actin-capping protein subunit alpha-2 | CAZA2 | CAPZA2 | ↓ | 7.82 × 10−11 |
| P51523 | Zinc finger protein 84 | ZNF84 | ZNF84 | ↓ | 4.79 × 10−20 |
| Accession Number | Protein Name | Protein | Gene Name | Change | p Value |
|---|---|---|---|---|---|
| A0A0B4J2D9 | Immunoglobulin kappa variable 1D-13 | KVD13 | IGKV1D-13 | ↑ | 4.98 × 10−21 |
| A5D6W6 | Fat storage-inducing transmembrane protein 1 | FITM1 | FITM1 | ↑ | 1.91 × 10−89 |
| P02730 | Band 3 anion transport protein | B3AT | SLC4A1 | ↑ | 3.33 × 10−75 |
| P0DOX3 | Immunoglobulin delta heavy chain | IGD | ↑ | 3.06 × 10−55 | |
| P10124 | Serglycin | SRGN | SRGN | ↑ | 1.95 × 10−49 |
| P11166 | Solute carrier family 2, facilitated glucose transporter member 1 | GTR1 | SLC2A1 | ↑ | 0.00029 |
| P27105 | Stomatin | STOM | STOM | ↑ | 4.1 × 10−101 |
| Q14CN4 | Keratin, type II cytoskeletal 72 | K2C72 | KRT72 | ↑ | 0.00112 |
| Q5THR3 | EF-hand calcium-binding domain-containing protein 6 | EFCB6 | EFCAB6 | ↑ | 0.000927 |
| P01701 | Immunoglobulin lambda variable 1–51 | LV151 | IGLV1-51 | ↓ | 2.12 × 10−58 |
| P01871 | Immunoglobulin heavy constant mu | IGHM | IGHM | ↓ | 1.8 × 10−107 |
| P02768 | Albumin | ALBU | ALB | ↓ | 1.1 × 10−124 |
| Q01804 | OTU domain-containing protein 4 | OTUD4 | OTUD4 | ↓ | 8.57 × 10−5 |
| Q8TF62 | Probable phospholipid-transporting ATPase IM | AT8B4 | ATP8B4 | ↓ | 3.41 × 10−25 |
| Q99453 | Paired mesoderm homeobox protein 2B | PHX2B | PHOX2B | ↓ | 1.17 × 10−57 |
| Q9BXR6 | Complement factor H-related protein 5 | FHR5 | CFHR5 | ↓ | 1.5 × 10−105 |
| Q9NTU7 | Cerebellin-4 | CBLN4 | CBLN4 | ↓ | 3.24 × 10−26 |
| Q9UBF6 | RING-box protein 2 | RBX2 | RNF7 | ↓ | 0.000191 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, P.; Li, L.; He, F.; Meng, F.; Liu, X.; Su, Y.; Su, X.; Luo, B.; Peng, G. Identification of Candidate Biomarkers of Alzheimer’s Disease via Multiplex Cerebrospinal Fluid and Serum Proteomics. Int. J. Mol. Sci. 2023, 24, 14225. https://doi.org/10.3390/ijms241814225
Liu P, Li L, He F, Meng F, Liu X, Su Y, Su X, Luo B, Peng G. Identification of Candidate Biomarkers of Alzheimer’s Disease via Multiplex Cerebrospinal Fluid and Serum Proteomics. International Journal of Molecular Sciences. 2023; 24(18):14225. https://doi.org/10.3390/ijms241814225
Chicago/Turabian StyleLiu, Ping, Lingxiao Li, Fangping He, Fanxia Meng, Xiaoyan Liu, Yujie Su, Xinhui Su, Benyan Luo, and Guoping Peng. 2023. "Identification of Candidate Biomarkers of Alzheimer’s Disease via Multiplex Cerebrospinal Fluid and Serum Proteomics" International Journal of Molecular Sciences 24, no. 18: 14225. https://doi.org/10.3390/ijms241814225
APA StyleLiu, P., Li, L., He, F., Meng, F., Liu, X., Su, Y., Su, X., Luo, B., & Peng, G. (2023). Identification of Candidate Biomarkers of Alzheimer’s Disease via Multiplex Cerebrospinal Fluid and Serum Proteomics. International Journal of Molecular Sciences, 24(18), 14225. https://doi.org/10.3390/ijms241814225