
Integrating the Soil Microbiota and Metabolome Reveals the Mechanism through Which Controlled Release Fertilizer Affects Sugarcane Growth


Abstract
1. Introduction
2. Results
2.1. Response of Agronomic Traits of Sugarcane to Different CRF Application Rates
2.2. Response of Sugarcane Rhizosphere Soil Nutrients to Different Fertilizer Applications
2.3. Composition of Sugarcane Rhizosphere Microbial Communities and Metabolites
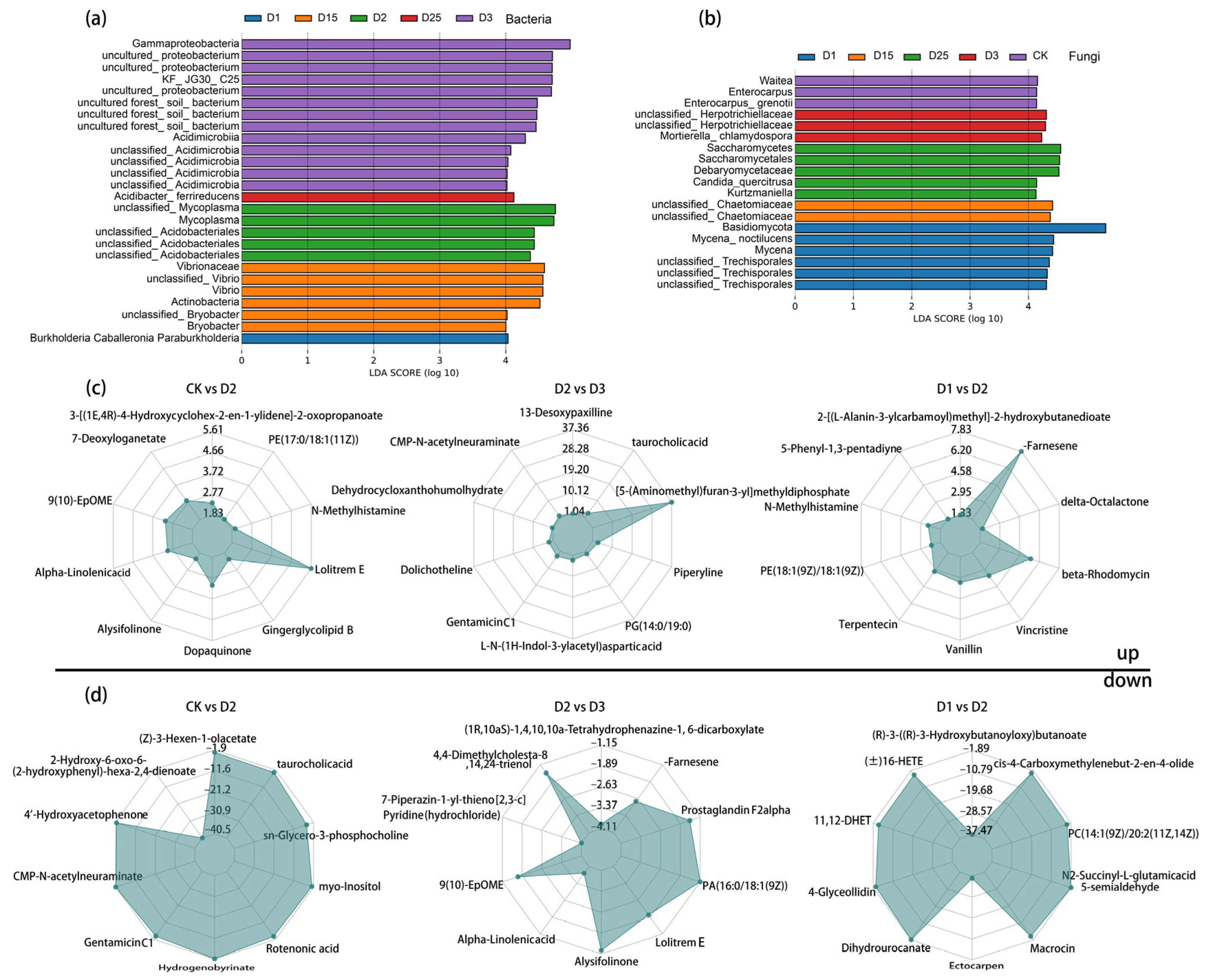
2.4. Screening for Differential Microorganisms and Metabolites in Sugarcane Rhizosphere Soil
2.5. Soil Nutrients Drive the Formation of Rhizosphere Microbial Communities and Metabolite Assemblages
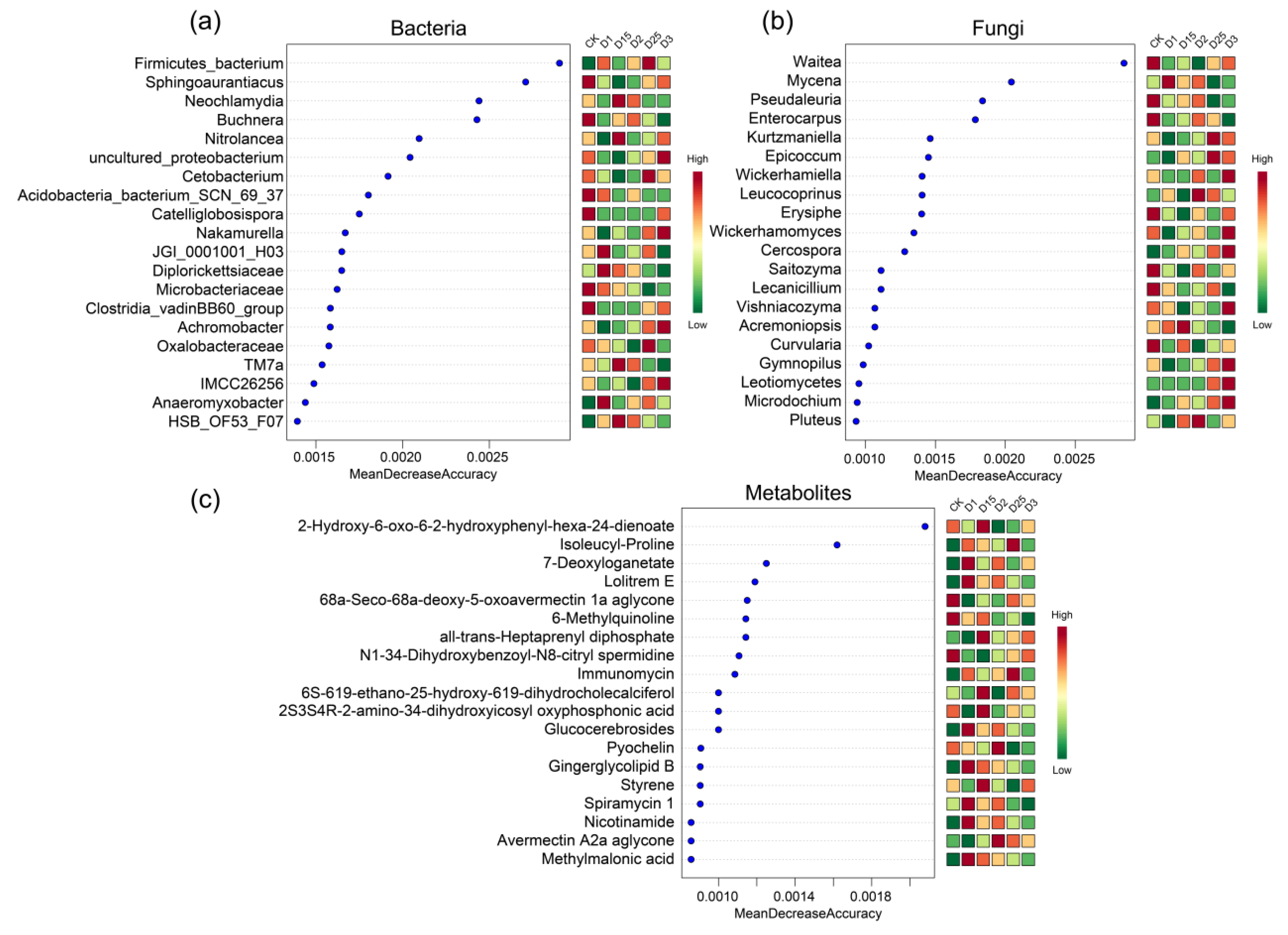
2.6. Random Forest Screening for Characteristic Microorganisms and Metabolites under Different Fertilizer Application Conditions
2.7. Association of Dominant Microorganisms and Metabolites in Sugarcane Rhizosphere Soils
3. Discussion
3.1. Response of Sugarcane Phenotypic Traits and Rhizosphere Conventional Soil Nutrients to CRF Applications
3.2. Response of Sugarcane Rhizosphere Microorganisms to Fertilizer Differences
3.3. Response of Characterized Microorganisms and Metabolites Screened by Random Forests to CRF Application
3.4. Dominant Microorganisms and Metabolite Associations under CRF Application Conditions
4. Materials and Methods
4.1. Experimental Site and Materials
4.2. Experimental Design and Sample Collection
4.3. Measurement of Phenotypic Traits and Rhizosphere Soil Physico-Chemical Indicators in Sugarcane
4.4. Soil Total DNA Extraction and High-Throughput Sequencing
4.5. Quality Control and Filtering of Microbiological Data
4.6. Non-Target Metabolomics Analysis
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, R.D.; Zhang, Y.B.; Liu, S.C.; Guo, J.W.; Wang, Y.Y. Present and Prospect of Sugarcane Production Technology in the World. Sugar Crops China 2009, 3, 54–56+64. [Google Scholar]
- Li, Y.R. Development of Sugar Industry in China. In Proceedings of the International Symposium on Sustainable Sugarcane and Sugar Production Technology, Nanning, China, 29 November–2 December 2004. [Google Scholar]
- Lou, T.; Long, H.Y.; Yang, L.J.; Chen, B.H.; Zhou, S.L.; Zhen, M.U. The effect of fertilizer ratio and rate on soil quality and crop yields in the farmland of excessive use of nitrogenous fertilizers. Soil Fertil. Sci. China 2010, 2, 11–15+34. [Google Scholar]
- Kannika, S.; Kanittha, B.; Thanakorn, W.; Pennapa, M.; Wiyong, K. A tunable controlled-release urea fertilizer coated with a biodegradable polyurethane-nanoclay composite layer. J. Coat. Technol. Res. 2023, 20, 635–646. [Google Scholar]
- Li, J. Effects of Different Slow Controlled Release Fertilizers on Yield of Rice Xiushui 134 and Its Benefits. Grain Sci. Technol. Econ. 2018, 43, 99–102. [Google Scholar]
- Fred, H. The benefits of applying controlled release fertilizers: CRFs offer growers another way to increase efficiency and profitability while promoting sustainability. Greenh. Manag. 2011, 31, 66–68. [Google Scholar]
- Jain, D.S.K.; Agrawal, G.P.; Kumar, M.M.; Anande, M.N. Controlled Release Fertilizers: Trends and Technologies. Pharm. Rev. 2007, 5. Available online: www.pharmainfo.net (accessed on 2 August 2023).
- Zhao, X.; Zhang, B.; Liu, S.; Yang, X. Evaluation of efficiency of controlled-release N fertiliser on tartary buckwheat production. Plant Soil Environ. 2021, 67, 399–407. [Google Scholar] [CrossRef]
- Souza, A.D.G.; Smiderle, O.J.; Montenegro, R.A.; Moriyama, T.K.; Dias, T.J. Controlled-Release Fertiliser and Substrates on Seedling Growth and Quality in Agonandra brasiliensis in Roraima. J. Agric. Stud. 2020, 8, 70. [Google Scholar] [CrossRef][Green Version]
- Wang, N.N.; Xiong, H.R.; Xiong, Y.F.; Li, X.; Wen, Z.Y.; Yuan, X.H. Slowly/controlled release fertilizers and its application in rice. Chem. Eng. 2013, 27, 30–34. [Google Scholar]
- Chen, L.; Schwier, M.; Krumbach, J.; Kopriva, S.; Jacoby, R.P. Chapter Five—Metabolomics in plant-microbe interactions in the roots. In Advances in Botanical Research; Pétriacq, P., Bouchereau, A., Eds.; Academic Press: Cambridge, MA, USA, 2021; Volume 98, pp. 133–161. [Google Scholar]
- Egamberdieva, D. Microbially Derived Phytohormones in Plant Adaptation Against Abiotic Stress; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016. [Google Scholar]
- Asghari, B.; Mussarat, F. Salt tolerance in Zea mays (L). following inoculation with Rhizobium and Pseudomonas. Biol. Fertil. Soils: Coop. J. Int. Soc. Soil Sci. 2009, 45, 405–413. [Google Scholar]
- Liu, H.; Brettell, L.E.; Qiu, Z.; Singh, B.K. Microbiome-Mediated Stress Resistance in Plants. Trends Plant Sci. 2020, 25, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Saeed, Q.; Xiukang, W.; Haider, F.U.; Kučerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M.; et al. Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms. Int. J. Mol. Sci. 2021, 22, 10529. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Mao, H.; Makinia, J.; Drewnowski, J.; Wu, B.; Xu, J.; Xie, L.; Lu, X. Impact of soluble organic matter and particulate organic matter on anammox system: Performance, microbial community and N2O production. J. Environ. Sci. 2023, 124, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, F.S.; Flora, L.V.D.; Henrique, I.G.; Silva, D.C.D.; Mercedes, A.P.; Silva, A.C.; Oliveira, A.S.D.; Silva, M.P.B.D.; Ronning, B.P.F.; Dreher, D.R. Arbuscular Mycorrhizal Fungi (Rhizophagus clarus) and Rhizobacteria (Bacillus subtilis) Can Improve the Clonal Propagation and Development of Teak for Commercial Plantings. Front. Plant Sci. 2021, 12, 1008. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, D.J.; Delgado-Baquerizo, M.; Travers, S.K.; Val, J.; Oliver, I.; Hamonts, K.; Singh, B.K. Competition drives the response of soil microbial diversity to increased grazing by vertebrate herbivores. Ecology 2017, 98, 1922–1931. [Google Scholar] [CrossRef] [PubMed]
- Shastri, B.; Kumar, R. Chapter 6—Microbial secondary metabolites and plant–microbe communications in the rhizosphere. In New and Future Developments in Microbial Biotechnology and Bioengineering; Singh, J.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 93–111. [Google Scholar]
- Sharma, A.; Sharma, S.; Kumar, A.; Kumar, V.; Sharma, A.K. Plant Secondary Metabolites: An Introduction of Their Chemistry and Biological Significance with Physicochemical Aspect. In Plant Secondary Metabolites: Physico-Chemical Properties and Therapeutic Applications; Sharma, A.K., Sharma, A., Eds.; Springer Nature Singapore: Singapore, 2022; pp. 1–45. [Google Scholar]
- Anjali; Kumar, S.; Korra, T.; Thakur, R.; Arutselvan, R.; Kashyap, A.S.; Nehela, Y.; Chaplygin, V.; Minkina, T.; Keswani, C. Role of plant secondary metabolites in defence and transcriptional regulation in response to biotic stress. Plant Stress 2023, 8, 100154. [Google Scholar] [CrossRef]
- Rao, M.J.; Duan, M.; Yang, M.; Li, M.; Wang, L. Sugarcane Rind Secondary Metabolites and Their Antioxidant Activities in Eleven Cultivated Sugarcane Varieties. Sugar Tech. 2022, 24, 1570–1582. [Google Scholar] [CrossRef]
- Liu, Y.; Li, D.; Gao, H.; Li, Y.; Chen, W.; Jiao, S.; Wei, G. Regulation of soil micro-foodwebs to root secondary metabolites in cultivated and wild licorice plants. Sci. Total Environ. 2022, 828, 154302. [Google Scholar] [CrossRef]
- Sun, Y.; Duan, C.; Cao, N.; Ding, C.; Huang, Y.; Wang, J. Biodegradable and conventional microplastics exhibit distinct microbiome, functionality, and metabolome changes in soil. J. Hazard. Mater. 2022, 424, 127282. [Google Scholar] [CrossRef]
- Huang, W.; Sun, D.; Chen, L.; An, Y. Integrative analysis of the microbiome and metabolome in understanding the causes of sugarcane bitterness. Sci. Rep. 2021, 11, 6024. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, D.; Yuan, X.; Xu, Y.; Chen, C.; Zhao, L. Soil microbiome and metabolome analysis reveals beneficial effects of ginseng–celandine rotation on the rhizosphere soil of ginseng-used fields. Rhizosphere 2022, 23, 100559. [Google Scholar] [CrossRef]
- Cock, J.H.; Luna, C.A.; Palma, A. The trade-off between total harvestable production and concentration of the economically useful yield component: Cane tonnage and sugar content. Field Crops Res. 2000, 67, 257–262. [Google Scholar] [CrossRef]
- Maharjan, G.R.; Prescher, A.K.; Nendel, C.; Ewert, F.; Mboh, C.M.; Gaiser, T.; Seidel, S.J. Approaches to model the impact of tillage implements on soil physical and nutrient properties in different agro-ecosystem models. Soil Tillage Res. 2018, 180, 210–221. [Google Scholar] [CrossRef]
- Farouque, G.; Takeya, H. Farmers’ Perception of Integrated Soil Fertility and Nutrient Management for Sustainable Crop Production: A Study of Rural Areas in Bangladesh. J. Agric. Educ. 2007, 48, 111–122. [Google Scholar] [CrossRef]
- Field, D.J. Identifying the Interrelationship between Soil Properties that Influence the Function of the Soil; Regional Institute Ltd.: New South Wales, Australia, 2004. [Google Scholar]
- Neina, D.; Trevisan, M. The Role of Soil pH in Plant Nutrition and Soil Remediation. Appl. Environ. Soil Sci. 2019, 2019, 5794869. [Google Scholar] [CrossRef]
- Xing, Y.; Jiang, Z.P.; Tan, Y.M.; Liao, Q.; Meng, Y.C.; Yang, S.E.; Chen, G.F.; Liang, P.X.; Huang, T.Q.; Pan, L.P. Effects of silicon-calcium-phosphorus fertilizer on sugarcane growth and soil fertility of acidified filed. J. South. Agric. 2016, 47, 1495–1499. [Google Scholar]
- Jariwala, H.; Santos, R.M.; Lauzon, J.D.; Dutta, A.; Wai Chiang, Y. Controlled release fertilizers (CRFs) for climate-smart agriculture practices: A comprehensive review on release mechanism, materials, methods of preparation, and effect on environmental parameters. Environ. Sci. Pollut. Res. 2022, 29, 53967–53995. [Google Scholar] [CrossRef] [PubMed]
- Vejan, P.; Khadiran, T.; Abdullah, R.; Ahmad, N. Controlled release fertilizer: A review on developments, applications and potential in agriculture. J. Control. Release 2021, 339, 321–334. [Google Scholar] [CrossRef]
- Yang, J.I.; Haiyang, Y.U.; Ralf, C.; Hua, X.U. Effect of the Coupling Technique of Intermittent irrigation and Controlledrelease Fertilizer on the Abundance of Soil Microbial Communities from Rice-wheat Rotation Ecosystem. Ecol. Environ. Sci. 2017, 26, 1696–1703. [Google Scholar]
- Niu, T.; Xie, J.; Li, J.; Zhang, J.; Zhang, X.; Ma, H.; Wang, C. Response of rhizosphere microbial community of Chinese chives under different fertilization treatments. Front. Microbiol. 2022, 13, 1031624. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, J.; Mi, W.; Mu, W.; Tao, W.; Chen, C.; Mi, W.; Su, L. Effects of slow- and controlled-release nitrogen fertilizers on bacterial and fungal community composition in a double-cropping rice system. Soil Tillage Res. 2023, 230, 105714. [Google Scholar] [CrossRef]
- Romero-Olivares, A.L.; Morrison, E.W.; Pringle, A.; Frey, S.D. Linking Genes to Traits in Fungi. Microb. Ecol. 2021, 82, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Finestone, J.; Templer, P.H.; Bhatnagar, J.M. Soil Fungi Exposed to Warming Temperatures and Shrinking Snowpack in a Northern Hardwood Forest Have Lower Capacity for Growth and Nutrient Cycling. Front. For. Glob. Chang. 2022, 5. [Google Scholar] [CrossRef]
- Khan, A.; Jiang, H.; Bu, J.; Adnan, M.; Gillani, S.W.; Hussain, M.A.; Zhang, M. Untangling the Rhizosphere Bacterial Community Composition and Response of Soil Physiochemical Properties to Different Nitrogen Applications in Sugarcane Field. Front. Microbiol. 2022, 13, 856078. [Google Scholar] [CrossRef] [PubMed]
- Rajan, M.; Shahena, S.; Chandran, V.; Mathew, L. Chapter 3—Controlled release of fertilizers—Concept, reality, and mechanism. In Controlled Release Fertilizers for Sustainable Agriculture; Lewu, F.B., Volova, T., Thomas, S., Rakhimol, R.K., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 41–56. [Google Scholar]
- Kielak, A.M.; Barreto, C.C.; Kowalchuk, G.A.; van Veen, J.A.; Kuramae, E.E. The Ecology of Acidobacteria: Moving beyond Genes and Genomes. Front. Microbiol. 2016, 7, 744. [Google Scholar] [CrossRef]
- Bender, S.F.; Conen, F.; Van der Heijden, M.G.A. Mycorrhizal effects on nutrient cycling, nutrient leaching and N2O production in experimental grassland. Soil Biol. Biochem. 2015, 80, 283–292. [Google Scholar] [CrossRef]
- Guasch, H.; Bonet, B.; Bonnineau, C.; Barral, L. Microbial Biomarkers. In Microbial Ecotoxicology; Cravo-Laureau, C., Cagnon, C., Lauga, B., Duran, R., Eds.; Springer International Publishing: Cham, Switerland, 2017; pp. 251–281. [Google Scholar]
- Wang, S.; Xiong, W.; Wang, Y.; Nie, Y.; Wu, Q.; Xu, Y.; Geisen, S. Temperature-Induced Annual Variation in Microbial Community Changes and Resulting Metabolome Shifts in a Controlled Fermentation System. mSystems 2020, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Jia, R.; Li, L.; Jiang, R.; Qu, D. Community structure of Anaeromyxobacter in Fe(III) reducing enriched cultures of paddy soils. J. Soils Sediments 2020, 20, 1621–1631. [Google Scholar] [CrossRef]
- Masuda, Y.; Yamanaka, H.; Xu, Z.X.; Shiratori, Y.; Aono, T.; Amachi, S.; Senoo, K.; Itoh, H. Diazotrophic Anaeromyxobacter Isolates from Soils. Appl. Environ. Microbiol. 2020, 86, e00956-20. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Zhang, Z.; Ding, Q.; Yang, Y.; Bindelle, J.; Ran, C.; Zhou, Z. Intestinal Cetobacterium and acetate modify glucose homeostasis via parasympathetic activation in zebrafish. Gut Microbes 2021, 13, 1–15. [Google Scholar] [CrossRef]
- Korenblum, E.; Dong, Y.; Szymanski, J.; Panda, S.; Jozwiak, A.; Massalha, H.; Meir, S.; Rogachev, I.; Aharoni, A. Rhizosphere microbiome mediates systemic root metabolite exudation by root-to-root signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 3874–3883. [Google Scholar] [CrossRef] [PubMed]
- Jain, J.D.S. Underlying mechanism of plant-microbe crosstalk in shaping microbial ecology of the rhizosphere. Acta Physiol. Plant 2020, 42, 1–13. [Google Scholar] [CrossRef]
- Shavit, U.; Shaviv, A.; Shalit, G.; Zaslavsky, D. Release characteristics of a new controlled release fertilizer. J. Control. Release 1997, 43, 131–138. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, Z. 3 Pezizomycotina: Sordariomycetes and Leotiomycetes. In Systematics and Evolution: Part B.; McLaughlin, D.J., Spatafora, J.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 57–88. [Google Scholar]
- Vaghefi, N.; Shivas, R.G.; Sharma, S.; Nelson, S.C.; Pethybridge, S.J. Phylogeny of cercosporoid fungi (Mycosphaerellaceae, Mycosphaerellales) from Hawaii and New York reveals novel species within the Cercospora beticola complex. Mycol. Prog. 2021, 20, 261–287. [Google Scholar] [CrossRef]
- Palma, C.F.F.; Castro-Alves, V.; Morales, L.O.; Rosenqvist, E.; Ottosen, C.; Hyötyläinen, T.; Strid, Å. Metabolic changes in cucumber leaves are enhanced by blue light but differentially affected by UV interactions with light signalling pathways in the visible spectrum. Plant Sci. 2022, 321, 111326. [Google Scholar] [CrossRef] [PubMed]
- Pathak, R.K.; Singh, D.B.; Sharma, H.; Pandey, D.; Dwivedi, S. Chapter 21—Calcium uptake and translocation in plants. In Calcium Transport Elements in Plants; Upadhyay, S.K., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 373–386. [Google Scholar]
- Matsumoto, H.; Fan, X.; Wang, Y.; Kusstatscher, P.; Duan, J.; Wu, S.; Chen, S.; Qiao, K.; Wang, Y.; Ma, B.; et al. Bacterial seed endophyte shapes disease resistance in rice. Nat. Plants 2021, 7, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Zhang, S.; Wang, X.; Xu, X.; Ai, C.; Liang, G.; Zhu, P.; Zhou, W. Enzymatic stoichiometry reveals phosphorus limitation-induced changes in the soil bacterial communities and element cycling: Evidence from a long-term field experiment. Geoderma Int. J. Soil Sci. 2022, 426, 116124. [Google Scholar] [CrossRef]
- Cui, Y.; Chun, S.J.; Ko, S.R.; Lee, H.G.; Srivastava, A.; Oh, H.M.; Ahn, C.Y. Reyranella aquatilis sp. nov., an alphaproteobacterium isolated from a eutrophic lake. Int. J. Syst. Evol. Microbiol. 2017, 67, 3496–3500. [Google Scholar] [CrossRef]
- Primieri, S.S.J.C. Nodule-associated bacteria alter the mutualism between arbuscular mycorrhizal fungi and N-2 fixing bacteria. Soil Biol. Biochem. 2021, 154, 108149. [Google Scholar] [CrossRef]
- Kang, L.; Ji, C.Y.; Kim, S.H.; Ke, Q.; Park, S.; Kim, H.S.; Lee, H.; Lee, J.S.; Park, W.S.; Ahn, M.; et al. Suppression of the β-carotene hydroxylase gene increases β-carotene content and tolerance to abiotic stress in transgenic sweetpotato plants. Plant Physiol. Biochem. 2017, 117, 24–33. [Google Scholar] [CrossRef]
- Saghai, A.; Wittorf, L.; Philippot, L.; Hallin, S. Loss in soil microbial diversity constrains microbiome selection and alters the abundance of N-cycling guilds in barley rhizosphere. Appl. Soil Ecol. 2022, 169, 104224. [Google Scholar] [CrossRef]
- Gao, S.; Yang, Y.; Wang, C.; Guo, J.; Zhou, D.; Wu, Q.; Su, Y.; Xu, L.; Que, Y. Transgenic Sugarcane with a cry1Ac Gene Exhibited Better Phenotypic Traits and Enhanced Resistance against Sugarcane Borer. PLoS ONE 2016, 11, e153929. [Google Scholar] [CrossRef]
- Bao, S. Soil Agrochemical Analysis, 3rd ed.; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Wang, K.; Zhang, Y.; Tang, Z.; Shangguan, Z.; Chang, F.; Jia, F.; Chen, Y.; He, X.; Shi, W.; Deng, L. Effects of grassland afforestation on structure and function of soil bacterial and fungal communities. Sci. Total Environ. 2019, 676, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, J41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; David, B.; Paul, B.; Eva, Z.; Sue, F.M.; Nadine, A.; Marie, B.; Knowles, J.D.; Antony, H.; Haselden, J.N. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2018, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, T.; Shen, X.; Liu, J.; Zhao, D.; Sun, Y.; Wang, L.; Liu, Y.; Gong, X.; Liu, Y. Serum metabolomics for early diagnosis of esophageal squamous cell carcinoma by UHPLC-QTOF/MS. Metabolomics 2016, 12, 116. [Google Scholar] [CrossRef]
- Wishart, D.S.; Djoumbou, F.Y.; Ana, M.; Chi, G.A.; Kevin, L.; Rosa, V.; Tanvir, S.; Daniel, J.; Carin, L.; Naama, K. HMDB 4.0: The human metabolome database for 2018. Nucleic. Acids. Res. 2017, 46, D608–D617. [Google Scholar] [CrossRef]
- Hiroyuki, O.; Susumu, G.; Kazushige, S.; Wataru, F.; Hidemasa, B.; Minoru, K. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef]
- Eoin, F.; Manish, S.; Dawn, C.; Shankar, S. LIPID MAPS online tools for lipid research. Nucleic. Acids. Res. 2007, 35, W606–W612. [Google Scholar]
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the Human Adult Urinary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height | Stem Diameter | Sucrose Content | Stem Weight | Effective Stems | Cane Yield | Sugar Yield |
|---|---|---|---|---|---|---|---|
| (cm) | (cm) | (%) | (kg) | (Numbers/hm−2) | (kg/hm−2) | (kg/hm−2) | |
| CK | 287.67 ± 17.18 b | 2.68 ± 0.92 b | 10.85 ± 0.64 ab | 1.46 ± 0.13 bc | 55,030 ± 6808 a | 79,749.21 ± 6745.89 abc | 8652.44 ± 893.44 bc |
| D1 | 287.92 ± 6.74 b | 2.75 ± 0.24 b | 10.43 ± 0.26 ab | 1.54 ± 0.23 bc | 40,080 ± 2324 b | 68,961.30 ± 5467.62 bc | 6789.10 ± 797.34 bc |
| D15 | 288.83 ± 7.75 b | 2.92 ± 0.04 ab | 10.50 ± 0.43 ab | 1.74 ± 0.09 ab | 40,950 ± 5877 b | 70,961.30 ± 8467.84 bc | 7441.10 ± 949.13 bc |
| D2 | 302.30 ± 5.07 ab | 2.97 ± 0.14 a | 10.24 ± 0.31 ab | 1.90 ± 0.21 a | 47,845 ± 1045 ab | 90,357.16 ± 8398.16 ab | 9232.12 ± 597.13 ab |
| D25 | 314.03 ± 9.74 a | 3.02 ± 0.11 a | 11.32 ± 0.39 a | 2.04 ± 0.21 a | 49,125 ± 2288 ab | 100,657.59 ± 13,961.44 a | 11,430.61 ± 1939.49 a |
| D3 | 304.13 ± 5.72 ab | 3.07 ± 0.20 a | 9.70 ± 1.13 b | 2.03 ± 0.27 a | 50,035 ± 9070 ab | 101,594.66 ± 24,240.53 a | 9827.33 ± 2404.75 ab |
| Treatment | pH | SOC (g/kg) | AN (mg/kg) | AP (mg/kg) | AK (mg/kg) |
|---|---|---|---|---|---|
| CK | 4.34 ± 0.16 b | 37.93 ± 6.64 b | 107.33 ± 17.96 ab | 378.77 ± 136.18 a | 196.36 ± 46.56 b |
| D1 | 4.97 ± 0.29 a | 48.70 ± 12.75 ab | 119.00 ± 37.86 ab | 168.15 ± 37.70 b | 170.45 ± 54.00 b |
| D15 | 4.61 ± 0.21 ab | 56.06 ± 9.64 a | 169.17 ± 67.26 a | 159.69 ± 20.46 b | 142.09 ± 25.56 b |
| D2 | 4.26 ± 0.24 b | 40.19 ± 7.94 b | 115.50 ± 35.00 ab | 138.77 ± 5.29 b | 181.01 ± 13.22 b |
| D25 | 4.37 ± 0.21 b | 40.38 ± 7.08 b | 101.50 ± 29.90 b | 184.30 ± 38.22 b | 200.99 ± 65.36 b |
| D3 | 4.38 ± 0.15 b | 42.65 ± 5.14 ab | 117.17 ± 1.76 ab | 273.73 ± 35.86 a | 302.43 ± 86.76 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Z.; Liu, Q.; Mo, L.; Pang, Z.; Hu, C. Integrating the Soil Microbiota and Metabolome Reveals the Mechanism through Which Controlled Release Fertilizer Affects Sugarcane Growth. Int. J. Mol. Sci. 2023, 24, 14086. https://doi.org/10.3390/ijms241814086
Yuan Z, Liu Q, Mo L, Pang Z, Hu C. Integrating the Soil Microbiota and Metabolome Reveals the Mechanism through Which Controlled Release Fertilizer Affects Sugarcane Growth. International Journal of Molecular Sciences. 2023; 24(18):14086. https://doi.org/10.3390/ijms241814086
Chicago/Turabian StyleYuan, Zhaonian, Qiang Liu, Lifang Mo, Ziqin Pang, and Chaohua Hu. 2023. "Integrating the Soil Microbiota and Metabolome Reveals the Mechanism through Which Controlled Release Fertilizer Affects Sugarcane Growth" International Journal of Molecular Sciences 24, no. 18: 14086. https://doi.org/10.3390/ijms241814086
APA StyleYuan, Z., Liu, Q., Mo, L., Pang, Z., & Hu, C. (2023). Integrating the Soil Microbiota and Metabolome Reveals the Mechanism through Which Controlled Release Fertilizer Affects Sugarcane Growth. International Journal of Molecular Sciences, 24(18), 14086. https://doi.org/10.3390/ijms241814086