

Single-Cell RNA-Sequencing Reveals Peripheral T Helper Cells Promoting the Development of IgG4-Related Disease by Enhancing B Cell Activation and Differentiation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
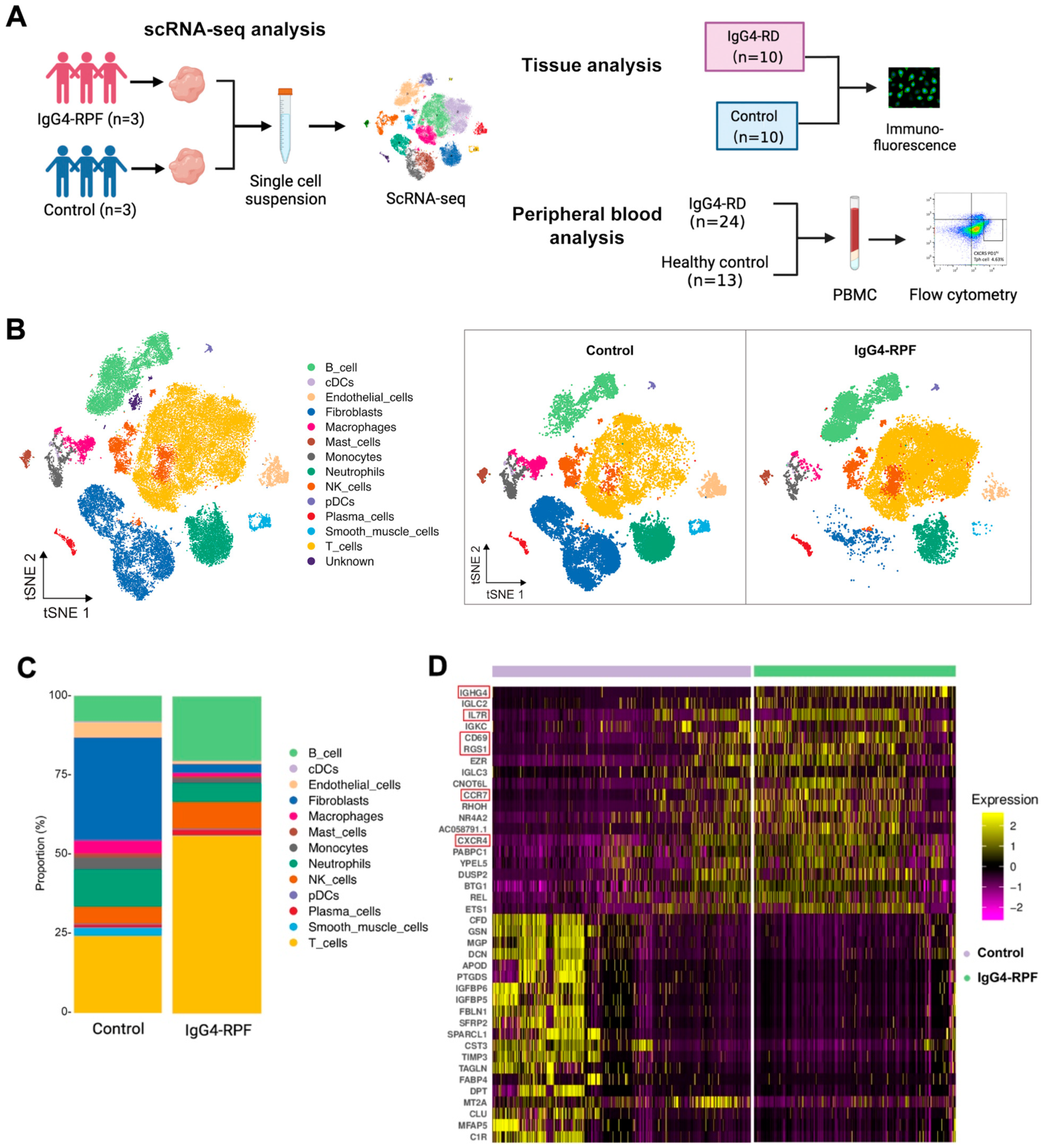
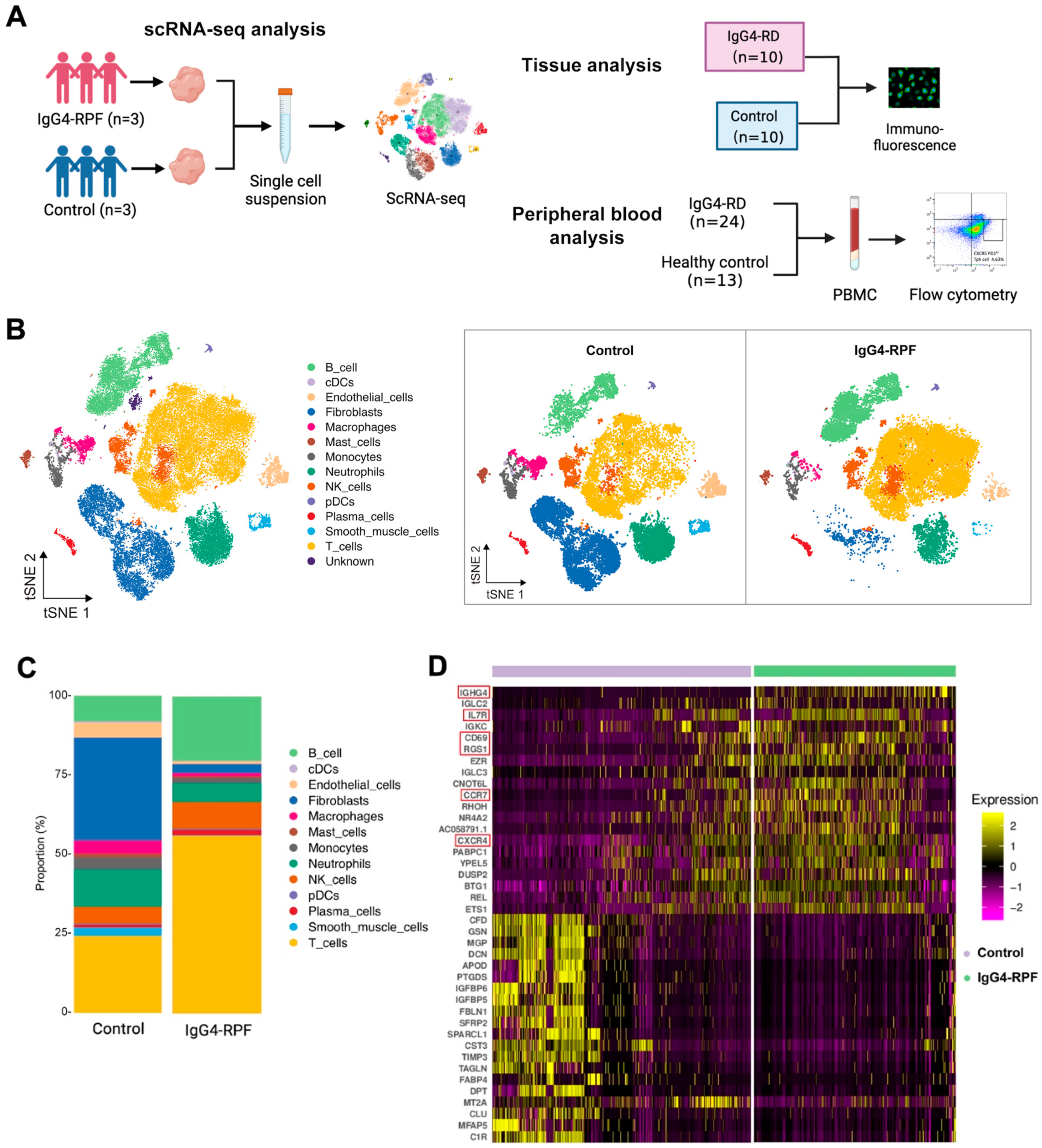
2.1. Study Population
2.2. B and T Cell Proportions Are Increased in the Tissue of IgG4-RPF Patients
2.3. The Active B Cells Are Increased in the Tissue of IgG4-RPF Patients
2.4. Gene Set Enrichment Analysis Reveals Enrichment of Cell Activation, Cytokine Secretion, and Receptor Binding Pathways in Active B Cells
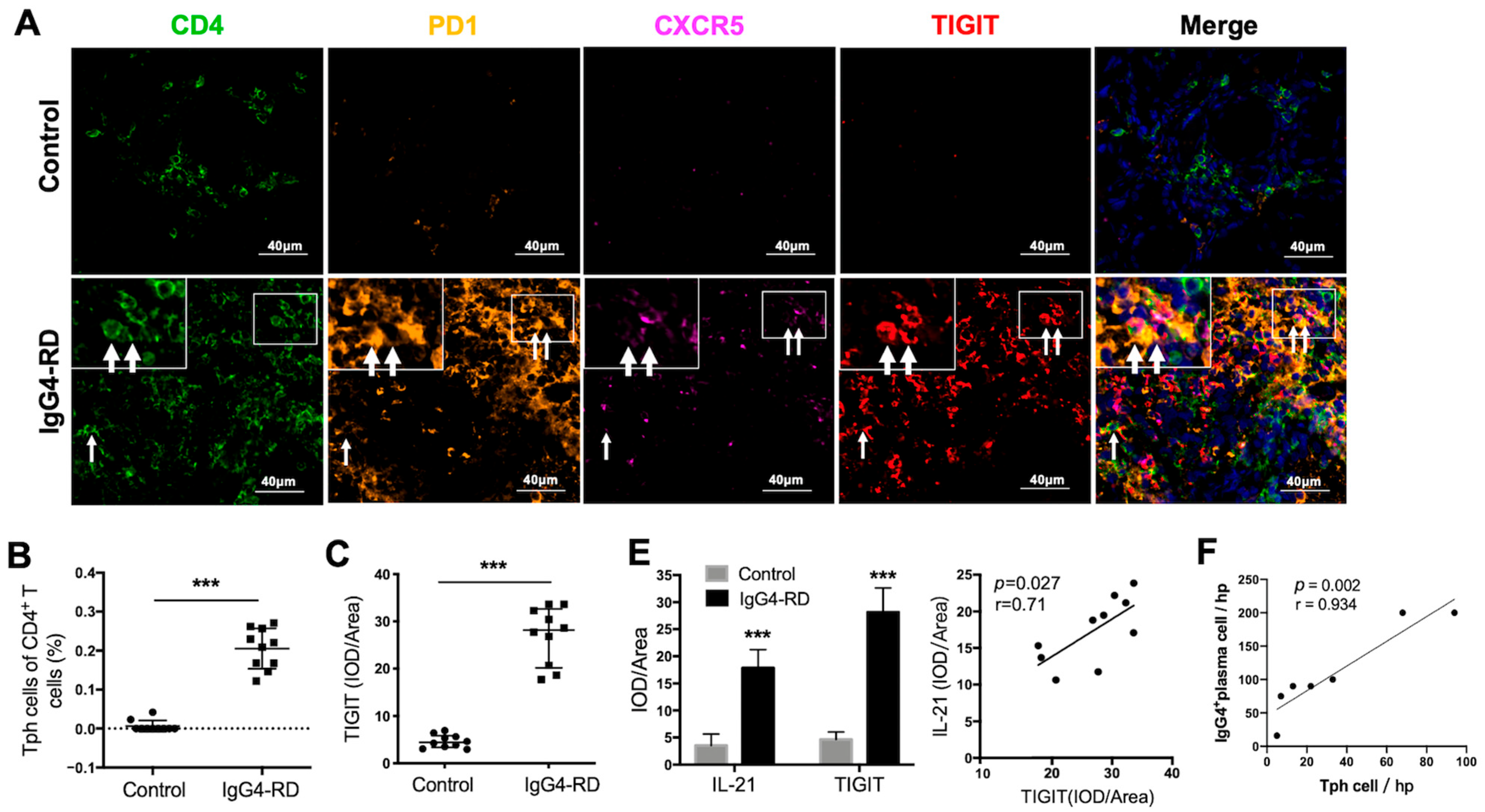
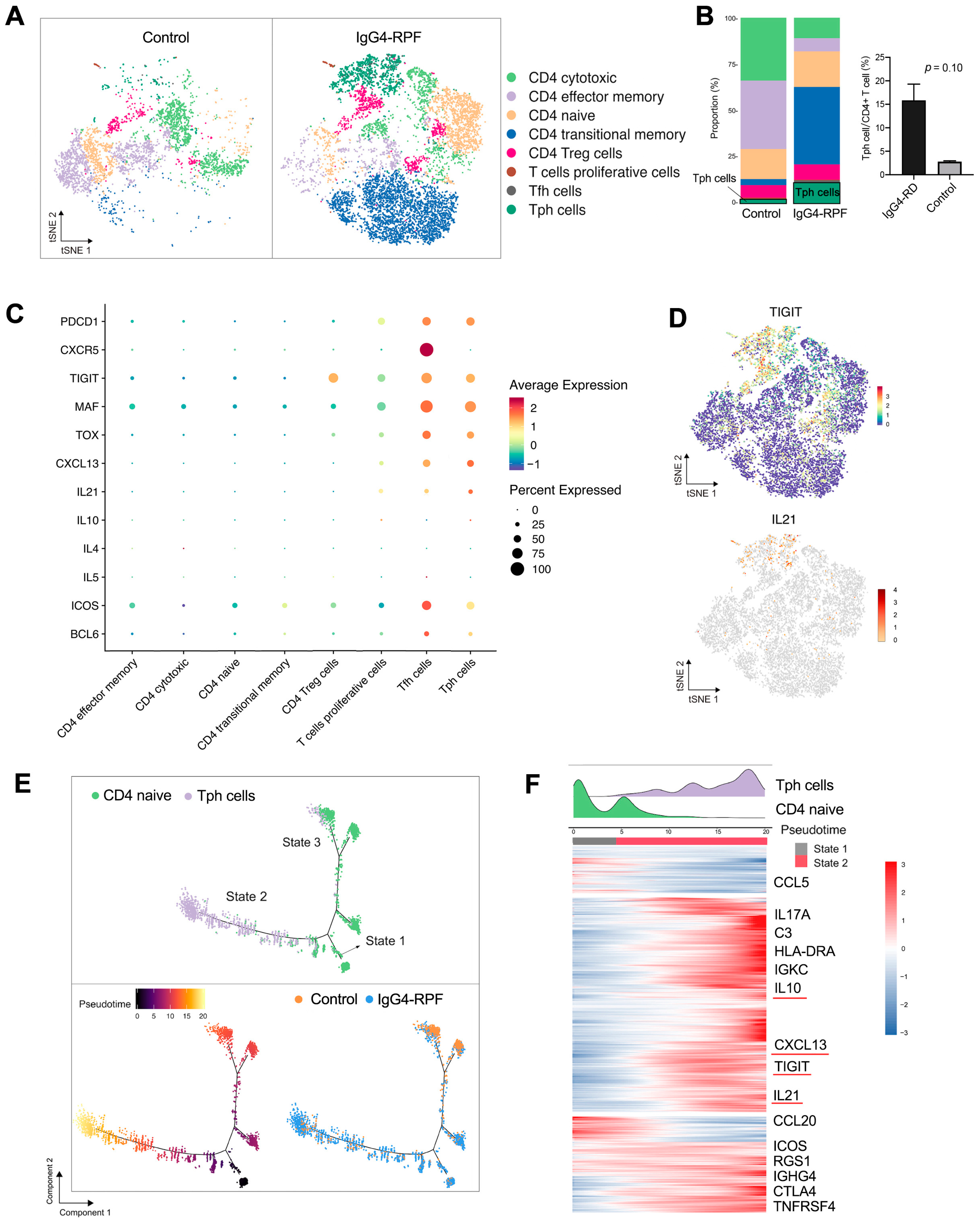
2.5. Tph Cells Are Increased in the Tissue of IgG4-RPF Patients
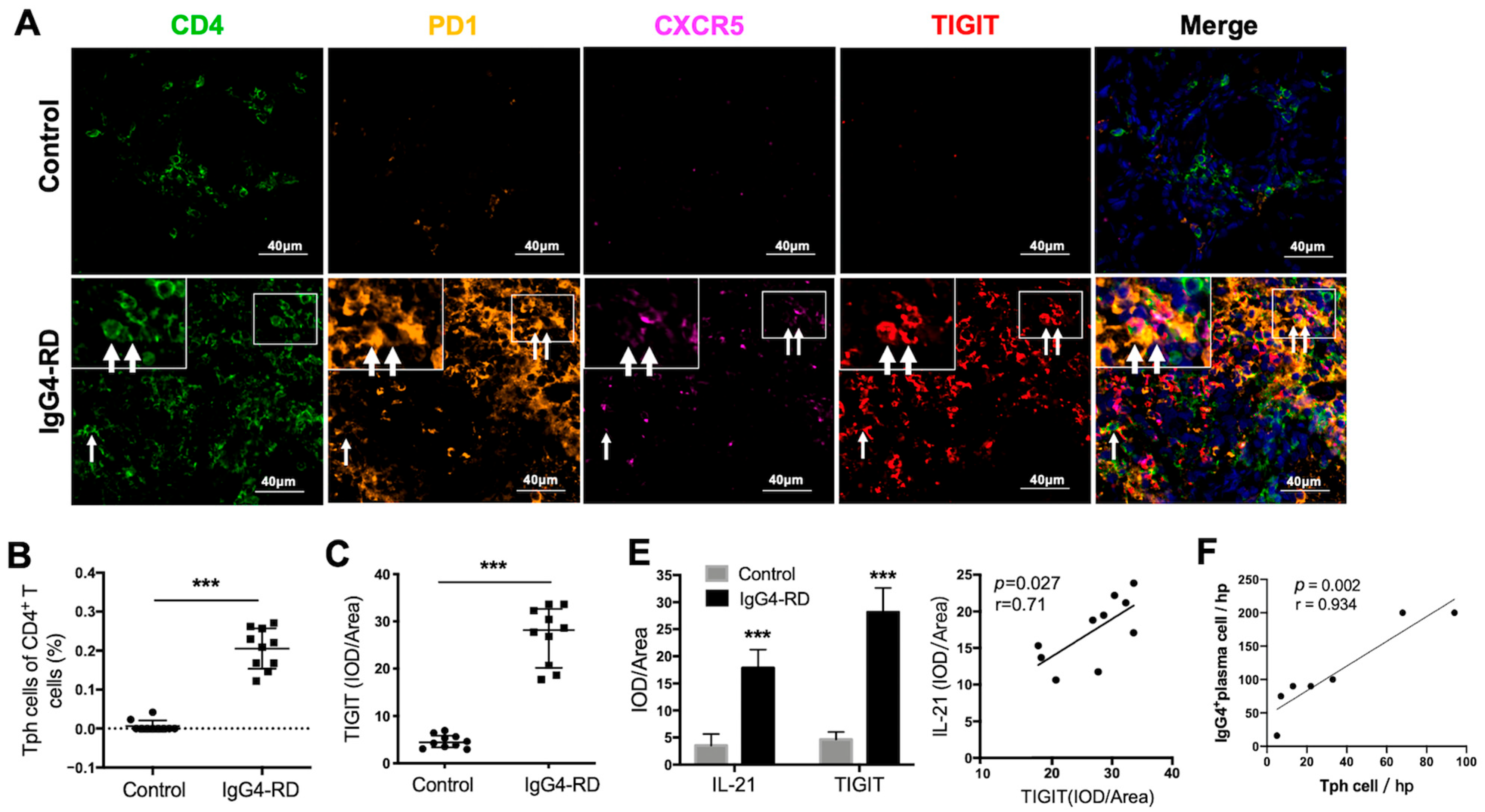
2.6. Tph Cells Were Increased in Both the Retroperitoneum Tissues and Salivary Glands in IgG4-RD Patients
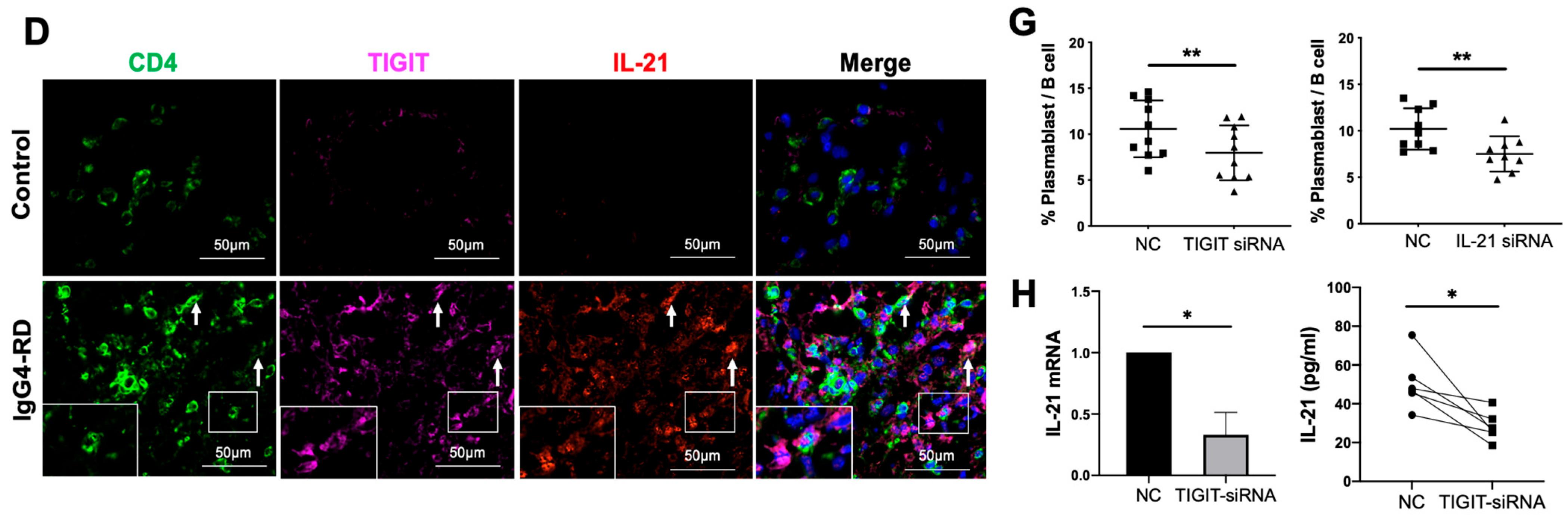
2.7. Tph Cells Promote B Cell Differentiation by TIGIT-IL-21 Axis In Vitro
2.8. Peripheral Tph Cells and TIGIT+ Tph Cells Are Positively Correlated with IgG4-RD Activity
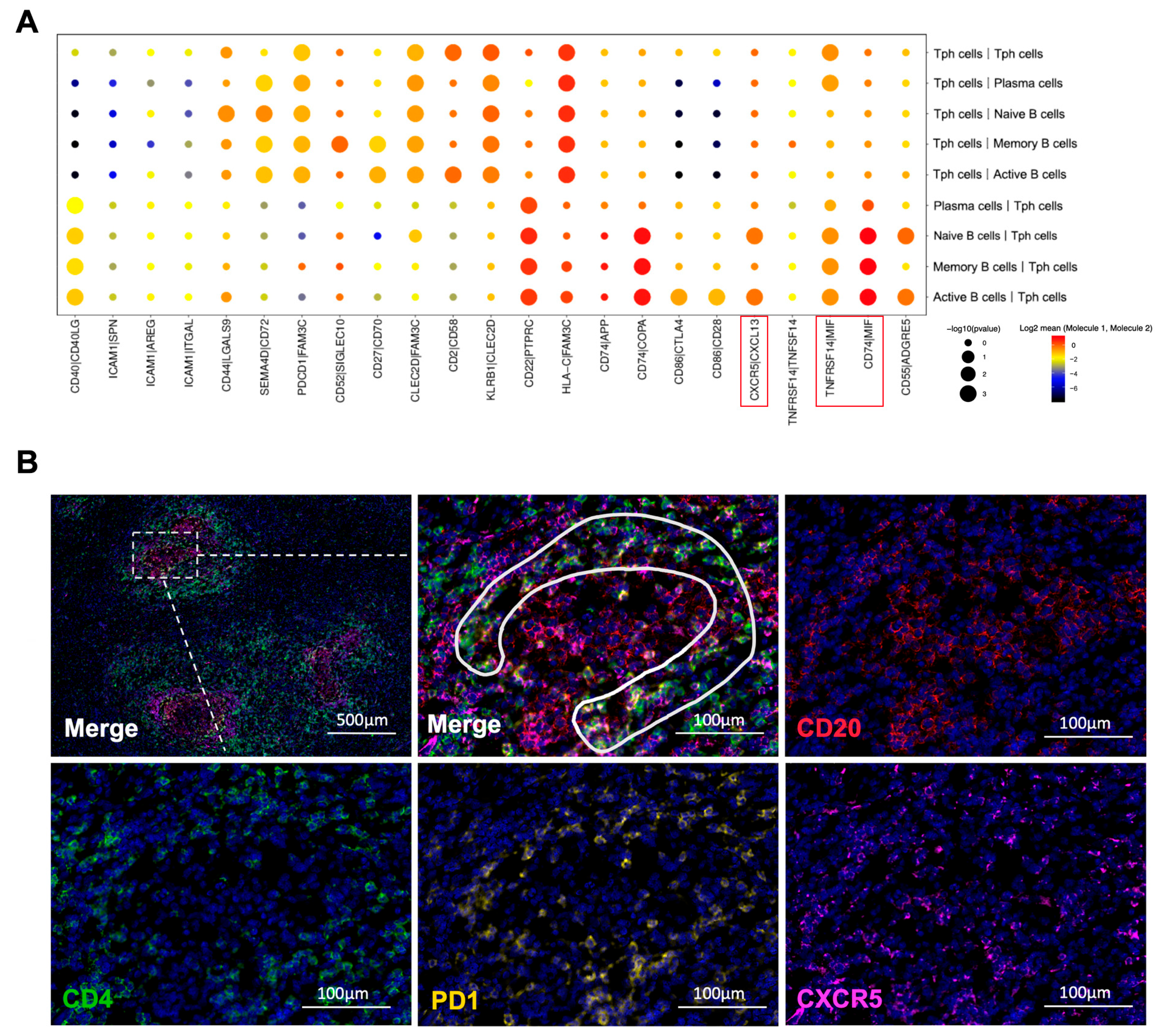
2.9. Tph Cell and B Cell Interactions Are Upregulated in IgG4-RPF
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. ScRNA-Seq
4.2.1. Preparation of Single-Cell Suspensions
4.2.2. Data Acquisition and Quality Control of the Single Cells
4.2.3. Gene Analysis, Clustering, and Identification of the Single Cells
4.2.4. Pseudotime Analysis
4.2.5. Cell–Cell Communication Analysis and Gene Set Variation Analysis (GSVA)
4.3. Flow Cytometry
4.4. Immunohistochemistry and Immunofluorescence Staining
4.5. Isolation of CD4+ T and B Cells and Cell Culture
4.6. Transfection of Small Interfering RNA (siRNA)
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. RNA Extraction and Real-Time Quantitative PCR
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takahashi, H.; Yamamoto, M.; Suzuki, C.; Naishiro, Y.; Shinomura, Y.; Imai, K. The birthday of a new syndrome: IgG4-related diseases constitute a clinical entity. Autoimmun. Rev. 2010, 9, 591–594. [Google Scholar] [CrossRef] [PubMed]
- Wallace, Z.S.; Miles, G.; Smolkina, E.; Petruski-Ivleva, N.; Madziva, D.; Cook, C.; Fu, X.; Zhang, Y.; Stone, J.H.; Choi, H.K. Incidence, prevalence and mortality of IgG4-related disease in the USA: A claims-based analysis of commercially insured adults. Ann. Rheum. Dis. 2023, 82, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Masamune, A.; Shimosegawa, T.; Okazaki, K. Prevalence of IgG4-Related Disease in Japan Based on Nationwide Survey in 2009. Int. J. Rheumatol. 2012, 2012, 358371. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, P.; Zhang, X.; Lin, W.; Tang, H.; Li, J.; Wang, M.; Liu, X.; Fei, Y.; Chen, H.; et al. Sex disparities in clinical characteristics and prognosis of immunoglobulin G4-related disease: A prospective study of 403 patients. Rheumatology 2019, 58, 820–830. [Google Scholar] [CrossRef]
- Wallace, Z.S.; Zhang, Y.; Perugino, C.A.; Naden, R.; Choi, H.K.; Stone, J.H.; ACR/EULAR IgG4-RD Classification Criteria Committee. Clinical phenotypes of IgG4-related disease: An analysis of two international cross-sectional cohorts. Ann. Rheum. Dis. 2019, 78, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Lian, L.; Wang, C.; Tian, J.L. IgG4-related retroperitoneal fibrosis: A newly characterized disease. Int. J. Rheum. Dis. 2016, 19, 1049–1055. [Google Scholar] [CrossRef]
- Umehara, H.; Nakajima, A.; Nakamura, T.; Kawanami, T.; Tanaka, M.; Dong, L.; Kawano, M. IgG4-related disease and its pathogenesis-cross-talk between innate and acquired immunity. Int. Immunol. 2014, 26, 585–595. [Google Scholar] [CrossRef]
- Maehara, T.; Moriyama, M.; Nakamura, S. Pathogenesis of IgG4-related disease: A critical review. Odontology 2019, 107, 127–132. [Google Scholar] [CrossRef]
- Mbengue, M.; Goumri, N.; Niang, A. IgG4-related kidney disease: Pathogenesis, diagnosis, and treatment. Clin. Nephrol. 2021, 95, 292–302. [Google Scholar] [CrossRef]
- Zongfei, J.; Rongyi, C.; Xiaomeng, C.; Lili, M.; Lingying, M.; Xiufang, K.; Xiaomin, D.; Zhuojun, Z.; Huiyong, C.; Ying, S.; et al. In vitro IL-6/IL-6R Trans-Signaling in Fibroblasts Releases Cytokines That May Be Linked to the Pathogenesis of IgG4-Related Disease. Front. Immunol. 2020, 11, 1272. [Google Scholar] [CrossRef]
- Takeuchi, M.; Sato, Y.; Ohno, K.; Tanaka, S.; Takata, K.; Gion, Y.; Orita, Y.; Ito, T.; Tachibana, T.; Yoshino, T. T helper 2 and regulatory T-cell cytokine production by mast cells: A key factor in the pathogenesis of IgG4-related disease. Mod. Pathol. 2014, 27, 1126–1136. [Google Scholar] [CrossRef]
- Arai, Y.; Yamashita, K.; Kuriyama, K.; Shiokawa, M.; Kodama, Y.; Sakurai, T.; Mizugishi, K.; Uchida, K.; Kadowaki, N.; Takaori-Kondo, A.; et al. Plasmacytoid Dendritic Cell Activation and IFN-α Production Are Prominent Features of Murine Autoimmune Pancreatitis and Human IgG4-Related Autoimmune Pancreatitis. J. Immunol. 2015, 195, 3033–3044. [Google Scholar] [CrossRef]
- Akiyama, M.; Suzuki, K.; Yasuoka, H.; Kaneko, Y.; Yamaoka, K.; Takeuchi, T. Follicular helper T cells in the pathogenesis of IgG4-related disease. Rheumatology 2018, 57, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Della-Torre, E.; Rigamonti, E.; Perugino, C.; Baghai-Sain, S.; Sun, N.; Kaneko, N.; Maehara, T.; Rovati, L.; Ponzoni, M.; Milani, R.; et al. B lymphocytes directly contribute to tissue fibrosis in patients with IgG4-related disease. J. Allergy Clin. Immunol. 2020, 145, 968–981.e14. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Moriyama, M.; Nakashima, H.; Miyake, K.; Hayashida, J.N.; Maehara, T.; Shinozaki, S.; Kubo, Y.; Nakamura, S. Th2 and regulatory immune reactions contribute to IgG4 production and the initiation of Mikulicz disease. Arthritis Rheum. 2012, 64, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Akitake, R.; Watanabe, T.; Zaima, C.; Uza, N.; Ida, H.; Tada, S.; Nishida, N.; Chiba, T. Possible involvement of T helper type 2 responses to Toll-like receptor ligands in IgG4-related sclerosing disease. Gut 2010, 59, 542–545. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, N.; Moriyama, M.; Furusho, K.; Furukawa, S.; Shibata, T.; Murakami, Y.; Chinju, A.; Haque, A.; Gion, Y.; Ohta, M.; et al. Activated M2 Macrophages Contribute to the Pathogenesis of IgG4-Related Disease via Toll-like Receptor 7/Interleukin-33 Signaling. Arthritis Rheumatol. 2020, 72, 166–178. [Google Scholar] [CrossRef]
- Li, P.H.; Ko, K.L.; Ho, C.T.; Lau, L.L.; Tsang, R.K.; Cheung, T.T.; Leung, W.K.; Lau, C.S. Immunoglobulin G4-related disease in Hong Kong: Clinical features, treatment practices, and its association with multisystem disease. Hong Kong Med. J. 2017, 23, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, M.; Nakamura, S. Th1/Th2 Immune Balance and Other T Helper Subsets in IgG4-Related Disease. Curr. Top. Microbiol. Immunol. 2017, 401, 75–83. [Google Scholar] [CrossRef]
- Munemura, R.; Maehara, T.; Murakami, Y.; Koga, R.; Aoyagi, R.; Kaneko, N.; Doi, A.; Perugino, C.A.; Della-Torre, E.; Saeki, T.; et al. Distinct disease-specific Tfh cell populations in 2 different fibrotic diseases: IgG4-related disease and Kimura disease. J. Allergy Clin. Immunol. 2022, 150, 440–455.e17. [Google Scholar] [CrossRef]
- Nakashima, H.; Miyake, K.; Moriyama, M.; Tanaka, A.; Watanabe, M.; Abe, Y.; Sato, H.; Nakamura, S.; Saito, T. An amplification of IL-10 and TGF-beta in patients with IgG4-related tubulointerstitial nephritis. Clin. Nephrol. 2010, 73, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.A.; Gurish, M.F.; Marshall, J.L.; Slowikowski, K.; Fonseka, C.Y.; Liu, Y.; Donlin, L.T.; Henderson, L.A.; Wei, K.; Mizoguchi, F.; et al. Pathologically expanded peripheral T helper cell subset drives B cells in rheumatoid arthritis. Nature 2017, 542, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Bocharnikov, A.V.; Keegan, J.; Wacleche, V.S.; Cao, Y.; Fonseka, C.Y.; Wang, G.; Muise, E.S.; Zhang, K.X.; Arazi, A.; Keras, G.; et al. PD-1hiCXCR5− T peripheral helper cells promote B cell responses in lupus via MAF and IL-21. J. Clin. Investig. 2019, 4, e130062. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, M.; Chen, Y.; Li, J.; Liu, Z.; Lu, H.; Fei, Y.; Feng, R.; Zhao, Y.; Zeng, X.; et al. Expanded CD4+CXCR5−PD-1+ peripheral T helper like cells and clinical significance in IgG4-related disease. Clin. Immunol. 2022, 237, 108975. [Google Scholar] [CrossRef]
- Kamekura, R.; Takahashi, H.; Ichimiya, S. New insights into IgG4-related disease: Emerging new CD4+ T-cell subsets. Curr. Opin. Rheumatol. 2019, 31, 9–15. [Google Scholar] [CrossRef]
- Deshpande, V.; Zen, Y.; Chan, J.K.; Yi, E.E.; Sato, Y.; Yoshino, T.; Kloppel, G.; Heathcote, J.G.; Khosroshahi, A.; Ferry, J.A.; et al. Consensus statement on the pathology of IgG4-related disease. Mod. Pathol. 2012, 25, 1181–1192. [Google Scholar] [CrossRef]
- Corradi, D.; Maestri, R.; Palmisano, A.; Bosio, S.; Greco, P.; Manenti, L.; Ferretti, S.; Cobelli, R.; Moroni, G.; Dei Tos, A.P.; et al. Idiopathic retroperitoneal fibrosis: Clinicopathologic features and differential diagnosis. Kidney Int. 2007, 72, 742–753. [Google Scholar] [CrossRef]
- Fry, T.J.; Mackall, C.L. The many faces of IL-7: From lymphopoiesis to peripheral T cell maintenance. J. Immunol. 2005, 174, 6571–6576. [Google Scholar] [CrossRef]
- Caballero-Franco, C.; Kissler, S. The autoimmunity-associated gene RGS1 affects the frequency of T follicular helper cells. Genes. Immun. 2016, 17, 228–238. [Google Scholar] [CrossRef]
- Vance, B.A.; Harley, P.H.; Backlund, P.S.; Ward, Y.; Phelps, T.L.; Gress, R.E. Human CD69 associates with an N-terminal fragment of calreticulin at the cell surface. Arch. Biochem. Biophys. 2005, 438, 11–20. [Google Scholar] [CrossRef]
- Forster, R.; Davalos-Misslitz, A.C.; Rot, A. CCR7 and its ligands: Balancing immunity and tolerance. Nat. Rev. Immunol. 2008, 8, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Capecchi, R.; Croia, C.; Puxeddu, I.; Pratesi, F.; Cacciato, A.; Campani, D.; Boggi, U.; Morelli, L.; Tavoni, A.; Migliorini, P. CXCL12/SDF-1 in IgG4-Related Disease. Front. Pharmacol. 2021, 12, 750216. [Google Scholar] [CrossRef] [PubMed]
- Han, B.K.; Olsen, N.J.; Bottaro, A. The CD27-CD70 pathway and pathogenesis of autoimmune disease. Semin. Arthritis Rheum. 2016, 45, 496–501. [Google Scholar] [CrossRef]
- Capasso, M.; Durrant, L.G.; Stacey, M.; Gordon, S.; Ramage, J.; Spendlove, I. Costimulation via CD55 on human CD4+ T cells mediated by CD97. J. Immunol. 2006, 177, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Kwakkenbos, M.J.; Pouwels, W.; Matmati, M.; Stacey, M.; Lin, H.H.; Gordon, S.; van Lier, R.A.; Hamann, J. Expression of the largest CD97 and EMR2 isoforms on leukocytes facilitates a specific interaction with chondroitin sulfate on B cells. J. Leukoc. Biol. 2005, 77, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Nieto, P.; Elosua-Bayes, M.; Trincado, J.L.; Marchese, D.; Massoni-Badosa, R.; Salvany, M.; Henriques, A.; Nieto, J.; Aguilar-Fernandez, S.; Mereu, E.; et al. A single-cell tumor immune atlas for precision oncology. Genome Res. 2021, 31, 1913–1926. [Google Scholar] [CrossRef]
- Poch, T.; Krause, J.; Casar, C.; Liwinski, T.; Glau, L.; Kaufmann, M.; Ahrenstorf, A.E.; Hess, L.U.; Ziegler, A.E.; Martrus, G.; et al. Single-cell atlas of hepatic T cells reveals expansion of liver-resident naive-like CD4+ T cells in primary sclerosing cholangitis. J. Hepatol. 2021, 75, 414–423. [Google Scholar] [CrossRef]
- Tanemoto, S.; Sujino, T.; Miyamoto, K.; Moody, J.; Yoshimatsu, Y.; Ando, Y.; Koya, I.; Harada, Y.; Tojo, A.O.; Ono, K.; et al. Single-cell transcriptomics of human gut T cells identifies cytotoxic CD4+CD8A+ T cells related to mouse CD4 cytotoxic T cells. Front. Immunol. 2022, 13, 977117. [Google Scholar] [CrossRef]
- Perugino, C.A.; Kaneko, N.; Maehara, T.; Mattoo, H.; Kers, J.; Allard-Chamard, H.; Mahajan, V.S.; Liu, H.; Della-Torre, E.; Murphy, S.J.H.; et al. CD4+ and CD8+ cytotoxic T lymphocytes may induce mesenchymal cell apoptosis in IgG4-related disease. J. Allergy Clin. Immunol. 2021, 147, 368–382. [Google Scholar] [CrossRef]
- Jiang, Q.; Wang, J.; Jiang, H.; Li, W.; Sun, Y.; Shan, Y.; Wei, T.; Chi, X.; Yu, S.; Ma, X. Competitive binding of transcription factors underlies flexibility of T peripheral helper cells and T follicular helper cells in SLE. Rheumatology 2022, 61, 4547–4557. [Google Scholar] [CrossRef]
- Shan, Y.; Nakayamada, S.; Nawata, A.; Yamagata, K.; Sonomoto, K.; Tanaka, H.; Satoh-Kanda, Y.; Nguyen, M.P.; Todoroki, Y.; Nagayasu, A.; et al. TGF-β3 in differentiation and function of Tph-like cells and its relevance to disease activity in patients with systemic lupus erythematosus. Rheumatology 2023, 62, 2464–2474. [Google Scholar] [CrossRef]
- Shibuya, A.; Shibuya, K. DNAM-1 versus TIGIT: Competitive roles in tumor immunity and inflammatory responses. Int. Immunol. 2021, 33, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; van Besouw, N.M.; Shi, Y.; Hoogduijn, M.J.; Wang, L.; Baan, C.C. The Biological Effects of IL-21 Signaling on B-Cell-Mediated Responses in Organ Transplantation. Front. Immunol. 2016, 7, 319. [Google Scholar] [CrossRef] [PubMed]
- Alampour-Rajabi, S.; El Bounkari, O.; Rot, A.; Muller-Newen, G.; Bachelerie, F.; Gawaz, M.; Weber, C.; Schober, A.; Bernhagen, J. MIF interacts with CXCR7 to promote receptor internalization, ERK1/2 and ZAP-70 signaling, and lymphocyte chemotaxis. FASEB J. 2015, 29, 4497–4511. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Peng, Y.; Li, J.; Zhang, P.; Liu, Z.; Lu, H.; Peng, L.; Zhou, J.; Fei, Y.; Zeng, X.; et al. Single-Cell Sequencing of Immune Cell Heterogeneity in IgG4-Related Disease. Front. Immunol. 2022, 13, 904288. [Google Scholar] [CrossRef]
- Cai, S.; Chen, Y.; Hu, Z.; Zhou, T.; Huang, Y.; Lin, S.; Gao, R.; Zhong, J.; Dong, L. The landscape of T and B lymphocytes interaction and synergistic effects of Th1 and Th2 type response in the involved tissue of IgG4-RD revealed by single cell transcriptome analysis. J. Autoimmun. 2022, 133, 102944. [Google Scholar] [CrossRef]
- Khosroshahi, A.; Wallace, Z.S.; Crowe, J.L.; Akamizu, T.; Azumi, A.; Carruthers, M.N.; Chari, S.T.; Della-Torre, E.; Frulloni, L.; Goto, H.; et al. International Consensus Guidance Statement on the Management and Treatment of IgG4-Related Disease. Arthritis Rheumatol. 2015, 67, 1688–1699. [Google Scholar] [CrossRef]
- Akiyama, M.; Yasuoka, H.; Yoshimoto, K.; Takeuchi, T. Interleukin-4 contributes to the shift of balance of IgG subclasses toward IgG4 in IgG4-related disease. Cytokine 2018, 110, 416–419. [Google Scholar] [CrossRef]
- Martin-Nares, E.; Hernandez-Molina, G.; Priego-Ranero, A.A.; Chan-Campos, I.; Herrera-Noguera, G.S.; Lopez-Verdugo, F.; Furuzawa-Carballeda, J. Peripheral Immunophenotype in IgG4-Related Disease and Its Association with Clinical Phenotypes and Disease Activity. Cells 2023, 12, 670. [Google Scholar] [CrossRef]
- Grados, A.; Ebbo, M.; Piperoglou, C.; Groh, M.; Regent, A.; Samson, M.; Terrier, B.; Loundou, A.; Morel, N.; Audia, S.; et al. T Cell Polarization toward TH2/TFH2 and TH17/TFH17 in Patients with IgG4-Related Disease. Front. Immunol. 2017, 8, 235. [Google Scholar] [CrossRef]
- Touzani, F.; Pozdzik, A. New insights into immune cells cross-talk during IgG4-related disease. Clin. Immunol. 2019, 198, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Holm Hansen, R.; Hojsgaard Chow, H.; Talbot, J.; Buhelt, S.; Nickelsen Hellem, M.N.; Nielsen, J.E.; Sellebjerg, F.T.; von Essen, M.R. Peripheral helper T cells in the pathogenesis of multiple sclerosis. Mult. Scler. 2022, 28, 1340–1350. [Google Scholar] [CrossRef] [PubMed]
- Makiyama, A.; Chiba, A.; Noto, D.; Murayama, G.; Yamaji, K.; Tamura, N.; Miyake, S. Expanded circulating peripheral helper T cells in systemic lupus erythematosus: Association with disease activity and B cell differentiation. Rheumatology 2019, 58, 1861–1869. [Google Scholar] [CrossRef]
- Chen, W.; Yang, F.; Lin, J. Tph Cells Expanded in Primary Sjogren’s Syndrome. Front Med. 2022, 9, 900349. [Google Scholar] [CrossRef] [PubMed]
- Yoshitomi, H.; Ueno, H. Shared and distinct roles of T peripheral helper and T follicular helper cells in human diseases. Cell Mol. Immunol. 2021, 18, 523–527. [Google Scholar] [CrossRef]
- Marks, K.E.; Rao, D.A. T peripheral helper cells in autoimmune diseases. Immunol. Rev. 2022, 307, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.S.K. The link between circulating follicular helper T cells and autoimmunity. Nat. Rev. Immunol. 2022, 22, 567–575. [Google Scholar] [CrossRef]
- Manzo, A.; Vitolo, B.; Humby, F.; Caporali, R.; Jarrossay, D.; Dell’accio, F.; Ciardelli, L.; Uguccioni, M.; Montecucco, C.; Pitzalis, C. Mature antigen-experienced T helper cells synthesize and secrete the B cell chemoattractant CXCL13 in the inflammatory environment of the rheumatoid joint. Arthritis Rheum. 2008, 58, 3377–3387. [Google Scholar] [CrossRef]
- Godefroy, E.; Zhong, H.; Pham, P.; Friedman, D.; Yazdanbakhsh, K. TIGIT-positive circulating follicular helper T cells display robust B-cell help functions: Potential role in sickle cell alloimmunization. Haematologica 2015, 100, 1415–1425. [Google Scholar] [CrossRef]
- Maehara, T.; Moriyama, M.; Nakashima, H.; Miyake, K.; Hayashida, J.N.; Tanaka, A.; Shinozaki, S.; Kubo, Y.; Nakamura, S. Interleukin-21 contributes to germinal centre formation and immunoglobulin G4 production in IgG4-related dacryoadenitis and sialoadenitis, so-called Mikulicz’s disease. Ann. Rheum. Dis. 2012, 71, 2011–2019. [Google Scholar] [CrossRef]
- Okazaki, K.; Umehara, H. Are Classification Criteria for IgG4-RD Now Possible? The Concept of IgG4-Related Disease and Proposal of Comprehensive Diagnostic Criteria in Japan. Int. J. Rheumatol. 2012, 2012, 357071. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Lu, S.; Chen, H.; Wu, Q.; Fei, Y.; Li, M.; Zhang, X.; Tian, X.; Zheng, W.; Leng, X.; et al. Clinical characteristics of immunoglobulin G4-related disease: A prospective study of 118 Chinese patients. Rheumatology 2015, 54, 1982–1990. [Google Scholar] [CrossRef] [PubMed]
- Yunyun, F.; Yu, C.; Panpan, Z.; Hua, C.; Di, W.; Lidan, Z.; Linyi, P.; Li, W.; Qingjun, W.; Xuan, Z.; et al. Efficacy of Cyclophosphamide treatment for immunoglobulin G4-related disease with addition of glucocorticoids. Sci. Rep. 2017, 7, 6195. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411–420. [Google Scholar] [CrossRef]
- McGinnis, C.S.; Murrow, L.M.; Gartner, Z.J. DoubletFinder: Doublet Detection in Single-Cell RNA Sequencing Data Using Artificial Nearest Neighbors. Cell Syst. 2019, 8, 329–337.e4. [Google Scholar] [CrossRef]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef]
- Moncada, R.; Barkley, D.; Wagner, F.; Chiodin, M.; Devlin, J.C.; Baron, M.; Hajdu, C.H.; Simeone, D.M.; Yanai, I. Author Correction: Integrating microarray-based spatial transcriptomics and single-cell RNA-seq reveals tissue architecture in pancreatic ductal adenocarcinomas. Nat. Biotechnol. 2020, 38, 1476. [Google Scholar] [CrossRef]
- Mabbott, N.A.; Baillie, J.K.; Brown, H.; Freeman, T.C.; Hume, D.A. An expression atlas of human primary cells: Inference of gene function from coexpression networks. BMC Genom. 2013, 14, 632. [Google Scholar] [CrossRef]
- Yost, K.E.; Satpathy, A.T.; Wells, D.K.; Qi, Y.; Wang, C.; Kageyama, R.; McNamara, K.L.; Granja, J.M.; Sarin, K.Y.; Brown, R.A.; et al. Clonal replacement of tumor-specific T cells following PD-1 blockade. Nat. Med. 2019, 25, 1251–1259. [Google Scholar] [CrossRef]
- Zhang, F.; Wei, K.; Slowikowski, K.; Fonseka, C.Y.; Rao, D.A.; Kelly, S.; Goodman, S.M.; Tabechian, D.; Hughes, L.B.; Salomon-Escoto, K.; et al. Defining inflammatory cell states in rheumatoid arthritis joint synovial tissues by integrating single-cell transcriptomics and mass cytometry. Nat. Immunol. 2019, 20, 928–942. [Google Scholar] [CrossRef]
- Trapnell, C.; Cacchiarelli, D.; Grimsby, J.; Pokharel, P.; Li, S.; Morse, M.; Lennon, N.J.; Livak, K.J.; Mikkelsen, T.S.; Rinn, J.L. The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat. Biotechnol. 2014, 32, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Efremova, M.; Vento-Tormo, M.; Teichmann, S.A.; Vento-Tormo, R. CellPhoneDB: Inferring cell-cell communication from combined expression of multi-subunit ligand-receptor complexes. Nat. Protoc. 2020, 15, 1484–1506. [Google Scholar] [CrossRef] [PubMed]
- Hanzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, Z.; Lu, W.; Wu, S.; Zhang, Y.; Meng, D.; Zhang, X.; Dai, X.; Chen, H.; Ma, L.; Sun, Y.; et al. Single-Cell RNA-Sequencing Reveals Peripheral T Helper Cells Promoting the Development of IgG4-Related Disease by Enhancing B Cell Activation and Differentiation. Int. J. Mol. Sci. 2023, 24, 13735. https://doi.org/10.3390/ijms241813735
Ji Z, Lu W, Wu S, Zhang Y, Meng D, Zhang X, Dai X, Chen H, Ma L, Sun Y, et al. Single-Cell RNA-Sequencing Reveals Peripheral T Helper Cells Promoting the Development of IgG4-Related Disease by Enhancing B Cell Activation and Differentiation. International Journal of Molecular Sciences. 2023; 24(18):13735. https://doi.org/10.3390/ijms241813735
Chicago/Turabian StyleJi, Zongfei, Weiqi Lu, Sifan Wu, Yong Zhang, Dan Meng, Xiao Zhang, Xiaojuan Dai, Huiyong Chen, Lili Ma, Ying Sun, and et al. 2023. "Single-Cell RNA-Sequencing Reveals Peripheral T Helper Cells Promoting the Development of IgG4-Related Disease by Enhancing B Cell Activation and Differentiation" International Journal of Molecular Sciences 24, no. 18: 13735. https://doi.org/10.3390/ijms241813735
APA StyleJi, Z., Lu, W., Wu, S., Zhang, Y., Meng, D., Zhang, X., Dai, X., Chen, H., Ma, L., Sun, Y., Jiang, L., & Kong, X. (2023). Single-Cell RNA-Sequencing Reveals Peripheral T Helper Cells Promoting the Development of IgG4-Related Disease by Enhancing B Cell Activation and Differentiation. International Journal of Molecular Sciences, 24(18), 13735. https://doi.org/10.3390/ijms241813735